헬리코프리온
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
헬리코프리온은 고생대 페름기에서 중생대 트라이아스기까지 생존했던 멸종된 연골어류로, 날카로운 나선형의 치아를 특징으로 한다. 이빨은 턱에 나선형으로 배열되어 있으며, 턱을 절단하는 방식으로 먹이를 섭취했을 것으로 추정된다. 헬리코프리온은 척추골 잔해를 통해 방추형 몸체와 삼각형 가슴 지느러미를 가졌을 것으로 추정되며, 전 세계적으로 화석이 발견되었다. 헬리코프리온은 진연골두류에 속하며, 에우게네오돈트목에 속하는 것으로 분류된다.
더 읽어볼만한 페이지
| 헬리코프리온 | |
|---|---|
| 개요 | |
 | |
| 학명 | Karpinsky, 1899 |
| 어원 | 'Helico(나선)' + 'prion(톱)' |
| 보존 상태 | 화석 |
| 속 | 멸종 |
| 분류 | |
| 계 | 동물계 |
| 문 | 척삭동물문 |
| 강 | 연골어강 |
| 아강 | 전두어아강 |
| 목 | †에우게네오돈트목 |
| 과 | 헬리코프리온과 |
| 종 | |
| 타입 종 | Helicoprion bessonowi Karpinsky, 1899 |
| 기타 종 | H. davisii (Woodward, 1886), 원래 Edestus davisii H. ergassaminon Bendix-Almgreen, 1966 H. karpinskii Obruchev, 1953 H. mexicanus Mullerried, 1945 H. svalis Siedlecki, 1970 H.? clerci Karpinsky, 1916 |
| 분포 | |
| 시대 | 시수랄기 ~ 과달루페기 (아르틴스크절 ~ 로디안절), 백만 년 전 |
| 이명 | |
| H. davisii 이명 | H. ferrieri (Hay, 1907), 원래 Lissoprion ferrieri H. jingmenense Chen, Cheng, & Yin, 2007 H. sierrensis Wheeler, 1939 |
| H. bessonowi 이명 | H. nevadensis Wheeler, 1939 |
2. 특징
헬리코프리온은 고생대 페름기부터 중생대 트라이아스기 초까지 생존했던 멸종한 연골어류의 일종이다. 가장 두드러진 특징은 아래턱 중앙에 위치한 나선형의 독특한 치아 뭉치로, 이름 역시 그리스어로 '나선형 톱날'을 의미한다.[5] 다른 연골어류처럼 골격이 연골로 이루어져 있어 완전한 형태의 화석이 드물지만, 발견된 이빨 나선과 유지어류(Eugeneodontida)에 속하는 가까운 친척들의 화석을 통해 연구되었다.[3][4][37]
이 독특한 나선형 치아는 평생 동안 만들어진 모든 이빨이 안쪽으로 말려 들어가며 보존되는 구조이다.[19][39] CT 스캔 연구 등을 통해 이 치아 나선이 아래턱 전체를 차지하며, 위턱에는 기능적인 큰 이빨이 없었다는 사실이 밝혀졌다.[40] 이를 바탕으로 헬리코프리온은 먹이를 씹기보다는 나선형 이빨을 이용해 주로 부드러운 몸체의 먹이를 자르거나 베어 사냥했을 것으로 추정된다.[8][40] 주된 먹이는 오징어와 같은 두족류나 물고기였을 가능성이 높다.[8]
분류학적으로 헬리코프리온은 연골어류 내에서 진연골두류(Euchondrocephali)에 속하며, 더 구체적으로는 에우게네오돈트목(Eugeneodontida)의 헬리코프리온과(Helicoprionidae)로 분류된다.[3][5][37] 이는 현존하는 상어나 가오리보다는 은상어(키메라)와 계통적으로 더 가깝다는 것을 의미하지만, 현생 은상어와는 모습이나 생태가 매우 다른 독자적인 그룹이다.[9][2]
2. 1. 형태
다른 연골어류와 마찬가지로, ''헬리코프리온''과 다른 유지어류(Eugeneodontida)는 연골로 이루어진 골격을 가졌다. 이 때문에 라거슈테트와 같이 연골까지 온전히 보존되는 특별한 환경이 아니면 사체가 분해되어, 전체적인 모습을 정확히 복원하기 어렵다.[3][4][37] 그러나 펜실베이니아기부터 트라이아스기 시대의 근연종인 ''Caseodus'', ''Fadenia'', ''Romerodus'' 등 일부 유지어류에서 척추뼈를 포함한 비교적 완전한 화석이 발견되어, 이를 바탕으로 ''헬리코프리온''의 몸 형태를 추정할 수 있다.[3][4][37]근연종 화석을 통해 추정된 ''헬리코프리온''의 몸은 방추형(유선형)이며, 삼각형의 가슴 지느러미를 가졌다. 지느러미 가시가 없는 크고 삼각형의 등지느러미가 하나 있었고, 높고 갈라진 꼬리 지느러미는 외형상 동형꼬리(두 엽의 크기가 같음) 형태였을 것이다. 이러한 몸 형태는 참치, 새치, 청상아리처럼 빠르고 활동적으로 넓은 바다를 헤엄치는 포식성 어류와 유사하다. 또한 유지어류는 골반 지느러미와 뒷지느러미가 없었으며, ''Romerodus''처럼 몸 측면을 따라 꼬리지느러미까지 이어지는 넓은 용골(keel)이 있었을 수도 있다. ''Fadenia''는 잘 드러난 아가미 구멍 5개와 흔적적인 여섯 번째 아가미 구멍이 있었던 것으로 보인다. 현생 키메라에서 나타나는 특수한 아가미 바구니나 살덩이 형태의 아가미 덮개의 증거는 없다.[3][4]
''헬리코프리온''의 크기는 특징적인 이빨 소용돌이의 크기로 추정한다. 레베데프는 지름 35cm에서 40cm 크기의 이빨 소용돌이를 가진 개체가 현생 돌묵상어와 비슷한 몸길이 5m에서 8m에 달했을 수 있다고 제안했다.[37] 현재까지 알려진 가장 큰 ''헬리코프리온'' 이빨 소용돌이(표본 IMNH 49382, 종 미상)는 지름이 56cm이고 가장 큰 치아 높이가 14cm에 달하며, 이는 몸길이 7.6m가 넘는 개체의 것으로 추정된다.[5][19]

''헬리코프리온''의 가장 두드러진 특징은 그리스어로 '나선형 톱날'이라는 이름처럼, 전기 톱 날과 유사한 나선형의 치아 뭉치("치아 나선", "tooth whorl")이다. 대부분의 ''헬리코프리온'' 표본은 이 치아 나선만 발견된다. 이 구조는 로그 나선 형태의 치근 위에 수십 개의 에나멜질로 덮인 이빨들이 배열된 형태이다. 나선 중심부의 가장 어리고 작은 이빨은 갈고리 모양이지만, 나머지 이빨들은 대체로 삼각형 모양에 옆으로 납작하며 종종 가장자리에 톱니가 있다.[5] 이빨 크기는 나선 중심에서 바깥쪽으로 갈수록 커져서 가장 큰 것은 길이가 10cm를 넘기도 한다. 각 이빨 아랫부분은 바로 앞 이빨의 머리(치관) 아래를 덮는 돌출부를 형성한다. 에나멜질 아래 치근의 가장 아랫부분("샤프트")은 나선의 이전 회전을 감싸는 연골 위에 놓인다. 완전한 치아 나선의 가장 바깥쪽 부분은 치관의 중간과 윗부분이 없는 확장된 뿌리로 끝난다.[19] ''헬리코프리온''을 포함한 에우게네오두스류는 좌우 턱의 이빨이 정중선에서 합쳐져(Symphyseal teeth) 하나의 구조를 이루는 특징이 있으며, ''헬리코프리온''은 이 치아 나선을 아래턱 중앙에 가지고 있었다.[39] 새로운 이빨이 나선 앞쪽 끝에 계속 더해지고 오래된 이빨은 안쪽으로 말려 들어가므로, 개체가 평생 만든 모든 이빨이 이 나선 안에 보존된다.
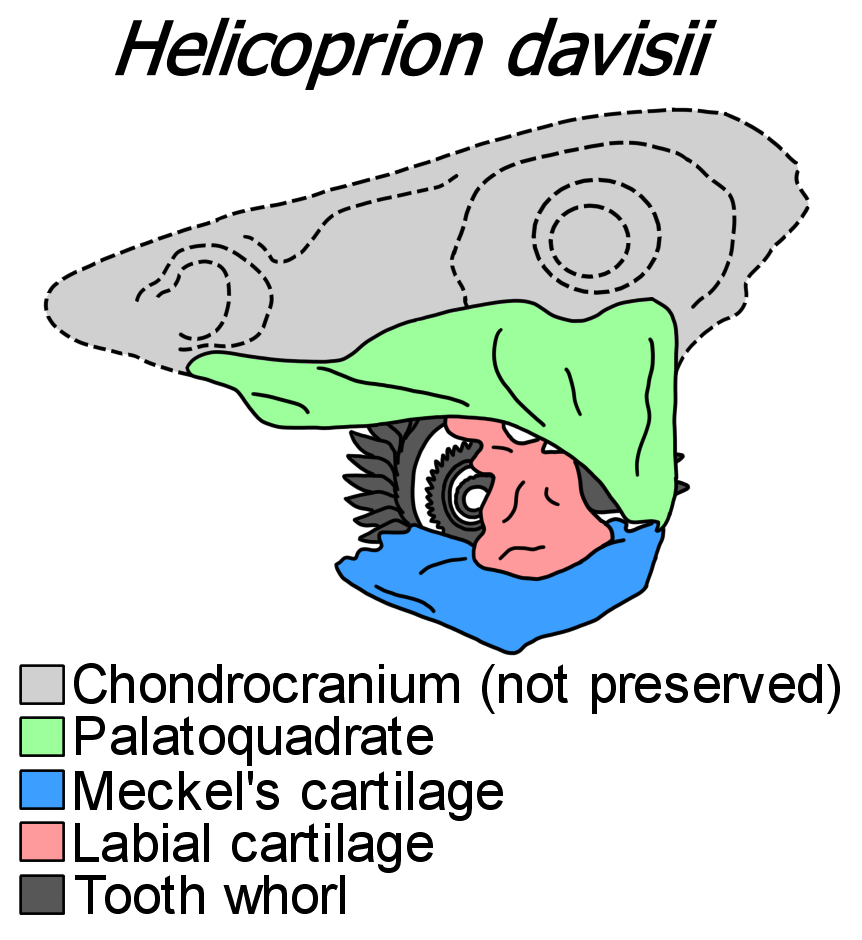
치아 나선 외 다른 부분이 보존된 표본은 매우 드물다. 가장 잘 보존된 표본은 아이다호에서 발견된 ''Helicoprion davisii''의 표본 IMNH 37899("아이다호 4")이다.[6] 2013년 CT 스캔을 이용한 재연구를 통해 연골 구조가 더 자세히 밝혀졌는데,[2][8] 이 표본에는 치아 나선과 함께 위턱을 이루는 구개사각골(palatoquadrate), 아래턱을 이루는 메켈 연골(Meckel's cartilage), 그리고 치아 나선을 지지하는 튼튼한 입술 연골(labial cartilage)이 거의 완전하게 보존되어 있었다. 이 모든 구조는 현생 연골어류처럼 각주형 석회화 연골로 이루어져 있다. 뇌와 감각 기관을 감쌌을 연골두개(neurocranium)는 보존되지 않았다. 턱은 옆으로 상당히 납작하게 눌려 있는데, 이는 화석화 과정에서의 변형일 수 있다.[2][8]
''헬리코프리온''은 '자동이형성'(autodiastylic) 턱 지지 구조를 가졌는데, 이는 구개사각골 안쪽 가장자리가 두 지점에서 연골두개에 단단히 붙어있지만 완전히 합쳐지지는 않았음을 의미한다. 이 두 부착 지점은 구개사각골 앞쪽의 돔 모양 돌기와 위쪽 뒷모서리의 넓적한 돌기이다.[2] 이런 형태의 턱은 초기 진연골두류(Euchondrocephali)에서 흔하며, 현생 동물 중에서는 배아 상태의 키메라에서만 볼 수 있다.[7] 또 다른 표본(USNM 22577+494391)에서는 구개사각골 안쪽 표면이 수많은 작은(약 2mm 너비) 이빨들로 덮여 있음이 확인되었다.[5] 이 위턱 이빨들은 낮고 둥글며, 아래턱의 치아 나선에 긁히면서 일종의 '포장'(pavement)을 형성했던 것으로 보인다.[8] 뒤쪽에서 구개사각골은 메켈 연골과 만나 턱 관절을 이룬다. 구개사각골과 설골(hyomandibula) 사이의 관절 증거는 없다.[2]
메켈 연골에는 턱 관절 바로 앞에 추가적인 돌기가 있는데, 이는 ''헬리코프리온'' 특유의 구조로, 턱이 닫힐 때 치아 나선이 연골두개를 뚫는 것을 막는 역할을 했을 것으로 추정된다. 또 다른 독특한 특징은 입술 연골이 메켈 연골 윗면과 연골결합(융합된 관절)을 형성한다는 점이다. 이 입술 연골은 치아 나선의 뿌리 근처에서 넓어져 측면 지지력을 제공하고, 턱이 닫힐 때 구개사각골에 맞물려 힘을 분산시키는 데 도움을 주며, 가장 안쪽의 어린 나선 뿌리를 보호하는 역할도 했다.[2][8][5]
위턱에 기능적인 절단용 이빨이 없었다는 점과 아래턱 치아 나선의 형태로 미루어 볼 때, ''헬리코프리온''은 먹이를 씹거나 으스러뜨리기보다는 아래턱의 치아 나선으로 먹이를 자르거나 베어서 삼켰을 것으로 생각된다.[40] 먹이로는 당시에 살았던 조개나 암모나이트 같은 단단한 껍질을 가진 동물보다는 오징어와 같은 부드러운 두족류나 물고기를 주로 먹었을 가능성이 높다.
''헬리코프리온''은 분류학상 현생 상어보다는 은상어에 더 가깝지만, 현재 은상어목만을 포함하는 두판류(Holocephali)는 고생대 동안 에우게네오두스목을 포함한 여러 목을 포함하며 매우 다양했다. 당시 두판류에는 현생 은상어와는 전혀 다른 형태의 종들(예: 스테타칸투스, ''Belantsea'', ''Iniopteryx'')이 많이 존재했다. 따라서 일부 오래된 복원도에서처럼 ''헬리코프리온''을 현생 은상어와 비슷한 몸체로 그리는 것은 근연종의 화석 증거와 맞지 않는 오류이다.[47][49][50]
2. 2. 치아 나선
대부분의 ''헬리코프리온'' 표본은 공통적인 로그 나선 형태의 치근 내에 매립된 수십 개의 에나멜질로 덮인 치아로 구성된 "치아 나선"에서만 알려져 있다. 나선 중심의 가장 젊고 첫 번째 치아는 갈고리 모양이지만, "어린 치아 아치"로 불리며, 다른 모든 치아는 일반적으로 삼각형 모양이며, 측면으로 압축되어 있고 종종 톱니 모양을 하고 있다.[5] 치아 크기는 나선의 중심(원심)에서 멀어질수록 증가하며, 가장 큰 치아는 길이가 10cm를 초과할 수 있다. 치아의 하단은 이전 치아의 치관 아래로 덮개 모양의 돌출부를 형성한다. 에나멜질 치아 돌출부 아래의 치근 최하단은 "샤프트"라고 하며, 나선의 이전 회전을 감싸는 연골 위에 놓여 있다. 완전한 치아 나선에서 나선의 가장 바깥쪽 부분은 치관의 중간 부분과 윗부분이 없는 확장된 뿌리로 끝난다.[19] 치아 직경이 40cm인 개체는 몸길이가 5m에서 8m까지 성장했을 것으로 추정되며[47], 발견된 치아 나선의 최대 직경은 약 56cm 정도였다.[38]
전기 톱을 연상시키는 나선형으로 감긴 치아는 헬리코프리온의 가장 큰 특징이다. 헬리코프리온이 속한 에우게네오두스류는 좌우 턱의 치아가 정중선상에서 유합(Symphyseal teeth)되어 있는 특징이 있으며, 헬리코프리온 역시 이 치아를 아래턱 중앙에 가지고 있었다.[39] 새롭고 큰 치아는 나선의 선단에 더해지고, 오래되고 작은 치아는 나선 안쪽으로 말려 들어가기 때문에, 개체가 태어나서 죽을 때까지 만들어낸 모든 치아가 이 나선 안에 수용되어 있다.

1세기 이상 헬리코프리온의 나선형 이빨이 아래턱의 어디에 위치하는지는 불명확했다. 1899년 발견 당시에는 위턱 바깥쪽에 위치한다고 생각되었고, 이후 아래턱에 배치하는 복원도 제안되었다. 심지어 등지느러미나 꼬리지느러미에 있는 방어 구조로 해석하는 설도 등장했다.[40] 1960년대에 같은 에우게네오두스류인 오르니토프리온의 두개골 화석이 발견되면서 나선형 이빨이 아래턱에 위치한다는 것이 밝혀졌다.[44] 1994년, 미술가 레이 트롤 등은 이 이빨을 아래턱의 끝에 배치하고 현대의 고블린 상어를 참고하여 가늘고 긴 주둥이를 가진 상어의 모습을 그렸다.[45] 2008년에는 메리 패리시가 스미소니언 박물관의 로버트 퍼디, 빅터 스프링어, 맷 캐러노 등의 감수 하에 이빨을 목구멍 안쪽에 배치한 복원도를 내놓았지만[46], 다른 연구에서는 이 결론을 받아들이지 않았다.[47][48]
2013년 CT 스캔을 이용한 연구에서 치아와 두개골의 위치 관계가 명확히 밝혀졌다. 이 연구에 따르면, 치아 나선은 턱 뒤쪽에 위치하며 턱뼈궁 전체를 차지하고 있었다. 또한 완두아강 특유의 특징인, 턱궁과 신경두개가 두 곳에서 관절하는 구조(Autodiastylic)도 확인되었다. 중요한 점은 위턱에는 치아가 존재하지 않았다는 사실이다. 이를 통해 헬리코프리온은 먹이를 씹어 으깨는 방식이 아니라, 아래턱의 치아 나선을 이용해 먹이를 절단하여 삼켰을 것으로 추정된다.[40]
다른 화석 어류 중에도 비슷한 형태의 치아를 가진 경우가 있는데, 예를 들어 오니코두스는 턱의 선단에 이와 유사한 몇 개의 치아를 가지고 있다.
2. 3. 두개골
다른 연골어류처럼 ''헬리코프리온''의 골격은 연골로 이루어져 있어, 특별한 보존 환경이 아니면 화석으로 남기 어려웠다. 이 때문에 두개골을 포함한 전체 모습을 정확히 파악하기 어려웠다.[3][4]
치아 구조 외에 두개골 일부가 함께 보존된 ''헬리코프리온'' 표본은 매우 드물다. 가장 잘 보존된 표본은 아이다호에서 발견된 IMNH 37899(일명 "아이다호 4")로, ''Helicoprion davisii'' 종에 속한다. 이 표본은 1950년에 발견되어 1966년에 처음 기술되었고,[6] 2013년 CT 스캔을 이용한 연구를 통해 연골 구조가 더 자세히 밝혀졌다. CT 스캔 결과, 거의 완전한 턱 구조가 3차원 형태로 보존되어 있음이 확인되었다. 이 표본에는 나선형 치아뿐만 아니라 위턱을 이루는 구개사각골, 아래턱을 이루는 메켈 연골, 그리고 치아를 지지하는 입술 연골이 포함되어 있다. 이 구조들은 모두 현대 연골어류처럼 각주형 석회화 연골로 이루어져 있다. 하지만 뇌와 감각 기관을 감쌌을 연골두개는 보존되지 않았다.[2][8]
''헬리코프리온''의 턱은 자동이형성(autodiastylic) 구조를 가지는데, 이는 위턱(구개사각골)이 연골두개와 두 지점에서 단단히 연결되지만 완전히 융합되지는 않은 형태를 의미한다. 이 연결 지점은 구개사각골 앞쪽의 돔형 돌기와 위쪽 뒷부분의 플랜지형 돌기이다.[2] 이러한 턱 구조는 초기 진연골두류에서 흔하며, 현생 동물 중에서는 배아 상태의 키메라에서만 관찰된다.[7] 또 다른 표본(USNM 22577+494391)에서는 위턱(구개사각골) 안쪽 표면이 수많은 작은 이빨(약 2mm 너비)로 덮여 있음이 밝혀졌다.[5] 이 위턱 이빨들은 낮고 둥글며, 아래턱의 나선형 치아가 움직일 때 마찰하며 먹이를 처리하는 데 도움을 주었을 것으로 보인다.[8] 턱 관절은 위턱(구개사각골)과 아래턱(메켈 연골) 사이에서 형성되며, 위턱과 설궁(hyoid arch) 사이의 관절 증거는 발견되지 않았다.[2]
아래턱(메켈 연골)에는 위턱과의 관절 바로 앞에 독특한 추가 돌기가 있는데, 이는 턱이 닫힐 때 나선형 치아가 연골두개를 손상시키는 것을 막는 역할을 했을 것으로 추정된다. 또한 ''헬리코프리온''의 특징적인 점은 입술 연골이 아래턱 윗면과 연골결합(synchondrosis, 융합된 관절)을 이룬다는 것이다. 이 입술 연골은 나선형 치아의 뿌리 부분에서 넓어져 치아를 옆에서 지지하고, 턱이 닫힐 때 가해지는 힘을 분산시키는 데 도움을 주었다. 입술 연골의 뒤쪽은 컵 모양으로, 새로 자라나는 가장 어린 치아의 뿌리를 보호하는 역할도 했다.[2][8][5]
나선형 치아의 정확한 위치는 100년 이상 논쟁거리였다. 1899년 처음 발견되었을 때는 위턱 바깥쪽에 위치한다고 생각되었으나, 이후 아래턱에 위치한다는 복원도 제안되었다. 심지어 등지느러미나 꼬리지느러미의 방어용 구조물이라는 설도 있었다.[40] 1960년대에 가까운 관계인 ''Ornithoprion''의 두개골 화석이 발견되면서 나선형 치아가 아래턱에 위치한다는 것이 확인되었다.[44] 1994년에는 미술가 레이 트롤 등이 아래턱 끝에 치아가 돌출된 모습으로 복원하기도 했다.[45] 2008년 스미스소니언 박물관의 감수를 받은 복원에서는 치아가 목구멍 안쪽에 위치하는 것으로 그려졌으나,[46] 이 해석은 다른 연구에서 받아들여지지 않았다.[47][48]
2013년 CT 스캔 연구를 통해 나선형 치아가 아래턱 중앙의 치아유합부(symphysis)에 위치하며, 턱뼈 전체를 차지한다는 사실이 최종적으로 밝혀졌다. 이 연구는 또한 ''헬리코프리온''이 완악류(현생 키메라 포함)의 특징인 자동이형성 턱 구조를 가졌음을 확인했다. 위턱에는 큰 이빨이 없었기 때문에, 먹이를 으깨기보다는 아래턱의 나선형 치아를 이용해 먹이를 자르거나 베어서 삼켰을 것으로 추정된다.[40]
2. 4. 비늘
카자흐스탄에서 발견된 ''H. 베소노비''의 유해와 함께 이빨 모양의 연골어류 비늘, 특히 치상돌기(placoid scale)로 알려진 비늘이 발견되었다. 이 비늘은 ''사르코프리온''이나 ''오르니토프리온''과 같은 다른 유게네오돈트의 비늘과 대체로 유사한 특징을 보인다.[37]헬리코프리온의 비늘은 오목한 아랫면을 가진 뚜껑 모양의 기저부(base)를 가지고 있다. 비늘의 윗부분인 크라운(crown)은 원뿔 모양이며, 표면에는 톱니 모양의 세로 융기(ridge)가 덮여 있다. 비늘은 하나의 기저부에 하나의 크라운이 있는 단일 치상돌기 형태일 수도 있고, 여러 치상돌기가 융합되어 여러 개의 크라운이 묶인 다중 치상돌기 형태일 수도 있다. 다른 유게네오돈트와 비교했을 때, 헬리코프리온의 비늘은 더 강하게 뾰족한 형태를 띠는 것이 특징이다.[37]
3. 고생물학

카자흐스탄에서 발견된 ''H. 베소노비''의 유해와 함께 이빨 모양의 연골어류 비늘, 특히 치상돌기가 발견되었다. 이 비늘은 ''사르코프리온''과 ''오르니토프리온''과 같은 다른 유게네오돈트류의 비늘과 대체로 유사하다. 비늘은 오목한 아랫면을 가진 뚜껑 모양의 기저부를 가지며, 크라운은 원뿔 모양이고 톱니 모양의 세로 융기로 덮여 있다. 비늘은 단일 치상돌기(기저부당 하나의 크라운) 또는 다중 치상돌기(여러 치상돌기가 융합되어 여러 개의 크라운이 묶인 형태)일 수 있다. 다른 유게네오돈트류와 비교했을 때, ''헬리코프리온''의 비늘은 더 뾰족한 형태를 띤다.[37]
''헬리코프리온''은 전기 톱을 연상시키는 나선형으로 감긴 독특한 치아 구조를 가지고 있다. 이빨 소용돌이는 아래턱 중앙에 위치하며, 이는 ''헬리코프리온''이 속한 유게네오돈트류의 공통적인 특징으로, 좌우 턱의 치아가 정중선상에서 합쳐져 형성된 것이다.[39] 새롭고 큰 이빨은 나선 바깥쪽에 계속 추가되고, 오래되고 작은 이빨은 나선 안쪽으로 말려 들어가기 때문에, 개체가 태어나서 죽을 때까지 사용한 모든 이빨이 이 소용돌이 안에 보존된다.
2013년의 연구에서는 CT 스캔을 통해 이빨 소용돌이와 두개골의 정확한 위치 관계가 밝혀졌다. 이빨 소용돌이는 턱 뒤쪽에 위치하며 턱뼈궁 전체를 차지하고 있었다. 또한, 턱궁과 신경두개가 두 곳에서 관절하는 완두아강 특유의 구조(Autodiastylic)도 확인되었다. 위턱에는 이빨이 존재하지 않는 것으로 밝혀졌는데, 이를 통해 ''헬리코프리온''은 먹이를 씹어 으깨는 방식이 아니라, 아래턱의 이빨 소용돌이를 이용해 먹이를 자르고 삼켰을 것으로 추정된다.[40]
특이한 톱날 모양의 이빨 소용돌이와 이빨에 마모 흔적이 거의 없다는 점은 ''헬리코프리온''이 주로 부드러운 몸체를 가진 먹이를 먹었음을 시사한다. 단단한 껍질을 가진 먹이는 이빨 구조상 입에서 빠져나가기 쉬웠을 것이다. 턱의 폭이 좁아 흡입 섭식은 비효율적이었을 것으로 보이며, 대신 물어뜯는 방식으로 먹이를 섭취했을 가능성이 높다. Ramsay 등이 2015년에 수행한 생체역학 모델링 연구에 따르면, 이빨 소용돌이 내의 이빨들은 나선형 위치에 따라 기능이 분화되어 있었다. 가장 앞쪽의 이빨은 먹이를 걸어 입 안으로 당기는 역할을 했고, 중간 이빨은 먹이를 찌르고, 뒤쪽 이빨은 먹이를 꿰뚫어 목구멍으로 밀어 넣는 역할을 했다. 이 과정에서 먹이는 이빨 소용돌이와 위턱의 구개사각골 사이에서 짓눌렸을 것으로 추정된다. 입술 연골은 이빨 소용돌이를 지지하는 역할을 했다.[8]
''헬리코프리온''은 먹이를 잡을 때 처음에는 턱을 넓게 벌렸다가, 이후 현대의 물어뜯는 상어처럼 턱을 작게 열고 닫는 동작을 반복하며 먹이를 입 안쪽으로 운반했을 수 있다. 현대 상어는 먹이를 자르기 위해 머리를 좌우로 흔들지만, ''헬리코프리온''은 호 모양으로 움직이는 아래턱 이빨 때문에 턱을 열 때 오히려 먹이를 더 효과적으로 잘랐을 것으로 보인다. 이는 칼로 베는 동작과 유사하다. ''헬리코프리온''은 빠르고 강력하게 턱을 닫아 먹이를 포획하고 입 안 깊숙이 밀어 넣은 다음, 턱을 주기적으로 열고 닫으며 먹이를 톱질하듯 잘랐을 가능성이 높다.[8]
Ramsay 등은 이빨 소용돌이가 초기 페름기 바다에 풍부했던 암모나이트나 앵무조개와 같은 단단한 껍질을 가진 두족류의 껍질을 제거하는 데 효과적이었을 수 있다고 제안했다. 만약 이들을 정면으로 물었다면, 이빨 소용돌이가 부드러운 몸체를 껍질 밖으로 끌어당기는 역할을 했을 수 있다. 턱이 닫힐 때 구개사각골과 이빨 소용돌이는 역3점 굽힘 시험과 유사한 3점 지지 시스템을 형성하여, 부드러운 부분을 고정시키고 절단 효율을 높이며 단단한 껍질에 대한 지렛대 역할을 했을 것이다. 이 세 지점에서의 추정 물기 힘은 1,192 ~ 2,391 N 사이였고, 초기 먹이 접촉 시의 추정 물기 응력은 397 ~ 797 MPa 범위였다. 이 강력한 물기 힘은 ''헬리코프리온''이 껍질 없는 경골어류나 다른 연골어류의 뼈를 자를 수 있게 하여, 식단을 척추동물까지 확장할 수 있었음을 시사한다.[8]
''헬리코프리온''의 크기는 이빨 소용돌이의 직경을 통해 추정된다. 이빨 소용돌이 직경이 40cm인 개체는 몸길이가 5m에서 8m에 달했을 것으로 추정되며[47], 발견된 가장 큰 이빨 소용돌이의 직경은 약 56cm에 이른다.[38]
다른 화석 어류 중에서도 유사한 형태의 이빨을 가진 경우가 있다. 예를 들어, 오니코두스는 턱 끝부분에 이와 비슷한 형태의 이빨 몇 개를 가지고 있었다.
4. 생존 시기, 서식지 및 화석 발견
(내용 없음)
4. 1. 생존 시기 및 서식지
헬리코프리온은 고생대 페름기부터 중생대 트라이아스기까지, 약 3억 년 전부터 2억 년 전까지 생존했던 어종이다.[41] 생존 당시에는 주로 중국과 일본을 중심으로 하는 바다에 서식했을 것으로 추정된다.최초의 화석은 1899년 러시아의 우랄산맥에서 고생물학자 카르핀스키에 의해 발견되었다. 이후 전 세계적으로 광범위한 지역에서 화석이 발견되었으며, 주요 발견 지역은 다음과 같다.
헬리코프리온은 석탄기 후기의 해성층에서 처음 나타나기 시작하여 페름기에 그 수가 크게 증가했다.[41] 페름기에서 초기 트라이아스기까지 매우 번성했으나, 이후 다른 해양 파충류나 해룡과의 경쟁에서 밀려 멸종했을 것으로 추정된다. 화석은 페름기에 형성된 지층에서 가장 많이 발견된다.
이 속은 곤드와나 대륙의 남쪽 해안에서 처음 나타나 분화한 뒤, 이후 판게아 대륙 전역으로 퍼져나간 것으로 보인다.[41] 일본에서는 군마현 미도리시 (구 세타군)의 아시오층군에서 ''H. bessonowi'' 종의 화석이 발견되었으며,[42] 미야기현 게센누마시에서도 발견 보고가 있다.[43] 근연속인 ''Sinohelicoprion''과 ''Hunanohelicoprion''은 중국에서 함께 발견되었다.[41]
4. 2. 화석 발견
헬리코프리온 화석은 1899년 러시아의 우랄산맥에서 고생물학자 카르핀스키에 의해 처음 발견되었다.[1] 이후 중국, 일본, 서부 오스트레일리아, 카자흐스탄, 라오스, 노르웨이, 캐나다, 미국 등 북아메리카 지역과 멕시코, 유럽 국가 등 전 세계적으로 광범위하게 발견되었다.[1] 화석은 주로 고생대 페름기에 형성된 지층에서 가장 많이 발견된다.[1]다른 연골어류처럼 헬리코프리온과 다른 유지어류는 연골로 이루어진 골격을 가졌다. 이 때문에 라거슈테트와 같이 특별한 보존 환경이 아니라면 몸 전체가 쉽게 분해되어 완전한 형태의 화석을 찾기 어렵다.[3][4][37] 따라서 헬리코프리온의 정확한 전체 모습을 파악하는 데 한계가 있다. 하지만 펜실베이니아기부터 트라이아스기까지 살았던 케이스오돈토이드류인 ''Caseodus'', ''Fadenia'', ''Romerodus'' 등 일부 유지어류에서 척추뼈 화석이 발견되어 이를 통해 몸의 형태를 추정할 수 있다.[3][4][37]
이들 유지어류의 화석을 통해 추정된 헬리코프리온의 몸은 방추형(유선형)으로, 삼각형 모양의 가슴지느러미를 가졌을 것으로 보인다. 또한 지느러미 가시가 없는 크고 삼각형 모양의 등지느러미 하나와, 외부적으로 두 엽의 크기가 같은 동형꼬리 형태의 높고 갈라진 꼬리지느러미를 가졌을 것이다.[3][4] 이는 참치, 새치, 청상아리와 같이 활동적으로 넓은 바다를 헤엄치는 포식성 어류와 유사한 형태이다. 유지어류는 골반 지느러미와 뒷지느러미가 없으며, ''Romerodus''의 경우 몸 측면을 따라 꼬리지느러미까지 넓은 용골이 있었을 가능성이 있다.[3][4] ''Fadenia''는 잘 드러난 5개의 아가미 구멍과 함께 흔적적인 여섯 번째 아가미 구멍이 있었을 수 있으며, 현생 키메라에서 보이는 특수한 아가미 바구니나 살덩이 형태의 아가미 덮개는 없었던 것으로 보인다.[3][4]
헬리코프리온의 크기는 이빨 소용돌이 화석의 크기를 바탕으로 추정된다. 레베데프는 지름이 35cm 에서 40cm에 달하는 이빨 소용돌이를 가진 개체가 현생 돌묵상어와 비슷한 크기인 5m 에서 8m 길이에 달했을 수 있다고 제안했다.[37] 현재까지 알려진 가장 큰 헬리코프리온 이빨 소용돌이 표본(IMNH 49382)은 지름 56cm, 치관 높이 14cm에 달하며, 이는 몸길이가 7.6m가 넘는 거대한 개체의 것으로 추정된다.[5][19]
헬리코프리온은 석탄기 후기의 해성층에서 처음 나타나기 시작하여 페름기에 그 수가 크게 증가했다. 화석은 우랄 산맥, 서호주, 중국(근연종인 ''Sinohelicoprion''과 ''Hunanohelicoprion'' 포함),[41] 북아메리카 서부(캐나다에서 멕시코까지) 등 넓은 지역에서 발견된다. 이는 헬리코프리온 속이 곤드와나 대륙 남쪽 해안에서 처음 나타나 이후 판게아 대륙 전역으로 퍼져나갔음을 시사한다.
일본에서도 군마현 세타군의 아시오층군에서 ''H. bessonowi'' 종의 화석이 발견되었으며,[42] 미야기현 게센누마시에서도 발견 보고가 있다.[43]
5. 분류
IMNH 37899 표본의 두개골 데이터는 헬리코프리온이 연골어류의 하위 그룹인 진연골두류에 속한다는 것을 보여준다. 이는 통합된 설골(혀뼈 지지 구조)이 없고 턱 구조가 분리되는 형태(자가분리형 턱 현탁액)와 같은 특징들로 확인된다.[9] 진연골두류는 대부분 멸종했으며, 현존하는 종류는 전두어강의 키메라목(키메라 또는 쥐상어)뿐이다. 이들은 상어나 가오리 등이 속하는 판새아강과는 자매군 관계이지만, 헬리코프리온과 현존 키메라의 관계는 매우 멀다.[9][2]
헬리코프리온은 더 구체적으로 데본기부터 트라이아스기까지 살았던 상어와 유사한 진연골두류 목인 에우게네오돈트목에 속한다. 에우게네오돈트류는 단순하고 분리된 형태의 두개골, 가장자리 치아 감소, 그리고 턱 중앙선에 칼날 모양으로 확대된 톱니(치열)를 가진 것이 특징이다.
에우게네오돈트목 내에서 헬리코프리온은 키가 크고 각진 융합 치아를 가진 에데스토이데아 그룹에 포함된다. 에데스토이데아는 치열 형태에 따라 두 과로 나뉜다. 하나는 에데스투스과로, 비교적 짧은 이빨 날과 뒤로 기울어진 뿌리를 가진다.[3][5]
헬리코프리온이 속하는 다른 과는 헬리코프리온과이다. 이 과는 때때로 속 ''아가시조두스''의 이름을 따 아가시조돈과(Agassizodontidae)로 불리기도 하지만,[3][5] 헬리코프리온과라는 명칭이 70년 먼저 사용되었다. 헬리코프리온과는 크고 강하게 아치형을 이루며 연골로 지지되는 소용돌이 모양의 치열을 가진다. 이들은 이빨을 탈락시키지 않고, 소용돌이 위쪽에서 앞으로 기울어진 기저부를 가진 새로운 이빨을 계속 추가하는 방식으로 치열이 성장한다.[3][37] 대부분의 에우게네오돈트류 화석은 단편적인 이빨 잔해로만 발견되기 때문에, 이 그룹 내의 구체적인 계통 발생 관계는 아직 불분명하다.[5]
5. 1. 헬리코프리온 베소노위 (H. bessonowi)

''헬리코프리온 베소노위''(''H. bessonowi'')는 1899년 알렉산드르 카르핀스키가 러시아 우랄 산맥의 아르틴스키안(페름기 전기) 석회암 지층인 Divya Formation에서 발견된 여러 표본을 바탕으로 처음 논문에 기술한 종이다.[16][51] 비록 처음 기술된 ''헬리코프리온'' 종은 아니었지만, 완전한 치열 화석이 처음 발견되어 ''헬리코프리온''이 ''에데스투스''와 다르다는 것을 명확히 보여주었기 때문에[16] ''H. bessonowi''는 ''헬리코프리온'' 속의 기준종 역할을 한다.[17] ''H. bessonowi'' 표본은 일본의 Tanukihara Formation[18] 과 카자흐스탄의 아르틴스키안 시대 지층에서도 발견된다.[37] 이 종은 짧고 좁게 간격을 둔 치열, 뒤쪽으로 향하는 치아 끝, 둔각을 이루는 치아 기저부, 그리고 지속적으로 좁은 치열 축으로 다른 ''헬리코프리온'' 종들과 구별된다.[19]
1939년 휠러가 기술한 ''H. nevadensis''는 1929년 네바다의 한 광산에서 발견된 단일 부분 화석(UNMMPC 1001, 현재 분실됨)을 기반으로 한다.[20] 휠러는 이 화석이 아르틴스키안 시대의 로체스터 트라키테 퇴적층에서 유래했다고 보고했으나, 해당 지층은 실제로는 트라이아스기이며 화석의 실제 연대는 불분명하다. 휠러는 치열 팽창 패턴과 치아 높이로 ''H. nevadensis''를 ''H. bessonowi''와 구별했지만, 2013년 라이프 타파닐라와 제시 프루잇은 이 특징들이 특정 발달 단계에서의 ''H. bessonowi''와 일치한다고 주장하며 동종이명일 가능성을 제기했다.[19]
노르웨이 스피츠베르겐 섬에서 발견된 표본을 바탕으로 1970년 스타니스와프 시에들레키가 기술한 ''H. svalis''는 매우 큰 치열(모식 표본 PMO A-33961)을 가지며, 치아가 서로 접촉하지 않는 특징이 보고되었다. 그러나 이는 치아 중앙 부분만 보존되었기 때문일 수 있다. 치열 축이 부분적으로 가려져 있어 확실한 분류는 어렵지만, 여러 비율 면에서 ''H. bessonowi''와 유사하다. 최대 회전 높이가 72mm인 ''H. svalis''는 가장 큰 ''H. bessonowi'' (최대 회전 높이 76mm)와 크기가 비슷하다.[19]
1999년, ''H. bessonowi''의 모식 표본이 도난당하는 사건이 있었으나, 익명의 화석 거래상을 통해 곧 회수되었다.[21]
''헬리코프리온''의 치열 위치에 대한 해석은 오랫동안 논쟁거리였다. 20세기 중반까지는 일반적으로 아래턱에 위치한다고 여겨졌으나, 카르핀스키의 발표 직후부터 의문이 제기되었다. 1900년대 중반, "F. 존"이라는 예술가는 치열이 아래턱에서 바깥으로 말려 내려오는 모습으로 묘사했으며, 존 A. 롱, 토드 마샬, 캐런 카 등 현대의 고생물학자 및 예술가들도 유사한 재구성을 제시했다. 이들은 치열이 톱상어처럼 먹이를 치는 데 사용되었을 것으로 추정했다.[37][2]
1966년, 라이너 장겔이 신종 유게노돈트 ''오르니토프리온''을 기술하면서 치열 위치에 대한 중요한 단서가 나왔다. ''오르니토프리온''은 아래턱 기저부 중앙, 즉 턱뼈가 만나는 접합 부위에 작은 치열을 가지고 있었다.[32] 같은 해, 벤딕스-알름그린은 ''헬리코프리온''의 치열 역시 메켈 연골 사이에 끼워진 접합 구조로 재해석했다. 그는 치열이 아래턱 뒤쪽 접합부에서 형성되어 위로, 앞으로, 아래로, 안쪽으로 나선형으로 이동하며 축적되고, 연골 고리에 의해 지지된다고 설명했다. 이 재구성에 따르면 치열은 위턱에 대고 먹이를 자르는 역할을 했을 것이다.[6]
1994년, 브래드 매첸과 예술가 레이 트롤은 벤딕스-알름그린의 연구를 바탕으로, 위턱에는 압착용 치아 외에는 다른 치아가 없고, 고블린 상어처럼 긴 코를 가진 모습으로 ''헬리코프리온''을 재구성했다.[33] 1996년 필립 자비에의 교과서에서도 유사하게 위턱 앞쪽에 날카로운 치아, 뒤쪽에 압착용 치아를 가진 모습으로 묘사했다.[34]
2008년, 메리 패리쉬는 스미소니언 자연사 박물관 전시를 위해 치열을 목구멍 깊숙이 위치시키는 새로운 재구성을 제시했다. 이는 치아에 마모 흔적이 없고, 턱 앞쪽에 치열이 있으면 저항을 유발할 것이라는 가정에 기반하여, 치열이 삼키는 것을 돕는 구조라고 주장했다.[35] 그러나 이 재구성은 단순히 삼키는 기능만을 위해 지나치게 복잡한 구조라는 비판을 받았다.[36]
2009년, 레베데프는 카자흐스탄에서 발견된 치열의 긁힌 자국과 함께 발견된 ''캄포두스''와 유사한 넓은 결절 치아를 근거로, 벤딕스-알름그린과 트롤의 재구성을 지지했다. 그는 긴 아래턱 앞쪽 접합부에 연골로 보호된 치열이 있고, 입을 다물면 위턱의 홈에 들어가며, 양쪽 턱 가장자리에는 ''캄포두스'' 같은 치아가 덮여 있었을 것으로 보았다. 이는 ''사르코프리온'' 등 다른 관련 속에서 보고된 것과 유사하다. 그는 치아가 아래턱 끝에 집중된 현대의 향유고래, 꼬마향유고래 및 피그미향유고래, 리스 돌고래, 부리고래 등과 생태적으로 비교했다.[37] 레베데프의 재구성은 현대적인 관점에 가깝지만, 가설적인 긴 턱과 측면 치아는 이후 CT 데이터 분석을 통해 수정되었다.[2]
5. 2. 헬리코프리온 다비시 (H. davisii)
헬리코프리온 다비시(''Helicoprion davisii'')의 최초 표본은 서호주 박물관(WAMAG) 9080으로,[19] 서호주 가스코인 강의 지류에서 발견된 15개의 치아 조각이다. 1886년 헨리 우드워드는 이 화석을 묘사하고 발견자의 이름을 따 ''에데스투스 데이비시(Edestus davisii)''라고 명명했다.[10] 1899년 알렉산더 카르핀스키는 ''H. 베소노비(H. bessonowi)''를 명명하면서 ''E. 데이비시''를 ''헬리코프리온(Helicoprion)'' 속으로 재분류했다.[16] 1902년 찰스 R. 이스트먼은 ''H. 데이비시''를 자신이 새로 만든 속인 ''캄필로프리온''(Campyloprion)으로 옮기려 했으나, 이 제안은 널리 받아들여지지 않았다.[11] 카르핀스키가 ''E. 데이비시''를 ''헬리코프리온''의 한 종으로 확인한 것은 1930년대 후반 커트 타이터트에 의해 지지되었는데, 그는 서호주의 완다지 지층에서 더 완전한 여러 개의 치열 화석을 묘사했다.[12]
1907년과 1909년에 올리버 페리 헤이는 인산염이 풍부한 포스포리아 지층에서 발견된 여러 화석을 바탕으로 새로운 속과 종인 ''리스소프리온 페리에리(Lissoprion ferrieri)''를 묘사했다. 그는 또한 ''H. 데이비시''를 자신이 명명한 새로운 속 및 종과 동일한 것으로 간주했다.[13] 그러나 1911년 카르핀스키는 두 종을 다시 분리하고, ''H. 페리에리''를 ''헬리코프리온'' 속으로 옮겼다.[14] ''H. 페리에리''는 처음에는 치아 각도와 높이 측정값을 기준으로 ''H. 데이비시''와 구별되었지만, 2013년 타파닐라와 프루이트는 이러한 특징들이 종 내에서도 다양하게 나타날 수 있다고 보았다. 그 결과, 그들은 ''H. 페리에리''를 ''H. 데이비시''의 주니어 동의어로 재분류했다. ''H. 데이비시'' 표본은 포스포리아 지층 외에도 멕시코, 텍사스, 캐나다 (누나부트와 앨버타)에서도 발견되었다. 이 종은 키가 크고 간격이 넓은 치아 고리를 특징으로 하며, 이는 나이가 들수록 더 뚜렷해진다. 또한 치아가 눈에 띄게 앞으로 휘어져 있다.[19]

1939년 출판물에서 해리 E. 휠러는 캘리포니아와 네바다에서 발견된 화석을 바탕으로 헬리코프리온의 두 가지 새로운 종을 묘사했다. 이 중 하나인 ''헬리코프리온 시에렌시스(Helicoprion sierrensis)''는 동부 캘리포니아의 빙하 모레인 퇴적물에서 발견된 표본(UNMMPC 1002)을 기준으로 묘사되었으며, 이 표본은 굿휴 지층에서 유래한 것으로 추정된다.[20] 타파닐라와 프루이트는 ''H. 시에렌시스''를 구별하는 특징으로 제시된 샤프트 범위가 ''H. 데이비시''에서 관찰되는 변이 범위 내에 있다고 판단하여, ''H. 시에렌시스'' 역시 ''H. 데이비시''의 동의어로 처리했다.[19]
''H. 징멘엔세(H. jingmenense)''는 2007년에 거의 완전한 치열(YIGM V 25147, 4회전 이상) 화석을 바탕으로 묘사되었다. 이 표본은 중국 후베이 성의 하부 페름기 치샤 지층을 통과하는 도로 건설 중에 발견되었다. 이 종은 ''H. 페리에리'' 및 ''H. 베소노비''와 매우 유사하지만, 전자는 더 넓은 절단면과 짧은 복합 뿌리를 가진 치아를 가지고 있고, 후자는 회전당 치아 수가 39개 미만이라는 점에서 차이가 있다고 보고되었다.[15] 그러나 타파닐라와 프루이트는 이 표본이 주변 암석에 의해 부분적으로 가려져 치아 높이가 실제보다 낮게 평가되었을 수 있다고 주장했다. 그들은 종 내 변이를 고려하여 ''H. 징멘엔세'' 또한 ''H. 데이비시''의 동의어로 간주했다.[19]
5. 3. 헬리코프리온 에르가사미논 (H. ergassaminon)

''H. davisii''와 마찬가지로, ''Helicoprion ergassaminon''은 아이다호주의 포스포리아 층군에서 발견되었지만, 비교적 훨씬 희귀하다. 이 종은 1966년 스벤 에리크 벤딕스-알름그린(Svend Erik Bendix-Almgreen)의 모노그래프에서 처음 명명되고 자세히 설명되었다.[6] 모식 표본("Idaho 5")은 먹이를 먹는 데 사용되었음을 보여주는 파손 및 마모 흔적이 있다. 포스포리아 층군에서 발견된 다른 여러 표본에서도 ''H. ergassaminon''이 확인되었지만, 이들 중 마모 흔적을 보이는 것은 없다.
''H. ergassaminon''은 ''H. bessonowi''와 ''H. davisii''가 보여주는 두 가지 대조적인 형태 사이의 중간적인 특징을 가진다. 이빨은 키가 크지만 간격이 좁고, 완만하게 굽어 있으며, 이빨 기저부는 둔각으로 각진 형태를 보인다.[19] 이 종의 모식 표본은 한때 분실된 것으로 여겨졌으나, 2023년에 재발견되어 아이다호 자연사 박물관의 소장품으로 돌아왔다.[22]
참조
[1]
웹사이트
Ancient shark relative had buzzsaw mouth
http://science.nbcne[...]
2013-02-27
[2]
논문
Jaws for a spiral-tooth whorl: CT images reveal novel adaptation and phylogeny in fossil ''Helicoprion''
[3]
서적
Chondrichthyes I – Paleozoic Elasmobranchii
Gustav Fischer Verlag
[4]
논문
New eugeneodontid sharks from the Lower Triassic Sulphur Mountain Formation of Western Canada
https://www.research[...]
2008-01-01
[5]
논문
Saws, Scissors, and Sharks: Late Paleozoic Experimentation with Symphyseal Dentition
2020
[6]
논문
New investigations on ''Helicoprion'' from the Phosphoria Formation of south-east Idaho, USA
https://www.royalaca[...]
1966
[7]
논문
Description of the chimaerid jaw and its phylogenetic origins
https://onlinelibrar[...]
1999-01-01
[8]
논문
Eating with a saw for a jaw: Functional morphology of the jaws and tooth-whorl in ''Helicoprion davisii'': Jaw and Tooth Function in ''Helicoprion''
https://www.research[...]
2014-09-02
[9]
논문
Relationships of the Chimaeriformes and the basal radiation of the Chondrichthyes
https://www.research[...]
1997-03-01
[10]
논문
On a Remarkable Ichthyodorulite from the Carboniferous Series, Gascoyne, Western Australia
https://www.biodiver[...]
1886
[11]
논문
II.—On ''Campyloprion'', a New Form of ''Edestus''-like Dentition
https://www.biodiver[...]
1902-04-01
[12]
논문
Helicoprion in the Permian of Western Australia
1940
[13]
논문
On the nature of Edestus and related genera, with descriptions of one new genus and three new species
https://www.biodiver[...]
1909
[14]
논문
"Замѣчанія о Helicoprion и о другихъ едестидахъ"
https://www.biodiver[...]
1911
[15]
논문
The first record of ''Helicoprion'' Karpinsky (Helicoprionidae) from China
2007-08-01
[16]
논문
Объ остаткахъ едестидъ и о новомъ ихъ родѣ Helicoprion
https://www.biodiver[...]
1899
[16]
논문
Ueber die Reste von Edestiden und die neue Gattung Helicoprion
https://books.google[...]
1899
[17]
논문
''Helicoprion'' from Elko County, Nevada
[18]
논문
On a Fusulina-Limeston with Helicoprion in Japan
1903
[19]
논문
Unravelling species concepts for the ''Helicoprion'' tooth whorl
http://geology.isu.e[...]
2017-07-26
[20]
논문
''Helicoprion'' in the Anthracolithic (Late Paleozoic) of Nevada and California, and its Stratigraphic Significance
[21]
서적
The Dinosaur Dealers
Allen & Unwin
[22]
웹사이트
Fossil Found: Idaho No. 5 Returns to Idaho Museum of Natural History after 60+ Years
https://www.isu.edu/[...]
2024-10-30
[23]
논문
Étude géologique sur le centre de l'Indochine entre Tourane et le Mekong
1933
[24]
논문
A revision of ''Campyloprion'' Eastman, 1902 (Chondrichthyes, Helicoprionidae), including new occurrences from the Upper Pennsylvanian of New Mexico and Texas, USA
https://geojournals.[...]
2018
[25]
논문
''Helicoprion'' (''Parahelicoprion'' n.g.) ''clerci''
[26]
논문
Artinskian (Early Permian) Sea Basin and Its Biota (Krasnoufimsk, Cis-Urals)
[27]
논문
Karpinsky's Genus ''Helicoprion''
https://www.journals[...]
1900
[28]
논문
Ueber die Reste von Edestiden und die neue Gattung Helicoprion
https://www.biodiver[...]
1899-11-21
[29]
논문
Ce que doit signifier la spirale de Helicoprion
https://www.biodiver[...]
1899-11-21
[30]
논문
Note Sur l'Helicoprion et les Edestides
https://www.biodiver[...]
1899-12-19
[31]
논문
Note sur ''Helicoprion bessonowi'' (Karpinsky)
https://www.biodiver[...]
1899-12-19
[32]
논문
A new shark of the family Edestidae, ''Ornithoprion hertwigi'', from the Pennsylvanian Mecca and Logan quarry shales of Indiana
https://www.biodiver[...]
Chicago Field Museum of Natural History
1966-03-17
[33]
서적
Planet Ocean: A Story of Life, the Sea and Dancing to the Fossil Record
Ten Speed Press
[34]
서적
Early Vertebrates
Oxford University Press
[35]
웹사이트
The Orthodonty of ''Helicoprion''
http://paleobiology.[...]
Smithsonian
2008-02-29
[36]
잡지
Unraveling the Nature of the Whorl-Toothed Shark
https://www.wired.co[...]
Wired
2012-09-23
[37]
학술지
A new specimen of ''Helicoprion'' Karpinsky, 1899 from Kazakhstanian Cisurals and a new reconstruction of its tooth whorl position and function
https://www.research[...]
[38]
학술지
Unraveling species concepts for the Helicoprion tooth whorl
https://www.research[...]
2013-11
[39]
웹사이트
Eugnathostomata GEOL 431 Vertebrate Paleobiology
http://www.geol.umd.[...]
2014-11-10
[40]
학술지
Jaws for a spiral-tooth whorl: CT images reveal novel adaptation and phylogeny in fossil ''Helicoprion''
[41]
학술지
The first record of ''Helicoprion'' Karpinsky (Helicoprionidae) from China
http://link.springer[...]
2007-08
[42]
학술지
群馬県桐生市の足尾帯のペルム系からクテナカントゥス科サメ類の新産出
[43]
학술지
宮城県気仙沼市より軟骨魚類ヘリコプリオン属化石の発見
[44]
논문
A new shark of the family Edestidae, Ornithoprion hertwigi, from the Pennslyvanian Mecca and Logan quarry shales of Indiana
https://archive.org/[...]
Field Museum of Natural History
[45]
기사
Planet Ocean: A Story of Life, the Sea, and Dancing to the Fossil Record
2012-10-25
[46]
웹사이트
The Orthodonty of Helicoprion
http://paleobiology.[...]
Smithsonian Institution
2008-02-29
[47]
학술지
A new specimen of ''Helicoprion'' Karpinsky, 1899 from Kazakhstanian Cisurals and a new reconstruction of its tooth whorl position and function
[48]
웹사이트
Unraveling the Nature of the Whorl-Toothed Shark
http://www.wired.com[...]
Wired
2008-02-29
[49]
서적
石炭紀・ペルム紀の生物
技術評論社
[50]
서적
大昔の生きもの
https://www.worldcat[...]
ポプラ社
2014
[51]
학술지
''Helicoprion'' from Elko County, Nevada
http://www.jstor.org[...]
[52]
학술지
The first record of Helicoprion Karpinsky (Helicoprionidae) from China
[53]
학술지
''Helicoprion'' in the Anthracolithic (Late Paleozoic) of Nevada and California, and it’s Stratigraphic Significance
http://www.jstor.org[...]
[54]
학술지
Eating with a saw for a jaw: Functional morphology of the jaws and tooth-whorl in Helicoprion davisii
https://www.research[...]
2015
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com