염색체 역위
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
염색체 역위는 염색체의 일부가 잘린 후 뒤집혀 재결합되는 염색체 구조적 이상을 의미한다. 1921년 알프레드 스터티번트가 초파리에서 최초로 발견했으며, 진핵생물에서 널리 발견된다. 역위는 중심절 포함 여부에 따라 완간 역위와 완내 역위로 나뉘며, 감수 분열 시 재조합 억제를 통해 유전자의 독립적인 진화를 가능하게 하고, 국소 적응과 종 분화에 기여할 수 있다. 염색체 역위는 세포유전학적 기술, 유전자 분석, 비교 유전체학 등을 통해 탐지할 수 있으며, ISCN 명명법에 따라 표기된다. 역위 보인자는 자손에게 염색체 이상을 유발할 수 있으며, 특정 역위는 울프-허쉬호른 증후군과 같은 유전 질환을 일으키기도 한다.

더 읽어볼만한 페이지
- 돌연변이 - 조로증
조로증은 DNA 복구 과정 결함으로 아동기 또는 청소년기에 노화 징후를 보이는 희귀 유전 질환으로, RecQ 헬리카제, NER 단백질, 라민 등 DNA 복구 단백질 유전자 돌연변이가 주요 원인이며, 베르너 증후군, 블룸 증후군, 로스문드-톰슨 증후군, 콕케인 증후군, 색소피부건조증, 허친슨-길포드 프로제리아 증후군 등이 대표적이다. - 돌연변이 - 돌연변이원
돌연변이원은 DNA에 변화를 일으켜 유전독성을 유발하는 물질 또는 방사선으로, 에임스 검사로 검출이 가능하며 암을 비롯한 질병을 유발할 수 있고, 물리적, 화학적, 생물학적 요인 등 다양한 종류가 존재하여 위험성을 줄이기 위한 연구가 진행 중이다. - 유전 정보 수정 - 형질전환
형질전환은 세균, 효모, 식물, 동물 등 생명체에 외부 DNA를 도입하여 유전적 형질을 변화시키는 기술이며, 자연적 또는 인공적으로 발생하며 유전자 조작에 활용된다. - 유전 정보 수정 - 염색체 전좌
염색체 전좌는 염색체 일부 또는 전체가 다른 염색체로 이동하는 구조 변화로, 유전 정보 변화를 초래하여 다양한 질병을 유발하며, 상호 전좌, 비상호 전좌, 로버트슨 전좌 등으로 분류되고, 가족력이 있는 경우 유전자 검사가 중요하다.
2. 역사
알프레드 스터티번트는 1921년에 초파리에서 염색체 역위 현상을 처음 발견했고,[9] 이후 모든 진핵생물에서 역위가 발견되었다.[3] 초기에는 이형접합체 초파리 유충의 타액선 속 다사 염색체에서 확인되었다.[3] 1970년 테오도시우스 도브잔스키는 역위 내부 유전자가 역위 외부 유전자보다 적합성이 높다고 언급했으나, 추가 연구가 필요하다.[10] 최근에는 키르크패트릭-바튼 모델(2006)을 통해 역위가 적응적 대립 유전자를 연결하여 선택적으로 유리하게 작용한다는 설명이 제시되었다.[11]
2. 1. 초기 연구
알프레드 스터티번트는 1921년에 초파리에서 염색체 역위에 대한 최초의 증거를 발견했다.[9] 이후, 모든 진핵생물에서 역위가 발견되었다.[3] 스터티번트가 발견했을 당시, 역위는 재조합 억제 영역으로 여겨졌다.원래 이러한 역위는 이형접합체 ''초파리'' 유충의 타액선 내 다사 염색체에서 확인되었다.[3] 1970년, 테오도시우스 도브잔스키는 역위 내 유전자가 역위 밖에서 발견되는 유전자보다 더 높은 적합성을 가진다는 것을 언급했지만, 이는 더 연구가 필요한 분야이다.[10]
역위에 대한 최근 모델 중 하나는 키르크패트릭-바튼 모델(2006)인데, 이 모델은 역위가 적응적 대립 유전자를 함께 연결함으로써 선택적으로 유리하다는 것을 명시한다. 여러 유전자에서 공진화된 변이를 역위의 개별 버전(haplotype)으로 물리적으로 연결함으로써, 선택은 이러한 변이를 집단 내에서 높은 빈도로 이끄는 데 더 효율적이어야 한다. 이는 적응적 및 부적응적 대립 유전자를 모두 운반할 수 있는 비역위 영역과는 대조적이다.[11]
2. 2. 도브잔스키의 연구
알프레드 스터티번트는 1921년에 초파리에서 염색체 역위에 대한 최초의 증거를 발견했다.[9] 그 이후, 모든 진핵생물에서 역위가 발견되었다.[3] 스터티번트가 발견했을 당시, 역위는 재조합 억제 영역으로 여겨졌다.원래 이러한 역위는 이형접합체 ''초파리'' 유충의 타액선 내 다사 염색체에서 확인되었다.[3] 테오도시우스 도브잔스키는 1970년에 역위 내 유전자가 역위 밖에서 발견되는 유전자보다 더 높은 적합성을 가진다는 것을 언급했지만, 이는 더 연구가 필요한 분야이다.[10]
2. 3. 현대 연구
알프레드 스터티번트는 1921년에 초파리에서 염색체 역위에 대한 최초의 증거를 발견했다.[9] 그 이후, 모든 진핵생물에서 역위가 발견되었다.[3] 스터티번트가 발견했을 당시, 역위는 재조합 억제 영역으로 여겨졌다.원래 이러한 역위는 이형접합체 ''초파리'' 유충의 타액선 내 다사 염색체에서 확인되었다.[3] 1970년, 테오도시우스 도브잔스키는 역위 내 유전자가 역위 밖에서 발견되는 유전자보다 더 높은 적합성을 가진다는 것을 언급했지만, 이는 더 연구가 필요한 분야이다.[10]
역위에 대한 최근 모델 중 하나는 키르크패트릭-바튼 모델(2006)인데, 이 모델은 역위가 적응적 대립 유전자를 함께 연결함으로써 선택적으로 유리하다는 것을 명시한다. 여러 유전자에서 공진화된 변이를 역위의 개별 버전(haplotype)으로 물리적으로 연결함으로써, 선택은 이러한 변이를 집단 내에서 높은 빈도로 이끄는 데 더 효율적이어야 한다. 이는 적응적 및 부적응적 대립 유전자를 모두 운반할 수 있는 비역위 영역과는 대조적이다.[11]
3. 기전
염색체 역위는 염색체의 구조적 이상 중 하나로, 대부분 정상 표현형을 나타내는 균형 재배열이다. 그러나 생식 세포의 감수분열 단계에서 역위 고리가 만들어지면 염색체의 결실 또는 중복을 보이는 재조합 염색체가 형성되어 자녀에게서 비정상 표현형이 나타날 수 있다. 예를 들어 4번 염색체의 구조적 이상에 동반된 4p 결실은 울프-허쉬호른 증후군을 초래할 수 있다.[22]
부모가 역위 보인자일 경우 자식 세대에 염색체 이상이 일어날 확률이 높아진다. 역위 보인자가 임신할 확률이나 아이에게 위험이 나타날 확률은 역위 부위와 역위된 부분의 크기에 따라 달라진다. Inv(9)와 Inv(2)와 같은 역위형은 많은 사람들이 갖고 있지만 임상소견이 없어 정상 변이형으로 판단하며, 임신이나 태아에 아무런 영향을 주지 않는다.[21]
염색체 역위를 포함하는 염색체의 구조적 이상에도 대부분 정상 표현형을 나타낼 수 있는 이유는 염색체가 곧 유전자는 아니기 때문이다. 이는 마치 컴퓨터의 하드디스크(염색체)가 외관상 변형되었음에도 저장되어 있는 정보들(유전자)이 파괴되지 않았다면 그 정보들을 큰 문제없이 활용할 수 있는 경우와 유사하다.
3. 1. 역위의 유형
염색체 역위는 역위된 부분에 중심절이 포함되는지 여부에 따라 완간 역위와 완내 역위로 나뉜다. 완간 역위는 중심체를 포함하므로 끊어진 부분이 장완과 단완에서 각각 발생한다. 발생 부위는 46,XY,inv(3)(p23q27)와 같이 나타낸다. 반면, 완내 역위는 장완 혹은 단완 내에서만 절단되므로 46,XY,inv(1)(p12p31)와 같이 나타낸다.[21]염색체 역위를 가진 염색체가 비역위 동계 염색체와 짝을 이루는 경우(역위 이형접합체), 감수 분열 동안 제대로 시냅스되지 않아 역위 루프가 형성된다. 루프 내에서 염색체 교차가 일어나면 불균형적인 배우자가 만들어질 수 있다. 중심절 역위에서 유전적 재조합은 이중원심체 염색분체와 무원심체 염색분체를 각각 하나씩 생성한다. 후기 동안 두 재조합체는 모두 문제를 겪는다. 무원심체 염색분체는 한쪽 극으로 끌려가고, 이중원심체 재조합체는 양쪽으로 당겨지면서 이중원심체 다리(bridge)를 형성한다.[12]
주위 역위에서도 유사하게 불균형 염색체가 생성된다. 이러한 교차로 생성된 재조합 염색체에는 결실과 유전자 중복이 나타난다. 이러한 배우자로 인해 생성된 자손은 대부분 생존할 수 없기 때문에, 재조합은 역위된 영역 내에서 간접적으로 억제된다.[12]
3. 2. 감수분열과 영향
염색체 역위는 균형 재배열을 나타내는 염색체의 구조적 이상 중 하나로 대부분 정상 표현형을 나타낸다. 그러나 생식 세포의 감수분열 단계에서 역위 고리를 만들면 염색체의 결실 또는 중복을 보이는 재조합염색체가 형성되어 자녀에게서 비정상 표현형이 나타날 수 있다. 4번 염색체의 구조적 이상에 동반된 4p결실은 울프-허쉬호른 증후군을 초래할 수 있다.[22]부모가 역위 보인자일 경우 자식 세대에 염색체 이상이 일어날 확률이 매우 높아진다. 역위 보인자가 임신할 확률이나 아이에게 위험이 나타날 확률은 역위 부위와 역위된 부분의 크기에 따라 달라진다. 예를 들어 Inv(9)와 Inv(2)와 같은 역위형은 많은 사람들이 갖고 있지만 임상소견이 없어 정상 변이형으로 판단하며, 임신이나 태아에 아무런 영향을 주지 않는다.[21]
염색체 역위를 포함하는 염색체의 구조적 이상에도, 대부분 정상 표현형을 나타낼 수 있는 이유는 염색체가 곧 유전자는 아니기 때문이다.
염색체 역위를 가진 염색체가 비역위 동계 염색체와 짝을 이루는 경우(역위 이형접합체) 감수 분열 동안 적절하게 시냅스되지 않아 역위 루프가 형성된다. 루프 내에서 염색체 교차가 일어나면 불균형적인 배우자가 생성될 수 있다. 중부 역위에서 유전적 재조합은 하나의 이중원심체 염색분체와 하나의 무원심체 염색분체를 생성한다. 후기 동안 두 재조합체는 모두 문제에 직면한다. 무원심체 염색분체는 한쪽 극으로 끌려가고, 이중원심체 재조합체는 두 방향으로 당겨지면서 이중원심체 브리지를 생성한다.[12]
주위 역위에서도 유사한 불균형 염색체가 생성된다. 이러한 교차의 결과로 생성된 재조합 염색체에는 결실과 유전자 중복이 포함된다. 이러한 배우자에 의해 생성된 자손은 대부분 생존할 수 없으므로 재조합은 역위된 영역 내에서 간접적으로 억제된다.[12]
4. 진화적 결과
역위 이형접합체 사이의 재조합 억제는 조상 배열과 역위 배열의 독립적인 진화를 가능하게 한다. 역위 배열에 변이가 없더라도, 부동으로 인해 손실되지 않으면 시간이 지남에 따라 변이가 증가할 수 있다.[13]
염색체 역위는 국소 적응 및 종 분화에 잠재적인 역할을 하여 진화 연구에서 주목받는다. 비재조합 역위 반수체형은 여러 공동 적응 유전자 변이를 포함할 수 있어, 역위가 다른 환경에 대한 국소 적응을 촉진한다고 생각된다. 자연 선택이 이러한 연결된 적응 변이를 집단 내에서 높은 빈도로 유도하기 때문이다.[14] 그러나 역위 반수체형이 재조합되지 않으므로, 역위 내 공동 적응 유전자 변이의 존재를 입증하기는 어렵다. 또한, 염색체 역위의 적응 효과는 연결된 적응 유전자 변이가 실제로 공동 적응한다는 가정에 기반한다. 하지만 이 가정은 집단이 변동하는 선택을 경험하는 상황에서 위배될 수 있다. 따라서 염색체 역위의 적응 중요성은 여전히 열린 문제이다.[15]
역위 다형성은 유전 부동, 선택, 유전자 흐름, 균형 선택 등으로 확립될 수 있다.[13] 역위 염색체와 조상 염색체 간 적합성 차이는 다형성을 생성하거나, 한 염색체의 고정을 초래한다.[16]
역위는 성염색체 진화와 새로운 성염색체의 기원에 필수적이다. 포유류 Y 염색체는 X 염색체와 재조합할 수 없는데, 이는 역위 중첩 때문이다. 성 결정 좌위와 성 반대 유전자 간 재조합률 감소는 선택에 의해 선호된다. 이는 수컷 결정 좌위와 수컷에게 유리한 좌위의 대립 유전자 간 연관 불균형을 유발, 두 좌위를 모두 포함하는 비재조합 블록을 초래하는 역위로 이어진다.[16][17]
역위는 종 분화에 관여한다. 이형접합체 역위는 열등성을 나타내 잡종 적합성 손실, 접합 후 격리를 초래한다. 종 간 선택된 차이를 축적, 접합 전/후 격리를 유발한다.[16] 역위는 지리적 빈도 경사를 형성하며, 이는 국소 적응 역할을 암시한다. ''초파리 수컷''의 역위 3RP가 그 예이다.[16] 역위가 둘 이상 국소 적응 대립 유전자를 포함하면 선택되어 확산될 수 있다. 나비 ''헬리코니우스 누마''에서 색상 제어 유전자 18개가 역위로 연결되어 높은 적합성을 부여한다.[18]
4. 1. 재조합 억제와 진화
1921년, 알프레드 스터티번트는 초파리에서 염색체 역위의 첫 증거를 발견했다.[9] 이후, 모든 진핵생물에서 역위가 발견되었다.[3] 스터티번트가 발견했을 당시, 역위는 재조합 억제 영역으로 여겨졌다.원래 이러한 역위는 이형접합체 ''초파리'' 유충의 타액선 내 다사 염색체에서 확인되었다.[3] 1970년, 테오도시우스 도브잔스키는 역위 내 유전자가 역위 밖에서 발견되는 유전자보다 더 높은 적합성을 가진다고 언급했지만, 이는 더 많은 연구가 필요한 분야이다.[10]
2006년 키르크패트릭-바튼 모델은 역위가 적응적 대립 유전자를 함께 연결함으로써 선택적으로 유리하다고 명시한다. 여러 유전자에서 공진화된 변이를 역위의 개별 버전(haplotype)으로 물리적으로 연결하면, 선택은 이러한 변이를 집단 내에서 높은 빈도로 이끄는 데 더 효율적이다. 이는 적응적 및 부적응적 대립 유전자를 모두 운반할 수 있는 비역위 영역과는 대조적이다.[11] 역위 이형접합체 사이의 재조합 억제는 조상 배열과 역위 배열의 독립적인 진화를 가능하게 한다. 처음에는 역위 배열에 변이가 없는 반면, 조상 배열에는 변이가 있다. 만약 역위 반수체형이 부동 등으로 인해 손실되지 않으면, 역위 배열의 변이는 시간이 지남에 따라 증가할 수 있으며, 더 많은 동형 접합체가 도입됨에 따라 역위 영역의 재조합률이 다소 회복된다.[13]
염색체 역위는 국소 적응 및 종 분화에 잠재적인 역할을 하기 때문에 진화 연구에서 많은 주목을 받았다. 비재조합 역위 반수체형은 여러 공동 적응 유전자 변이를 포함할 수 있으므로, 역위는 다른 환경에 대한 국소 적응을 촉진하는 것으로 생각된다. 그 이유는 자연 선택이 이러한 연결된 적응 변이를 집단 내에서 높은 빈도로 유도하는 데 더 효율적이기 때문이다.[14] 그러나 역위 반수체형이 재조합되지 않기 때문에, 역위 내에서 연결된 공동 적응 유전자 변이의 존재를 경험적으로 입증하는 것은 어렵다. 또한, 다른 환경에 대한 적응에 대한 염색체 역위의 이러한 가능한 긍정적인 효과는, 별개의 역위 반수체형으로 연결된 적응 유전자 변이가 실제로 공동 적응한다는 가정에 근거한다. 그러나 이 아이디어는 집단이 공간적으로 또는 시간적으로 변동하는 선택을 경험하는 상황에서 위배될 가능성이 높다. 역위 연결 변이에 대한 변동 선택으로 인해, 별개의 유전자 변이를 포함하는 역위 반수체형 간의 재조합이 없으면, 별개의 환경에 대한 적응을 돕기보다는 제한할 수 있다.[15] 따라서, 다른 환경에 대한 적응에서 염색체 역위의 중요성은 진화 유전학에서 여전히 열린 경험적 문제로 남아있다.
역위 다형성은 두 가지 방법으로 확립될 수 있다. 유전 부동 또는 선택은 국소 집단에서 역위의 고정을 초래할 수 있다. 역위 다형성은 이 집단과 역위가 없는 집단 간의 유전자 흐름의 결과로 나타날 수 있다. 균형 선택은 빈도 의존성 또는 과우성에 의해 역위 다형성을 초래할 수도 있다.[13] 역위 염색체와 조상 염색체 간의 적합성 차이는 안정적인 다형성을 생성하거나, 한 염색체의 고정을 초래할 수 있다.[16]
역위는 성염색체 진화에 필수적이었다. 포유류에서 Y 염색체는 거의 전체 길이에 걸쳐 X 염색체와 재조합할 수 없다. 이 비재조합 부분은 일련의 역위가 중첩되어 나타난다. 성 결정 좌위와 성 반대 유전자 간의 재조합률 감소는 선택에 의해 선호된다. 이는 수컷 결정 좌위와 수컷에게 유익한 다른 좌위의 대립 유전자 간의 연관 불균형을 유발한다. 이는 포유류 Y 염색체의 경우처럼, 두 좌위를 모두 포함하는 비재조합 블록을 초래하는 역위를 통해 발생할 수 있다.[16]
역위는 또한 새로운 성염색체의 기원에 필수적일 수 있다. 역위는 성 결정 돌연변이와 성 반대 좌위 간의 연관 불균형을 유발하고, 상염색체로부터 새로운 성염색체를 만들 수 있다.[17]
역위는 여러 가지 방식으로 종 분화에 관여할 수 있다. 이형접합체 역위는 열등성을 나타낼 수 있으므로, 잡종 적합성 손실을 유발하여 접합 후 격리를 초래할 수 있다. 또한 종 간의 선택된 차이를 축적하여 접합 전 및 접합 후 격리를 모두 유발할 수 있다.[16]
역위는 종종 지리적 빈도 경사를 형성하며, 이는 국소 적응에서의 역할을 암시할 수 있다. 이러한 경사의 두드러진 예는 세 대륙에서 관찰될 수 있는 ''초파리 수컷''의 역위 3RP이다.[16] 역위가 두 개 이상의 국소 적응 대립 유전자를 포함하는 경우, 선택되어 확산될 수 있다. 예를 들어, 나비 ''헬리코니우스 누마''에서 색상을 제어하는 18개의 유전자가 역위에 의해 함께 연결되어, 함께 더 높은 적합성을 부여한다.[18]
4. 2. 국소 적응과 종 분화
알프레드 스터티번트가 1921년에 초파리에서 염색체 역위의 최초 증거를 발견한 이후,[9] 모든 진핵생물에서 역위가 발견되었다.[3] 염색체 역위는 국소 적응 및 종 분화에 잠재적인 역할을 하기 때문에 진화 연구에서 많은 주목을 받았다.역위는 종종 지리적 빈도 경사를 형성하는데, 이는 국소 적응에서의 역할을 암시한다. 예를 들어, 나비 ''헬리코니우스 누마''에서 색상을 제어하는 18개의 유전자가 역위에 의해 함께 연결되어, 함께 더 높은 적합성을 부여한다.[18]
키르크패트릭-바튼 모델(2006)에 따르면, 역위는 적응적 대립 유전자를 함께 연결함으로써 선택적으로 유리하다. 여러 유전자에서 공진화된 변이를 역위의 개별 버전(haplotype)으로 물리적으로 연결함으로써, 선택은 이러한 변이를 집단 내에서 높은 빈도로 이끄는 데 더 효율적이다. 비재조합 역위 반수체형은 여러 공동 적응 유전자 변이를 포함할 수 있으므로, 역위는 다른 환경에 대한 국소 적응을 촉진하는 것으로 생각된다.[11]
그러나 역위 반수체형이 재조합되지 않기 때문에, 역위 내에서 연결된 공동 적응 유전자 변이의 존재를 경험적으로 입증하는 것은 어렵다. 또한, 다른 환경에 대한 적응에 대한 염색체 역위의 이러한 가능한 긍정적인 효과는, 별개의 역위 반수체형으로 연결된 적응 유전자 변이가 실제로 공동 적응한다는 가정에 근거한다. 그러나 이 아이디어는 집단이 공간적으로 또는 시간적으로 변동하는 선택을 경험하는 상황에서 위배될 가능성이 높다. 따라서, 다른 환경에 대한 적응에서 염색체 역위의 중요성은 진화 유전학에서 여전히 열린 경험적 문제로 남아있다.[15]
역위 다형성은 유전 부동, 선택, 유전자 흐름, 균형 선택 등에 의해 확립될 수 있다.[13] 역위 염색체와 조상 염색체 간의 적합성 차이는 안정적인 다형성을 생성하거나, 한 염색체의 고정을 초래할 수 있다.[16]
역위는 종 분화에 여러 방식으로 관여할 수 있는데, 이형접합체 역위는 열등성을 나타낼 수 있으므로, 잡종 적합성 손실을 유발하여 접합 후 격리를 초래할 수 있다. 또한 종 간의 선택된 차이를 축적하여 접합 전 및 접합 후 격리를 모두 유발할 수 있다.[16]
4. 3. 역위 다형성
역위 이형접합체 사이의 재조합 억제는 조상 배열과 역위 배열이 독립적으로 진화할 수 있게 한다. 처음에는 역위 배열에 변이가 없지만, 조상 배열에는 변이가 존재한다. 만약 역위 반수체형이 부동과 같은 이유로 소실되지 않으면, 역위 배열의 변이는 시간이 지남에 따라 증가하며, 더 많은 동형 접합체가 도입되면서 역위 영역의 재조합률이 어느 정도 회복될 수 있다.[13]염색체 역위는 국소 적응 및 종 분화에 잠재적인 역할을 하기 때문에 진화 연구에서 많은 주목을 받고 있다. 비재조합 역위 반수체형은 여러 공동 적응 유전자 변이를 포함할 수 있으므로, 역위는 다른 환경에 대한 국소 적응을 촉진하는 것으로 여겨진다. 이는 자연 선택이 이러한 연결된 적응 변이를 집단 내에서 높은 빈도로 유도하는 데 더 효율적이기 때문이다.[14] 그러나 역위 반수체형이 재조합되지 않기 때문에, 역위 내에서 연결된 공동 적응 유전자 변이의 존재를 경험적으로 입증하는 것은 어렵다. 또한, 다른 환경에 대한 적응에 대한 염색체 역위의 긍정적인 효과는, 별개의 역위 반수체형으로 연결된 적응 유전자 변이가 실제로 공동 적응한다는 가정에 기반한다. 그러나 이 아이디어는 집단이 공간적으로 또는 시간적으로 변동하는 선택을 경험하는 상황에서 위배될 가능성이 높다. 역위 연결 변이에 대한 변동 선택으로 인해, 별개의 유전자 변이를 포함하는 역위 반수체형 간의 재조합이 없으면, 별개의 환경에 대한 적응을 돕기보다는 제한할 수 있다.[15] 따라서, 다른 환경에 대한 적응에서 염색체 역위의 중요성은 진화 유전학에서 여전히 해결해야 할 경험적 문제로 남아있다.
역위 다형성은 두 가지 방법으로 확립될 수 있다. 유전 부동 또는 선택은 국소 집단에서 역위 고정을 초래할 수 있다. 역위 다형성은 이 집단과 역위가 없는 집단 간의 유전자 흐름의 결과로 나타날 수 있다. 균형 선택은 빈도 의존성 또는 과우성에 의해 역위 다형성을 초래할 수도 있다.[13] 역위 염색체와 조상 염색체 간의 적합성 차이는 안정적인 다형성을 생성하거나, 한 염색체의 고정을 초래할 수 있다.[16]
역위는 성염색체 진화에 필수적이었다. 포유류에서 Y 염색체는 거의 전체 길이에 걸쳐 X 염색체와 재조합할 수 없다. 이 비재조합 부분은 일련의 역위가 중첩되어 나타난다. 성 결정 좌위와 성 반대 유전자 간의 재조합률 감소는 선택에 의해 선호된다. 이는 수컷 결정 좌위와 수컷에게 유익한 다른 좌위의 대립 유전자 간의 연관 불균형을 유발한다. 이는 포유류 Y 염색체의 경우처럼, 두 좌위를 모두 포함하는 비재조합 블록을 초래하는 역위를 통해 발생할 수 있다.[16]
역위는 또한 새로운 성염색체의 기원에 필수적일 수 있다. 역위는 성 결정 돌연변이와 성 반대 좌위 간의 연관 불균형을 유발하고, 상염색체로부터 새로운 성염색체를 만들 수 있다.[17]
역위는 여러 가지 방식으로 종 분화에 관여할 수 있다. 이형접합체 역위는 열등성을 나타낼 수 있으므로, 잡종 적합성 손실을 유발하여 접합 후 격리를 초래할 수 있다. 또한 종 간의 선택된 차이를 축적하여 접합 전 및 접합 후 격리를 모두 유발할 수 있다.[16]
역위는 종종 지리적 빈도 경사를 형성하며, 이는 국소 적응에서의 역할을 암시할 수 있다. 이러한 경사의 두드러진 예는 세 대륙에서 관찰될 수 있는 ''초파리 수컷''의 역위 3RP이다.[16] 역위가 두 개 이상의 국소 적응 대립 유전자를 포함하는 경우, 선택되어 확산될 수 있다. 예를 들어, 나비 ''헬리코니우스 누마''에서 색상을 제어하는 18개의 유전자가 역위에 의해 함께 연결되어, 함께 더 높은 적합성을 부여한다.[18]
4. 4. 성염색체 진화
염색체 역위는 성염색체 진화에 필수적인 역할을 한다. 포유류에서 Y 염색체는 거의 전체 길이에 걸쳐 X 염색체와 재조합할 수 없다. 이러한 비재조합 부분은 일련의 역위가 중첩되어 나타난다. 성 결정 좌위와 성 반대 유전자 간의 재조합률 감소는 선택에 의해 선호된다. 이는 수컷 결정 좌위와 수컷에게 유익한 다른 대립 유전자 간의 연관 불균형을 유발하며, 포유류 Y 염색체의 경우처럼, 두 좌위를 모두 포함하는 비재조합 블록을 초래하는 역위를 통해 발생할 수 있다.[16]역위는 새로운 성염색체의 기원에도 필수적일 수 있다. 역위는 성 결정 돌연변이와 성 반대 좌위 간의 연관 불균형을 유발하고, 상염색체로부터 새로운 성염색체를 만들 수 있다.[17]
4. 5. 종 분화 기여
알프레드 스터티번트가 1921년에 초파리에서 염색체 역위의 최초 증거를 발견한 이후,[9] 모든 진핵생물에서 역위가 발견되었다.[3] 염색체 역위는 국소 적응 및 종 분화에 잠재적인 역할을 하기 때문에 진화 연구에서 많은 주목을 받고 있다.역위는 여러 가지 방식으로 종 분화에 관여할 수 있다. 이형접합체 역위는 열등성을 나타낼 수 있으므로, 잡종 적합성 손실을 유발하여 접합 후 격리를 초래할 수 있다. 또한 종 간의 선택된 차이를 축적하여 접합 전 및 접합 후 격리를 모두 유발할 수 있다.[16] 역위는 종종 지리적 빈도 경사를 형성하며, 이는 국소 적응에서의 역할을 암시할 수 있다. 예를 들어, 나비 ''헬리코니우스 누마''에서 색상을 제어하는 18개의 유전자가 역위에 의해 함께 연결되어, 함께 더 높은 적합성을 부여한다.[18]
키르크패트릭-바튼 모델(2006)에 따르면, 역위가 적응적 대립 유전자를 함께 연결함으로써 선택적으로 유리하다. 여러 유전자에서 공진화된 변이를 역위의 개별 버전(haplotype)으로 물리적으로 연결함으로써, 선택은 이러한 변이를 집단 내에서 높은 빈도로 이끄는 데 더 효율적이다. 비재조합 역위 반수체형은 여러 공동 적응 유전자 변이를 포함할 수 있으므로, 역위는 다른 환경에 대한 국소 적응을 촉진하는 것으로 생각된다.[11][14]
역위 이형접합체 사이의 재조합 억제는 조상 배열과 역위 배열의 독립적인 진화를 위한 기회를 제공한다. 역위 반수체형이 부동으로 인해 손실되지 않으면, 역위 배열의 변이는 시간이 지남에 따라 증가할 수 있으며, 더 많은 동형 접합체가 도입됨에 따라 역위 영역의 재조합률이 다소 회복된다.[13]
역위 다형성은 유전 부동 또는 선택이 국소 집단에서 역위의 고정을 초래하고, 이 집단과 역위가 없는 집단 간의 유전자 흐름의 결과로 나타날 수 있다. 균형 선택은 빈도 의존성 또는 과우성에 의해 역위 다형성을 초래할 수도 있다.[13] 역위 염색체와 조상 염색체 간의 적합성 차이는 안정적인 다형성을 생성하거나, 한 염색체의 고정을 초래할 수 있다.[16]
역위는 성염색체 진화에 필수적이었다. 포유류에서 Y 염색체는 거의 전체 길이에 걸쳐 X 염색체와 재조합할 수 없다. 이 비재조합 부분은 일련의 역위가 중첩되어 나타난다. 성 결정 좌위와 성 반대 유전자 간의 재조합률 감소는 선택에 의해 선호된다. 이는 수컷 결정 좌위와 수컷에게 유익한 다른 좌위의 대립 유전자 간의 연관 불균형을 유발한다. 이는 포유류 Y 염색체의 경우처럼, 두 좌위를 모두 포함하는 비재조합 블록을 초래하는 역위를 통해 발생할 수 있다.[16] 역위는 성 결정 돌연변이와 성 반대 좌위 간의 연관 불균형을 유발하고, 상염색체로부터 새로운 성염색체를 만들 수 있다.[17]
5. 임상적 의의
염색체 역위는 대부분 정상 표현형을 나타내지만, 생식 세포의 감수분열 과정에서 문제가 발생하면 자녀에게 울프-허쉬호른 증후군과 같은 유전 질환을 유발할 수 있다.[22] 부모가 역위 보인자일 경우 자식 세대에서 염색체 이상이 발생할 확률이 높아지지만, Inv(9), Inv(2)와 같이 임상 소견이 없는 정상 변이형도 존재한다.[21] 염색체 역위가 발생해도 대부분 정상 표현형을 보이는 이유는 염색체(하드디스크) 외형이 변형되어도 유전자(저장된 정보)가 파괴되지 않으면 기능을 유지할 수 있기 때문이다.
5. 1. 유전 질환
염색체 역위는 균형 재배열을 나타내는 염색체의 구조적 이상 중 하나로, 대부분 정상 표현형을 나타낸다. 그러나 생식 세포의 감수분열 단계에서 역위 고리를 만들면 염색체의 결실 또는 중복을 보이는 재조합 염색체가 형성되어 자녀에게서 비정상 표현형이 나타날 수 있다. 4번 염색체의 구조적 이상에 동반된 4p 결실은 울프-허쉬호른 증후군을 초래할 수 있다.[22]부모가 역위 보인자일 경우 자식 세대에 염색체 이상이 일어날 확률이 매우 높아진다. 역위 보인자가 임신할 확률이나 아이에게 위험이 나타날 확률은 역위 부위와 역위된 부분의 크기에 따라 달라진다. 예를 들어 Inv(9)와 Inv(2)와 같은 역위형은 많은 사람들이 갖고 있지만 임상 소견이 없어 정상 변이형으로 판단하며, 임신이나 태아에 아무런 영향을 주지 않는다.[21]
염색체 구조적 이상에도 불구하고 대부분 정상 표현형을 나타낼 수 있는 이유는 염색체가 곧 유전자는 아니기 때문이다. 이는 컴퓨터 하드디스크(염색체)가 외관상 변형되었더라도 저장된 정보(유전자)가 파괴되지 않았다면 그 정보를 문제없이 활용할 수 있는 것과 같다.
5. 2. 보인자
염색체 역위는 균형 재배열을 나타내는 염색체의 구조적 이상 중 하나로, 대부분 정상 표현형을 나타낸다. 그러나 생식 세포의 감수분열 단계에서 역위 고리를 만들면 염색체의 결실 또는 중복을 보이는 재조합 염색체가 형성되어 자녀에게서 비정상 표현형이 나타날 수 있다. 4번 염색체의 구조적 이상에 동반된 4p 결실은 울프-허쉬호른 증후군을 초래할 수 있다.[22]부모가 역위 보인자일 경우 자식 세대에 염색체 이상이 일어날 확률이 매우 높아진다. 역위 보인자가 임신할 확률이나 아이에게 위험이 나타날 확률은 역위 부위가 어디인지, 얼마나 많은 부분이 역위되었는지에 따라 달라진다. 예를 들어 Inv(9)와 Inv(2)와 같은 역위형은 많은 사람들이 갖고 있지만 임상 소견이 없어 정상 변이형으로 판단하며, 임신이나 태아에 아무런 영향을 주지 않는다.[21]
역위를 포함하는 염색체의 구조적 이상에도 대부분 정상 표현형을 나타낼 수 있는 이유는 염색체가 곧 유전자는 아니기 때문이다. 이는 컴퓨터 하드디스크(염색체)가 외관상 변형되었어도 저장된 정보(유전자)가 파괴되지 않았다면 그 정보를 문제없이 활용할 수 있는 경우와 같다.
5. 3. 유전 상담
염색체 역위는 균형 재배열을 나타내는 염색체의 구조적 이상 중 하나로, 대부분 정상 표현형을 나타낸다. 그러나 생식 세포의 감수분열 단계에서 역위 고리를 만들면 염색체의 결실 또는 중복을 보이는 재조합 염색체가 형성되어 자녀에게서 비정상 표현형이 나타날 수 있다. 4번 염색체의 구조적 이상에 동반된 4p 결실은 울프-허쉬호른 증후군을 초래할 수 있다.[22]부모가 역위 보인자일 경우 자식 세대에 염색체 이상이 일어날 확률이 매우 높아진다. 한편, 역위 보인자가 임신할 확률이나 아이에게 위험이 나타날 확률은 역위 부위와 역위된 부분의 크기에 따라 달라진다. 예를 들어 Inv(9)와 Inv(2)와 같은 역위형은 많은 사람들이 갖고 있지만, 임상 소견이 없어 정상 변이형으로 판단하며 임신이나 태아에 아무런 영향을 주지 않는다.[21]
6. 탐지
세포유전학 기술 및 유전자 분석을 통해 염색체 역위를 탐지하거나 추론할 수 있다. 그러나 대부분의 종에서 작은 역위는 감지되지 않는다. 최근에는 게놈 매핑을 통한 비교 유전체학이나, 연관 불평형 영역을 활용한 집단 유전체학으로 염색체 역위를 감지한다.[6][7] 역위 보균자 가능성이 있는 가족에게는 유전 상담과 유전자 검사가 제공될 수 있다.[8]
6. 1. 세포유전학 기술
세포유전학 기술은 역위를 감지할 수 있으며, 유전자 분석을 통해 역위를 추론할 수도 있다. 그럼에도 불구하고, 대부분의 종에서 작은 역위는 감지되지 않는다. 더 최근에는, 게놈을 매핑하여 염색체 역위를 감지하기 위해 비교 유전체학이 사용되었다.[6][7] 집단 유전체학은 또한 가능한 역위 부위의 지표로 높은 연관 불평형 영역을 사용하여 역위를 감지하는 데 사용될 수 있다. 역위 보균자일 수 있는 인간 가족에게는 유전 상담과 유전자 검사가 제공될 수 있다.[8]6. 2. 유전체 분석
세포유전학 기술은 역위를 감지할 수 있으며, 유전자 분석을 통해 역위를 추론할 수도 있다. 그럼에도 불구하고, 대부분의 종에서 작은 역위는 감지되지 않는다. 더 최근에는, 게놈을 매핑하여 염색체 역위를 감지하기 위해 비교 유전체학이 사용되었다.[6][7] 집단 유전체학은 또한 가능한 역위 부위의 지표로 높은 연관 불평형 영역을 사용하여 역위를 감지하는 데 사용될 수 있다. 역위 보균자일 수 있는 인간 가족에게는 유전 상담과 유전자 검사가 제공될 수 있다.[8]7. 명명법
염색체가 역위된 부분에 중심절이 포함되면 완간 역위, 그렇지 않으면 완내 역위로 구분된다. 완간 역위의 경우 중심체를 포함하므로 끊어진 부분이 장완과 단완에서 각각 발생하므로 발생부위를 46,XY,inv(3)(p23q27)와 같이 나타낸다. 완내 역위는 장완 혹은 단완 내에서만 절단되므로 46,XY,inv(1)(p12p31)와 같이 나타낸다.[21]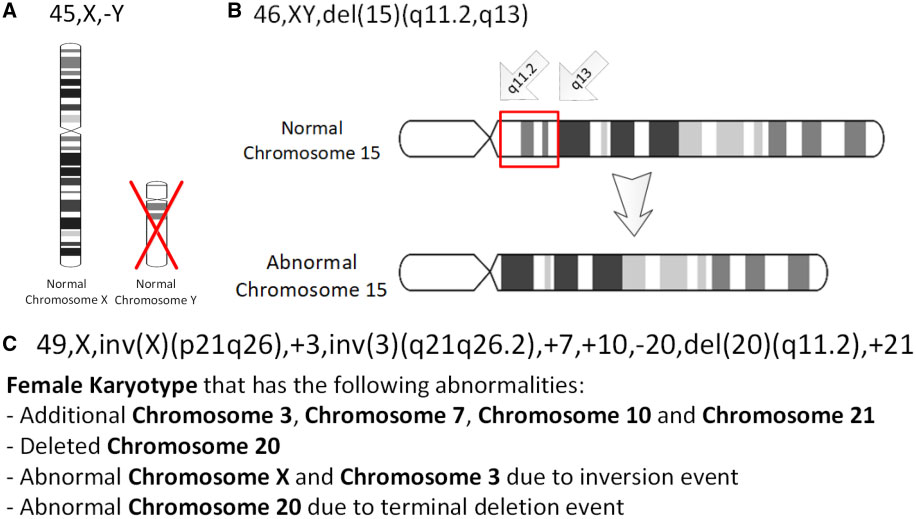
인간 세포유전체 명명법 국제 시스템(ISCN)은 인간 염색체 명명법에 대한 국제 표준으로, 인간 염색체 및 염색체 이상에 대한 설명에 사용되는 밴드 이름, 기호 및 약어를 포함한다. 약어에는 역위를 나타내는 ''inv''가 포함된다.[20]
8. 주목할 만한 사례
브렌던 애덤스는 한때 최장신 십대 기록으로 기네스 세계 기록을 보유했다. 그의 키는 12번 염색체 역위로 인해 발생했다. 곤충 ''Coelopa frigida''에서 염색체 역위의 예를 볼 수 있다. 이 종의 ''Coelopa''는 일련의 신체적 차이를 만들 수 있도록 하는 염색체 역위 변이를 가지고 있다. 더 큰 개체 ''C. frigida''는 염색체 역위를 겪지 않는 반면, 더 작은 개체는 염색체 역위를 겪는다.[1]
참조
[1]
논문
Fine-Mapping Complex Inversion Breakpoints and Investigating Somatic Pairing in the ''Anopheles gambiae'' Species Complex Using Proximity-Ligation Sequencing
2019-12
[2]
논문
Recurrent inversion toggling and great ape genome evolution
2020-08
[3]
논문
Eco-Evolutionary Genomics of Chromosomal Inversions
2018-06
[4]
논문
Frequency, Origins, and Evolutionary Role of Chromosomal Inversions in Plants
2020
[5]
논문
How and why chromosome inversions evolve
2010-09
[6]
논문
Frequency, Origins, and Evolutionary Role of Chromosomal Inversions in Plants
2020
[7]
논문
How and why chromosome inversions evolve
2010-09
[8]
서적
Chromosome Abnormalities and Genetic Counseling
Oxford University Press
[9]
논문
How and why chromosome inversions evolve
2010-09
[10]
논문
Frequency, Origins, and Evolutionary Role of Chromosomal Inversions in Plants
2020
[11]
논문
Frequency, Origins, and Evolutionary Role of Chromosomal Inversions in Plants
2020
[12]
웹사이트
Concepts of Genetics
https://www.pearson.[...]
2022-12-05
[13]
논문
Evolving Inversions
2019-03
[14]
논문
Chromosomal rearrangements and speciation
2001
[15]
논문
Chromosomal inversions can limit adaptation to new environments.
https://doi.org/10.1[...]
2022
[16]
논문
How and why chromosome inversions evolve
2010-09
[17]
논문
Turnover of sex chromosomes induced by sexual conflict
2007-10
[18]
논문
Chromosomal rearrangements maintain a polymorphic supergene controlling butterfly mimicry
2011-08
[19]
논문
A fully computational and reasonable representation for karyotypes
2019-12
[20]
웹사이트
ISCN Symbols and Abbreviated Terms
https://www.coriell.[...]
2022-10-27
[21]
웹사이트
세포유전학적 이상 소견의 생성기전
http://labmed.hallym[...]
한림대학교
2014-10-27
[22]
논문
Recombinant Chromosome 4 with Partial 4p Deletion and 4q Duplication Inherited from Paternal Pericentric Inversion(부모의 유전자 역위로 인해 4번 염색체 재조합에서 일어나는 유전자 중복과 유전자 결실에 대해)
2010
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com