보조 인자는 효소의 활성에 필요한 비단백질성 분자로, 유기 보조 인자와 무기 보조 인자로 분류된다. 유기 보조 인자는 조효소, 보결분자단 등으로 세분화되며, 비타민은 조효소의 전구체 역할을 하거나 조효소 자체로 작용한다. 무기 보조 인자로는 금속 이온과 철-황 클러스터 등이 있으며, 아포효소와 홀로효소의 관계를 통해 효소 활성에 필수적임을 알 수 있다. 보조 인자는 물질대사 과정에서 작용기 전달 중간생성물로 재사용되며, 단백질의 번역 후 변형에 의해 형성되기도 한다. 또한, 비효소 보조 인자는 생물학의 다른 영역에서 단백질의 기능을 조절하며, ATP, NADH와 같은 유기 보조 인자는 생명체의 진화 과정에서 중요한 역할을 해왔다.
더 읽어볼만한 페이지
보조 인자 - 피롤로퀴놀린 퀴논 피롤로퀴놀린 퀴논(PQQ)은 세균에서 발견되는 산화환원 반응 보조 인자로서, 여러 효소 활성에 관여하고 단백질과 결합하여 퀴노프로테인을 형성하며, 항산화 및 신경 보호 작용을 하는 것으로 알려졌지만 포유류 필수 영양소 여부는 불확실하고 코코아, 모유, 낫토 등에 존재한다.
보조 인자 - 필로퀴논 필로퀴논은 비타민 K₁이라고도 불리는 지용성 비타민으로, 동물에서는 혈액 응고와 뼈 형성에, 식물과 시아노박테리아에서는 광합성에 중요한 역할을 하며, 의학적으로는 출혈 장애 치료와 비타민 K 결핍증 예방에 사용된다.
효소 - 활성 부위 활성 부위는 효소 분자 내에서 기질과 결합하여 촉매 작용을 하는 특정 부위이며, 다양한 모델로 설명되고, 결합 부위, 촉매 부위, 보조인자를 포함하며, 저해제와 알로스테릭 부위와 관련되어 신약 개발에 활용된다.
효소 - 제한 효소 제한 효소는 DNA의 특정 염기서열을 인식하여 절단하는 효소로, 유전체학과 생명공학 발전에 기여하여 노벨상을 수상했으며, 인식 서열 특이성 등에 따라 다양한 유형으로 분류되어 여러 연구 및 응용 분야에 활용된다.
헴 (헤모글로빈의 산소 운반 보조 인자) 플라빈 조효소 A 피리독살 인산 금속 이온: 구리, 철, 망간, 몰리브덴, 아연
관련 질병
특정 보조 인자 결핍은 질병 유발 가능 (예: 비타민 결핍증)
2. 정의 및 분류
보조 인자는 효소의 활성에 필요한 비단백질 분자로, 크게 유기 보조 인자와 무기 보조 인자로 나뉜다.
유기 보조 인자는 플라빈, 헴과 같이 탄소를 포함하는 분자이며, 조효소, 보결분자단으로 더 세분화되기도 한다. 조효소는 효소의 기능적 특성을 강조하는 용어이며, 보결분자단은 효소와의 결합 강도(단단하게 또는 공유 결합)와 같은 구조적 특성을 나타내는 용어이다. 하지만 문헌에 따라 이러한 용어들의 정의가 조금씩 다르며, 종종 느슨하게 사용되기도 한다.[109] 예를 들어, 어떤 문헌에서는 단단히 결합된 유기 분자를 조효소가 아닌 보결분자단으로 간주하는 반면, 다른 문헌에서는 효소 활성에 필요한 모든 비단백질 유기 분자를 조효소로 정의하고 단단하게 결합된 것을 조효소 보결분자단으로 분류한다.
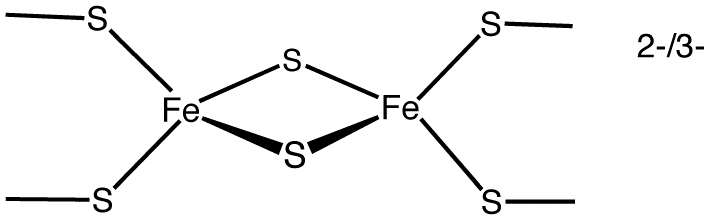
2개의 철 원자와 2개의 황 원자를 포함하는 단순한 [Fe2S2] 클러스터는 4개의 시스테인 잔기에 의해 조정된다.
철-황 클러스터는 시스테인 잔기에 의해 단백질 내에 포함된 철과 황 원자의 복합체로, 전자전달, 산화환원 감지, 구조적 역할 등 다양한 기능을 수행한다.
4. 1. 금속 이온
금속 이온은 일반적인 보조 인자이다.[110] 이러한 보조 인자에 대한 연구는 생물무기화학 영역에 속한다. 영양에서 필수 미량 원소의 목록은 보조 인자로서의 역할을 반영한다. 사람의 경우 이 목록에는 일반적으로 철, 마그네슘, 망가니즈, 코발트, 구리, 아연, 몰리브데넘이 포함된다.[111]
크로뮴 결핍은 내당능장애를 유발하지만, 크로뮴을 보조 인자로 사용하는 사람의 효소는 확인되지 않았다.[112][113]아이오딘은 필수 미량 원소이지만, 효소의 보조 인자가 아닌 갑상샘 호르몬 구조의 일부로 사용된다.[114]칼슘은 사람의 식단에 필요한 구성 요소이며, 산화질소 생성효소, 단백질 인산가수분해효소, 아데닐산 키네이스 등 많은 효소들의 완전한 활성을 위해 필요하지만, 알로스테릭 조절에서 이러한 효소들을 활성화시키고 종종 칼모듈린과의 복합체에서 이들 효소에 결합한다.[115] 따라서 칼슘은 세포 신호전달 분자이며, 일반적으로 칼슘이 조절하는 효소의 보조 인자로 간주되지 않는다.[116]
어떤 생물들은 효소 보조 인자로 추가적인 금속을 필요로 한다. 아조토박터속 질소고정세균의 질소고정효소에 있는 바나듐,[117] 호열성 고세균인 ''Pyrococcus furiosus''의 알데하이드 페레독신 산화환원효소에 있는 텅스텐,[118] 해양 규조류인 ''Thalassiosira weissflogii''의 탄산무수화효소에 있는 카드뮴 등이 여기에 해당한다.[119][120]
많은 경우 보조 인자는 무기 및 유기 성분을 모두 포함한다. 철에 배위된 포르피린 고리로 구성된 헴 단백질이 그러한 예시이다.[121]
철-황 클러스터는 시스테인잔기에 의해 단백질 내에 포함된 철과 황원자의 복합체이다. 철-황 클러스터는 전자전달, 산화환원 감지 및 구조 모듈을 포함한 구조적, 기능적 역할을 모두 수행한다.[122][22]
5. 유기 보조 인자
유기 보조 인자는 효소에 느슨하게 또는 단단하게 결합하여 반응에 직접적으로 참여할 수 있는 작은 유기 분자(일반적으로 1,000 Da 미만의 분자량을 가짐)이다.[106][123][124][125] 효소에 단단히 결합하여 효소를 변성시키지 않고는 제거하기 어려운 유기 보조 인자를 보결분자단이라고 부른다.
여기서, 느슨하게 결합된 보조 인자와 단단하게 결합된 보조 인자 사이에 뚜렷한 구분이 없다는 점을 강조할 필요가 있다.[106] 실제로 NAD+와 같은 많은 보조 인자들은 일부 효소에는 단단히 결합될 수 있지만, 다른 효소에서는 느슨하게 결합될 수 있다.[106] 또 다른 예로 티아민 피로인산(TPP)은 트랜스케톨레이스나 피루브산 탈카복실화효소에는 단단히 결합되어 있지만, 피루브산 탈수소 효소에는 상대적으로 느슨하게 결합되어 있다.[126]플라빈 아데닌 다이뉴클레오타이드(FAD), 비오틴, 리포아마이드와 같은 다른 조효소들은 단단히 결합되어 있다.[127]
일반적으로 단단히 결합된 보조 인자는 동일한 반응 주기 동안 재생되는 반면, 느슨하게 결합된 보조 인자는 다른 효소에 의해 촉매되는 후속 반응에서 재생될 수 있다. 느슨하게 결합된 보조 인자의 경우 기질 또는 보조 기질로 간주될 수도 있다.
비타민은 많은 유기 보조 인자(예: 비타민 B1, 비타민 B2, 비타민 B6, 비타민 B12, 니아신, 폴산) 또는 조효소 그 자체(예: 비타민 C)의 전구체 역할을 할 수 있다. 비타민은 신체에서 다른 기능을 수행할 수도 있다.[128] 많은 유기 보조 인자들은 전자 운반체인 NAD+ 및 FAD와 아실기를 운반하는 조효소 A를 포함하고 있다. 이러한 보조 인자들의 대부분은 매우 다양한 생물종에서 발견되며, 일부 보조 인자들은 모든 생명체에 보편적으로 존재한다. 예외적으로 고세균 그룹으로 제한되는 메테인 생성균에서 진화한 고유한 보조 인자 그룹이 있다.[129]
5. 1. 비타민과 그 유도체
비타민은 많은 유기 보조 인자(예: 비타민 B1, B2, B6, B12, 니아신, 엽산) 또는 조효소 자체(예: 비타민 C)로 작용할 수 있다. 그러나 비타민은 신체 내에서 다른 기능도 수행한다.[28]
비타민과 그 유도체는 다양한 화학 그룹 전달에 관여하며, 생물체 내에서 널리 분포한다. 다음은 주요 비타민 유도체와 그 역할에 대한 표이다.
물질대사는 광범위한 화학 반응을 포함하지만, 대부분은 작용기의 전달을 포함하는 몇 가지 기본적인 반응 유형들로 구분할 수 있다.[158] 이러한 화학 반응들은 세포가 작은 세트의 대사 중간생성물들을 사용하여 서로 다른 반응들 사이에 화학 작용기들을 운반할 수 있도록 한다.[159] 이러한 작용기 전달 중간생성물은 보통 조효소라고 부르는 느슨하게 결합된 유기 보조 인자이다.
작용기 전달 반응의 각 부류는 작용기를 생성하는 효소 세트의 기질인 특정 보조 인자와 이러한 작용기를 소비하는 효소 세트에 의해 수행된다. 이러한 반응의 예는 니코틴아마이드 아데닌 다이뉴클레오타이드(NAD+)를 보조 인자로 사용하는 탈수소효소이다. 수백 가지 유형의 효소가 기질로부터 전자를 NAD+로 전달하여 NADH로 환원시킨다. 이러한 환원된 보조 인자는 기질을 환원시키기 위해 전자를 필요로 하는 세포 내 환원효소의 기질이 된다.[160]
따라서 이러한 보조 인자는 물질대사의 일부로 지속적으로 재사용된다. 예를 들어 인체의 ATP의 총량은 약 0.1몰이다. 이러한 ATP는 계속해서 ADP로 분해되고, 다시 ATP로 합성된다. 따라서 어떤 주어진 시간에서 ATP+ADP의 총량은 상당히 일정하다. 사람이 매일 사용하는 에너지는 100~150몰의 ATP가 가수분해될 때 나오는 에너지이며, 이는 약 50~75 kg에 해당하는 양이다. 일반적인 상황에서 사람은 하루 동안 자신의 체중 만큼의 ATP를 소모한다.[161] 이는 각 ATP 분자가 매일 1,000~1,500회 재활용된다는 것을 의미한다.
6. 단백질 유래 보조 인자
많은 효소에서 보조 인자로 작용하는 부분은 단백질 서열 일부의 번역 후 변형에 의해 형성된다. 이것은 종종 단백질의 기능을 위해 금속 이온과 같은 외부 결합 인자의 필요성을 대체한다. 잠재적인 변형은 방향족 잔기의 산화, 잔기 간의 결합, 절단 또는 고리 형성이 될 수 있다.[178] 이러한 변형은 아미노산이 일반적으로 새로운 기능을 획득한다는 점에서 인산화, 메틸화, 글리코실화와 같은 다른 번역 후 변형과는 다르다. 이것은 단백질의 기능을 증가시킨다. 변형되지 않은 아미노산은 일반적으로 산-염기 반응으로 제한되며, 잔기의 변형은 단백질의 친전자성 부위 또는 자유 라디칼을 안정화시키는 능력을 제공할 수 있다.[178] 보조 인자 생성의 예로는 두 개의 트립토판곁사슬에서 유래된 트립토판 트립토필퀴논(TTQ)[179]과 Ala-Ser-Gly 모티프에서 유래된 4-메틸리덴-이미다졸-5-온(MIO)이 있다.[180] 단백질 유래 보조 인자의 특성화는 X선 결정학 및 질량 분석법을 사용하여 수행된다. 시퀀싱은 변경된 부위를 쉽게 식별하지 못하기 때문에 구조 데이터가 필요하다.
7. 비효소 보조 인자
비효소 보조 인자는 생물학에서 단백질 기능을 활성화, 불활성화하거나, 단백질 기능에 필요한 비단백질(또는 단백질) 분자를 광범위하게 지칭하는 용어이다. 예를 들어 수용체 단백질에 결합하여 활성화시키는 호르몬과 같은 리간드는 보조 인자 또는 보조활성인자(coactivator)라고 하는 반면, 수용체 단백질을 억제하는 분자는 보조억제인자(corepressor)라고 한다.[181] 그러한 예 중 하나는 감각 뉴런에서 자주 발견되는 수용체의 G 단백질 연결 수용체 패밀리이다. 수용체에 결합하는 리간드는 G 단백질을 활성화한 다음 효소를 활성화시켜 효과인자를 활성화시킨다.[181] 혼동을 피하기 위해 리간드 결합 매개 활성화 또는 억제하는 단백질을 공동조절인자(coregulator)로 하자는 것이 제안되었다.[182]
8. 진화
ATP 및 NADH와 같은 유기 보조 인자는 알려진 모든 형태의 생명체에 존재하며 물질대사의 핵심적인 부분을 담당한다. 이러한 보편성은 이들 분자가 생명 역사의 초기부터 진화해 왔다는 것을 나타낸다.[162] 따라서 현존하는 보조 인자들 중 일부는 약 38억년 전에 출현한 최초의 생명체에 존재했을 수도 있다.[163][164]
유기 보조 인자는 지구 상의 생명체의 역사에서 훨씬 더 일찍 존재했을 수도 있다.[165]아데노신의 뉴클레오타이드는 메틸기, 아실기, 포스포릴기 전달과 같은 기본적인 대사 반응과 산화환원반응을 촉매하는 보조 인자에 존재한다. 따라서 이러한 보편적인 화학적 골격은 RNA 월드의 잔재로 제안되었으며, 초기 리보자임은 제한된 세트의 뉴클레오타이드 및 관련 화합물에 결합하도록 진화해 왔다.[166][167] 아데노신 기반의 보조 인자는 원래 다른 보조 인자에 결합하도록 진화했던 기존의 아데노신 결합 도메인의 작은 변형을 통해 효소와 리보자임이 새로운 보조 인자에 결합할 수 있도록 하는 상호교환 가능한 어댑터 역할을 한 것으로 생각된다.[168] 새로운 사용을 위해 미리 진화된 구조를 적응시키는 과정을 굴절적응(exaptation)이라고 한다.
효소가 조효소를 필요로 한다면, 조효소는 어떻게 진화했을까? 가장 가능성이 높은 시나리오는 효소가 처음에 조효소 없이 기능을 수행하고 나중에 조효소를 획득했다는 것이다. 비록 촉매 반응이 효율적이거나 빠르지 않더라도 말이다. 예로는 알코올 탈수소 효소 (조효소: NAD⁺),[73] 젖산 탈수소 효소 (NAD⁺),[74] 글루타티온 환원 효소 (NADPH)가 있다.[75]
컴퓨터 프로그램인 IPRO는 최근 ''Candida boidinii''의 자일로스 환원효소의 보조 인자인 NADPH에 대한 특이성을 NADH로 바꾸는 돌연변이를 예측했다.[169]
9. 역사
최초로 발견된 유기 보조 인자는 1906년 아서 하든과 윌리엄 영이 확인한 NAD+이다.[170] 이들은 가열하지 않은 효모 추출물에 가열한 효모 추출물을 첨가하면 알코올 발효가 크게 가속화된다는 사실을 발견하고, 가열해도 파괴되지 않는 미확인 인자를 "coferment"라고 불렀다. 한스 폰 오일러켈핀은 효모 추출물을 어렵게 정제하여 이러한 열안정 인자가 당뉴클레오타이드 인산염이라는 것을 확인했다.[171]
1929년에 칼 로만(Karl Lohmann)은 ATP를 분리했고,[172] 1945년에는 프리츠 앨버트 리프먼이 조효소 A를 발견했다.[173]
1936년 오토 하인리히 바르부르크는 수소화물 전달에서 NAD+의 기능을 확인했다.[174] 1940년대 초, 헤르만 칼카르(Herman Kalckar)는 당의 산화와 ATP 생성 사이의 연관성을 확립했으며,[175] 이는 1941년 프리츠 앨버트 리프먼이 제안한 에너지 전달에서 ATP의 중심적인 역할을 확인시켜 주었다.[176] 이후 1949년 모리스 프리드킨(Morris Friedkin)과 앨버트 레스터 레닌저는 NAD+가 시트르산 회로 및 ATP 합성과 같은 대사 경로들을 연결한다는 것을 증명했다.[177]
참조
[1]
논문
Coenzyme, Cofactor and Prosthetic Group — Ambiguous Biochemical Jargon
http://eprints.um.ed[...] [2]
웹사이트
Coenzymes and cofactors
http://academic.broo[...]
2007-11-17
[3]
웹사이트
Enzyme Cofactors
http://www.elmhurst.[...]
2007-11-17
[4]
서적
Lehninger Principles of Biochemistry
W.H. Freeman and Company
[5]
서적
Biochemistry: the chemical reactions of living cells
https://archive.org/[...]
Harcourt/Academic Press
[6]
서적
Glossary of Terms Used in Bioinorganic Chemistry
http://publications.[...]
Pure & Applied Chemistry
[7]
서적
Thiamine: catalytic mechanisms in normal and disease states
Marcel Dekker
[8]
뉴스
Pyruvate Dehydrogenase Complex
https://chem.librete[...]
2013-10-02
[9]
논문
SAM – semantics and misunderstandings
1979-03
[10]
웹사이트
Biochemistry: Enzymes: Classification and catalysis (Cofactors)
http://vle.du.ac.in/[...]
2018-02-07
[11]
논문
Physiology and metabolism of essential trace elements: an outline
1985-08
[12]
논문
Is chromium a trace essential metal?
[13]
논문
The biochemistry of chromium
2000-04
[14]
논문
Iodine metabolism and thyroid physiology: current concepts
1997-04
[15]
논문
Calcium signaling
[16]
논문
Ca2+ signaling and intracellular Ca2+ binding proteins
1996-10
[17]
논문
The vanadium-containing nitrogenase of Azotobacter
1988-07
[18]
논문
Structure of a hyperthermophilic tungstopterin enzyme, aldehyde ferredoxin oxidoreductase
1995-03
[19]
논문
A biological function for cadmium in marine diatoms
2000-04
[20]
논문
Biochemistry: a cadmium enzyme from a marine diatom
[21]
논문
Structural analysis of heme proteins: implications for design and prediction
2011-03
[22]
논문
Iron-sulfur protein folds, iron-sulfur chemistry, and evolution
2008-02
[23]
서적
Understanding enzymes
https://archive.org/[...]
Horwood
[24]
서적
Lehninger principles of biochemistry
https://archive.org/[...]
Worth Publishers
[25]
서적
Biochemistry
Brooks Cole
[26]
논문
Studies on the nature of the binding of thiamine pyrophosphate to enzymes
http://www.jbc.org/c[...]
1968-06
[27]
논문
Conservation of the Enzyme–Coenzyme Interfaces in FAD and NADP Binding Adrenodoxin Reductase-A Ubiquitous Enzyme
2017-12
[28]
논문
Vitamins: not just for enzymes
[29]
논문
Novel biochemistry of methanogenesis
1988-06
[30]
논문
Structure, mechanism and catalytic duality of thiamine-dependent enzymes
[31]
논문
Pyridoxal phosphate enzymes: mechanistic, structural, and evolutionary considerations
[32]
논문
The many faces of vitamin B12: catalysis by cobalamin-dependent enzymes
https://digitalcommo[...] [33]
논문
The biotin enzyme family: conserved structural motifs and domain rearrangements
[34]
논문
Coenzyme A: back in action
[35]
논문
Folic acid
2001-06
[36]
논문
Microbial ubiquinones: multiple roles in respiration, gene regulation and oxidative stress management
http://mic.sgmjourna[...]
1999-08
[37]
논문
Vitamin C. Biosynthesis, recycling and degradation in mammals
[38]
논문
Flavoenzymes
[39]
논문
Riboflavin analogs and inhibitors of riboflavin biosynthesis
[40]
서적
An introduction to enzyme and coenzyme chemistry
https://archive.org/[...]
Blackwell Science
[41]
논문
S-Adenosylmethionine and methylation
1996-03
[42]
논문
Structure of component B (7-mercaptoheptanoylthreonine phosphate) of the methylcoenzyme M methylreductase system of Methanobacterium thermoautotrophicum
1986-06
[43]
논문
Structure and methylation of coenzyme M(HSCH2CH2SO3)
1974-08
[44]
논문
Specificity and biological distribution of coenzyme M (2-mercaptoethanesulfonic acid)
1979-01
[45]
논문
Biochemical functions of coenzyme Q10
http://www.jacn.org/[...]
2001-12
[46]
서적
Biochemistry & molecular biology of plants
https://archive.org/[...]
American society of plant physiology
[47]
서적
Significance of glutathione in plant adaptation to the environment
https://books.google[...]
Springer
[48]
논문
Glutathione
[49]
논문
Biology of heme in health and disease
[50]
논문
The active species of 'CO2' utilized by formylmethanofuran dehydrogenase from methanogenic Archaea
1997-09
[51]
논문
Molybdoenzymes and molybdenum cofactor in plants
2002-08
[52]
논문
Cell biology of molybdenum
[53]
논문
Comparative biochemistry of nucleotide-linked sugars
[54]
논문
Structure and function of sulfotransferases
https://zenodo.org/r[...]
2001-06
[55]
논문
A novel coenzyme from bacterial primary alcohol dehydrogenases
1979-08
[56]
논문
Tetrahydrobiopterin biosynthesis, regeneration and functions
2000-04
[57]
논문
Unusual coenzymes of methanogenesis
[58]
논문
The Ninth Sir Hans Krebs Lecture. Compartmentation and communication in living systems. Ligand conduction: a general catalytic principle in chemical, osmotic and chemiosmotic reaction systems
1979-03
[59]
논문
Mechanisms of enzyme-catalyzed group transfer reactions
[60]
논문
The power to reduce: pyridine nucleotides—small molecules with a multitude of functions
[61]
논문
Estimating ATP resynthesis during a marathon run: a method to introduce metabolism
http://advan.physiol[...] [62]
논문
Ribozyme catalysis of metabolism in the RNA world
[63]
서적
How did bacteria come to be?
[64]
논문
The emergence of major cellular processes in evolution
1996-07
[65]
논문
Coenzymes as fossils of an earlier metabolic state
1976-03
[66]
논문
The tyranny of adenosine recognition among RNA aptamers to coenzyme A
[67]
논문
Coenzymes as coribozymes
[68]
논문
Adenine recognition: a motif present in ATP-, CoA-, NAD-, NADP-, and FAD-dependent proteins
2001-08
[69]
논문
Prebiotic synthesis of propiolaldehyde and nicotinamide
https://pubmed.ncbi.[...]
1970-09-25
[70]
논문
A possible prebiotic synthesis of pantetheine, a precursor to coenzyme A
https://pubmed.ncbi.[...]
1995-02-23
[71]
논문
Life's utilization of B vitamins on early Earth
https://onlinelibrar[...]
2017-01
[72]
논문
Computational design of Candida boidinii xylose reductase for altered cofactor specificity
2009-10
[73]
서적
Comprehensive Enzyme Kinetics
https://link.springe[...]
2004
[74]
서적
Biochemistry
Brooks/Cole, Cengage Learning
2010
[75]
간행물
"[59] Glutathione reductase"
https://linkinghub.e[...]
Elsevier
2024-09-21
[76]
논문
The Alcoholic Ferment of Yeast-Juice
1906-10-24
[77]
웹사이트
Fermentation of sugars and fermentative enzymes: Nobel Lecture, May 23, 1930
http://nobelprize.or[...]
Nobel Foundation
2007-09-30
[78]
논문
Über die Pyrophosphatfraktion im Muskel
1929-08
[79]
논문
Acetylation of sulfanilamide by liver homogenates and extracts
1945-09-01
[80]
논문
Pyridin, the hydrogen-transferring component of the fermentation enzymes (pyridine nucleotide)
[81]
논문
Origins of the concept oxidative phosphorylation
1974-11
[82]
서적
A Source Book in Chemistry, 1900-1950
[83]
논문
Esterification of inorganic phosphate coupled to electron transport between dihydrodiphosphopyridine nucleotide and oxygen
http://www.jbc.org/c[...] [84]
논문
Protein-Derived Cofactors. Expanding the Scope of Post-Translational Modifications†
[85]
논문
Posttranslational biosynthesis of the protein-derived cofactor tryptophan tryptophylquinone
[86]
논문
A new member of the 4-methylideneimidazole-5-one-containing aminomutase family from the enediyne kedarcidin biosynthetic pathway
2013-05
[87]
서적
Molecular Cell Biology
https://www.ncbi.nlm[...]
2000-01-01
[88]
논문
Coactivators and corepressors: what's in a name?
2008-10
[89]
웹사이트
生物無機化学で使われる用語集(IUPAC推奨 1997): 補因子
http://www.chem.qmul[...]
International Union of Pure and Applied Chemistry
1997
[90]
웹사이트
coenzymes and cofactors
http://academic.broo[...]
2007-11-17
[91]
웹사이트
Enzyme Cofactors
http://www.elmhurst.[...]
2007-11-17
[92]
논문
Physiology and metabolism of essential trace elements: an outline
[93]
논문
Is chromium a trace essential metal?
[94]
논문
The biochemistry of chromium
http://jn.nutrition.[...] [95]
논문
Iodine metabolism and thyroid physiology: current concepts
[96]
논문
Calcium signaling
[97]
논문
Ca2+ signaling and intracellular Ca2+ binding proteins
[98]
논문
The vanadium-containing nitrogenase of Azotobacter
[99]
논문
Structure of a hyperthermophilic tungstopterin enzyme, aldehyde ferredoxin oxidoreductase
[100]
논문
A biological function for cadmium in marine diatoms
http://www.pnas.org/[...] [101]
논문
Biochemistry: a cadmium enzyme from a marine diatom
[102]
서적
Coenzyme, Cofactor and Prosthetic Group – Ambiguous Biochemical Jargon
Biochemical Education
[103]
웹인용
coenzymes and cofactors
http://academic.broo[...]
2007-11-17
[104]
웹인용
Enzyme Cofactors
http://www.elmhurst.[...]
2007-11-17
[105]
서적
Lehninger Principles of Biochemistry
https://archive.org/[...]
W.H. Freeman and Company
[106]
서적
Biochemistry: the chemical reactions of living cells
https://archive.org/[...]
Harcourt/Academic Press
[107]
서적
Thiamine: catalytic mechanisms in normal and disease states
https://archive.org/[...]
Marcel Dekker
[108]
뉴스
Pyruvate Dehydrogenase Complex
https://chem.librete[...]
2013-10-02
[109]
저널
SAM – semantics and misunderstandings
1979-03
[110]
웹인용
Biochemistry: Enzymes: Classification and catalysis (Cofactors)
http://vle.du.ac.in/[...]
2018-02-07
[111]
저널
Physiology and metabolism of essential trace elements: an outline
1985-08
[112]
저널
Is chromium a trace essential metal?
[113]
저널
The biochemistry of chromium
2000-04
[114]
저널
Iodine metabolism and thyroid physiology: current concepts
1997-04
[115]
저널
Calcium signaling
[116]
저널
Ca2+ signaling and intracellular Ca2+ binding proteins
1996-10
[117]
저널
The vanadium-containing nitrogenase of Azotobacter
1988-07
[118]
저널
Structure of a hyperthermophilic tungstopterin enzyme, aldehyde ferredoxin oxidoreductase
1995-03
[119]
저널
A biological function for cadmium in marine diatoms
2000-04
[120]
저널
Biochemistry: a cadmium enzyme from a marine diatom
[121]
저널
Structural analysis of heme proteins: implications for design and prediction
2011-03
[122]
저널
Iron-sulfur protein folds, iron-sulfur chemistry, and evolution
2008-02
[123]
서적
Understanding enzymes
https://archive.org/[...]
Horwood
[124]
서적
Lehninger principles of biochemistry
https://archive.org/[...]
Worth Publishers
[125]
서적
Biochemistry
Brooks Cole
[126]
저널
Studies on the nature of the binding of thiamine pyrophosphate to enzymes
http://www.jbc.org/c[...]
1968-06
[127]
저널
Conservation of the Enzyme–Coenzyme Interfaces in FAD and NADP Binding Adrenodoxin Reductase-A Ubiquitous Enzyme
2017-12
[128]
저널
Vitamins: not just for enzymes
[129]
저널
Novel biochemistry of methanogenesis
http://www.jbc.org/c[...]
2021-04-15
[130]
저널
Structure, mechanism and catalytic duality of thiamine-dependent enzymes
[131]
저널
Pyridoxal phosphate enzymes: mechanistic, structural, and evolutionary considerations
[132]
저널
The many faces of vitamin B12: catalysis by cobalamin-dependent enzymes
https://digitalcommo[...] [133]
저널
The biotin enzyme family: conserved structural motifs and domain rearrangements
[134]
저널
Coenzyme A: back in action
[135]
저널
Folic acid
2001-06
[136]
저널
Microbial ubiquinones: multiple roles in respiration, gene regulation and oxidative stress management
http://mic.sgmjourna[...]
2021-04-15
[137]
저널
Vitamin C. Biosynthesis, recycling and degradation in mammals
[138]
저널
Flavoenzymes
[139]
저널
Riboflavin analogs and inhibitors of riboflavin biosynthesis
[140]
서적
An introduction to enzyme and coenzyme chemistry
https://archive.org/[...]
Blackwell Science
[141]
저널
S-Adenosylmethionine and methylation
1996-03
[142]
저널
Structure of component B (7-mercaptoheptanoylthreonine phosphate) of the methylcoenzyme M methylreductase system of Methanobacterium thermoautotrophicum
1986-06
[143]
저널
Structure and methylation of coenzyme M(HSCH2CH2SO3)
http://www.jbc.org/c[...]
2021-04-15
[144]
저널
Specificity and biological distribution of coenzyme M (2-mercaptoethanesulfonic acid)
1979-01
[145]
저널
Biochemical functions of coenzyme Q10
http://www.jacn.org/[...]
2001-12
[146]
서적
Biochemistry & molecular biology of plants
https://archive.org/[...]
American society of plant physiology
[147]
서적
Significance of glutathione in plant adaptation to the environment
https://books.google[...]
Springer
[148]
저널
Glutathione
[149]
저널
Biology of heme in health and disease
[150]
저널
The active species of 'CO2' utilized by formylmethanofuran dehydrogenase from methanogenic Archaea
1997-09
[151]
저널
Molybdoenzymes and molybdenum cofactor in plants
http://jxb.oxfordjou[...]
2002-08
[152]
저널
Cell biology of molybdenum
[153]
저널
Comparative biochemistry of nucleotide-linked sugars
[154]
저널
Structure and function of sulfotransferases
https://zenodo.org/r[...]
2001-06
[155]
저널
A novel coenzyme from bacterial primary alcohol dehydrogenases
1979-08
[156]
저널
Tetrahydrobiopterin biosynthesis, regeneration and functions
2000-04
[157]
저널
Unusual coenzymes of methanogenesis
[158]
저널
The Ninth Sir Hans Krebs Lecture. Compartmentation and communication in living systems. Ligand conduction: a general catalytic principle in chemical, osmotic and chemiosmotic reaction systems
1979-03
[159]
저널
Mechanisms of enzyme-catalyzed group transfer reactions
[160]
저널
The power to reduce: pyridine nucleotides—small molecules with a multitude of functions
[161]
저널
Estimating ATP resynthesis during a marathon run: a method to introduce metabolism
http://advan.physiol[...]
2021-04-15
[162]
저널
Ribozyme catalysis of metabolism in the RNA world
[163]
서적
How did bacteria come to be?
[164]
저널
The emergence of major cellular processes in evolution
1996-07
[165]
저널
Coenzymes as fossils of an earlier metabolic state
1976-03
[166]
저널
The tyranny of adenosine recognition among RNA aptamers to coenzyme A
[167]
저널
Coenzymes as coribozymes
[168]
저널
Adenine recognition: a motif present in ATP-, CoA-, NAD-, NADP-, and FAD-dependent proteins
2001-08
[169]
저널
Computational design of Candida boidinii xylose reductase for altered cofactor specificity
2009-10
[170]
저널
The Alcoholic Ferment of Yeast-Juice
1906-10-24
[171]
웹인용
Fermentation of sugars and fermentative enzymes: Nobel Lecture, May 23, 1930
http://nobelprize.or[...]
Nobel Foundation
2007-09-30
[172]
저널
Über die Pyrophosphatfraktion im Muskel
1929-08
[173]
저널
Acetylation of sulfanilamide by liver homogenates and extracts
http://www.jbc.org/c[...]
2021-04-15
[174]
저널
Pyridin, the hydrogen-transferring component of the fermentation enzymes (pyridine nucleotide)
[175]
저널
Origins of the concept oxidative phosphorylation
1974-11
[176]
서적
A Source Book in Chemistry, 1900-1950
[177]
저널
Esterification of inorganic phosphate coupled to electron transport between dihydrodiphosphopyridine nucleotide and oxygen
http://www.jbc.org/c[...] [178]
저널
Protein-Derived Cofactors. Expanding the Scope of Post-Translational Modifications†
[179]
저널
Posttranslational biosynthesis of the protein-derived cofactor tryptophan tryptophylquinone
[180]
저널
A new member of the 4-methylideneimidazole-5-one-containing aminomutase family from the enediyne kedarcidin biosynthetic pathway
2013-05
[181]
저널
G Protein –Coupled Receptors and Their Effectors
https://www.ncbi.nlm[...]
2000-01-01
[182]
저널
Coactivators and corepressors: what's in a name?
2008-10
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.