소과
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
소과는 1821년 존 에드워드 그레이가 명명한 우제목의 과로, 143종의 현존하는 종과 300여 종의 멸종된 종을 포함한다. 소과 동물은 뭉툭한 주둥이, 뿔, 뚜렷한 목과 사지, 꼬리를 가지고 있으며, 크기와 털 색깔이 다양하다. 뿔의 형태는 종에 따라 다르며, 사회적 행동과 서식지, 먹이 습성도 다양하다. 소과는 미오세에 사슴과와 기린과에서 분화되었으며, 아프리카와 유라시아에서 진화하여 하위 분류로 8개의 아과로 나뉜다. 소는 유제품, 쇠고기, 가죽, 털 등 다양한 형태로 인간에게 유용하며, 가축화되어 농업에 기여했다.
더 읽어볼만한 페이지
- 소과 - 염소 (동물)
염소는 인류가 최초로 가축화한 소과 동물 중 하나로, 우유, 고기, 가죽, 털 등을 얻기 위해 사육되었으며, 양과 유사하지만 턱수염, 꼬리, 식성 등에서 차이를 보이고, 뛰어난 적응력으로 전 세계에 분포하지만, 야생화되어 생태계를 파괴하기도 한다. - 소과 - 영양 (동물)
영양은 소과에 속하는 초식 포유류로, 크기, 뿔의 유무와 모양, 분포 지역, 행동 양상 등이 종에 따라 다르며, 일부 종은 멸종 위기에 놓여 있고 인간과의 다양한 관계를 맺는다. - 존 에드워드 그레이가 명명한 분류군 - 투아타라
투아타라는 뉴질랜드 고유종 파충류이자 린코케팔리아목의 유일한 현존종으로, 오랜 진화 역사를 지니고 완전한 아래 측두궁 두개골, 첨생치, 두정안 등의 특징을 가지며 멸종 위기에 처해 뉴질랜드 정부의 보호를 받는다. - 존 에드워드 그레이가 명명한 분류군 - 코뿔소
코뿔소는 아프리카와 아시아에 서식하며 1~2개의 뿔과 두꺼운 피부를 가진 대형 초식성 포유류이지만, 밀렵으로 인해 5종 모두 멸종 위기에 처해 있다.
| 소과 - [생물]에 관한 문서 | |
|---|---|
| 과 정보 | |
 | |
| 학명 | Bovidae |
| 명명자 | Gray, 1821 |
| 모식속 | Bos |
| 모식속 명명자 | Linnaeus, 1758 |
| 하위 분류군 | |
| 아과 | 임팔라아과(Aepycerotinae) (1 속) 하테비스트아과(Alcelaphinae) (4 속) 영양아과(Antilopinae) (3 족 및 14 속) 소아과(Bovinae) (3 족 및 15 속) 양아과(Caprinae) (3 족 및 15 속) 다이커영양아과(Cephalophinae) (3 속) 말영양아과(Hippotraginae) (3 속) 네소트라구스아과(Nesotraginae) (1 속) 바위타기영양아과(Oreotraginae) (1 속) 리드벅아과(Reduncinae) (3 속) †힙소돈티나에아과(Hypsodontinae) |
| 대체 분류군 | 영양아과(Antilopinae) (10 족 및 45 속) 소아과(Bovinae) (3 족 및 15 속) †힙소돈티나에아과(Hypsodontinae) |
2. 명칭과 어원
"소과"라는 이름은 1821년 영국의 동물학자 존 에드워드 그레이가 명명했다.[1] "Bovidae"라는 단어는 접두사 'bov-' (라틴어 'bos', "소"에서 유래, 후기 라틴어 'bovinus'를 거쳐)와 접미사 '-idae'의 합성어이다.[2]
모든 소과 동물은 뭉툭한 주둥이, 한 쌍 이상의 뿔, 뚜렷한 목과 사지, 다양한 길이의 꼬리 등 비슷한 기본 형태를 띤다.[25] 대부분 성적 이형성을 보이며, 수컷이 암컷보다 크고 무겁다. 중대형 소과 동물에서 성적 이형성이 두드러진다. 모든 소과 동물은 각 발에 네 개의 발가락을 가지고 있으며, 가운데 두 개의 굽으로 걷고, 바깥쪽 두 개의 부제는 작고 거의 땅에 닿지 않는다.[3]
초기 미오세에 소과는 사슴과와 기린과에서 분화되기 시작했다. 초기 미오세 후반(20 Mya)에 아프리카와 유라시아에서 발견된 가장 초기의 소과는 현대의 가젤과 다소 유사한 작은 동물이었으며, 아마도 삼림 환경에서 살았을 것이다.[22] 가장 초기의 소과로 알려진 ''Eotragus''는 톰슨가젤과 거의 같은 크기였다.[21]
소과의 하위 분류는 다음과 같다.
3. 형태
소과 동물은 크기가 매우 다양하다. 가우르는 1500kg 이상, 어깨 높이가 2.2m에 달하는 반면,[26] 로열안텔로프는 키가 25cm이고 무게는 최대 3kg에 불과하다.[27] 바위산양은 어깨 높이가 45cm에서 60cm이고 무게는 10kg에서 20kg 정도이다.[28]
털 색깔은 아라비아오릭스처럼 옅은 흰색에서[29] 검은누처럼 검은색까지 다양하다.[30] 갈색이나 적갈색(갈대영양 등)이 일반적이다.[31] 누처럼 털에 줄무늬가 있거나, 아닥스처럼 계절에 따라 털 색깔이 변하기도 한다.[32] 일부 종은 암컷과 어린 개체가 밝은색 털을, 수컷은 나이가 들면서 짙은 색 털을 보인다. 향선과 피지선이 종종 존재한다.[25]
겜스복, 세이블영양, 그랜트가젤 등은 눈에 띄는 눈을 가리기 위해 파괴적 색채로 위장한다.[33] 가젤처럼 많은 종은 역음영을 통해 배경에 섞인다.[34] 많은 소과 동물의 윤곽은 뚜렷하게 대비되는 패턴으로 끊어져 포식자의 인식을 늦춘다.[35] 히포트라기네족(겜스복 포함)은 밝은 몸과 눈에 띄는 얼굴 무늬를 가지는데, 이는 의사소통에 기능할 수 있다. ''소'', ''양'', 본테복과 겜스복처럼 다리 색깔이 강하게 대비되는 것은 소과에서만 일반적인데, 이는 의사소통 기능일 가능성이 높다.[36]
일부 가축화된 형태를 제외하고 모든 수컷 소과 동물은 뿔을 가지고 있으며, 많은 종에서 암컷도 뿔을 가진다. 뿔의 크기와 모양은 다양하지만, 기본 구조는 갈라지지 않은 한 쌍의 뼈 돌기로, 케라틴 외피로 덮여 있다. 네 뿔 영양[37]과 제이콥양[38][39]처럼 예외도 있다. 뿔 구조는 다른 반추류와 구별되는 소과 동물의 특징이다.[40][41] 뿔 형태와 싸움 행동 사이에는 상관관계가 있다. 긴 뿔은 레슬링과 펜싱, 굽은 뿔은 들이받는 데 사용된다.[42] 안쪽으로 향하는 뿔을 가진 수컷은 일부일처제, 바깥쪽으로 향하는 뿔을 가진 수컷은 일부다처제 경향이 있다. 이는 신체 크기와 무관하다.[43]
수컷의 뿔 발달은 성 선택과 관련이 있다.[44][45] 일부일처제인 디커와 다른 작은 영양은 뿔이 작은 가시 모양인 반면, 일부다처제는 뿔이 크고 정교하게 형성된다(예: 자이언트엘런드처럼 나선형). 뿔은 종 내 수컷 간의 경쟁 정도를 나타낸다.[31] 암컷의 뿔은 자연 선택에 의한 것일 가능성이 높다.[44][46] 암컷의 뿔은 보통 수컷보다 작고 모양도 다르다. 암컷 소과 동물의 뿔은 포식자 방어 또는 영역 표시를 위해 진화한 것으로 여겨진다. 비영역형 암컷은 은폐를 통해 포식자를 피할 수 있어 뿔이 없는 경우가 많다.[46] 암컷이 뿔을 가진 경우는 소과 속의 절반에 불과하며, 이 속의 암컷은 더 무겁다. 암컷은 주로 찌르기 위해 뿔을 사용한다.[47]
소과 동물은 세 번째와 네 번째 중족골이 주상골로 결합된다. 자뼈와 종아리뼈는 축소되어 각각 요골과 경골에 융합된다. 긴 어깨뼈가 존재하지만 쇄골은 없다. 반추동물이기 때문에 위는 반추위, 제3위, 벌집위, 진위의 네 개 방으로 구성되어 있다. 반추위의 섬모충과 세균은 복잡한 셀룰로스를 발효하여 더 단순한 지방산으로 만들고, 이는 반추위 벽을 통해 흡수된다. 소과 동물은 긴 소장을 가지고 있으며, 소의 소장 길이는 29m에서 49m이다. 체온은 하루 동안 변동한다. 예를 들어, 염소는 아침에 37°C에서 오후에 40°C로 약간 변한다. 체온은 소의 경우 발한, 염소는 헐떡거림으로 조절된다. 4~5개 엽으로 구성된 오른쪽 폐는 3개 엽을 가진 왼쪽 폐보다 약 1.5배 크다.[3][25]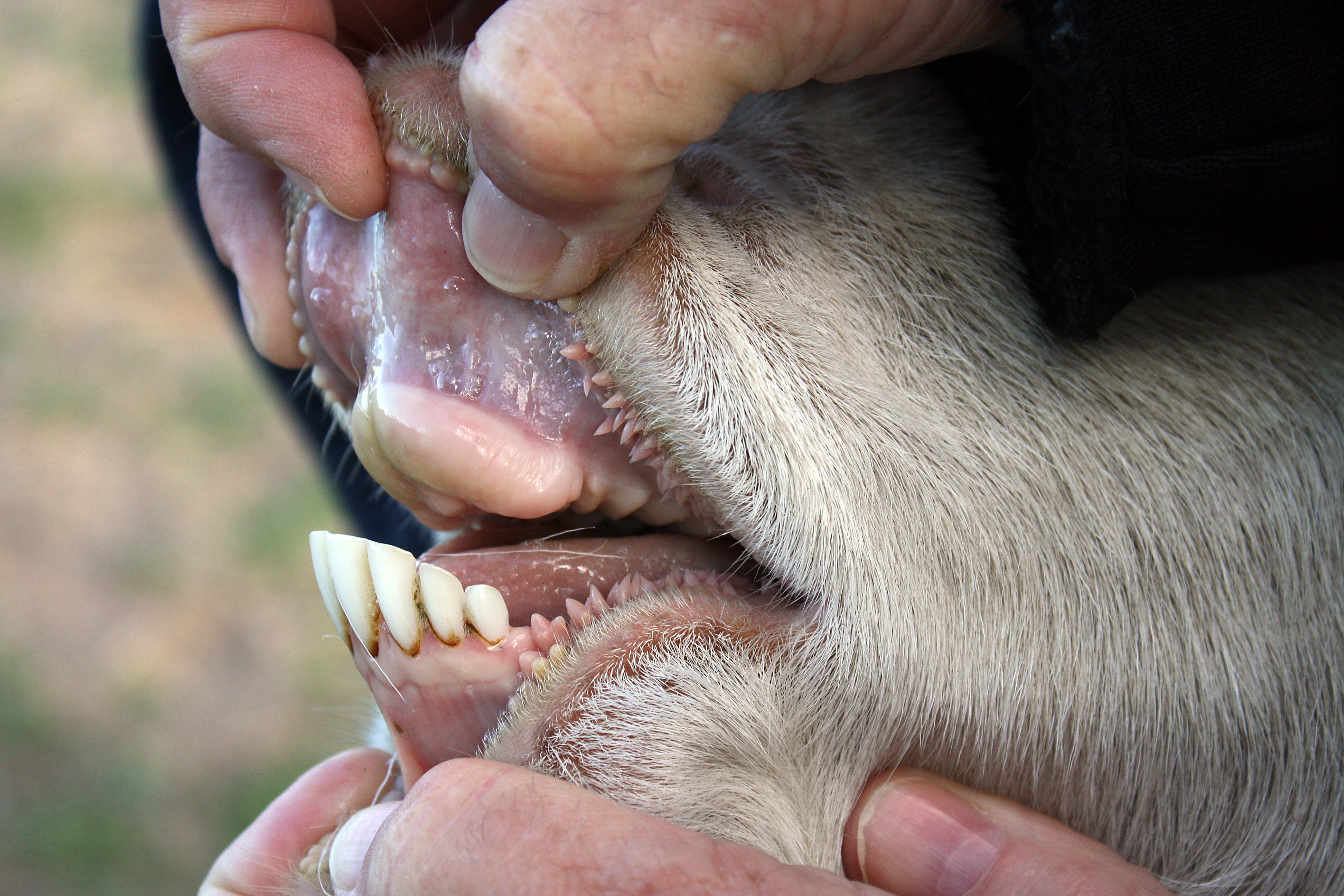
대부분의 소과 동물은 30~32개의 이빨을 가지고 있다.[31] 윗 앞니는 없지만, 윗 송곳니는 축소되었거나 없다. 윗 앞니 대신 두껍고 질긴 치상판을 가지고 있어 풀과 잎을 잡을 수 있다. 고치성이며 반달형인데, 어금니와 소구치가 낮고 초승달 모양 치첨을 가지기 때문이다. 아래 앞니와 송곳니는 앞으로 튀어나와 있다. 앞니 뒤에는 이개라고 알려진 긴 이빨 없는 틈이 있다.[48] 소과의 일반적인 치열은 0.0.2-3.33.1.3.3이다. 대부분 초식동물이지만, 대부분의 듀이커는 잡식동물이다. 다른 반추동물처럼 4개의 방으로 된 위를 가지고 있어 풀과 같은 식물 재료를 소화할 수 있다. 반추동물은 내장 미생물을 사용하여 발효를 통해 섬유소를 분해한다.[3] 소과에 공통적인 특징으로, 그 뿔은 영각이며, 각질의 뿔집과 골질의 심을 가지고 있다. 사슴과의 가지뿔처럼 골질부가 바뀌거나, 프로롱혼처럼 뿔집이 바뀌는 일은 없다.
측면의 발가락은 항상 불완전하며, 보통은 작은 발굽만 있고, 이 작은 발굽조차 가지지 않는 것도 있다.
4. 진화 역사

진화 초기에 소과는 Boodontia(유라시아 기원)와 Aegodontia(아프리카 기원)의 두 가지 주요 클레이드로 나뉘었다. Boodontia와 Aegodontia 사이의 이러한 초기 분화는 이러한 육지 사이의 대륙 분할로 여겨진다. 나중에 이러한 대륙이 다시 합쳐졌을 때 이 장벽이 제거되었고 두 그룹 모두 서로의 영토로 확장되었다.[16] Bovini와 Tragelaphini 부족은 초기 미오세에 분화되었다.[20] 소과는 베링 육교를 건너 플라이스토세에 아메리카에 도달한 것으로 알려져 있다.[21]
현재의 Alcelaphinae 속은 플리오세에 나타났다. 멸종된 Alcelaphine 속인 ''Paramularius''는 하테비스트와 같은 크기였으며 플리오세에 생겨났지만 플라이스토세 중기에 멸종된 것으로 여겨진다.[6] 여러 Hippotraginae 속이 플리오세와 플라이스토세부터 알려져 있다. 이 아과(subfamily)는 초기 미오세 후반에 Alcelaphinae에서 분화된 것으로 보인다.[20] Bovinae는 초기 미오세에 나머지 소과에서 분화된 것으로 여겨진다.[17] Boselaphini는 초기 플리오세에 아프리카에서 멸종되었다. 그들의 가장 최근 화석은 Langebaanweg(남아프리카)와 Lothagam(케냐)에서 발굴되었다.[18]
중기 마이오세는 소과의 중국과 인도 아대륙으로의 확산을 특징으로 한다.[21] Vrba에 따르면, 방산의 아과 알케라피나이는 중기 마이오세 후반에 시작되었다.[6] 염소아과는 아마도 초기 중기 마이오세에 분화되었을 것이다. 카프리니는 중기 마이오세에 등장했으며, 유라시아에서 다른 소과와 사슴과에 의해 대체된 것으로 보인다.[19] 영양아과의 가장 초기의 화석은 중기 마이오세에서 발견되었지만, 연구에 따르면 이 아과는 초기 마이오세부터 존재했다. 종분화는 중기 또는 후기 마이오세 동안, 주로 유라시아에서 족 영양족에서 일어났다. 네오트라기니족은 마이오세 말까지 아프리카에서 나타난 것으로 보이며, 플라이오세까지 널리 퍼지게 되었다.[20]
후기 마이오세, 약 1000만 년 전 무렵, 소과 동물은 빠르게 다양화를 이루어 70개의 새로운 속이 생겨났다.[21] 이러한 후기 마이오세 방산은 많은 소과 동물이 더 개방적인 초원 서식지에 적응하게 되었기 때문이기도 하다.[22] Aepycerotinae는 후기 마이오세에 처음 등장했으며, 원시 임팔라와 현대 임팔라의 크기에서 큰 차이점은 발견되지 않았다.[23] Caprinae의 일족인 ovibovine의 화석은 아프리카에서 후기 마이오세까지 거슬러 올라간다.[20] 가장 초기의 Hippotragine 화석은 후기 마이오세로 거슬러 올라가며, 로타감과 아와시 계곡과 같은 지역에서 발굴되었다.[20] Reduncinae의 최초 아프리카 화석은 600만 년에서 700만 년 전으로 거슬러 올라간다.[24] Reduncinae와 Peleinae는 중기 마이오세에 분화되었을 것이다.[6]
5. 하위 분류
6. 계통 분류
2003년 프랑스 국립 자연사 박물관의 알렉산드르 하사닌(Alexandre Hassanin)과 동료들은 미토콘드리아 및 핵 분석을 기반으로 계통발생학 연구를 진행했다. 이 연구에서 사향노루과와 소과는 사슴과와 자매인 분지군을 형성하며, 사슴과는 2700만에서 2800만 년 전에 소과-사향노루과 분지군으로부터 분기되었다는 사실이 밝혀졌다.[4]
분자 연구는 소과가 단계통군(한 조상 종과 모든 후손으로 구성된 생물군)임을 뒷받침한다.[5][6] 소과의 아과 수는 논쟁의 여지가 있지만, 분자, 형태학, 화석 증거는 8개의 뚜렷한 아과가 존재함을 나타낸다. 8개의 아과는 영양아과(임팔라만으로 구성), 누아과(본테복, 하테비스트, 누 및 친척), 영양아과(여러 영양, 가젤 및 친척), 소아과(소, 물소, 들소 및 기타 영양), 염소아과(염소, 양, 아이벡스, 세로우 및 친척), 듀이커아과(듀이커), 영양아과(아닥스, 오릭스 및 친척), 갈대영양아과(갈대영양 및 코브 영양)이다. 또한, 3개의 멸종된 아과도 알려져 있다.[7][8]
1992년, 런던 자연사 박물관의 앨런 W. 젠트리(Alan W. Gentry)는 소과의 8개 주요 아과를 진화적 역사를 기반으로 두 개의 주요 분지군으로 나누었다. 소과만 포함하는 Boodontia와 나머지 아과로 구성된 Aegodontia이다. Boodonts는 황소의 치아와 유사한 원시적인 치아를 가지며, aegodonts는 염소의 치아와 같은 더 발달된 치아를 가진다.[9]
''펠레아''와 ''Pantholops''를 각각 Peleinae와 Pantholopinae라는 아과로 인식하는 것에 대한 논쟁이 있다. 2000년, 미국의 생물학자 조지 셜러와 고생물학자 엘리자베스 브르바는 ''펠레아''를 갈대영양아과에 포함시키는 것을 제안했지만,[10] 회색리복은 ''펠레아''의 유일한 종으로 형태학적으로 코브와 갈대영양류와 매우 다르다.[11] ''Pantholops''는 이전에 영양아과로 분류되었지만, 나중에 자체 아과인 Pantholopinae에 속하게 되었다. 그러나 분자 및 형태학적 분석은 ''Pantholops''를 염소아과에 포함시키는 것을 지지한다.[12]
소과는 Boodontia(소이류)와 Aegedontia(아이게 치류)로 나뉜다.[85] Boodontia는 아시아에서, Aegedontia는 아프리카에서 진화했다. 소과는 12족으로 분류되며,[85][86][83] Boodontia에는 소아과의 3족이 속하고, Aegedontia에는 나머지 9족이 속한다. Aegedontia의 아과 분류는 일정하지 않아 1개[86][83] 또는 6개[87]에서 8개[84]의 아과로 분류되기도 하지만, 여기서는 7개의 아과(소과 전체에서는 8개의 아과)로 나누는 것으로 한다.[81] 단, 이 분류에서 블랙벅아과(블랙벅족 + 클립스프링어족 + 로열앤틸로프족)는 다계통이다.[85][86][83][84]
소과 내에서는 먼저 소아과가 분기되고, 다음으로 (임팔라아과 + 로열앤틸로프족)이 분기되었다.[83][84] 그 후의 분기는 불확실하다. 소과의 정확한 자매군은 불확실하지만, 소과 등 5과로 구성된 진반추하목은 단일 계통이다.[83]
6. 1. 2009년 계통 분류
2009년 스폴딩(Spaulding) 등의 연구에 기초한 소과의 계통 분류는 다음과 같다.[91]6. 2. 2019년 계통 분류
다음은 2019년 주라노(Zurano) 등의 연구에 기초한 소과의 계통 분류이다.[92]{| class="wikitable"
|-
! 소과
|-
|
{| class="wikitable"
|-
! 소아과
|-
|
{| class="wikitable"
|-
! 닐가이영양족
|-
|
|-
! 트라겔라푸스족
|-
|
{| class="wikitable"
|-
| 암멜라푸스속
|-
|
{| class="wikitable"
|-
| 니알라영양속
|-
|
{| class="wikitable"
|-
|
|-
| 트라겔라푸스속
|}
|}
|}
|-
! 소족
|-
|
{| class="wikitable"
|-
| 사올라속
|-
|
{| class="wikitable"
|-
|
7. 생태와 행동
대부분의 야생 소과 동물은 10~15년 동안 산다. 더 큰 종은 더 오래 사는 경향이 있는데,[3] 예를 들어 아메리카들소는 최대 25년, 가우르는 최대 30년까지 살 수 있다. 사육되는 개체의 평균 수명은 거의 10년이다. 예를 들어, 사육되는 염소는 평균 수명이 12년이다.
일반적으로 수컷은, 특히 일부다처제 종에서, 암컷보다 수명이 짧다. 이는 어린 수컷의 조기 분산, 공격적인 수컷 간의 싸움, 포식에 대한 취약성(특히 쿠두와 같이 수컷이 덜 민첩할 때), 영양 부족(큰 몸집 때문에 수컷은 높은 영양 요구량을 가지며 충족되지 않을 수 있음) 등 여러 가지 이유로 설명될 수 있다.[65][66] 리처드 데스파드 에스테스는 암컷이 우세한 수컷으로부터 수컷 새끼를 보호하기 위해 뿔과 같은 수컷의 2차 성징을 모방한다고 제안했다. 이 특징은 공격적인 수컷의 공격과 청소년기 동안 어린 수컷의 강제 분산으로 인한 수컷 사망률과 불균형한 성비를 방지하기 위해 강력하게 선택된 것으로 보인다.[67]
7. 1. 사회적 행동
블랙벅이나 가이알과 같은 소과 동물들은 단독 행동과 군집 행동, 영역 행동과 비영역 행동 등 다양한 사회적 조직과 사회적 행동 방식을 보인다.[31]클립스프링어, 오리비, 스틴복과 같은 작은 소과 동물들은 일반적으로 단독 생활을 하며 영역을 형성한다. 이들은 다른 종의 구성원이 들어갈 수 없는 작은 영역을 소유하고, 일부일처 쌍을 이룬다. 딕딕과 같은 많은 종들은 눈물샘에서 페로몬을 분비하고, 때로는 배설물을 사용하여 영역을 표시한다.[49] 새끼들은 청소년기에 흩어지며, 수컷은 짝짓기를 하기 전에 영역을 확보해야 한다.[3] 부시벅은 단독 생활을 하면서 영역을 갖지 않는 유일한 소과 동물이다. 이 영양은 거의 공격성을 보이지 않으며, 스스로 고립되거나 느슨한 무리를 형성하는 경향이 있지만, 유리한 서식지에서는 여러 부시벅이 서로 매우 가까이 발견될 수 있다.[50]
케팔로피네아과(듀이커), 트라겔라피네아과(나선형 뿔 영양) 및 네오트라기네아과를 제외한 대부분의 아프리카 소과 동물들은 군집 생활을 하며 영역을 형성한다. 수컷은 성적으로 성숙해지면 흩어져서 자신의 영역을 형성해야 하는 반면, 암컷은 그렇게 할 필요가 없다. 영역을 확보하지 못한 수컷들은 수컷 무리를 형성한다. 수컷들 사이에서는 지배권을 얻기 위한 경쟁이 벌어지며, 싸움은 제한된 발정기에 더 격렬해지는 경향이 있다. 이동하는 수컷을 제외하고, 수컷들은 일반적으로 평생 동안 같은 영역을 유지한다.[31] 워터벅에서는 "위성 수컷"으로 알려진 일부 수컷 개체들이 다른 수컷의 영역에 들어갈 수 있으며, 소유자가 늙을 때까지 기다려 그의 영역을 얻어야 할 수도 있다.[51] 수컷들이 모여 경쟁적으로 잠재적 짝에게 과시하는 렉 짝짓기는 토피, 코브, 레체에서 존재하는 것으로 알려져 있다.[52] 트라겔라피네아과, 소, 양, 염소는 군집 생활을 하며 영역을 형성하지 않는다. 이러한 종에서는 수컷이 다른 모든 수컷보다 절대적인 지배력을 얻어야 하며, 싸움은 영역에 국한되지 않는다. 따라서 수컷은 신체 성장에 여러 해를 소비한다.[31]
7. 2. 활동
대부분의 소과 동물은 주행성이지만, 물소, 부시벅, 리드벅, 그림스복 등 일부는 예외이다. 사회적 활동과 먹이 활동은 보통 새벽과 황혼 무렵에 절정에 달한다. 소과 동물들은 보통 새벽, 정오 무렵, 그리고 해가 진 후에 휴식을 취한다. 그루밍은 보통 혀로 핥는 방식으로 이루어진다. 사슴과 동물은 진흙이나 먼지 속에서 구르는 경우가 드물다. 누와 물소는 보통 진흙에서 뒹구는 반면, 하테비스트와 토피는 머리와 뿔을 진흙에 비빈 다음 몸 전체에 바른다. 소과 동물은 다양한 형태의 음성적, 후각적, 그리고 유형적 의사소통을 사용한다. 여기에는 성적 흥분, 감정 상태 또는 경고를 전달하기 위해 목, 머리, 뿔, 털, 다리, 귀의 다양한 자세가 포함된다. 그러한 표현 중 하나가 플레멘 반응이다. 소과 동물은 위험을 감지하면 머리를 높이 들고 의도적인 시선으로 움직이지 않고 서 있는 경향이 있다. 임팔라, 쿠두, 엘란과 같은 일부 종은 몇 피트 높이까지 뛰어오를 수도 있다.[31] 소과 동물은 다른 동물에게 경고하고 포식자를 쫓아내기 위해 포효하거나 으르렁거릴 수 있다.[3] 가젤과 같은 소과 동물은 포식자에 반응하여 스토팅을 하며, 다리를 꼿꼿하게 세우고 높이 뛰어올라, 포식자를 보았고 스토팅을 하는 개체가 강하며 쫓을 가치가 없다는 것을 정직하게 신호한다.[53]
짝짓기철에는 발정기 수컷이 암컷에게 자신의 존재를 알리기 위해 울부짖는다. 사향소는 수컷 간의 싸움 중에 포효하며, 수컷 사이가는 코를 통해 공기를 내뿜어 경쟁 수컷을 위협하고 암컷을 유혹하는 포효를 낸다. 어미는 또한 새끼가 떨어졌을 때 새끼를 찾기 위해 음성적 의사소통을 사용한다. 지배권을 위한 싸움 동안 수컷은 꼿꼿한 자세로 코를 수평으로 유지하며 자신을 과시하는 경향이 있다.[55][54]
싸움 기술은 소과 동물 가족에 따라 다르며, 체형에 따라 다르기도 하다. 하테비스트는 무릎을 꿇고 싸우는 반면, 다른 종은 보통 네 발로 싸운다. 다양한 크기의 가젤은 서로 다른 전투 방식을 사용한다. 가젤은 보통 복싱을 하며, 심각한 싸움에서는 짧은 거리에서 강한 타격을 주고받는 충돌과 펜싱을 할 수 있다. 아이벡스, 염소, 양 수컷은 똑바로 서서 아래로 서로 충돌한다. 누는 공격적인 충돌에서 강력한 머리 박치기를 사용한다. 뿔이 얽히면 상대방은 뿔을 풀기 위해 원을 그리며 움직인다. 사향소는 빠른 속도로 서로 돌진한다. 일반적으로, 동일한 체형과 방어 수준을 가진 두 소과 동물만이 싸움에 참여하며, 이는 두 개체 중 우위를 결정하기 위한 것이다. 다른 개체보다 명백히 열등한 개체는 싸우기보다는 도망가는 경향이 있다. 예를 들어, 미성숙한 수컷은 성숙한 수컷과 싸우지 않는다. 일반적으로 소과 동물은 몸보다는 상대방의 머리를 공격한다. 임팔라의 뿔과 같은 S자형 뿔은 치기, 잡기, 찌르기에 도움이 되는 다양한 부분이 있다. 부상으로 이어지는 심각한 싸움은 드물다.[31][55][56]
7. 3. 번식
대부분의 소과 동물은 일부다처제이다. 일부 종은 일부일처제를 보여 수컷 간의 공격성이 적고, 수컷의 큰 몸집에 대한 선택 압력이 줄어들어 성적 이형성이 거의 나타나지 않는다. 암컷이 수컷보다 약간 더 큰 경우가 있는데, 이는 암컷 간의 영역 획득 경쟁 때문일 수 있다. 이는 사슴과 다른 작은 소과 동물에서 나타난다.[68][64] 암컷과 수컷 모두 성적 성숙에 도달하는 데 걸리는 시간은 소과 동물 사이에서 다양하다. 예를 들어, 임팔라 수컷은 1살이 되면 성적으로 성숙하지만, 4살이 되어서야 짝짓기를 할 수 있다.[59] 반대로 바바리양 암컷은 성적 성숙을 하기 전에도 새끼를 낳을 수 있다.[60] 수컷의 성적 성숙 지연은 성적 이형성이 나타나는 종, 특히 레둔친에서 더 두드러지게 나타나는데, 이는 수컷 간의 경쟁 때문일 것이다.[3] 예를 들어, 누 암컷은 태어난 지 1~2년 이내에 번식 능력을 갖게 되지만, 수컷은 4살이 되어서야 성숙해진다.[30]
모든 소과 동물은 적어도 1년에 한 번 짝짓기를 하며, 작은 종은 두 번 짝짓기를 하기도 한다. 대부분 소과 동물의 짝짓기 시기는 보통 우기이며, 따라서 적도 지역에서는 번식이 두 번 최고조에 달할 수 있다. 양과 염소는 번식의 계절성이 뚜렷한데, 일년 중 일일 광주기의 주기가 결정적인 역할을 한다. 이 주기에 영향을 미치는 다른 요인으로는 주변 온도, 영양 상태, 사회적 상호 작용, 분만 날짜 및 수유 기간이 있다. 염소와 양은 단일 번식 동물이다. 대부분의 양 품종에서 짝짓기는 여름 또는 초가을에 시작된다.[61] 양의 짝짓기는 번식기 시작을 앞당기는 멜라토닌;[62]과 번식기를 종료시키는 티록신의 영향을 받는다.[63] 발정은 소과 동물에서 며칠 이상 지속되지 않으며, 소와 트라겔라핀을 제외한다. 하테비스트와 토피를 제외하고, 모든 소과 동물은 서골 기관을 사용하여 소변을 검사하여 암컷의 발정을 감지한다.[31] 수컷은 암컷이 발정 상태임을 확인하면 구애 행동을 시작한다. 처음에는 받아들이지 않던 암컷은 결국 다른 수컷보다 우위를 차지한 수컷과 짝짓기를 한다. 교미는 보통 몇 초밖에 걸리지 않는다.[31][68]
임신 기간은 소과 동물에 따라 다르다. 사슴의 임신 기간은 120~150일인 반면, 아프리카물소의 임신 기간은 300~330일이다. 보통 새끼 한 마리가 태어나며 (쌍둥이는 드물다), 태어난 지 한 시간 이내에 스스로 서서 달릴 수 있다. 일부일처제 종에서는 수컷이 새끼를 방어하는 데 도움을 주지만, 일부다처제 종에서는 그렇지 않다. 대부분의 신생 송아지는 1주에서 2개월 동안 숨어 지내며, 어미에게 정기적으로 젖을 먹는다. 일부 소과 동물 종에서는, 임팔라처럼, 신생아가 즉시 또는 며칠 이내에 어미를 따라다니기 시작한다.[59] 소과 동물은 새끼를 방어하기 위해 서로 다른 전략을 가지고 있다. 예를 들어, 누 어미는 자신의 새끼만 방어하는 반면, 물소는 집단 방어를 보인다. 이유 시기는 2개월(로열안텔로프) 또는 1년(사향소)이 될 수 있다.[68][64]
8. 분포
대부분의 야생 소과 동물은 10~15년 동안 산다. 더 큰 종은 더 오래 사는 경향이 있다.[3] 예를 들어, 아메리카들소는 최대 25년까지 살 수 있으며, 가우르는 최대 30년까지 산다. 사육되는 개체의 평균 수명은 거의 10년이다. 예를 들어, 사육되는 염소는 평균 수명이 12년이다.
일반적으로 수컷은, 특히 일부다처제 종에서, 암컷보다 수명이 짧다. 이는 여러 가지 이유로 설명될 수 있다. 어린 수컷의 조기 분산, 공격적인 수컷 간의 싸움, 포식에 대한 취약성(특히 쿠두와 같이 수컷이 덜 민첩할 때), 그리고 영양 부족(크기가 크기 때문에 수컷의 몸은 높은 영양 요구량을 가지며 충족되지 않을 수 있음) 등이 그 이유이다.[65][66] 리처드 데스파드 에스테스는 암컷이 우세한 수컷으로부터 수컷 새끼를 보호하기 위해 뿔과 같은 수컷의 2차 성징을 모방한다고 제안했다. 이 특징은 공격적인 수컷의 공격과 청소년기 동안 어린 수컷의 강제 분산으로 인한 수컷 사망률과 불균형한 성비를 방지하기 위해 강력하게 선택된 것으로 보인다.[67]
9. 인간과의 관계
소과는 인간에게 다양한 방식으로 이용되어 왔다. 젖소에게서는 우유, 버터, 기, 요거트, 버터밀크, 치즈와 같은 유제품을 얻는다. 특히 물소 젖은 이탈리아에서 모짜렐라 치즈를, 인도에서는 굴랍 자문 디저트를 만드는 데 사용되며,[70] 양젖은 프랑스에서 푸른색 로크포르 치즈를 만드는 데 사용된다.[71]
쇠고기는 아연, 셀레늄, 인, 철, 비타민 B가 풍부한 식품이다.[72] 들소 고기는 쇠고기보다 지방과 콜레스테롤 함량이 낮지만, 단백질 함량은 더 높다.[73]
소과 가죽은 질기고 튼튼하며, 부드러운 의류 가죽에서 단단한 신발 가죽에 이르기까지 다양한 두께로 만들 수 있다. 염소 가죽과 소 가죽은 널리 사용되는 반면, 양가죽은 의류 용도로만 적합하다.[74] 메리노 어린 양의 양털은 매우 가늘고 부드러워 가장 가치가 높다. 굵은 양털은 내구성이 뛰어나고 보풀에 강하여 튼튼한 의류[75]와 카펫을 만드는 데 사용된다.
골분은 칼슘, 인, 질소가 풍부한 중요한 비료이며, 토양 산도를 제거하는 데 효과적이다.[76] 소과 뿔은 고대부터 마시는 용기로 사용되어 왔다.[77]
9. 1. 가축화
소과의 다양한 종들은 대부분 아프리카에 서식한다. 최대 밀집 지역은 동아프리카의 사바나이다. 먹이 습성에 따라 여러 종들이 광대한 지역으로 적응 방산하여 뻗어나갔으며, 이로 인해 치아 및 사지 형태에서 다양한 변이가 관찰된다. 듀이커는 열대 우림에, 시타퉁가와 레체는 늪지 근처에, 일런드는 초원에, 스프링복과 오릭스는 사막에, 봉고와 아노아는 울창한 숲에, 산양과 타킨은 고산 지대에 서식한다.[31] 소과 종 중 일부는 유럽, 아시아, 북미에서도 발견된다. 양과 염소는 주로 유라시아에서 발견되지만, 바바리양과 아이벡스는 아프리카 동물군의 일부를 구성한다. 사향소는 북극 툰드라에 국한되어 서식한다. 여러 소과 종들은 인류에 의해 가축화되었다. 염소와 양의 가축화는 1만 년 전에 시작되었으며, 소는 약 7,500년 전에 가축화되었다.[3][68]9. 2. 문화
소과의 가축화는 인류가 수렵과 채집 생활에서 농업으로 전환하는 데 큰 영향을 주었다. 소과 동물 중에는 전 세계적으로 널리 퍼진 소, 양, 염소가 있는데, 이들은 모두 유라시아 지역이 원산지이다.[68] 물소(야생 물소에서 유래), 야크(야생 야크에서 유래), 제부(인도 오록스에서 유래), 가이알(가우어에서 유래), 발리 소(반텡에서 유래) 등은 가축화되었지만 널리 퍼지지는 않았다.[68]오릭스, 아닥스, 엘란드 및 멸종된 부발 하테비스트와 같은 일부 영양도 가축화되었다. 고대 이집트 벽화에는 오릭스, 아닥스, 부발 하테비스트가 묘사되어 있다.

소의 가축화에 대한 초기 증거는 기원전 8000년경으로 거슬러 올라가며, 키프로스와 유프라테스 분지에서 시작되었음을 보여준다.[69]
소과는 적어도 기원전 600년경 고대 그리스의 이솝 우화 시대부터 이야기에 등장했다. 이솝 우화에는 '까마귀와 양', '개구리와 황소', '늑대와 어린 양' 등이 있다.[78] 사자, 등에서 솟아오른 염소 머리, 뱀 머리로 끝나는 꼬리를 가진 신화 속 동물 키마이라는 티폰과 에키드나의 자손 중 하나이며, 케르베로스 및 레르네의 히드라와 같은 괴물의 형제였다.[79] 중국 신화에서 염소자리와 같은 의미인 양은 중국 십이지의 여덟 번째 동물이며, 효의 상징이다.[80]
참조
[1]
MSW3
[2]
웹사이트
Bovidae
http://www.merriam-w[...]
2014-10-07
[3]
웹사이트
Bovidae: antelopes, cattle, gazelles, goats, sheep, and relatives
http://animaldiversi[...]
University of Michigan Museum of Zoology
2014-10-07
[4]
논문
Molecular and morphological phylogenies of Ruminantia and the alternative position of the Moschidae
https://www.research[...]
2003
[5]
논문
A cladistic analysis of mitochondrial ribosomal DNA from the Bovidae
[6]
논문
A complete estimate of the phylogenetic relationships in Ruminantia: a dated species-level supertree of the extant ruminants
2005
[7]
서적
Paleontology and Geology of Laetoli Human Evolution in Context
Springer
2011
[8]
논문
First evidence of ''Tethytragus'' Azanza and Morales, 1994 (Ruminantia, Bovidae), in the Miocene of the Vallès-Penedès Basin (Spain)
[9]
서적
Neogene Paleontology of the Manonga Valley, Tanzania: A Window into the Evolutionary History of East Africa
Plenum Press
1997
[10]
서적
Antelopes, Deer, and Relatives: Fossil Record, Behavioral Ecology, Systematics, and Conservation
Yale University Press
2000
[11]
MSW3
[12]
MSW3
[13]
논문
Phylogenetic analyses and improved resolution of the family Bovidae based on complete mitochondrial genomes
https://www.scienced[...]
2013-06-01
[14]
논문
CytochromebPhylogeny of the Family Bovidae: Resolution within the Alcelaphini, Antilopini, Neotragini, and Tragelaphini
https://www.scienced[...]
1999-06-01
[15]
논문
Total Evidence Phylogenetic Analysis Supports New Morphological Synapomorphies for Bovidae (Mammalia, Artiodactyla)
https://bioone.org/j[...]
2021-06
[16]
논문
The tribal radiation of the family Bovidae (Artiodactyla) and the evolution of the mitochondrial cytochrome ''b'' gene
http://www.isem.cnrs[...]
[17]
논문
A multi-calibrated mitochondrial phylogeny of extant Bovidae (Artiodactyla, Ruminantia) and the importance of the fossil record to systematics
2013-08
[18]
논문
A new caprin bovid (Mammalia) from the late Miocene of Morocco
2012
[19]
서적
East African Mammals: An Atlas of Evolution in Africa (Volume III, Part C)
University of Chicago press
1989
[20]
서적
Homo Erectus: Pleistocene Evidence from the Middle Awash, Ethiopia
University of California Press
2008
[21]
서적
Horns, Tusks, and Flippers: the Evolution of Hoofed Mammals
Johns Hopkins University Press
2002
[22]
서적
Mammal Evolution: an illustrated guide
https://archive.org/[...]
Facts on File
[23]
서적
Living Fossils
Springer
1984
[24]
서적
Paleoclimate and Evolution, with Emphasis on Human Origins
Yale University Press
1995
[25]
서적
Fauna of Australia (Volume 1B)
Australian Government Publication Service
1989
[26]
서적
The Genetics of Cattle
Cabi
2015
[27]
웹사이트
Royal antelope
http://www.ultimateu[...]
2014-10-08
[28]
서적
Endangered Wildlife and Plants of the World
Marshall Cavendish
2001
[29]
EOL
Oryx leucoryx
[30]
웹사이트
"''Connochaetes gnou'': black wildebeest"
http://animaldiversi[...]
University of Michigan
2013-08-21
[31]
서적
The Behavior Guide to African Mammals: Including Hoofed Mammals, Carnivores, Primates
https://archive.org/[...]
University of California Press
2004
[32]
논문
"''Addax nasomaculatus''"
[33]
서적
Adaptive Coloration in Animals
Methuen
[34]
논문
Countershading: Universally deceptive or deceptively universal?
1998-01
[35]
서적
Adaptive Coloration in Animals
Methuen
[36]
논문
Contrasting colouration in terrestrial mammals
2009
[37]
논문
Tetracerus quadricornis (Artiodactyla: Bovidae)
2009-09-25
[38]
서적
Exotic Animal Field Guide: Nonnative Hoofed Mammals in the United States
Texas A&M University Press
2007
[39]
웹사이트
Jacob Sheep
http://www.albc-usa.[...]
American Livestock Breeds Conservancy
2011-05-05
[40]
논문
The fossil record and evolution of Bovidae: state of the field
http://palaeo-electr[...]
2010-11-14
[41]
논문
Phylogeny of the Bovidae (Artiodactyla, Mammalia), based on mitochondrial ribosomal DNA sequences
[42]
논문
Morphology of horns and fighting behavior in the family Bovidae
[43]
논문
Correlates of horn and antler shape in bovids and cervids
2003
[44]
논문
The intensity of sexual selection predicts weapon size in male bovids
[45]
Q
Q55899859
[46]
논문
Evolution of weaponry in female bovids
[47]
논문
Sexual Dimorphism: The Horns of African Antelopes
1983
[48]
서적
The Encyclopedia of Mammals
https://archive.org/[...]
Facts on File
[49]
서적
Pheromones and Animal Behaviour: Communication by Smell and Taste
https://archive.org/[...]
Cambridge University Press
2003
[50]
웹사이트
Bushbuck
http://animaldiversi[...]
University of Michigan Museum of Zoology
2014-10-28
[51]
웹사이트
Waterbuck
http://animaldiversi[...]
University of Michigan Museum of Zoology
2014-10-28
[52]
서적
Intraspecific Variation in the Social Systems of Wild Vertebrates
https://archive.org/[...]
Cambridge University Press
[53]
논문
Observations on the behaviour and feeding habits of the springbok ''Antidorcas marsupialis''
[54]
서적
Mammalogy
Jones and Bartlett Publishers
2011
[55]
서적
Grzimek's Encyclopedia of Mammals (Volume 5)
https://archive.org/[...]
McGraw-Hill Publishing
1990
[56]
논문
Climate change reduces reproductive success of an Arctic herbivore through trophic mismatch
2008-07
[57]
논문
Dietary preferences in extant African Bovidae
2000-05
[58]
논문
Diets of Southern African Bovidae: Stable Isotope Evidence
2003
[59]
서적
The Behavior Guide to African Mammals: Including Hoofed Mammals, Carnivores, Primates
https://archive.org/[...]
University of California Press
2004
[60]
논문
Ammotragus lervia
[61]
논문
Seasonality of reproduction in sheep
2003
[62]
논문
Photoperiodic and melatonin treatments for the control of seasonal reproduction in sheep and goats
1988
[63]
논문
Hypothalamic gene expression in reproductively photoresponsive and photorefractory Siberian hamsters
2002
[64]
서적
Behavioural Ecology: An Evolutionary Approach
Wiley-Blackwell
1997
[65]
논문
Comparative mortality rates of male and female kudus: the costs of sexual size dimorphism
1993
[66]
논문
Causes of Sex-Biased Adult Survival in Ungulates: Sexual Size Dimorphism, Mating Tactic or Environment Harshness?
2003
[67]
논문
The significance of horns and other male secondary sexual characters in female bovids
1991
[68]
서적
Mammalogy: Adaptation, Diversity, Ecology
https://books.google[...]
Johns Hopkins University Press
2016-11-07
[69]
서적
Documenting Domestication: New Genetic and Archaeological Paradigms
University of California Press
2006
[70]
웹사이트
Others' Milk
http://www.slate.com[...]
Slate.com
2013-07-24
[71]
서적
Gastronomie!: Food Museums and Heritage Sites of France
Bunker Hill Publishing
[72]
웹사이트
Beef, lean organic
http://whfoods.org/g[...]
WHFoods
2015-04-01
[73]
웹사이트
National Bison Association
http://www.bisoncent[...]
Bisoncentral.com
2015-04-01
[74]
서적
Why Leather?: The Material and Cultural Dimensions of Leather
Sidestone Press
2014
[75]
웹사이트
Merino Sheep in Australia
http://www.merinos.c[...]
2015-04-01
[76]
서적
Manures and fertilizers
Atlantic Publications
2007
[77]
학술지
Germanic Glass Drinking Horns
1975
[78]
웹사이트
Aesop's Fables
http://www.aesops-fa[...]
2014-10-10
[79]
웹사이트
Entry:Chimaera
https://www.perseus.[...]
2015-03-31
[80]
서적
A Dictionary of Chinese Symbols: Hidden Symbols in Chinese Life and Thought
https://archive.org/[...]
Routledge
[81]
문서
Order Artiodactyla
https://www.departme[...]
Johns Hopkins University Press
[82]
문서
世界哺乳類標準和名目録
https://doi.org/10.1[...]
日本哺乳類学会
[83]
간행물
Pattern and timing of diversification of Cetartiodactyla (Mammalia, Laurasiatheria), as revealed by a comprehensive analysis of mitochondrial genomes
http://www.sciencedi[...]
[84]
간행물
Phylogenetic analyses and improved resolution of the family Bovidae based on complete mitochondrial genomes
http://www.sciencedi[...]
[85]
간행물
A complete estimate of the phylogenetic
http://www.pmmv.com.[...]
[86]
간행물
SuperTRI: A new approach based on branch support analyses of multiple independent data sets for assessing reliability of phylogenetic inferences
http://www.normalesu[...]
[87]
서적
新図説 動物の起源と進化 書き換えられた系統樹
八坂書房
[88]
문서
ウシ科の分類
東京動物園協会
[89]
웹사이트
レイヨウ
平凡社
2024-12-23
[90]
웹사이트
FAMILY Bovidae
MSW3
[91]
문서
Relationships of Cetacea (Artiodactyla) Among Mammals: Increased Taxon Sampling Alters Interpretations of Key Fossils and Character Evolution.
[92]
문서
Cetartiodactyla: Updating a time-calibrated molecular phylogeny.
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com