무족영원목
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
무족영원목은 사지 없이 가늘고 긴 원통형 몸체를 가진 양서류로, 굴을 파는 생활 방식에 적합한 해부학적 특징을 지닌다. 대부분 폐를 가지고 있지만, 피부나 입으로도 산소를 흡수하며, 촉수를 통해 냄새를 감지한다. 전 세계 열대 지역에 분포하며, 10개의 과로 분류된다. 인간과의 관계에서는 일부 문화에서 혐오스럽거나 독성이 있는 존재로 여겨지기도 한다.
더 읽어볼만한 페이지
- 무족영원목 - 에오카에킬리아
- 요하네스 페터 뮐러가 명명한 분류군 - 별불가사리
별불가사리(Patiria pectinifera)는 동북아시아 해역에 분포하며 얕은 조간대에서 서식하고, 조류 등을 섭취하며, 난모세포 연구에 사용되는 모델 생물이다. - 요하네스 페터 뮐러가 명명한 분류군 - 황소상어
황소상어는 전 세계 열대, 아열대 해역, 기수역, 담수에서 서식하며, 4m까지 자라는 크고 뚱뚱한 체형에 짧고 둥근 주둥이를 가진 상어로, 뛰어난 삼투압 조절 능력으로 담수 생존이 가능하고 공격적인 성향을 보이며, 어획과 서식지 파괴로 IUCN 적색 목록에 취약종으로 등재되었다. - 표시 이름과 문서 제목이 같은 위키공용분류 - 라우토카
라우토카는 피지 비치레부섬 서부에 위치한 피지에서 두 번째로 큰 도시이자 서부 지방의 행정 중심지로, 사탕수수 산업이 발달하여 "설탕 도시"로 알려져 있으며, 인도에서 온 계약 노동자들의 거주와 미 해군 기지 건설의 역사를 가지고 있고, 피지 산업 생산의 상당 부분을 담당하는 주요 기관들이 위치해 있다. - 표시 이름과 문서 제목이 같은 위키공용분류 - 코코넛
코코넛은 코코넛 야자나무의 열매로 식용 및 유지로 사용되며, 조리되지 않은 과육은 100g당 354kcal의 열량을 내는 다양한 영양 성분으로 구성되어 있고, 코코넛 파우더의 식이섬유는 대부분 불용성 식이섬유인 셀룰로오스이며, 태국 일부 지역에서는 코코넛 수확에 훈련된 원숭이를 이용하는 동물 학대 문제가 있다.
| 무족영원목 - [생물]에 관한 문서 | |
|---|---|
| 개요 | |
![''[[Oscaecilia ochrocephala]]'' ([[Caeciliidae]])](https://cdn.onul.works/wiki/noimage.png) | |
| 학명 | Gymnophiona Müller, 1832 |
| 이명 | Gymnophia Rafinesque, 1814 |
| 어원 | 그리스어 gymnos(벌거벗은) + ophis(뱀) |
| 분포 | 열대 지역 |
| 하위 분류 | |
| 멸종된 속 | †Eocaecilia †Funcusvermis †Rubricacaecilia †Chinlestegophis? |
| 아포다 분기군 | Apoda Oppel, 1811 |
| 현존하는 과 | Caeciliidae (무족영원과) Chikilidae (치킬리과) Dermophiidae (피부무족영원과) Herpelidae (헤르펠스과) Ichthyophiidae (물고기무족영원과) Grandisoniidae (그랜디소니과) Rhinatrematidae (코무족영원과) Scolecomorphidae (스코레코모르푸스과) Siphonopidae (사이폰무족영원과) Typhlonectidae (수생무족영원과) |
| 생물학적 분류 | |
| 계 | 동물계 |
| 문 | 척삭동물문 |
| 아문 | 척추동물아문 |
| 강 | 양서강 |
| 아강 | 평활양서아강 |
| 목 | 무족영원목 (Gymnophiona) |
2. 형태 및 해부학적 특징
무족영원목은 겉모습이 지렁이나 뱀과 비슷하며, 다리가 없는 것이 특징이다. 몸에는 많은 주름이 있어 마디가 있는 것처럼 보인다.
무족영원목의 눈은 작거나 아예 없으며, 피부 아래에 묻혀 있는 경우도 있다. 시력은 빛과 어둠을 구별하는 정도로 제한되어 있다.[13][14] 머리 양쪽에는 눈과 콧구멍 사이에 촉수라는 한 쌍의 감각 기관이 있는데, 이는 후각 기능을 돕는 것으로 보인다.[11]
피부는 매끄럽고 어두운 색을 띠는 경우가 많지만, 일부 종은 화려한 색을 띠기도 한다. 피부 속에는 방해석 성분의 비늘이 있는데, 이는 Scolecomorphidae과와 Typhlonectidae과에는 없지만, ''Typhlonectes compressicauda''의 경우 몸 뒤쪽에서 미세한 비늘이 발견되었다.[15][21] 피부에는 독을 분비하는 샘이 있어 포식자로부터 몸을 보호한다.[11] ''Siphonops paulensis''의 피부 분비물은 용혈성 특성을 갖는 것으로 밝혀졌다.[22]
무족영원목의 두개골은 단단하며, 뼈 사이의 틈이 거의 없다. 주둥이는 뾰족해서 흙을 파고 들어가는 데 유리하다.[15] 입은 머리 아래쪽에 위치하며, 위턱에는 2열, 아래턱에는 1~2열의 이빨이 있다. 이빨은 먹이를 물고 놓치지 않는 데 사용된다. 턱을 닫는 근육은 두 쌍으로, 다른 양서류보다 발달되어 있어 턱을 강하게 닫을 수 있다.[11][20] 아래턱은 앞쪽에 이빨이 있는 의치와 뒤쪽에 턱 관절과 근육 부착부가 있는 의각으로 구성된다. 최대 20개의 이빨이 있는 추가 삽입 이빨 열은 턱의 주요 가장자리 이빨 열과 평행하게 놓여 있다.[30]
환무족영원목 (''Siphonops annulatus'')은 일부 뱀독과 도마뱀의 상동일 수 있는 치아선을 가지고 있다. 이 선의 기능은 알려져 있지 않다.[18]
중이는 등골뼈와 난원창으로만 구성되어 있으며, 이는 일부 파충류에서 볼 수 있는 재진입 유체 회로를 통해 진동을 내이로 전달한다. Scolecomorphidae과 성체는 등골과 난원창이 모두 없어 중이 장치의 모든 구성 요소가 없는 유일한 양서류이다.[19]
2. 1. 몸 구조
무족영원목의 해부학적 구조는 굴을 파는 생활 방식에 매우 적합하다. 원시적인 속인 ''Ichthyophis''에 속하는 몇몇 종에서는 퇴화된 사지의 흔적이 발견되었으며, ''Typhlonectes compressicauda''에서는 배아 발달 과정에서 사지 싹의 존재가 관찰되었다. 이들은 다른 부분에서는 완전히 사지가 없는 몸에 잔존해 있다.[4] 이로 인해 작은 종들은 벌레와 비슷하고, 최대 1.5m 길이의 ''Caecilia thompsoni''와 같은 대형 종은 뱀과 유사하다. 꼬리는 짧거나 없고, 총배설강은 몸의 끝 부분 근처에 있다.[5][6][15]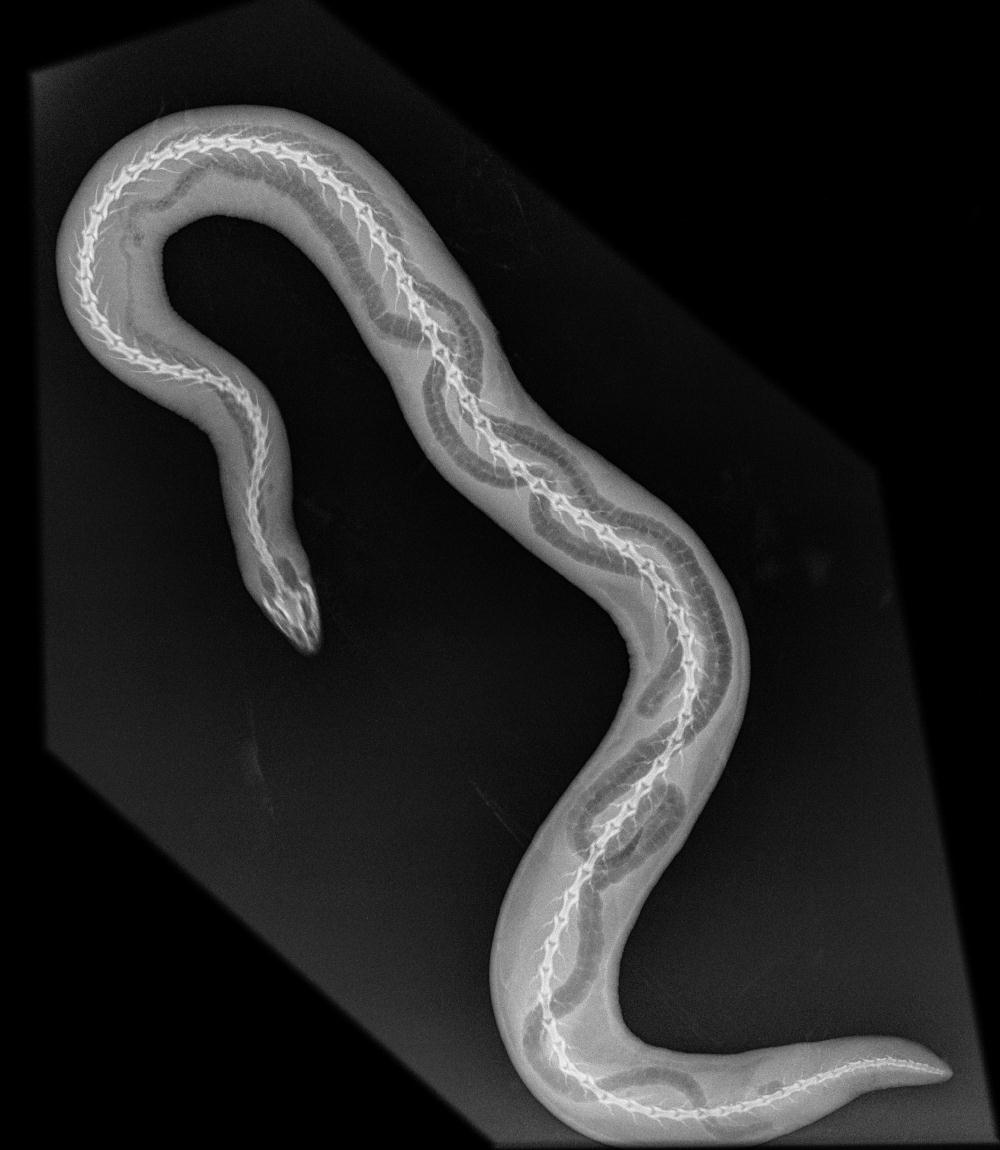
폐가 없는 한 종, ''Atretochoana eiselti''를 제외하고,[7] 모든 무족영원목은 폐를 가지고 있지만, 피부나 입을 사용하여 산소를 흡수하기도 한다. 종종 왼쪽 폐가 오른쪽 폐보다 훨씬 작으며, 이는 뱀에서도 발견되는 체형 적응의 일종이다.[8]
몸통 근육은 땅을 헤쳐나가는 데 적합하며, 척추와 그 근육은 피부에 밀착된 몸벽 근육의 바깥층 내부에서 피스톤 역할을 한다.[9] 바깥층 근육을 수축시키면 체강을 압착하여 몸을 길게 하는 강력한 정수압이 발생한다.[10] 이 근육 시스템을 통해 동물은 뒷부분을 고정하고 머리를 앞으로 밀어낸 다음, 파동처럼 몸의 나머지 부분을 끌어올릴 수 있다. 물이나 매우 묽은 진흙 속에서는 무족영원목은 뱀장어처럼 헤엄친다.[11] Typhlonectidae과에 속하는 무족영원목은 수생이며, 이 종류 중에서 가장 크다. 이 과의 대표 종들은 몸의 뒷부분을 따라 살로 된 지느러미가 있어 물속에서 추진력을 높인다.[12]
무족영원목의 몸 구조는 다음과 같다.
- 사지와 지대를 가지지 않으며, 몸은 가늘고 긴 원통형이며, 많은 체절적인 환상 피부의 주름(환대)을 가지고 있어 지렁이와 비슷하다. 환대를 구분하는 홈이 척추골의 말단과 일치할 때를 1차 환대, 1차 환대 사이에 있는 것을 2차 환대라고 한다.
- 긴 꼬리는 가지지 않으며, 총배설강은 몸의 말단 근처에 있다.
- 피부는 점액선이 풍부하다.
- 많은 경우 피부에 골질 비늘을 가진다. 조상으로 여겨지는 세룡류를 거쳐 멀리 어류로부터 물려받은 것인지, 2차적으로 획득한 형질인지는 설이 갈린다.
- 눈은 피부에 덮여 있다. 명암을 느끼는 정도밖에 도움이 되지 않는다.
- 감각 기관으로 촉수를 가진다. 안으로 집어넣을 수 있으며, 야콥슨 기관(서골비기)에 냄새를 운반한다.
- 고막을 가지지 않는다. 공기 중을 전달하는 소리는 매우 낮은 주파수의 것밖에 (상한 200Hz) 들을 수 없지만, 땅의 진동은 민감하게 느낄 수 있다.
- 위턱에 2열, 아래턱에 1~2열의 이빨 모양의 이가 있다. 물어뜯은 상대를 강력하게 유지하는 데 도움이 된다.
- 턱을 닫을 때, 다른 턱구류와 마찬가지로 하악거근에 의해 아래턱을 끌어올리는 것 외에도, 후하골간근에 의해 하악골의 관절 후돌기를 후하방으로 당기는 것으로 아래턱 전부를 상방으로 움직이고, 두장근을 수축시킴으로써 턱과 두개골을 굴곡시킨다. 이것에 의해 매우 강한 힘으로 턱을 닫을 수 있다.
- 두개골은 거의 완전히 뼈로 덮여 있다. 땅을 파고 들어가는 데 사용된다. 이것이 원시 양서류로부터 물려받은 특징인지, 지하 생활에 적응하여 새롭게 얻은 형질인지는 잘 알려져 있지 않다.
- 척추골은 일반적으로 보이는 것처럼 먼저 연골로서 형성되어 그것이 치환되는 것이 아니라, 척색 주위에 직접 형성된다. 이것은 공추아강과 공통되는 특징이다.
- 많은 경우, 좌폐는 흔적적이며, 우폐가 자루 모양으로 발달해 있다. 미즈아시나시이모리과는 좌우 폐가 모두 발달해 있다. 예외적으로 폐를 가지지 않는 종이 2종 (미즈아시나시이모리과의 ''Atretochoana eiselti'' (남미), ''Caecilia iwokramae'') 발견되었다. 피부와 입도 가스 교환에 사용된다.
2. 2. 두개골과 감각 기관
무족영원목은 작거나 없는 눈을 가지고 있으며, 알려진 유일한 종류의 광수용체가 있어 시력은 명암 감지에 제한되어 있다.[13][14] 다른 현대 양서류(개구리와 도롱뇽)와 달리 두개골은 튼튼하고 견고하며 판 모양의 두개골 뼈 사이에 큰 구멍이 거의 없다. 주둥이는 뾰족하고 탄알 모양으로 흙이나 진흙을 뚫고 나가는 데 사용된다. 대부분의 종에서 입은 머리 아래로 들어가 있어 주둥이가 입 위로 돌출되어 있다.[15]두개골의 뼈는 선사 시대 양서류 종에 비해 수가 줄어들었다. 두개골의 많은 뼈가 융합되어 있는데, 상악골과 구개골은 모든 현존하는 무족영원목에서 상악구개골로 융합되었고, 비골과 전상악골은 일부 과에서 비전상악골로 융합된다. 일부 과는 중격상악골, 전두골, 그리고/또는 안와(눈 구멍)를 둘러싼 후두개골 유사 뼈와 같은 특정 두개골 뼈의 유무로 구별될 수 있다. 뇌는 os basale이라고 하는 완전히 통합된 복합 뼈로 둘러싸여 있으며, 이는 두개골의 뒤쪽과 아래쪽 부분을 대부분 차지한다. 위에서 본 두개골에서는 일부 종에서 중사골이 두개골 지붕의 중앙선에 끼어 있는 것을 볼 수 있다.[16][30][17]
모든 무족영원목은 머리 양쪽에 눈과 콧구멍 사이에 위치한 촉수로 알려진 한 쌍의 독특한 감각 구조를 가지고 있다. 이들은 코를 기반으로 하는 정상적인 후각 외에 두 번째 후각 능력을 위해 사용될 것으로 보인다.[11]
환무족영원목 (''Siphonops annulatus'')은 일부 뱀독과 도마뱀의 상동일 수 있는 치아선를 가지고 있다. 이 선의 기능은 알려져 있지 않다.[18]
중이는 등골뼈와 난원창으로만 구성되어 있으며, 이는 일부 파충류에서 볼 수 있는 재진입 유체 회로를 통해 진동을 내이로 전달한다. Scolecomorphidae과 내의 종의 성체는 등골과 난원창이 모두 없어 중이 장치의 모든 구성 요소가 없는 유일한 알려진 양서류이다.[19]
아래턱은 무족영원목에서 특화되어 있다. 멸종된 종을 포함한 Gymnophionans는 턱의 두 가지 구성 요소만 가지고 있다. 의치 (앞쪽에 있으며 이빨을 지님)와 의각 (뒤쪽에 있으며 턱 관절과 근육 부착부를 지님)인데, 이 두 가지 구성 요소는 더 큰 뼈 세트가 융합된 후 남는 것이다. 최대 20개의 이빨이 있는 추가 삽입 이빨 열은 턱의 주요 가장자리 이빨 열과 평행하게 놓여 있다.[30]
가장 원시적인 무족영원목을 제외한 모든 무족영원목은 다른 양서류에서 발견되는 단일 쌍과 비교하여 턱을 닫기 위한 두 세트의 근육을 가지고 있다. 한 세트의 근육인 내전근은 턱 관절 앞의 의각 상단 가장자리에 삽입된다. 내전근은 척추동물에서 흔히 볼 수 있으며, 위쪽과 앞쪽으로 당겨 턱을 닫는다. 더 독특한 세트의 근육인 외전근은 턱 관절 아래와 뒤의 의각 뒤쪽 가장자리에 삽입된다. 그들은 뒤쪽과 아래쪽으로 당겨 턱을 닫는다. 턱 근육은 무족영원목 중 가장 효율적인 굴착기에서 더 잘 발달되어 있으며, 두개골과 턱을 견고하게 유지하는 데 도움이 되는 것으로 보인다.[11][20]
2. 3. 피부
무족영원류의 피부는 매끄럽고 보통 어둡지만, 일부 종은 다채로운 색을 띠기도 한다. 피부 안에는 방해석 비늘이 있다. 이러한 비늘 때문에 무족영원류는 한때 화석 Stegocephalia와 관련이 있다고 여겨졌지만, 현재는 이차적으로 발달한 것으로 여겨지며 두 그룹은 관련이 없을 가능성이 높다.[15] Scolecomorphidae 및 Typhlonectidae과에서는 비늘이 없지만, ''Typhlonectes compressicauda'' 종에서는 몸 뒤쪽 부위에서 미세한 비늘이 발견되었다.[21]피부에는 또한 몸을 부분적으로 감싸는 많은 링 모양의 주름 또는 환대가 있어 분절된 모습을 보인다. 다른 몇몇 현존하는 양서류와 마찬가지로, 피부에는 포식자를 억제하기 위해 독소를 분비하는 샘이 포함되어 있다.[11] ''Siphonops paulensis''의 피부 분비물은 용혈성 특성을 갖는 것으로 나타났다.[22]
2. 4. 턱과 이빨
무족영원목의 두개골은 튼튼하고 견고하며, 판 모양의 두개골 뼈 사이에 큰 구멍이 거의 없다. 주둥이는 뾰족하고 탄알 모양으로 흙이나 진흙을 뚫는 데 사용된다. 대부분의 종에서 입은 머리 아래로 들어가 있어 주둥이가 입 위로 돌출되어 있다.[15]두개골의 뼈는 선사 시대 양서류 종에 비해 수가 줄어들었다. 상악골과 구개골은 모든 현존하는 무족영원목에서 상악구개골로 융합되었고, 비골과 전상악골은 일부 과에서 비전상악골로 융합되는 등 두개골의 많은 뼈가 융합되어 있다. 일부 과는 중격상악골, 전두골, 그리고/또는 안와(눈 구멍)를 둘러싼 후두개골 유사 뼈와 같은 특정 두개골 뼈의 유무로 구별될 수 있다.[16][30][17] 뇌는 os basale이라고 하는 완전히 통합된 복합 뼈로 둘러싸여 있으며, 이는 두개골의 뒤쪽과 아래쪽 부분을 대부분 차지한다. 위에서 본 두개골에서는 일부 종에서 중사골이 두개골 지붕의 중앙선에 끼어 있는 것을 볼 수 있다.[16][17]
모든 무족영원목은 머리 양쪽에 눈과 콧구멍 사이에 위치한 촉수라는 한 쌍의 독특한 감각 구조를 가지고 있다. 이들은 코를 기반으로 하는 정상적인 후각 외에 두 번째 후각 능력을 위해 사용될 것으로 보인다.[11]
환무족영원목 (''Siphonops annulatus'')은 일부 뱀독과 도마뱀의 상동일 수 있는 치아선을 가지고 있다. 이 선의 기능은 알려져 있지 않다.[18]
아래턱은 무족영원목에서 특화되어 있다. 멸종된 종을 포함한 Gymnophionans는 턱의 두 가지 구성 요소만 가지고 있는데, 앞쪽에 있으며 이빨을 지닌 의치와 뒤쪽에 있으며 턱 관절과 근육 부착부를 지닌 의각이 그것이다. 이 두 가지 구성 요소는 더 큰 뼈 세트가 융합된 후 남는 것이다. 최대 20개의 이빨이 있는 추가 삽입 이빨 열은 턱의 주요 가장자리 이빨 열과 평행하게 놓여 있다.[30]
가장 원시적인 무족영원목을 제외한 모든 무족영원목은 다른 양서류에서 발견되는 단일 쌍과 비교하여 턱을 닫기 위한 두 세트의 근육을 가지고 있다. 한 세트의 근육인 내전근은 턱 관절 앞의 의각 상단 가장자리에 삽입된다. 내전근은 척추동물에서 흔히 볼 수 있으며, 위쪽과 앞쪽으로 당겨 턱을 닫는다. 더 독특한 세트의 근육인 외전근은 턱 관절 아래와 뒤의 의각 뒤쪽 가장자리에 삽입된다. 그들은 뒤쪽과 아래쪽으로 당겨 턱을 닫는다. 턱 근육은 무족영원목 중 가장 효율적인 굴착기에서 더 잘 발달되어 있으며, 두개골과 턱을 견고하게 유지하는 데 도움이 되는 것으로 보인다.[11][20]
위턱에 2열, 아래턱에 1~2열의 이빨 모양의 이가 있다. 물어뜯은 상대를 강력하게 유지하는 데 도움이 된다. 턱을 닫을 때, 다른 턱구류와 마찬가지로 하악거근에 의해 아래턱을 끌어올리는 것 외에도, 후하골간근에 의해 하악골의 관절 후돌기를 후하방으로 당기는 것으로 아래턱 전부를 상방으로 움직이고, 두장근을 수축시킴으로써 턱과 두개골을 굴곡시킨다. 이것에 의해 매우 강한 힘으로 턱을 닫을 수 있다.
3. 생태
]
무족영원목은 굴을 파는 생활 방식에 매우 적합한 해부학적 구조를 가지고 있다. 원시적인 속인 ''Ichthyophis''의 몇몇 종에서는 퇴화된 사지의 흔적이 발견되었으며, ''Typhlonectes compressicauda''에서는 배아 발달 과정에서 사지 싹이 관찰되기도 했다. 그러나 이들은 다른 부분에서는 완전히 사지가 없는 몸을 가지고 있다.[4]
많은 무족영원목 종은 외적에게 습격당했을 때 피부에서 대량의 점액을 분비하여 미끄러져 빠져나가려 한다. 소수의 종은 독액을 분비하는데, 그 독의 종류는 자세히 연구되지 않았지만 다른 양서류의 독과는 상당히 다른 것으로 보인다. 또한 대형 종은 상당한 힘으로 물 수 있으며, 무족영원과의 ''Geotrypetes seraphini''는 입에서 상당한 힘으로 물을 뿜어낸다고 한다.
3. 1. 서식지
무족영원목은 굴을 파는 생활 방식에 매우 적합한 신체 구조를 가지고 있다. 작은 종들은 벌레와 비슷하고, 큰 종은 뱀과 유사하다. 꼬리는 짧거나 없고, 총배설강은 몸 끝 부분 근처에 있다.[5][6][15]Atretochoana eiselti를 제외한[7] 모든 무족영원목은 폐를 가지고 있지만, 피부나 입을 사용하여 산소를 흡수하기도 한다. 종종 왼쪽 폐가 오른쪽 폐보다 훨씬 작으며, 이는 뱀에서도 발견되는 체형 적응의 일종이다.[8]
몸통 근육은 땅을 헤쳐나가는 데 적합하며, 척추와 그 근육은 피부에 밀착된 몸벽 근육의 바깥층 내부에서 피스톤 역할을 한다.[9] 바깥층 근육을 수축시키면 체강을 압착하여 몸을 길게 하는 강력한 정수압이 발생한다.[10] 이 근육 시스템을 통해 동물은 뒷부분을 고정하고 머리를 앞으로 밀어낸 다음, 파동처럼 몸의 나머지 부분을 끌어올릴 수 있다. 물이나 매우 묽은 진흙 속에서는 뱀장어처럼 헤엄친다.[11] Typhlonectidae과에 속하는 무족영원목은 수생이며, 이 종류 중에서 가장 크다. 이 과의 대표 종들은 몸의 뒷부분을 따라 살로 된 지느러미가 있어 물속에서 추진력을 높인다.[12]
지중해종은 환대 사이의 주름을 수축시켜 땅 속을 천천히 이동한다. 물가의 젖은 흙 속, 밭의 부드러운 흙 속, 숲 바닥의 낙엽 속 등에 서식한다. 야간에는 먹이가 되는 절지동물이나 소형 척추동물을 찾아 지상으로 나올 수도 있다고 한다. 수생종은 지느러미 모양이 된 몸통 후부를 사용하여 헤엄친다. 물 밑의 돌이나 쓰러진 나무 밑에 산다.
4. 분포
무족영원목은 동남아시아, 인도, 방글라데시, 네팔[26], 스리랑카, 동부 및 서아프리카, 인도양의 세이셸 제도, 중앙아메리카, 북부 및 동부 남아메리카의 습한 열대 지역에 서식한다. 아프리카에서는 기니비사우(''Geotrypetes'')에서 남부 말라위(''Scolecomorphus'')까지 발견되며, 짐바브웨 동부에서도 보고된 바 있지만 확인되지 않았다. 중앙 아프리카의 광대한 열대 우림 지역에서는 기록되지 않았다. 남아메리카에서는 아열대 동부 브라질을 거쳐 온대 북부 아르헨티나까지 분포한다. 파라나강의 홍수에 의해 북쪽에서 쓸려 내려와 부에노스아이레스까지 발견되기도 한다. 아메리카 대륙에서는 북쪽으로 멕시코 남부까지 분포한다. 가장 북쪽에 분포하는 종은 북부 인도의 ''Ichthyophis sikkimensis''이다. ''Ichthyophis''는 중국 남부와 베트남 북부에서도 발견된다. 동남아시아에서는 자바, 보르네오, 필리핀 남부까지 발견되지만, 윌리스선을 넘어 오스트레일리아나 인근 섬에서는 발견되지 않는다. 마다가스카르에는 무족영원목이 없는 것으로 알려져 있지만, 세이셸과 인도에 서식한다는 사실 때문에 그곳에 아직 발견되지 않은 멸종되었거나 현존하는 무족영원목이 있을 것이라는 추측이 제기되었다.[27]
2021년, 콜롬비아와 베네수엘라가 원산지인 무족영원목인 ''Typhlonectes natans''의 살아있는 표본이 플로리다 남부의 배수관에서 채집되었다. 이는 미국에서 야생에서 보고된 유일한 무족영원목으로, 아마도 야생동물 거래를 통해 유입된 도입종으로 간주된다. 해당 지역에 번식 집단이 형성되었는지는 알려지지 않았다.[28][29]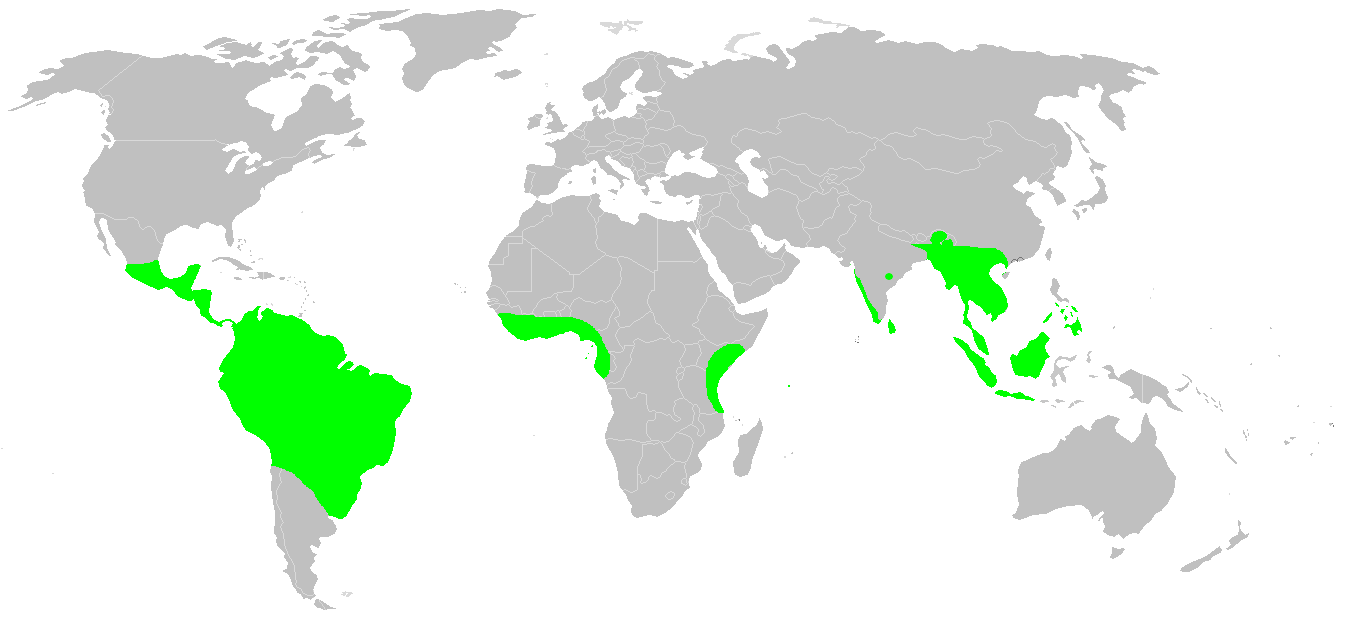

열대 지역에 널리 분포한다. 바다를 건널 수 없음에도 불구하고, 동남아시아, 인도, 중남미, 마다가스카르를 제외한 열대 아프리카에 분포하고 있으며, 아직 이들이 하나로 판게아 대륙을 형성하고 있던 트라이아스기 이전에 기원을 가지고 있음을 시사한다.
5. 진화
무족영원목의 진화 역사는 화석 기록이 매우 드물어 거의 알려져 있지 않다. 최초의 화석은 1972년에 발견된 고신세 척추뼈이다.[41] 이후 고신세와 후기 백악기 (세노마기) 퇴적층에서 현대 종의 특징을 가진 다른 척추뼈들이 발견되었다.[42]
계통 발생학적 증거에 따르면 무족영원목과 개구리류(개구리와 도롱뇽 포함)의 조상은 석탄기에 서로 갈라졌다. 이는 무족영원목의 추정 기원과 줄기 무족영원목의 가장 초기의 확실한 화석 사이의 7천만 년 이상의 간극을 남긴다.[43][51]
2023년 이전에는 줄기 무족영원목(개구리나 도롱뇽보다는 무족영원목에 더 가깝지만 현존하는 무족영원목 계통의 구성원은 아님)으로 여겨지는 가장 초기의 화석은 쥐라기 시대의 것이었다. 이 원시 속인 ''에오카에실리아''(Eocaecilia)는 작은 사지와 잘 발달된 눈을 가지고 있었다.[44] 
콜로라도의 후기 트라이아스기 친레 지층에서 발견된 스테레오스폰딜리 템노스폰딜인 ''친레스테고피스''(Chinlestegophis)는 2017년 논문에서 줄기 무족영원목으로 제안되었다.[43] 그러나 2020년 연구는 이러한 관계를 뒷받침하는 특성의 선택에 의문을 제기했으며,[48] 2019년에는 다른 위치가 지지되었다.[49] 2024년에는 ''친레스테고피스''가 리스암피비아 외부의 스테레오스폰딜리에 속하며 ''릴레이밀러루스''(Rileymillerus)의 자매 분류군으로 회수되었다.[50]
2023년 논문은 애리조나의 친레 지층에서 발견된 또 다른 양서류인 ''펑쿠스버미스''(Funcusvermis)에 대해 설명했다. ''펑쿠스버미스''는 수많은 두개골 및 턱 조각의 특징을 기반으로 줄기군 무족영원목으로 강력하게 지지되었다. 이 논문은 무족영원목의 기원에 대한 다양한 가설을 논의했지만, 가장 강력하게 지지된 가설은 리스암피비아를 ''게로바트라쿠스''와 밀접하게 관련이 있는 디소로포이드 템노스폰딜리의 단일 계통군으로 결합하는 것이었다.[51]
6. 분류
무족영원목(caecilian)이라는 이름은 "눈이 먼"을 의미하는 라틴어 단어 `caecus`에서 유래되었으며, 작거나 때로는 존재하지 않는 눈을 가리킨다. 이 이름은 칼 린네가 처음 기술한 종의 분류학적 이름에서 유래되었으며, 그는 이 종을 ''Caecilia tentaculata''라고 명명했다.[15]
역사적으로 무족영원목의 두 가지 주요 학명인 Apoda와 Gymnophiona의 사용에 대한 이견이 있었다. 일부 고생물학자들은 "관 그룹", 즉 모든 현대 무족영원목과 이러한 현대 계통의 멸종된 구성원을 포함하는 그룹을 지칭하기 위해 Apoda라는 이름을 사용하고, Gymnophiona라는 이름은 전체 그룹, 즉 개구리나 도롱뇽보다 현대 그룹과 더 밀접한 관련이 있는 모든 무족영원목과 무족영원목 유사 양서류를 지칭하기 위해 사용하는 것을 선호한다. 그러나 Apoda는 물고기, 해삼 그룹에 사용되었으며 나방의 속명이기도 하며, 무족영원목 분류학에서 계속 사용하면 혼란스러울 수 있다.
Wilkinson 외(2011)의 무족영원목 분류는 현존하는 무족영원목을 거의 200종을 포함하는 9개 과로 나누었다.[30] 2012년에는 열 번째 무족영원목 과인 Chikilidae가 새로 기술되었다.[31][32] 현존하는 무족영원목은 10개의 과로 나뉘며, 자세한 내용은 하위 과 섹션에서 다룬다.
무족영원목의 계통 분류는 다음과 같다.
- ''Funcusvermis'': 2023년에 기재된 멸종군. 트라이아스기 북미에 서식했던, 스템 그룹의 무족영원류. 1속 1종.[68]
- 에오카에키리아과(Eocaeiliaidae): 멸종군. 쥐라기 북미에 서식. 짧은 사지를 가지고 있었지만, 이미 공간이 없는 머리뼈와 촉수를 가지고 있었다. 1속.
- 오아시나시이무리과(Rhinatrematidae): 2속 9종. 남미 북부에 분포하는 소형 종으로, 최대 30cm 정도이며 가장 원시적인 형질을 가진다.
- 누메아시나시이무리과(Ichthyophiidae): 2속 39종. 인도, 중국 남부, 태국, 말레이 반도에 분포하는 중형 종으로, 최대 50cm 정도이며 원시적인 특징과 분화된 특징을 함께 가지고 있다.
- 케랄라아시나시이무리과(Uraeotyphlidae): 1속 5종. 인도 남부에 분포하는 소형 종으로, 최대 30cm 정도이며 누메아시나시이무리과와 유사하지만, 더욱 진보된 무족영원과와의 공통점도 가지고 있다.
- 아프리카아시나시이무리과(Scolecomorphidae): 2속 6종. 사하라 사막 이남의 열대 아프리카에 분포하는 중형 종으로, 최대 45cm 정도이다.
- 무족영원과(Caeciliidae): 26속 99종. 중남미, 아프리카, 인도, 세이셸 제도에 분포. 10cm 정도의 소형 종부터 150cm에 달하는 대형 종까지 포함하며 파생적인 특징을 가진다.
- 미즈아시나시이무리과(Typhlonectidae): 5속 13종. 남미에 분포하는 수생 대형 종으로, 몸통의 뒷부분은 지느러미 모양이 되어 있으며 가장 파생적인 특징을 가진다.
San Mauro 외(2014)는 분자 미토게놈 증거를 기반으로 ''Amazops''와 같이 최근에 기술된 속을 포함하도록 계통 분류를 수정했다.[38][39][40]
6. 1. 하위 과
무족영원목에는 10개의 과가 있으며, 아래 표와 같이 정리할 수 있다.[30][31][32]| 과 | 그림 | 분류학적 저자 | 속 | 종 | 일반명 | 지리적 분포 |
|---|---|---|---|---|---|---|
| 무족영원과(Caeciliidae) |  Caecilia subnigricans | Rafinesque, 1814 | 2 | 47 | 일반 무족영원목 | 중앙아메리카와 남아메리카(볼리비아 북부에서 코스타리카까지). |
| 키킬라과(Chikilidae) | style="text-align:center;"| | Kamei 외, 2012 | 1 | 4 | 인도 북동부 무족영원목 | 인도 북동부와 방글라데시, 미얀마에서도 발생 가능성. |
| 데르모피스과(Dermophiidae) |  Geotrypetes seraphini | Taylor, 1969 | 4 | 15 | 신열대 무족영원목 | 적도 아프리카(서아프리카, 탄자니아, 케냐), 중앙 및 남아메리카(콜롬비아 북부에서 멕시코까지). |
| Grandisoniidae (구 인도티플루스과(Indotyphlidae)) |  Grandisonia sechellensis | Lescure, Renous & Gasc, 1986 | 7 | 24 | 인도-아프리카 무족영원목 | 적도 아프리카(카메룬, 에티오피아), 세이셸, 서부 인도(서고츠). |
| 헤르펠레과(Herpelidae) |  Boulengerula taitana | 로랑, 1984 | 2 | 10 | 아프리카 무족영원목 | 적도 아프리카(나이지리아 남부에서 콩고 민주 공화국까지, 케냐 남부에서 말라위까지, 앙골라 및 잠비아에서도 발생 가능성). |
| 이크티오피스과(Ichthyophiidae) |  Ichthyophis kodaguensis | 테일러, 1969 | 2 | 57 | 아시아 꼬리 무족영원목 | 남아시아 및 동남아시아(서부 인도 북부에서 네팔까지, 동쪽으로 필리핀, 남부 중국 및 인도네시아까지). |
| 리나트레마과(Rhinatrematidae) |  Epicrionops sp. | 너스바움, 1977 | 3 | 14 | 아메리카 꼬리 무족영원목 | 남아메리카 북부(브라질 최북단 서부에서 베네수엘라, 콜롬비아, 에콰도르, 페루까지). |
| 스콜레코모르푸스과(Scolecomorphidae) |  Scolecomorphus kirkii | 테일러, 1969 | 2 | 6 | 땅속 눈 무족영원목 | 적도 아프리카(카메룬, 탄자니아, 말라위, 모잠비크). |
| 시포놉스과(Siphonopidae) |  Microcaecilia dermatophaga | 보나파르테, 1850 | 5 | 28 | 남아메리카 무족영원목 | 남아메리카(콜롬비아 남부에서 북부 아르헨티나, 파라과이, 최남단 브라질까지). |
| 티플로넥테스과(Typhlonectidae) |  Typhlonectes natans | 테일러, 1968 | 5 | 14 | 수생 무족영원목 | 남아메리카(콜롬비아와 베네수엘라 남부에서 북부 아르헨티나와 우루과이까지). |
이 과들은 형태학적, 분자적 증거를 바탕으로 분류되었으며,[33][34][35][36] 분류군 불확정성 문제를 해결하였다.[30][37]
6. 2. 계통 분류
2012년 현재 무족영원목의 계통 분류는 다음과 같다.[74]| 과 | 그림 | 분류학적 저자 | 속 | 종 | 일반명 | 지리적 분포 |
|---|---|---|---|---|---|---|
| 무족영원과(Caeciliidae) | Caecilia subnigricans | Rafinesque, 1814 | 2 | 47 | 일반 무족영원목 | 중앙아메리카와 남아메리카(볼리비아 북부에서 코스타리카까지). |
| 키킬라과(Chikilidae) | Kamei 외, 2012 | 1 | 4 | 인도 북동부 무족영원목 | 인도 북동부와 방글라데시, 미얀마에서도 발생 가능성. | |
| 데르모피스과(Dermophiidae) | Geotrypetes seraphini | Taylor, 1969 | 4 | 15 | 신열대 무족영원목 | 적도 아프리카(서아프리카, 탄자니아, 케냐), 중앙 및 남아메리카(콜롬비아 북부에서 멕시코까지). |
| 그란디소니아과(Grandisoniidae) (구 인도티플루스과(Indotyphlidae)) | Grandisonia sechellensis | Lescure, Renous & Gasc, 1986 | 7 | 24 | 인도-아프리카 무족영원목 | 적도 아프리카(카메룬, 에티오피아), 세이셸, 서부 인도(서고츠). |
| 헤르펠레과(Herpelidae) | Boulengerula taitana | 로랑, 1984 | 2 | 10 | 아프리카 무족영원목 | 적도 아프리카(나이지리아 남부에서 콩고 민주 공화국까지, 케냐 남부에서 말라위까지, 앙골라 및 잠비아에서도 발생 가능성). |
| 이크티오피스과(Ichthyophiidae) | Ichthyophis kodaguensis | 테일러, 1969 | 2 | 57 | 아시아 꼬리 무족영원목 | 남아시아 및 동남아시아(서부 인도 북부에서 네팔까지, 동쪽으로 필리핀, 남부 중국 및 인도네시아까지). |
| 리나트레마과(Rhinatrematidae) | Epicrionops sp. | 너스바움, 1977 | 3 | 14 | 아메리카 꼬리 무족영원목 | 남아메리카 북부(브라질 최북단 서부에서 베네수엘라, 콜롬비아, 에콰도르, 페루까지). |
| 스콜레코모르푸스과(Scolecomorphidae) | Scolecomorphus kirkii | 테일러, 1969 | 2 | 6 | 땅속 눈 무족영원목 | 적도 아프리카(카메룬, 탄자니아, 말라위, 모잠비크). |
| 시포놉스과(Siphonopidae) | Microcaecilia dermatophaga | 보나파르테, 1850 | 5 | 28 | 남아메리카 무족영원목 | 남아메리카(콜롬비아 남부에서 북부 아르헨티나, 파라과이, 최남단 브라질까지). |
| 티플로넥테스과(Typhlonectidae) | Typhlonectes natans | 테일러, 1968 | 5 | 14 | 수생 무족영원목 | 남아메리카(콜롬비아와 베네수엘라 남부에서 북부 아르헨티나와 우루과이까지). |
무족영원목의 가장 최근 계통 발생은 San Mauro 외(2014)가 검토한 분자 미토게놈 증거를 기반으로 하며, ''Amazops''와 같이 최근에 기술된 속을 포함하도록 수정되었다.[38][39][40]
; ''Funcusvermis''
: 2023년에 기재된 멸종군. 트라이아스기 북미에 서식했던, 스템 그룹의 무족영원류. 1속 1종.[68]
; 에오카에키리아과(Eocaeiliaidae)
: 멸종군. 쥐라기 북미에 서식. 짧은 사지를 가지고 있었지만, 이미 공간이 없는 머리뼈와 촉수를 가지고 있었다. 1속.
; 오아시나시이무리과(Rhinatrematidae)
: 2속 9종. 남미 북부에 분포하는 소형 종. 최대 30cm 정도. 가장 원시적인 형질을 가진다.
; 누메아시나시이무리과(Ichthyophiidae)
: 2속 39종. 인도, 중국 남부, 태국, 말레이 반도에 분포하는 중형 종. 최대 50cm 정도. 원시적인 특징과 분화된 특징을 함께 가지고 있다.
; 케랄라아시나시이무리과(Uraeotyphlidae)
: 1속 5종. 인도 남부에 분포하는 소형 종. 최대 30cm 정도. 누메아시나시이무리과와 유사하지만, 더욱 진보된 무족영원과와의 공통점도 가지고 있다.
; 아프리카아시나시이무리과(Scolecomorphidae)
: 2속 6종. 사하라 사막 이남의 열대 아프리카에 분포하는 중형 종. 최대 45cm 정도.
; 무족영원과(Caeciliidae)
: 26속 99종. 중남미, 아프리카, 인도, 세이셸 제도에 분포. 10cm 정도의 소형 종부터 150cm에 달하는 대형 종까지 포함한다. 파생적인 특징을 가진다.
; 미즈아시나시이무리과(Typhlonectidae)
: 5속 13종. 남미에 분포하는 수생 대형 종. 몸통의 뒷부분은 지느러미 모양이 되어 있다. 가장 파생적인 특징을 가진다.
7. 인간과의 관계
무족영원목은 은둔형 동물이기 때문에 인간의 몇몇 신화에만 등장하며, 전통적인 관습에서는 일반적으로 혐오스러운 존재로 여겨진다.
인도 특정 지역의 민속에서 무족영원목은 치명적인 독이 있다는 믿음에 근거하여 두려움과 혐오의 대상이 된다. 동히말라야의 무족영원목은 구어체로 "요통 뱀"이라고 불린다.[64] 서고츠에서는 ''Ichthyophis tricolor''가 킹코브라보다 더 독성이 강하다고 여겨진다.[65][66] 코브라와 다른 위험한 동물들에 대한 깊은 문화적 존경심에도 불구하고, 무족영원목은 발견 즉시 소금과 등유로 죽임을 당한다.[65] 이러한 신화는 인도 무족영원목의 보존 계획을 복잡하게 만들었다.[65][64][66]
카메룬 오쿠산에 서식하는 희귀종인 ''Crotaphatrema lamottei''는 오쿠족에 의해 케파-느티에(땅을 파는 생물)로 분류된다. 케파-느티에는 토착 두더지와 장님 뱀을 포함하는 용어로, 독성이 있어 마주치거나, 접촉하거나, 죽이면 고통스러운 궤양을 일으킨다고 여겨진다. 오쿠 전통에 따르면, 이 고통을 정화하는 의식에는 갈아놓은 허브, 팜유, 달팽이 껍질, 닭의 피로 구성된 물약을 만들어 왼쪽 엄지에 바르고 핥아 먹는 것이 포함된다.[67]
남미 무족영원목은 지역 문화와 다양한 관계를 가지고 있다.[66] 브라질 민속의 전설적인 웜형 짐승인 미뇨캉은 무족영원목에서 영감을 받았을 수 있다. 콜롬비아 민속에서는 수생 무족영원목인 ''Typhlonectes natans''가 가라앉은 병 속에 봉인된 머리카락 뭉치에서 나타날 수 있다고 한다. 멕시코 남부와 중앙 아메리카에서 ''Dermophis mexicanus''는 구어체로 "타팔쿠아"라고 불리며, 이는 그들이 집 위에 배변하는 부주의한 사람의 엉덩이에 파고든다는 믿음을 참조하는 이름이다. 이는 그들이 쓰레기 더미에 둥지를 트는 경향에서 영감을 받았을 수 있다.[66]
참조
[1]
웹사이트
Gymnophiona Müller, 1832
http://research.amnh[...]
American Museum of Natural History
2019-09-11
[2]
논문
Caecilians.
https://www.cell.com[...]
2012
[3]
문서
Caecilians. Current Biology, 22(17), R668-R669.
2012
[4]
문서
An Anatomical Feature of Caecilian Amphibians: The Lengthening of the Body and Organs
https://scienceforec[...]
[5]
서적
Introduction to Herpetology
W.H. Freeman and Company
[6]
서적
Die Blindwühlen
Westarp Wissenschaften
[7]
웹사이트
"''Atretochoana eiselti''"
http://www.nhm.ac.uk[...]
Natural History Museum
2012-02-22
[8]
논문
Reptilian Anatomy
1995-06
[9]
문서
The Ecology and Behavior of Amphibians
https://books.google[...]
[10]
문서
Caecilians - Current Biology
https://www.cell.com[...]
[11]
서적
Encyclopedia of Reptiles and Amphibians
Academic Press
[12]
서적
Extraordinary Animals: An Encyclopedia of Curious and Unusual Animals
Greenwood Press
[13]
논문
The Evolution of Amphibian Photoreception
2019-08-27
[14]
논문
Identification and characterization of visual pigments in caecilians (Amphibia: Gymnophiona), an order of limbless vertebrates with rudimentary eyes
[15]
문서
Caecilia
[16]
논문
Skulls of Gymnophiona and their significance in the taxonomy of the group
https://www.biodiver[...]
1969
[17]
논문
High-density three-dimensional morphometric analyses support conserved static (intraspecific) modularity in caecilian (Amphibia: Gymnophiona) crania
https://academic.oup[...]
2019-03-27
[18]
논문
Morphological Evidence for an Oral Venom System in Caecilian Amphibians
2020-07
[19]
문서
Hearing and Sound Communication in Amphibians
https://books.google[...]
[20]
논문
Caecilian jaw-closing mechanics: integrating two muscle systems
2008-05-15
[21]
논문
An insight into the skin glands, dermal scales and secretions of the caecilian amphibian Ichthyophis beddomei
[22]
논문
Occurrence of hemolytic activity in the skin secretion of the caecilian ''Siphonops paulensis''
[23]
뉴스
Bizarre, Wormlike and Oozing Milk for Their Babies
https://www.nytimes.[...]
2024-03-11
[24]
논문
Amphibian hatchlings find mother's milk
2024-03-07
[25]
논문
Milk provisioning in oviparous caecilian amphibians
https://www.science.[...]
2024-03-08
[26]
뉴스
Kutnjema Mareka Jiv Sarpa Hoina
http://kantipur.ekan[...]
Katipur
2016-05-05
[27]
문서
James D. Gardner, Jean-Claude Rage, The fossil record of lissamphibians from Africa, Madagascar, and the Arabian Plate, Palaeobiodiversity and Palaeoenvironments 96(1):1-52 · March 2016
[28]
웹사이트
Worm-Like, Limbless Amphibian Known As Caecilians Spotted in Florida For The First Time
https://conservation[...]
2021-08-04
[29]
논문
First record of a caecilian (order Gymnophiona, Typhlonectes natans) in Florida and in the United States
https://journals.ku.[...]
2021-07-15
[30]
논문
A nine-family classification of caecilians (Amphibia: Gymnophiona)
http://www.mapress.c[...]
[31]
논문
Discovery of a new family of amphibians from Northeast India with ancient links to Africa
[32]
뉴스
New amphibian family found in India
https://www.cbc.ca/n[...]
CBC News
2012-02-21
[33]
논문
Phylogeny of caecilian amphibians (Gymnophiona) based on complete mitochondrial genomes and nuclear RAG1
[34]
논문
Experimental design in caecilian systematics: phylogenetic information of mitochondrial genomes and nuclear rag1
[35]
논문
A mitogenomic perspective on the phylogeny and biogeography of living caecilians (Amphibia: Gymnophiona)
[36]
논문
Experimental design in phylogenetics: testing predictions from expected information
[37]
논문
The Amphibian Tree of Life
[38]
논문
A large-scale phylogeny of Amphibia including over 2800 species, and a revised classification of extant frogs, salamanders, and caecilians
[39]
논문
Life-history evolution and mitogenomic phylogeny of caecilian amphibians
[40]
웹사이트
AmphibiaWeb - Amazops amazops
https://amphibiaweb.[...]
2021-01-24
[41]
논문
The First Fossil Record of Caecilian Amphibians
1972-09-22
[42]
논문
A stem-group caecilian (Lissamphibia: Gymnophiona) from the Lower Cretaceous of North Africa
[43]
논문
Stem caecilian from the Triassic of Colorado sheds light on the origins of Lissamphibia
2017-07-03
[44]
논문
An Early Jurassic caecilian with limbs
1993-09-16
[45]
논문
A stem batrachian from the Early Permian of Texas and the origin of frogs and salamanders
[46]
논문
Cranial morphology of recumbirostrans (Lepospondyli) from the Permian of Kansas and Nebraska, and early morphological evolution inferred by micro-computed tomography
[47]
논문
A multilocus timescale for the origin of extant amphibians
[48]
논문
A review of the fossil record of caecilians (Lissamphibia; Gymnophionomorpha) with comments on its use to calibrate molecular timetrees
2020
[49]
논문
Phylogeny of Paleozoic limbed vertebrates reassessed through revision and expansion of the largest published relevant data matrix
2019
[50]
논문
The new problem of ''Chinlestegophis'' and the origin of caecilians (Amphibia, Gymnophionomorpha) is highly sensitive to old problems of sampling and character construction
https://fr.pensoft.n[...]
2024-01-04
[51]
논문
Triassic stem caecilian supports dissorophoid origin of living amphibians
2023-01-25
[52]
서적
Reproductive Biology and Phylogeny of Gymnophiona: Caecilians
https://books.google[...]
CRC Press
[53]
간행물
Caecilians - Cell Press
https://www.cell.com[...]
[54]
논문
Parental investment by skin feeding in a caecilian amphibian
https://deepblue.lib[...]
[55]
뉴스
'Yummy mummy' worms feed their skin to offspring
https://www.newscien[...]
2006-04-12
[56]
논문
Care and parentage in a skin-feeding caecilian amphibian
https://onlinelibrar[...]
2008
[57]
논문
One hundred million years of skin feeding? Extended parental care in a Neotropical caecilian (Amphibia: Gymnophiona)
2008-06-11
[58]
논문
A New Species of Skin-Feeding Caecilian and the First Report of Reproductive Mode in Microcaecilia (Amphibia: Gymnophiona: Siphonopidae)
2013-03-06
[59]
뉴스
These Amphibians Have a Taste for Their Mom's Skin
https://www.nytimes.[...]
2023-10-22
[60]
논문
Parental Care Contributes to Vertical Transmission of Microbes in a Skin-Feeding and Direct-Developing Caecilian
BioMed Central
2023-05-15
[61]
논문
An unusual diet of Ichthyophis caecilians
https://www.jstor.or[...]
2024-03-15
[62]
논문
An unusual diet of Ichthyophis caecilians
https://www.jstor.or[...]
2024-03-15
[63]
서적
Feeding: Form, Function, and Evolution in Tetrapod Vertebrates
Elsevier
[64]
간행물
Conservation of Caecilians in the Eastern Himalayas Region
https://www.cepf.net[...]
2009
[65]
간행물
Deep-rooted myths and their impact on the population of gymnophionan amphibians among the inhabited areas of Kerala, India
https://www.academia[...]
2008-08
[66]
서적
Eye of Newt and Toe of Frog, Adder's Fork and Lizard's Leg
http://dx.doi.org/10[...]
University of Chicago Press
2015
[67]
논문
Traditional indigenous perspectives on soil-dwelling vertebrates in Oku, Cameroon, with special reference to the caecilian ''Crotaphatrema lamottei''
https://www.research[...]
2011
[68]
논문
Triassic stem caecilian supports dissorophoid origin of living amphibians
https://www.nature.c[...]
2023-02
[69]
논문
Stem caecilian from the Triassic of Colorado sheds light on the origins of Lissamphibia
https://www.pnas.org[...]
2017-07-03
[70]
논문
Phylogeny of Paleozoic limbed vertebrates reassessed through revision and expansion of the largest published relevant data matrix
2019
[71]
논문
On Defining and Finding Islands of Trees and Mitigating Large Island Bias
http://dx.doi.org/10[...]
2021-03-22
[72]
논문
Enigmatic amphibians in mid-Cretaceous amber were chameleon-like ballistic feeders
http://dx.doi.org/10[...]
2020-11-06
[73]
논문
A Triassic stem-salamander from Kyrgyzstan and the origin of salamanders
2020-05-11
[74]
저널
Phylogenetic view of Amphibia
http://www.amphibiaw[...]
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com