오르토믹소바이러스과
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
오르토믹소바이러스과는 RNA 바이러스를 포함하는 분류군으로, 인플루엔자 바이러스를 포함한다. 이 과에는 인플루엔자 A, B, C, D 바이러스와 연어빈혈증 바이러스, 토고토바이러스, 콰란자바이러스 등이 속한다. 인플루엔자 바이러스는 구형 또는 실 모양의 외피를 가지며, 헤마글루티닌(HA)과 뉴라미니다제(NA) 단백질을 통해 숙주 세포에 결합한다. 바이러스는 6~8개의 RNA 세그먼트를 포함하며, 캡 스내칭 과정을 통해 숙주 세포의 mRNA를 이용하여 복제한다. 인플루엔자 A 바이러스는 다양한 숙주에 감염되며, 변이를 통해 유행을 일으킬 수 있다. B형은 사람, C형은 사람과 돼지, D형은 소를 주로 감염시킨다. 감염은 에어로졸, 체액 접촉을 통해 이루어지며, 백신과 항바이러스제로 예방 및 치료가 가능하다.
더 읽어볼만한 페이지
- 오르토믹소바이러스과 - 인플루엔자바이러스 B형
인플루엔자바이러스 B형은 RNA 바이러스의 일종으로, A형과는 다른 항원 교차 반응을 보이며, B/야마가타와 B/빅토리아 계통으로 나뉘고, A형보다 느린 속도로 돌연변이를 일으켜 매년 백신 성분이 변경된다. - 오르토믹소바이러스과 - 인플루엔자바이러스 A형
인플루엔자바이러스 A형은 헤마글루티닌(H)과 뉴라미니다아제(N) 조합에 따라 아형이 나뉘며 조류, 돼지, 사람 등 다양한 종에 감염될 수 있고, 높은 변이율로 팬데믹을 일으킬 수 있어 지속적인 감시가 필요한 바이러스이다. - 바이러스 과 - 코로나바이러스과
코로나바이러스과는 표면의 스파이크 단백질 모양이 코로나와 유사한 RNA 바이러스로, 숙주 세포 침투 및 복제 과정을 거쳐 유전적 재조합을 통해 진화하며, 레토바이러스아과와 코로나바이러스아과로 나뉘고 다양한 속과 종으로 분류되며, 사람에게 감기부터 SARS, MERS, 코로나19와 같은 심각한 호흡기 질환을 일으킨다. - 바이러스 과 - 파르보바이러스
파르보바이러스는 선형 단일 가닥 DNA 게놈을 가진 바이러스로, NS 유전자와 VP 유전자를 포함하며, 롤링 헤어핀 복제 방식으로 게놈을 복제하고, 질병을 유발하거나 유전자 치료에 활용된다. - 인플루엔자 - 백신
백신은 약화된 병원체나 일부를 활용하여 인체의 면역계를 활성화, 특정 질병에 대한 항체 생성을 유도하는 의약품으로 감염병 예방에 효과적이지만, 부작용 발생 가능성과 효과의 다양성, 백신 거부와 오해로 인한 접종률 저하 및 감염병 확산의 위험이 존재하며, 지속적인 연구개발을 통해 안전성과 효능이 향상되고 새로운 기술을 이용한 백신들이 개발되고 있다. - 인플루엔자 - 인플루엔자 백신
인플루엔자 백신은 A형 및 B형 인플루엔자 바이러스 감염으로 인한 질병 예방을 위해 사용되는 능동 면역 백신으로, 1918년 스페인 독감 유행 이후 개발되기 시작하여 현재는 다양한 제조 방식의 3가 또는 4가 백신이 사용되고 있으며 매년 유행하는 균주에 맞춰 재구성되지만 효능은 변동될 수 있고 부작용 가능성이 존재하여 지속적인 연구가 진행되고 있다.
| 오르토믹소바이러스과 - [생물]에 관한 문서 | |
|---|---|
| 바이러스 정보 | |
 | |
| 분류 계급 | 속 |
| 하위 분류 | 알파인플루엔자바이러스 베타인플루엔자바이러스 감마인플루엔자바이러스 델타인플루엔자바이러스 이사바이러스 콰란자비루스 토고토바이러스 |
| 어원 | 그리스어 ὀρθός (orthós, 곧은) + μύξα (mýxa, 점액) |
| 추가 정보 | |
| Jones LD, Nuttall PA의 연구 | https://doi.org/10.1016/0035-9203(89)90405-7 |
| Ely의 연구 | http://www.nimr.mrc.ac.uk/MillHillEssays/1999/isa.htm |
| Raynard RS, Murray AG, Gregory A의 연구 | https://doi.org/10.3354/dao046093 |
| |
2. 분류
RNA 바이러스는 계통발생학 기반 분류법에서 음성 가닥 ssRNA 바이러스 하위 범주를 포함하며, 여기에는 ''Articulavirales'' 목과 ''Orthomyxoviridae'' 과가 포함된다. 오르토믹소바이러스과의 속 관련 종 및 혈청형은 다음 표와 같다.
| 속 | 종 (*는 기준종을 나타냄) | 혈청형 또는 아형 | 숙주 |
|---|---|---|---|
| 인플루엔자바이러스 A형 | 인플루엔자 A 바이러스* | H1N1, H1N2, H2N2, H3N1, H3N2, H3N8, H5N1, H5N2, H5N3, H5N8, H5N9, H7N1, H7N2, H7N3, H7N4, H7N7, H7N9, H9N2, H10N7 | 사람, 돼지, 조류, 말, 박쥐 |
| 인플루엔자바이러스 B형 | 인플루엔자 B 바이러스* | 빅토리아, 야마가타[59] | 사람, 물개 |
| 인플루엔자바이러스 C형 | 인플루엔자 C 바이러스* | 사람, 돼지, 개 | |
| 인플루엔자바이러스 D형 | 인플루엔자 D 바이러스* | 돼지, 소 | |
| 연어빈혈증바이러스 | 연어빈혈증바이러스* | 대서양 연어 | |
| 토고토바이러스 | 토고토바이러스* | 진드기, 모기, 포유류 (사람 포함) | |
| 도리 바이러스 | 바트켄 바이러스, 버번 바이러스, 조스 바이러스 | ||
| 콰란자 바이러스[60] | |||
| 콰란필 바이러스,* 존스톤 환초 바이러스 |
3. 구조
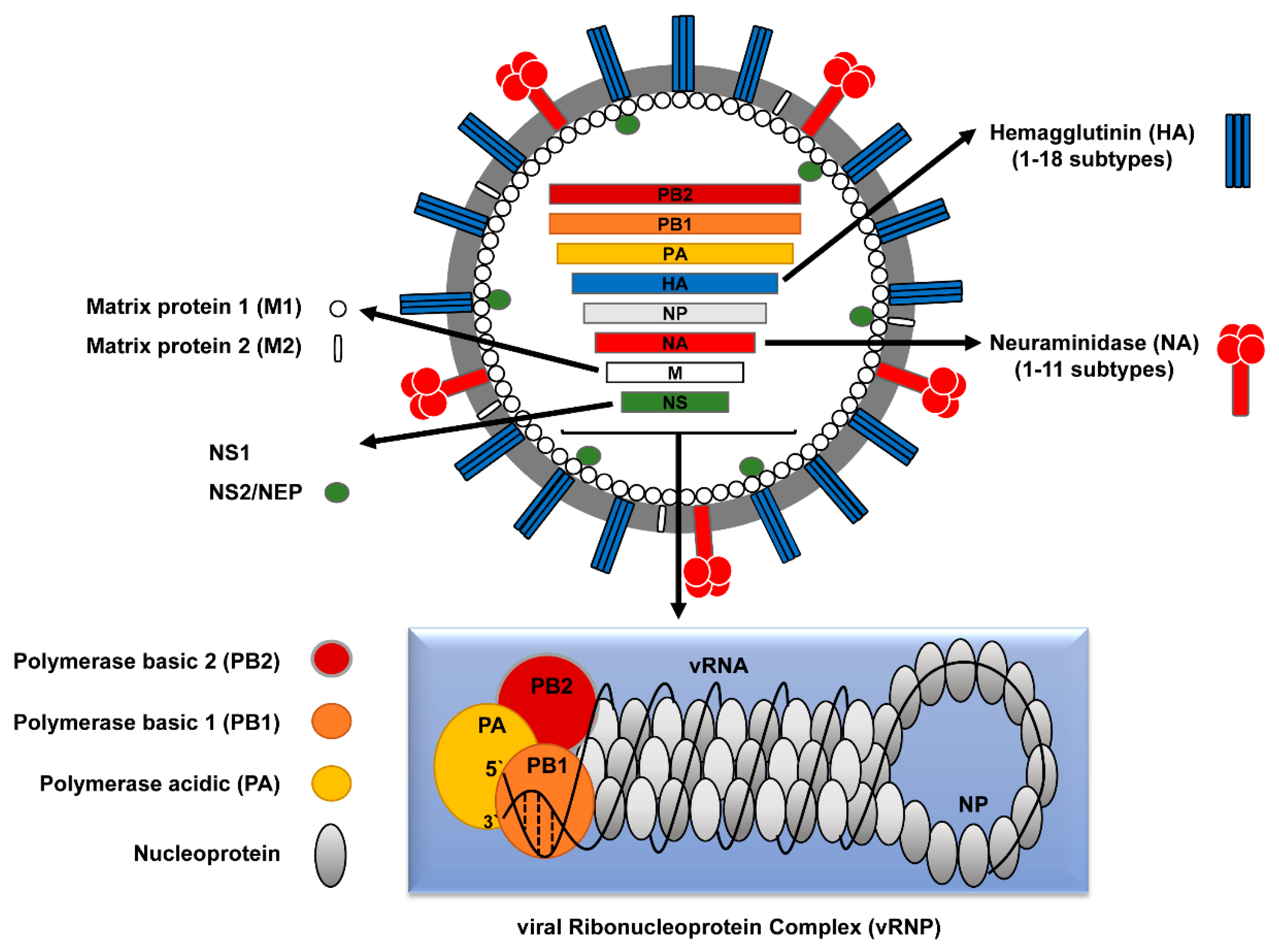
인플루엔자 바이러스의 비리온은 다형성을 띤다. 바이러스 외피는 구형과 실 모양으로 나타날 수 있다. 일반적으로 바이러스의 형태는 타원형이며 입자 직경은 100–120 nm이고, 실 모양의 입자는 직경이 80–100 nm이고 최대 20 μm 길이이다.[5] 외피에는 표면에서 10–14 nm 돌출된 약 500개의 스파이크 모양 표면 돌기가 있으며, 표면 밀도가 다양하다. 주요 당단백질(HA) 스파이크는 뉴라미니다아제 (NA) 스파이크 뭉치에 의해 불규칙하게 삽입되며, HA와 NA의 비율은 약 10:1이다.[6]
글리코 단백질 스파이크가 고정된 지질 이중층 막으로 구성된 바이러스 외피는 뉴클레오캡시드를 둘러싸고 있으며, 각 끝에 고리가 있는 크기가 다른 뉴클레오단백질을 포함한다. 비리온 내의 배열은 불확실하다. 리보핵단백질은 실 모양이며 길이가 50–130 nm, 직경이 9–15 nm이며 나선형 대칭을 이룬다.
3. 1. 게놈
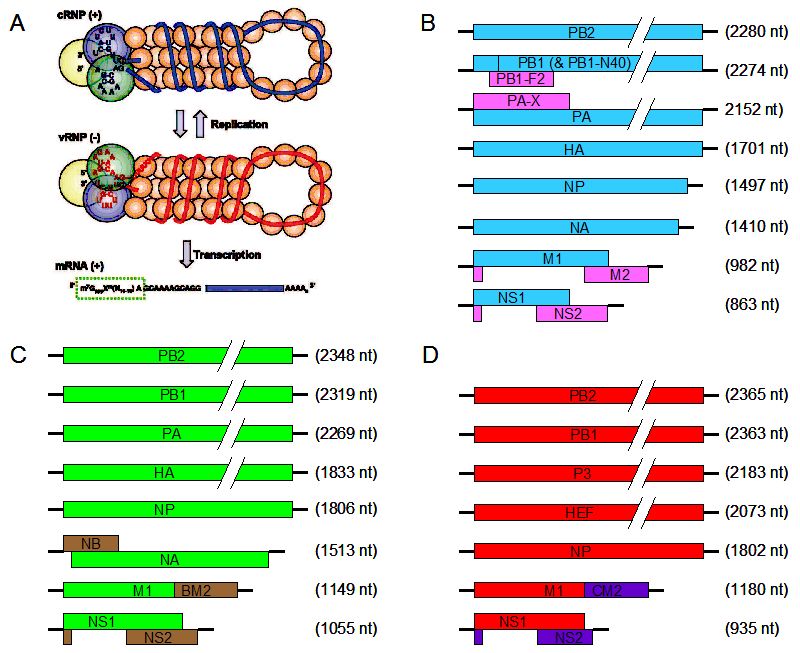
''오르토믹소바이러스과'' 바이러스는 6~8개의 선형 음성 가닥 단일 가닥 RNA 세그먼트를 포함한다. 총 게놈 길이는 10,000~14,600 뉴클레오타이드 (nt)이다.[7] 예를 들어, 인플루엔자 A 게놈은 8개의 분절된 음성 가닥 RNA 조각(총 13.5 킬로베이스)을 가지고 있다.[8]
가장 잘 특성화된 인플루엔자바이러스 단백질은 바이러스 입자 외부에 있는 두 개의 큰 당단백질인 헤마글루티닌과 뉴라미니다제이다. 헤마글루티닌은 바이러스가 표적 세포에 결합하고 바이러스 게놈이 표적 세포에 진입하는 것을 매개하는 렉틴이다.[9] 반대로 뉴라미니다제는 성숙한 바이러스 입자를 결합하는 당을 절단하여 감염된 세포로부터 자손 바이러스를 방출하는 데 관여하는 효소이다. 헤마글루티닌(H) 및 뉴라미니다제(N) 단백질은 항체 및 항바이러스제의 주요 표적이며,[10][11] 인플루엔자 A 바이러스의 서로 다른 혈청형을 분류하는 데 사용되므로 ''H5N1''의 ''H''와 ''N''이 사용된다.
게놈 서열은 말단 반복 서열을 가지고 있으며, 양쪽 끝에서 반복된다. 5' 말단의 말단 반복 서열은 12~13 뉴클레오타이드 길이이다. 3' 말단의 뉴클레오타이드 서열은 동일하며, 동일한 과의 속, 대부분의 RNA(세그먼트) 또는 모든 RNA 종에서 동일하다. 3' 말단의 말단 반복 서열은 9~11 뉴클레오타이드 길이이다. 캡슐화된 핵산은 순전히 게놈이다. 각 바이리온은 결함 간섭 복사본을 포함할 수 있다. 인플루엔자 A(H1N1)에서 PB1-F2는 PB1의 대체 리딩 프레임에서 생성된다. M 및 NS 유전자는 대체 스플라이싱을 통해 두 개의 다른 유전자를 생성한다.[12]
4. 인플루엔자 바이러스 종류
인플루엔자 바이러스에는 4개의 속이 있으며, 각 속은 단일 종 또는 유형을 포함한다. A형 인플루엔자 바이러스와 C형 인플루엔자 바이러스는 다양한 종(사람 포함)에 감염되는 반면, B형 인플루엔자 바이러스는 거의 사람에게만 감염되고, 인플루엔자 D 바이러스는 소와 돼지에게 감염된다.[26][27][28]
- A형 인플루엔자 바이러스
- B형 인플루엔자 바이러스
- C형 인플루엔자 바이러스
- 이사바이러스 (''Isavirus'')
- 쿠아란자바이러스
- 토고토바이러스
4. 1. 인플루엔자 A 바이러스

포유류와 조류를 숙주로 삼으며, 대부분의 경우 비병원성이지만 일부 아종의 경우 숙주에게 인플루엔자를 일으킨다.[64] 인플루엔자바이러스 A형은 보통 큰 병으로 확산되지 않는데, 특정 변이가 닭이나 오리 등의 가정 가금류와 인간에게 심각한 질병을 가져오는 경우가 있다. 때때로 바이러스는 야생 수생 조류에서 가금류로 전염되고, 이는 인플루엔자 전염병을 일으킬 수 있다.[65][66] 이들은 유전물질로 단일가닥, 분절형의 RNA를 가지고 있다.
A형 인플루엔자 바이러스는 바이러스 표면 단백질인 혈구 응집소(HA 또는 H)와 뉴라미니다아제 (NA 또는 N)를 기반으로 추가 분류된다. 자연계에서 18개의 HA 아형(또는 혈청형)과 11개의 NA 아형의 A형 인플루엔자 바이러스가 분리되었다. 이 중 HA 아형 1-16과 NA 아형 1-9는 야생 물새와 해안 조류에서 발견되며, HA 아형 17-18과 NA 아형 10-11은 박쥐에서만 분리되었다.[29][30]
추가적인 변이가 존재하며, 따라서 특정 인플루엔자 균주 분리주는 바이러스 유형, 숙주 종(사람이 아닌 경우), 최초 분리된 지리적 위치, 실험실 참조, 분리 연도 및 HA 및 NA 아형을 지정하는 ''인플루엔자 바이러스 명명법''에 의해 식별된다.[31][32][33]
명명법의 예는 다음과 같다.
# A/브리즈번/59/2007 (H1N1) - 사람에게서 분리
# A/돼지/사우스다코타/152B/2009 (H1N2) - 돼지에서 분리
A형 인플루엔자 바이러스는 세 가지 인플루엔자 유형 중 인간에게 가장 치명적인 병원체이며 가장 심각한 질병을 유발한다. 모든 인플루엔자 A 바이러스가 유행 또는 대유행을 일으키는 것은 야생 조류에서 기원하는 것으로 생각된다.[34] 1900년대 이후 모든 A형 인플루엔자 바이러스 대유행은 다른 인플루엔자 균주와의 재조합을 통해 조류 인플루엔자에 의해 발생했으며, 이는 인간에게 영향을 미치는 경우(계절성 독감) 또는 다른 동물에게 영향을 미치는 경우(2009년 돼지 인플루엔자 대유행 참조)가 있다.[35]
- H1N1은 1918년 "스페인 독감"과 2009년 "돼지 독감"을 유발했다.[36]
- H2N2는 "아시아 독감"을 유발했다.
- H3N2는 "홍콩 독감"을 유발했다.
- H5N1은 "조류" 또는 "조류 독감"으로 불린다.[37]
- H7N7은 특이한 인수 공통 감염 가능성을 가지고 있다.[38]
- H1N2는 돼지와 인간에게 감염된다.[39]
- H9N2, H7N2, H7N3, H10N7도 존재한다.
| 대유행 이름 | 날짜 | 사망자 수 | 치사율 | 관련 아형 | 대유행 심각도 지수 |
|---|---|---|---|---|---|
| 1889–1890년 인플루엔자 대유행 (아시아 또는 러시아 독감)[42] | 1889–1890 | 100만 | 0.15% | 아마도 H3N8 또는 H2N2 | |
| 1918년 인플루엔자 대유행 (스페인 독감)[43] | 1918–1920 | 2천만~1억 | 2% | H1N1 | 5 |
| 아시아 독감 | 1957–1958 | 100만~150만 | 0.13% | H2N2 | 2 |
| 홍콩 독감 | 1968–1969 | 75만~100만 | <0.1% | H3N2 | 2 |
| 러시아 독감 | 1977–1978 | H1N1 | |||
| 2009년 인플루엔자 대유행[44][45] | 2009–2010 | 105,700–395,600[46] | 0.03% | H1N1 |
4. 1. 1. 인플루엔자 A 바이러스에 의해 발생한 질병
- H1N1은 1918년 "스페인 독감"과 2009년 "돼지 독감"을 유발했다.[36]
- H2N2는 "아시아 독감"을 유발했다.
- H3N2는 "홍콩 독감"을 유발했다.
- H5N1은 "조류" 또는 "조류 독감"으로 불린다.[37]
- H7N7은 특이한 인수 공통 감염 가능성을 가지고 있다.[38]
- H1N2는 돼지와 인간에게 감염된다.[39]
- H9N2, H7N2, H7N3, H10N7도 존재한다.
A형 인플루엔자 바이러스는 바이러스 표면 단백질인 혈구 응집소(HA 또는 H)와 뉴라미니다아제 (NA 또는 N)를 기반으로 추가 분류된다.
4. 2. 인플루엔자 B 바이러스
B형 인플루엔자 바이러스는 사람과 물개만을 숙주로 삼아 인플루엔자를 일으킨다.[67] 숙주 범위가 좁아 다양한 숙주를 가질 수 있는 A형 인플루엔자 바이러스에 비해 범유행을 일으키는 경우가 매우 적다.[68][69][70]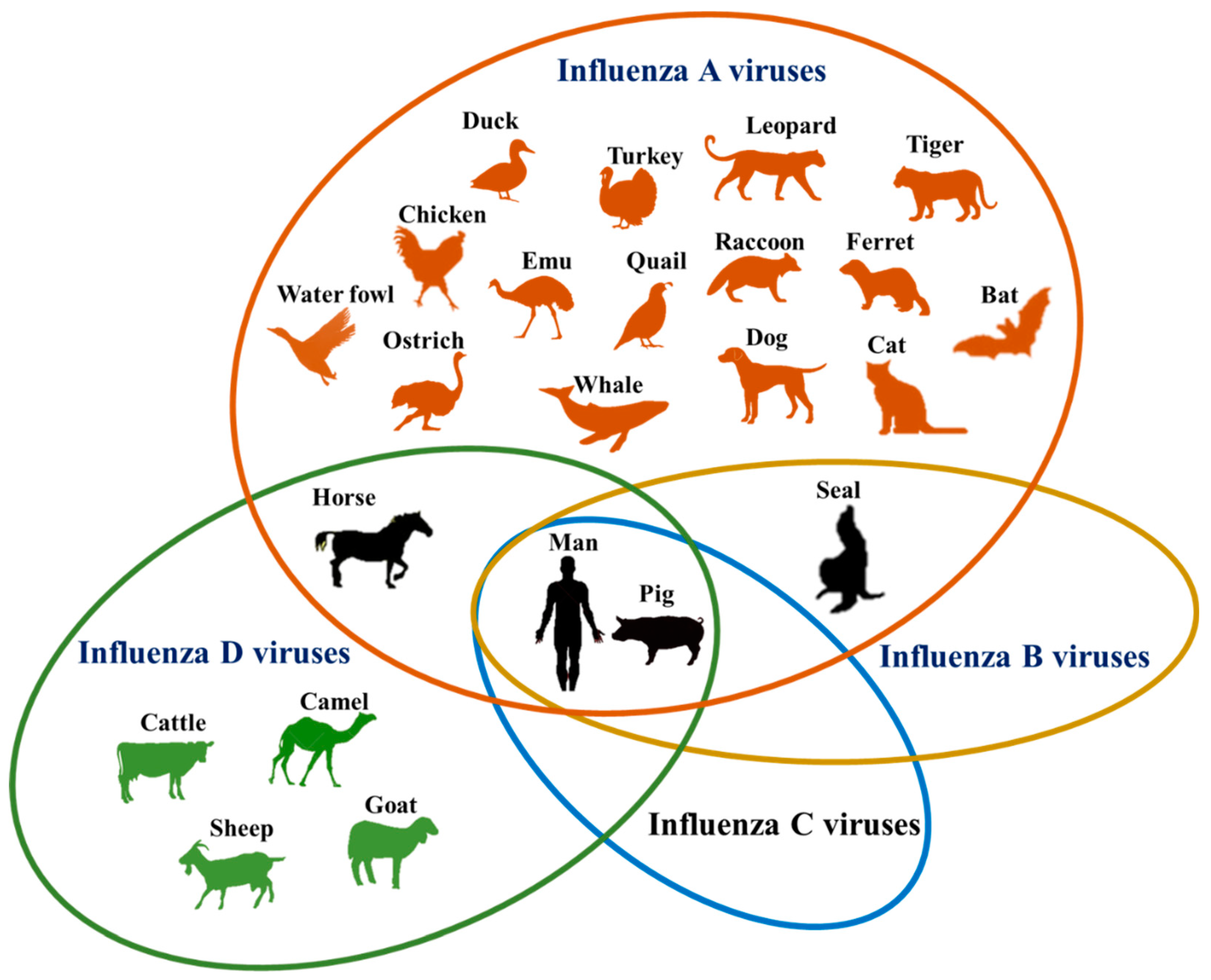
B형 인플루엔자 바이러스는 거의 전적으로 인간 병원체이며, A형 인플루엔자보다 흔하지 않다. B형 인플루엔자 감염에 취약한 것으로 알려진 유일한 다른 동물은 물범이다.[47] 이 유형의 인플루엔자는 A형보다 2~3배 낮은 속도로 변이하며[48] 유전적으로 덜 다양하며, 단 하나의 B형 인플루엔자 혈청형만 존재한다.[26] 이러한 항원 다양성의 부족으로 인해 B형 인플루엔자에 대한 어느 정도의 면역력은 일반적으로 어린 나이에 획득된다. 그러나 B형 인플루엔자는 영구적인 면역이 불가능할 정도로 충분히 변이한다.[49] 이러한 항원 변화율 감소는 제한된 숙주 범위(종간 항원 이동을 억제)와 결합하여 B형 인플루엔자 대유행이 발생하지 않도록 한다.[50]
4. 3. 인플루엔자 C 바이러스
C형 인플루엔자 바이러스는 사람과 돼지를 감염시키며 심각한 질병과 지역 유행을 일으킬 수 있다.[51] 하지만 다른 유형에 비해 덜 흔하며 일반적으로 어린이에게서 경미한 질병을 유발한다.[52][53] A형이나 B형에 비해 발병 빈도는 적지만, 일단 감염되면 심각한 상태를 일으키고, 국소적인 유행을 일으킬 수도 있다. 그러나 다른 인플루엔자 바이러스에 비해서 변이 속도가 느리기 때문에 범유행을 일으키지는 않는다. 인플루엔자바이러스 C형에 대한 백신은 2010년 3월 현재 개발된 것이 없다.[71]4. 4. 인플루엔자 D 바이러스
인플루엔자 D 바이러스는 2011년 돼지에게서 처음 분리되었으며, 2016년 오르토믹소바이러스과가 만들어지면서 새로 분류되었다.[72][73] 이전에는 인플루엔자바이러스 C형의 아류로 여겨져 인플루엔자바이러스 C형속에 속했었다.[72] C형 인플루엔자와 가장 밀접한 관련이 있는 것으로 보이며, 수백 년 전에 분기된 것으로 추정된다.[55] 주요 숙주는 소로 보이지만, 돼지도 감염되는 것으로 알려져 있다.[56]5. 증식
오르토믹소바이러스는 주로 기침이나 재채기를 통해 생성되는 에어로졸, 감염된 조류의 배설물, 타액, 비강 분비물, 대변 및 혈액을 통해 전염된다.[13][14][15] 이러한 체액이나 오염된 표면과의 접촉을 통해 감염이 발생하며, 바이러스는 인체 온도에서 약 1주일, 에서 30일 이상, 매우 낮은 온도에서는 무기한으로 감염성을 유지할 수 있다. 소독제와 세제로 쉽게 비활성화할 수 있다.[13][14][15]
바이러스는 헤마글루티닌 당단백질과 폐, 인후 상피 세포 표면의 시알산 당의 상호 작용을 통해 세포에 결합한다.[16] 엔도사이토시스를 통해 세포 내로 유입된 후, 산성 엔도솜에서 헤마글루티닌 단백질의 일부가 바이러스 외피를 소포 막과 융합시켜 바이러스 RNA(vRNA), 부속 단백질, RNA 의존성 RNA 중합 효소를 세포질로 방출한다.[17] 이들은 세포 핵으로 이동하여 RNA 의존성 RNA 중합 효소가 상보적인 양성 가닥 cRNA를 전사한다.[18] cRNA는 세포질로 나가 번역되거나 핵에 남아있게 된다. 새로 합성된 바이러스 단백질은 골지체를 통해 세포 표면으로 분비되거나, 핵으로 다시 수송되어 vRNA에 결합하여 새로운 바이러스 게놈 입자를 형성한다. 다른 바이러스 단백질은 숙주 세포 내에서 세포 mRNA 분해, 방출된 뉴클레오티드를 vRNA 합성에 사용, 숙주 세포 mRNA 번역 억제 등 여러 작용을 한다.[19]
음성 가닥 vRNA, RNA 의존성 RNA 전사 효소, 기타 바이러스 단백질은 비리온으로 조립된다. 헤마글루티닌과 뉴라미니다제 분자는 세포막에서 뭉쳐지고, vRNA와 바이러스 코어 단백질은 핵을 떠나 막 돌출부로 들어간다. 성숙한 바이러스는 숙주 인지질 막의 구체에서 세포에서 싹이 트며 막 외피로 헤마글루티닌과 뉴라미니다제를 획득한다.[20] 뉴라미니다제가 숙주 세포에서 시알산 잔기를 절단하면 성숙한 바이러스가 분리된다.[16] 이후 숙주 세포는 죽는다.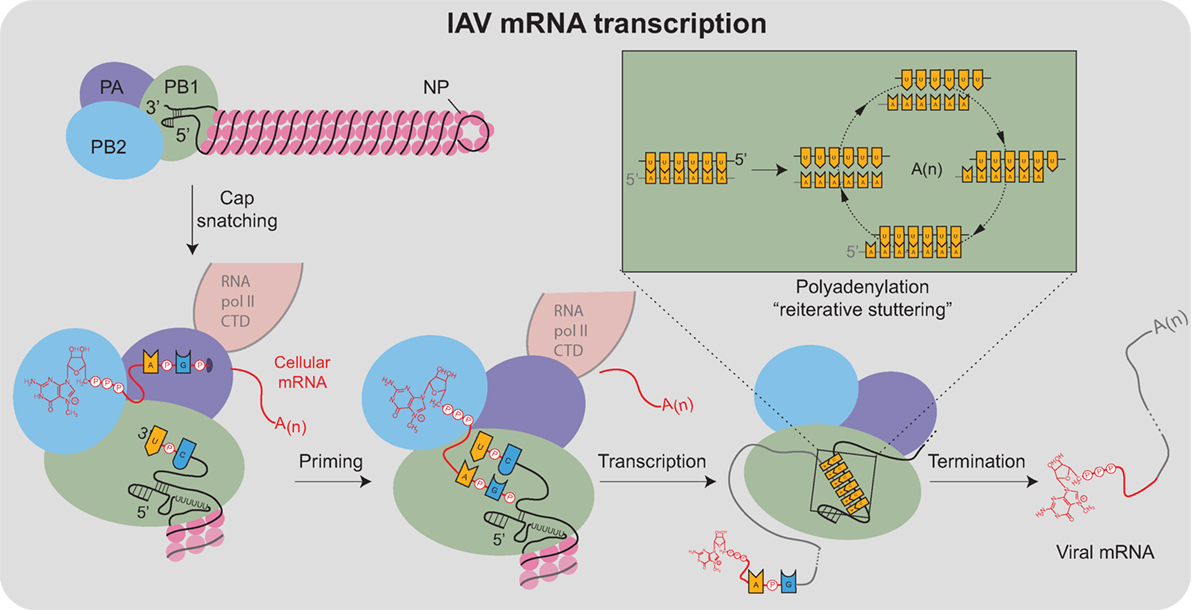
오르토믹소바이러스는 핵에서 복제되는 RNA 바이러스로, 자체 mRNA를 만들 수 없어 캡 스내칭이라는 과정을 통해 세포 RNA를 사용한다.[21] 핵 내에서 RNA 중합 효소 단백질 PB2가 세포 pre-mRNA의 5′ 캡 끝에 결합하고, RNA 중합 효소 PA가 5′ 끝 근처의 세포 mRNA를 절단하여 이 캡된 단편을 바이러스 RNA 게놈 전사를 위한 프라이머로 사용한다.[22] mRNA가 세포 리보솜에서 번역되려면 5′ 캡이 필요하기 때문이다.
RNA 교정 효소가 없어 RNA 의존성 RNA 전사 효소는 약 10,000개의 뉴클레오티드마다 오류를 발생시켜, 거의 모든 새로 제조된 인플루엔자 바이러스는 게놈에 돌연변이를 포함한다.[23] 게놈이 8개의 개별 vRNA 세그먼트로 분리되어 있어, 여러 종류의 인플루엔자 바이러스가 동일 세포를 감염(중복 감염)시키면 유전자 혼합(재분류)이 가능하다. 이는 새로운 특성을 가진 바이러스 출현으로 이어질 수 있으며, 항원 이동을 유발할 수 있다.[10]
6. 백신 및 치료
인플루엔자 바이러스 감염의 예방 및 치료를 위해 백신과 약물이 사용 가능하다. 백신은 H1N1 및 H3N2 사람 인플루엔자 A 바이러스와 인플루엔자 B 바이러스의 비활성화 또는 약독화된 바이리온으로 구성된다. 야생 바이러스의 항원성이 진화하기 때문에 백신은 시드 균주를 업데이트하여 매년 재조합된다.[58]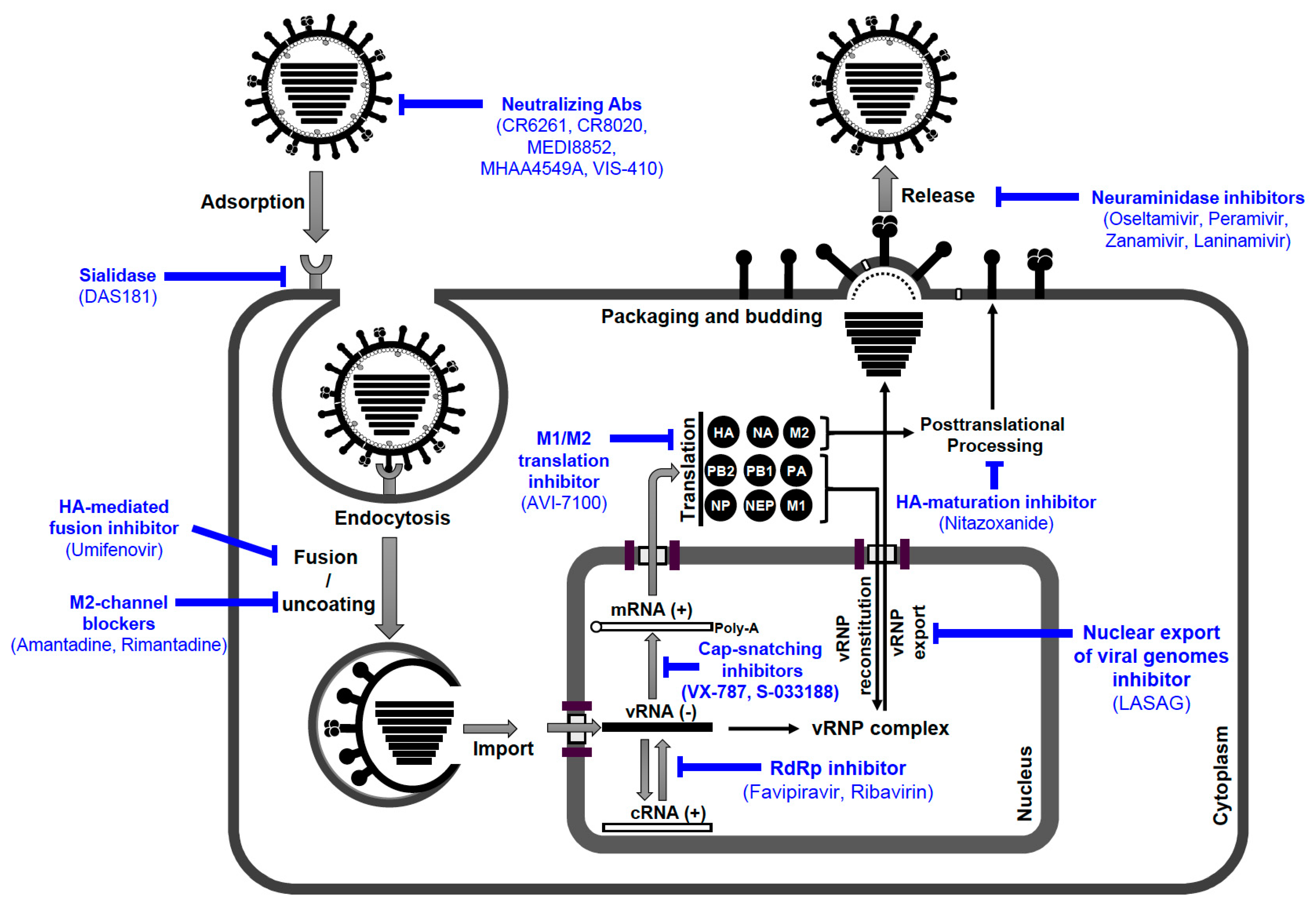
시드 균주와 야생 바이러스의 항원성이 일치하지 않으면 백신은 백신 접종자를 보호하지 못한다. 또한 일치하더라도 탈출 변이체가 자주 생성된다.[58]
인플루엔자 치료에 사용할 수 있는 약물에는 아만타딘과 리만타딘이 있으며, 이는 M2 양성자 채널을 방해하여 바이리온의 코팅 해제를 억제하고, 오셀타미비르(타미플루)(Tamiflu)라는 브랜드 이름으로 판매됨), 자나미비르, 페라미비르는 NA를 방해하여 감염된 세포에서 바이리온의 방출을 억제한다. 그러나 전자의 약물에 대해서는 탈출 변이체가 자주 생성되고, 후자의 약물에 대해서는 덜 자주 생성된다.[58]
참조
[1]
간행물
International Committee on Taxonomy of Viruses
https://www.ncbi.nlm[...]
Columbia University, New York
2006
[2]
논문
Non-viraemic transmission of Thogoto virus: influence of time and distance
[3]
웹사이트
Infectious Salmon Anaemia
http://www.nimr.mrc.[...]
National Institute for Medical Research
2007-09-14
[4]
논문
Infectious salmon anaemia virus in wild fish from Scotland
[5]
논문
Native morphology of influenza virions
2012-01-03
[6]
논문
SnapShot: Influenza by the Numbers
https://www.cell.com[...]
2020-07-23
[7]
웹사이트
ICTV Ninth Report; 2009 Taxonomy Release: ''Orthomyxoviridae''
https://ictv.global/[...]
2020-09-19
[8]
논문
Large-scale sequencing of human influenza reveals the dynamic nature of viral genome evolution
2005-10
[9]
논문
Sialobiology of influenza: molecular mechanism of host range variation of influenza viruses
2005-03
[10]
논문
Realities and enigmas of human viral influenza: pathogenesis, epidemiology and control
2002-08
[11]
논문
Recent strategies in the search for new anti-influenza therapies
2003-07
[12]
논문
The biology of influenza viruses
2008-09
[13]
논문
The effect of various disinfectants on detection of avian influenza virus by real time RT-PCR
[14]
웹사이트
Avian Influenza (Bird Flu) Implications for Human Disease. Physical characteristics of influenza A viruses.
http://www.cidrap.um[...]
University of Minnesota
2024-03-12
[15]
뉴스
Flu viruses 'can live for decades' on ice
http://www.nzherald.[...]
2006-11-30
[16]
논문
Functional balance between haemagglutinin and neuraminidase in influenza virus infections
2002-05
[17]
논문
Visualizing infection of individual influenza viruses
2003-08-05
[18]
논문
Trafficking of viral genomic RNA into and out of the nucleus: influenza, Thogoto and Borna disease viruses
2003-09
[19]
논문
Hijacking of the host-cell response and translational control during influenza virus infection
2006-07
[20]
논문
Assembly and budding of influenza virus
2004-12
[21]
웹사이트
Cap Snatching
http://viralzone.exp[...]
expasy
2014-09-11
[22]
논문
The cap-snatching endonuclease of influenza virus polymerase resides in the PA subunit
2009-04
[23]
논문
Rates of spontaneous mutation among RNA viruses
1993-05-01
[24]
논문
Differentiation of influenza B virus lineages Yamagata and Victoria by real-time PCR
http://edoc.rki.de/o[...]
2010-04
[25]
문서
ICTV Taxonomy History
http://www.ictvonlin[...]
ICTV
2006-06-06
[26]
논문
The evolution of human influenza viruses
2001-12-29
[27]
웹사이트
Avian Influenza (Bird Flu)
https://www.cdc.gov/[...]
Centers for Disease Control and Prevention
2007-09-15
[28]
논문
The emerging influenza virus threat: status and new prospects for its therapy and control
2018-04
[29]
논문
An overview of the epidemiology of avian influenza
https://pubmed.ncbi.[...]
2007-07-26
[30]
논문
Bat-derived influenza-like viruses H17N10 and H18N11
2014-04
[31]
논문
A revision of the system of nomenclature for influenza viruses: a WHO Memorandum
1980
[32]
서적
Epidemiology and Prevention of Vaccine-Preventable Diseases
https://www.cdc.gov/[...]
Centers for Disease Control and Prevention
[33]
웹사이트
Avian Influenza (Bird Flu): Implications for Human Disease
http://www.cidrap.um[...]
Center for Infectious Disease Research & Policy, University of Minnesota
2007-09-14
[34]
논문
Evolution and ecology of influenza A viruses
1992-03
[35]
논문
Influenza: the once and future pandemic
2010-04
[36]
논문
Unraveling the Mystery of Swine Influenza Virus
2009-06
[37]
논문
Pandemic influenza – including a risk assessment of H5N1
2009-04
[38]
논문
Avian influenza A virus (H7N7) associated with human conjunctivitis and a fatal case of acute respiratory distress syndrome
[39]
논문
Emergence of a novel swine-origin influenza A virus (S-OIV) H1N1 virus in humans
2009-07
[40]
논문
A history of influenza
2001-10
[41]
웹사이트
Ten things you need to know about pandemic influenza
https://www.who.int/[...]
World Health Organization
2005-10-14
[42]
논문
Transmissibility and geographic spread of the 1889 influenza pandemic
2010-05
[43]
논문
Transmissibility of 1918 pandemic influenza
2004-12
[44]
논문
Mortality from pandemic A/H1N1 2009 influenza in England: public health surveillance study
[45]
웹사이트
ECDC Daily Update – Pandemic (H1N1) 2009 – January 18, 2010
http://ecdc.europa.e[...]
European Centre for Disease Prevention and Control
2010-01-18
[46]
Submitted manuscript
Estimated global mortality associated with the first 12 months of 2009 pandemic influenza A H1N1 virus circulation: a modelling study
https://zenodo.org/r[...]
2012-09
[47]
논문
Influenza B virus in seals
2000-05
[48]
논문
Comparison of the mutation rates of human influenza A and B viruses
2006-04
[49]
논문
Evolution and ecology of influenza A viruses
1992-03
[50]
논문
Epidemiology and pathogenesis of influenza
1999-11
[51]
논문
Antigenic and genetic characterization of influenza C viruses which caused two outbreaks in Yamagata City, Japan, in 1996 and 1998
[52]
논문
Clinical features of influenza C virus infection in children
2006-05-01
[53]
논문
An outbreak of type C influenza in a children's home
1983-07
[54]
논문
Isolation of a novel swine influenza virus from Oklahoma in 2011 which is distantly related to human influenza C viruses
2013-02
[55]
논문
Genomic and evolutionary characterization of a novel influenza-C-like virus from swine
2014-02
[56]
논문
Cocirculation of two distinct genetic and antigenic lineages of proposed influenza D virus in cattle
2015-01
[57]
웹사이트
Influenza
http://www.cfsph.ias[...]
Iowa State University
2016-02
[58]
논문
Natural selection on the influenza virus genome
https://academic.oup[...]
Oxford Academic
2006-10
[59]
저널
Differentiation of influenza B virus lineages Yamagata and Victoria by real-time PCR
http://edoc.rki.de/o[...]
2010-04
[60]
간행물
ICTV Taxonomy History
http://www.ictvonlin[...]
ICTV
2006-06-06
[61]
저널
The evolution of human influenza viruses
https://www.ncbi.nlm[...]
2001-12-29
[62]
웹인용
Information on Avian Influenza
https://www.cdc.gov/[...]
2019-03-21
[63]
저널
The emerging influenza virus threat: status and new prospects for its therapy and control
https://www.ncbi.nlm[...]
2018-04
[64]
웹인용
Avian influenza (" bird flu")
http://www.who.int/m[...]
2010-02-26
[65]
서적
Animal viruses : molecular biology
https://www.worldcat[...]
2008
[66]
서적
Influenza virology : current topics
https://www.worldcat[...]
2006
[67]
저널
Influenza B virus in seals
[68]
저널
The evolution of human influenza viruses
https://www.ncbi.nlm[...]
2001-12-29
[69]
저널
Genetic diversity of influenza B virus: the frequent reassortment and cocirculation of the genetically distinct reassortant viruses in a community
https://www.ncbi.nlm[...]
2004-09
[70]
저널
Comparative analysis of evolutionary mechanisms of the hemagglutinin and three internal protein genes of influenza B virus: multiple cocirculating lineages and frequent reassortment of the NP, M, and NS genes
https://www.ncbi.nlm[...]
1999-05
[71]
서적
부모도 반의사가 되어야 한다
청문각
1998-04-01
[72]
저널
Novel Influenza D virus: Epidemiology, pathology, evolution and biological characteristics
2017-08-25
[73]
웹인용
New virus gets official name, influenza D
https://www.scienced[...]
ScienceDaily
2016-09-01
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com