큰박쥐류
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
큰박쥐류는 박쥐목에 속하는 대형 박쥐로, 모든 종이 과일이나 꿀을 먹는 채식성이다. 반향정위를 사용하지 않고 시각과 후각에 의존하며, 개와 같은 얼굴, 두 번째 발가락의 발톱, 단순한 귀 모양을 특징으로 한다. 가장 큰 종은 날개폭이 1.7m에 달하고, 작은 종은 14.2g의 무게를 갖는 등 크기가 다양하다.
큰박쥐류는 아프리카, 아시아, 오스트레일리아, 인도양, 오세아니아 등 구세계 열대 지역에 널리 분포하며, 서식지 파괴, 사냥, 질병 매개체 역할 등으로 인해 위협받고 있다. 종자 살포에 중요한 역할을 하며, 일부 문화에서 식용 또는 의례용으로 사용되기도 한다. 큰박쥐과는 6개의 아과로 분류되며, 혀를 사용하여 원시적인 반향정위를 하는 종도 있다.
더 읽어볼만한 페이지
- 큰박쥐과 - 망치머리박쥐
망치머리박쥐는 아프리카에 서식하며 수컷의 독특한 머리 형태로 이름 붙여진 과일박쥐로, 렉 시스템을 통해 번식하고 과일을 섭취하며 에볼라 바이러스의 잠재적 매개체로 연구되기도 하는 종이다. - 큰박쥐과 - 스테드패스트관코과일박쥐
- 생물 계통도에 관한 - 눈표범
눈표범은 고양이과 동물로 중앙아시아와 남아시아의 고산 지대에 서식하며, 길고 두꺼운 털, 큰 발, 긴 꼬리가 특징이며, 유전자 분석 결과 호랑이와 가장 가까운 종으로 밝혀졌으며, 밀렵과 서식지 파괴로 인해 개체 수가 감소하여 여러 국가에서 보호 노력이 진행 중이다. - 생물 계통도에 관한 - 벵골호랑이
벵골호랑이는 인도아대륙에서 서식하는 호랑이 아종으로, 노란색~주황색 털과 검은 줄무늬를 가지며, 멧돼지, 사슴 등 우제류를 먹이로 하며, 서식지 파괴와 밀렵으로 인해 제한적인 지역에서 서식한다. - 생물 분류에 관한 - 다람쥐
다람쥐는 등 쪽에 줄무늬가 있는 다람쥐속 설치류로, 홀로 생활하며 겨울잠을 자고 씨앗, 견과류, 곤충 등을 먹으며 맹금류 등의 먹이가 되고, 특히 한반도 서식 다람쥐는 새로운 종으로 분류될 가능성이 있으며, 유럽에서는 침입 외래종으로 지정되기도 하고 라임병을 옮길 수 있어 주의가 필요한 동물이다. - 생물 분류에 관한 - 황금랑구르
황금랑구르는 인도 아삼 주와 부탄에 분포하며 크림색에서 황금색 털을 가진 멸종위기종 영장류로, 서식지 파괴로 개체수가 감소하여 보호받고 있다.
| 큰박쥐류 - [생물]에 관한 문서 | |
|---|---|
| 분포 | |
| 분포 지역 | 전 세계 |
| 분류 | |
| 학명 | Pteropodidae |
| 명명자 | Gray, 1821 |
| 이명 | Pteropidae (Gray, 1821) Pteropodina C. L. Bonaparte, 1837 |
| 상위 분류 | Pteropodoidea |
| 아목 | Yinpterochiroptera 또는 큰박쥐아목(Megachiroptera) |
| 하위 분류 | Nyctimeninae Cynopterinae Harpyionycterinae Pteropodinae Rousettinae Eidolinae |
| 모식속 | Pteropus |
| 모식속 명명자 | Brisson, 1762 |
| 생물학적 특징 | |
| 최대 몸무게 | 1.45 kg |
| 최대 몸길이 | 1.7 m |
| 최소 몸무게 | 50 g |
| 영어 이름 | Megabat Fruit bat Flying fox |
| 이미지 | |
![다양한 아과의 큰박쥐 종; 시계 방향으로 위쪽 왼쪽부터 [[짧은코과일박쥐|짧은코과일박쥐]], [[인도날여우박쥐|인도날여우박쥐]], [[밀짚색과일박쥐|밀짚색과일박쥐]], [[이집트과일박쥐|이집트과일박쥐]]](https://cdn.onul.works/wiki/noimage.png) | |
| |
| |
2. 특징
큰박쥐류는 다른 박쥐들과 달리 반향정위를 하지 않고, 발달된 시각과 후각을 이용하여 이동하고 먹이를 찾는다.
큰박쥐류는 머리 앞쪽에 큰 눈을 가지고 있는데,[63] 이는 박쥐의 공통 조상보다 더 크다.[61] 일반꽃박쥐는 지름 5.03mm로 가장 작은 눈을 가졌고, 큰과일박쥐는 지름 12.34mm로 가장 큰 눈을 가졌다.[61] 홍채는 보통 갈색이지만, 루손큰박쥐, 피지여우박쥐, 여우박쥐속, 큰박쥐속 일부 종은 붉은색이나 주황색을 띠기도 한다.[62]
밝은 곳에서는 사람보다 시력이 떨어지지만, 어두운 곳에서는 더 뛰어나다.[63] 루셋박쥐속, 켄쇼박쥐속, 스트로박쥐속, 큰박쥐속 일부 종을 대상으로 한 연구에서, 큰박쥐속을 제외한 세 속은 눈에 반사판을 가지고 있어 어두운 곳에서 시력을 보완한다.[64] 연구된 모든 종은 망막에 간상세포와 추상세포를 모두 가지고 있었지만, 큰박쥐속만이 단파장을 감지하는 S추상세포를 가지고 있었다. 옵신의 스펙트럼 조정을 확인할 수 없어, 큰박쥐속의 S추상세포가 청색광을 감지하는지, 자외선을 감지하는지는 불분명하다. 큰박쥐속은 두 종류의 추상세포를 가진 이색시이며, 다른 세 속은 S추상세포가 없어 색을 볼 수 없다. 모든 속은 간상세포 밀도가 매우 높아 빛에 대한 감도가 높다. 큰박쥐속과 루셋박쥐속의 간상세포 밀도는 1mm2당 35만~80만 개로, 집쥐, 고양이, 토끼 등 다른 야행성 또는 박명성 동물과 비슷하거나 더 높다.[64]
큰박쥐류는 과일이나 꽃꿀과 같은 먹이를 찾기 위해 후각을 사용하며,[66] 그 예민함은 개와 맞먹는다.[65] 동부관코박쥐와 같은 관코과일박쥐는 입체 후각을 통해 냄새의 흐름을 3차원적으로 추적할 수 있다.[65] 다른 대부분의 박쥐처럼, 큰박쥐류 어미와 새끼는 서로를 인식하기 위해 냄새를 사용한다.[66] 수컷은 어깨에 안드로겐에 민감한 피지선이 발달되어 번식기에 냄새로 영역을 표시하며, 종에 따라 분비물이 다르다.[67] 또한 수컷은 자신의 소변을 몸에 바르는 행동을 한다.[67][68]
큰박쥐류는 ''TAS1R2'' 유전자를 가지고 있어 단맛을 감지할 수 있다. 다른 모든 박쥐와 마찬가지로 ''TAS1R1'' 유전자가 없어 감칠맛을 느낄 수 없다.[66] 또한 여러 개의 ''TAS2R'' 유전자를 가지고 있어 쓴맛을 감지할 수 있다.[69]
2. 1. 형태
큰박쥐류는 그 이름처럼 큰 몸집을 가지고 있다. 가장 큰 종인 큰검은박쥐는 최대 1.6kg의 무게가 나가며, 큰박쥐속(Pteropus)과 ''Acerodon'' 속의 일부 종은 날개폭이 최대 1.7m에 달한다.[93] 하지만 모든 큰박쥐류가 흡혈박쥐류보다 큰 것은 아니다. 예를 들어 얼룩날개과일박쥐는 14.2g에 불과하다.[38] 큰박쥐속과 검은박쥐속의 큰 종들이 큰박쥐류 전체를 대표하는 것처럼 여겨지기도 하지만, 이는 예외적인 경우이며, 실제로는 대부분의 큰박쥐류가 이들보다 훨씬 작다.[4] 2004년 연구에 따르면 큰박쥐류 종의 28%가 50g 미만이다.[38]큰박쥐류는 개와 비슷한 얼굴, 두 번째 발가락의 발톱, 단순한 모양의 귀를 가지고 있어 흡혈박쥐류와 구별된다.[39] 귀가 단순한 이유는 흡혈박쥐류에서 흔히 발견되는 귓바퀴가 없기 때문이다.[39] 검은박쥐속은 얼굴이 짧고 콧구멍이 관 모양이라 개와 같은 모습이 덜하다.[58] 2011년 연구에 따르면, 167종의 큰박쥐류 중 63%는 털 색깔이 균일하지만, 역광채색(4%), 목띠 또는 털뭉치(5%), 줄무늬(10%), 반점(19%) 등 다양한 무늬를 가진 종들도 있다.[40]
흡혈박쥐류와 달리 큰박쥐류는 뒷다리 사이의 비행막인 후비막이 매우 작다.[51] 꼬리 또한 없거나 매우 짧은데,[58] 긴꼬리과일박쥐(Notopteris)는 예외적으로 긴 꼬리를 가지고 있다.[41] 대부분의 큰박쥐류는 날개가 몸통 측면에 붙어 있지만, 도브소니아속은 날개가 척추 가까이에 붙어 있어 등이 벗겨진 것처럼 보인다.[51]
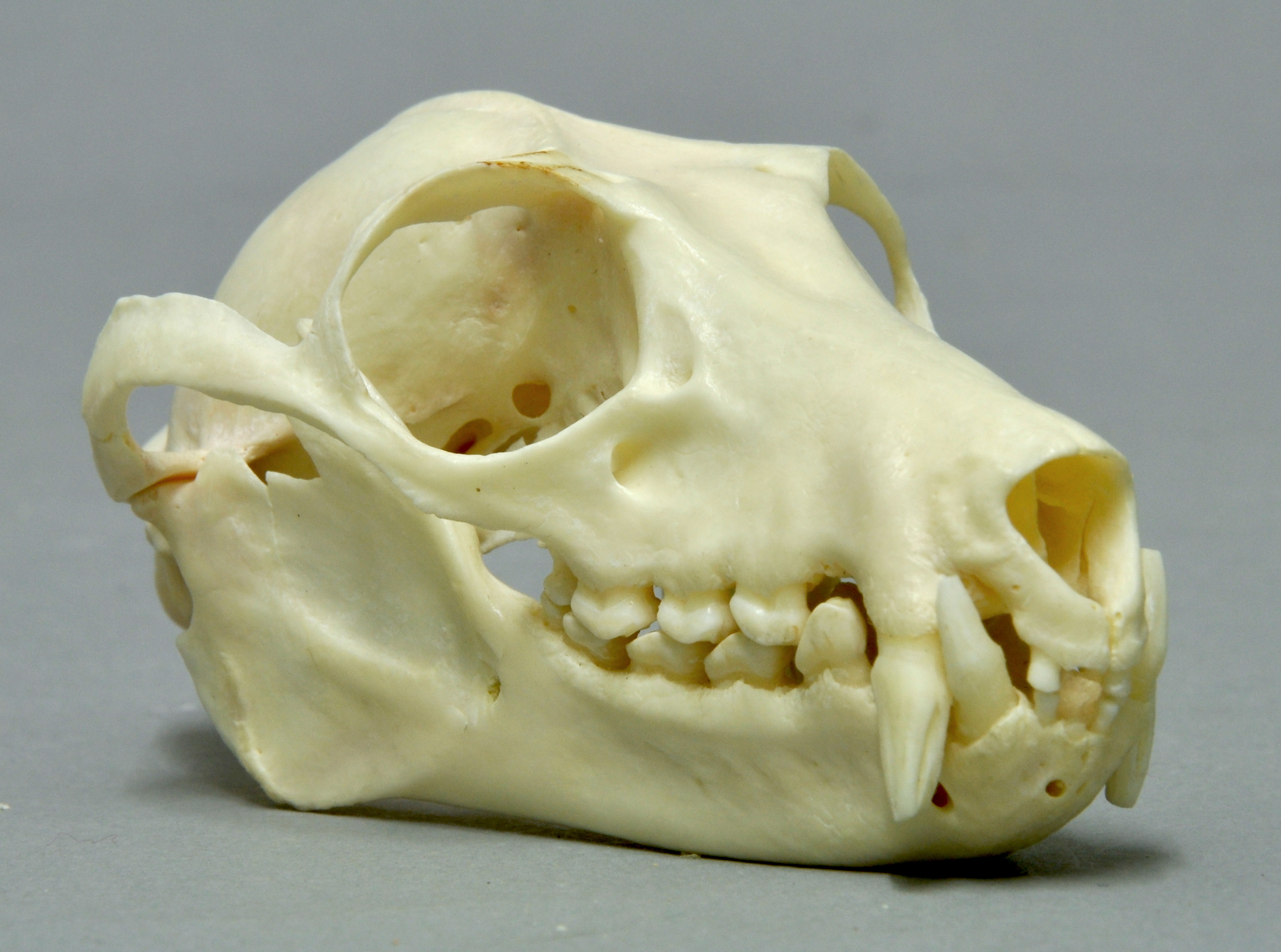
큰박쥐류는 크고 잘 발달된 후안와돌기로 둘러싸인 큰 안와를 가지고 있으며, 후안와돌기는 때때로 합쳐져 후안와봉합을 형성한다. 주둥이는 단순하며, 속에 따라 길이가 다르다. 상악골은 잘 발달되어 있고, 상악골과 융합되지 않고 인대를 통해 관절을 이루어 자유롭게 움직일 수 있다.[43][44] 상악골에는 항상 구개 가지가 없다.[3] 주둥이가 긴 종은 두개골이 활처럼 휘어져 있지만, 얼굴이 짧은 속(''Penthetor'', ''Nyctimene'', ''Dobsonia'', ''Myonycteris'')에서는 거의 휘어지지 않거나 전혀 휘어지지 않는다.[45]
큰박쥐류는 비교적 작은 앞니와 큰 송곳니를 가지고 있으며, 작은어금니와 어금니는 과일을 부수고 뚫는 데 적합하다.[46] 가장 완전한 치식은 I2/2, C 1/1, P3/3, M2/3 x 2 = 34이다.[47] 34개의 치아는 큰박쥐류의 상동형질이다.[49] 치아의 총 개수는 종에 따라 24개에서 34개까지 다양하다. 일부 종은 아래턱 양쪽에 3개가 아닌 2개의 어금니만 있거나, 위턱이나 아래턱의 앞니가 부족할 수 있다.[49] 불머과일박쥐는 앞니가 아예 없고,[49] 상투메칼라과일박쥐는 위턱에 2개, 아래턱에 3개의 앞니를 가지고 있어 비대칭적인 치식을 가진 유일한 포유류이다.[48] 모든 종은 위턱과 아래턱에 각각 2개의 송곳니를 가지고 있으며, 작은어금니는 위턱과 아래턱에 각각 4개 또는 6개가 있다. 첫 번째 위턱과 아래턱 큰어금니는 항상 존재하며, 나머지 큰어금니는 존재하거나, 축소되어 있거나, 없을 수 있다.[49] 큰어금니와 작은어금니는 단순화되어 첨두와 능선이 감소하고 평평한 치관을 형성한다.[52]
대부분의 포유류처럼 큰박쥐류는 이중치성으로, 어린 개체는 유치를 가지며, 이는 영구치로 교체된다. 대부분의 종에서 유치는 20개이며, 포유류의 일반적인 특징처럼 유치에는 큰어금니가 없다.[50][52]

2. 2. 골격
안와는 크고, 그 뒤쪽에는 잘 발달된 후안와돌기가 있다. 후안와돌기는 때때로 합쳐져 안와 후벽을 형성한다. 주둥이는 외관상 단순하며, 다른 박쥐들처럼 큰 변형은 없다.[212] 주둥이의 길이는 속에 따라 다르다. 상악골은 잘 발달되어 있으며,[167] 상악골과 유합되어 있지 않고, 인대를 통해 관절을 이루어 자유롭게 움직일 수 있다.[213][214] 상악골에는 구개지가 없다.[167] 주둥이가 긴 종에서는 두개골이 일반적으로 활처럼 휘어져 있다. 주둥이가 짧은 루카스과일박쥐속, 코큰과일박쥐속, 민과일박쥐속, 흰목큰박쥐속에서는 두개골이 거의 또는 전혀 휘어져 있지 않다.[215]치아의 수는 종에 따라 다르며, 총 24~34개이다. 상악과 하악의 절치는 각각 2개 또는 4개이지만, 뉴기니아과일박쥐처럼 절치가 전혀 없는 종[217]이나 상악에 2개, 하악에 3개의 절치를 가진 상투메과일박쥐 등의 예외가 있다. 이 종은 비대칭적인 치식을 가진 유일한 포유류이다.[216]
모든 종은 상하에 2개의 견치를 가지고 있다. 소구치의 수는 상하 각각 4개 또는 6개이다. 상하에 제1대구치를 가지며, 모든 종은 최소 4개의 대구치를 가지고 있다. 나머지 대구치는 존재하거나, 작거나, 또는 존재하지 않는다.[217] 대구치와 소구치는 단순화되어 있으며, 교두와 융기가 적고 더 평평하다.[220]
대부분의 포유류와 마찬가지로, 새끼는 유치를 가지며, 그 후 영구치로 바뀐다. 대부분의 종에서 유치는 20개이다. 전형적인 포유류와 마찬가지로,[218] 유치에는 대구치가 포함되지 않는다.[220]
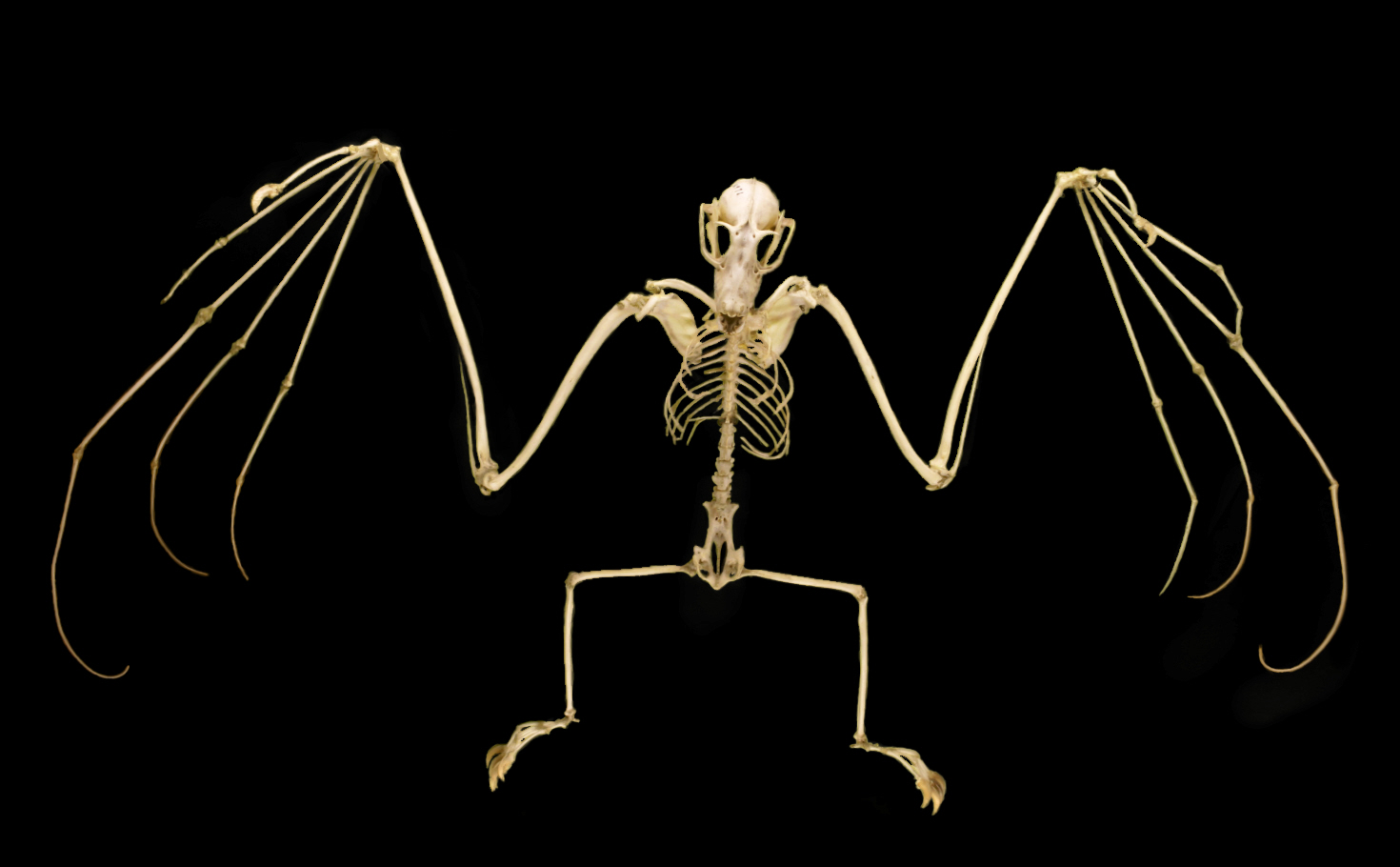
큰박쥐류의 견갑골은 박쥐류 중 가장 원시적인 것으로 여겨진다.[220] 전체적으로 단순한 구조이지만, 몇 가지 특징적인 특징이 있다. 견갑골과 견갑설골근(肩甲舌骨筋)의 연결 부위가 쇄골에서 바깥쪽으로 치우쳐 있으며, 이러한 특징은 긴코박쥐과에서도 수렴진화의 예로 볼 수 있다. 어깨에는 잘 발달된 가느다란 근육띠가 있으며, 목덜미에서 엄지손가락 밑동까지 이어지는 occipito-pollicalis 근의 건을 피부에 고정하고 있다.[226][219]
다른 박쥐류와 달리, 엄지손가락에만 발톱이 있는 것이 아니라, 대부분의 큰박쥐류는 두 번째 손가락에도 발톱이 있다.[220] 단, ''Cynopterus''속, ''Megaerops''속, ''Balionycteris''속, ''Styloctenium''속은 두 번째 손가락에 발톱이 없다.[221] 첫 번째 손가락이 가장 짧고, 세 번째 손가락이 가장 길다. 두 번째 손가락은 구부릴 수 없다.[220] 큰박쥐류의 엄지손가락은 다른 박쥐류보다 전완에 비해 길다.[219]
뒷다리는 인간과 같은 골격 구조를 가지고 있다. 대부분의 큰박쥐류는 종골에서 종골돌기가 나온다.[223] 일부 연구자들은 구조가 다른 다른 박쥐류의 종골돌기와 구분하기 위해 이 구조를 "uropatagial spur (꼬리막 돌기)"라고 부른다. 이 구조는 꼬리막을 안정시키기 위해 존재하며, 비행 중에 막의 휘어짐을 조절할 수 있다. ''Eonycteris''속, ''Syconycteris''속, ''Harpyionycteris''속 등은 종골돌기가 없다.[222] 일반적인 포유류와 비교하면, 다리 전체가 고관절에서 회전하고, 무릎이 뒤쪽을 향하고 있다. 발가락은 모두 시상면 방향으로 구부러지며, 새처럼 반대 방향으로 구부러지는 발가락은 없다.[223]
2. 3. 내부 기관
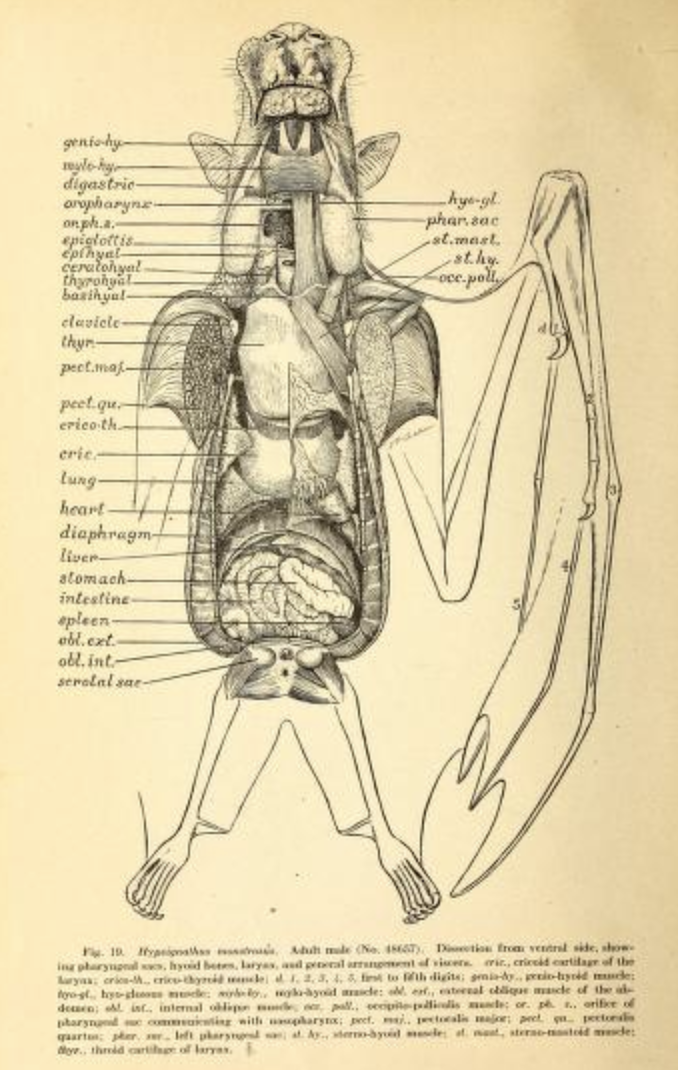
비행은 많은 에너지를 소모하므로 심혈관계에는 비행에 적응한 몇 가지 특징이 있다. 비행 중에는 산소 소비량을 지속적으로 20배 이상 증가시킬 수 있는데, 이는 인간 운동선수가 수 분 동안 최대로 증가시킬 수 있는 양과 비슷하다.[224] 1994년 작은검은박쥐와 흡혈박쥐 연구에서는 이산화탄소 생성량과 산소 사용량의 비율이 약 0.78로 나타났다. 비행 중 최대 심박수는 큰갈색관박쥐 분당 476회, 이집트과일박쥐 분당 728회였다. 분당 최대 호흡수는 큰갈색관박쥐 163회, 작은검은박쥐 316회였다.[225] 몸 크기에 비해 폐 용적이 매우 큰데, 땃쥐과와 같은 육상 포유류는 체중 1g당 0.03cm3인 반면, 황금털박쥐 등은 체중 1g당 0.13cm3로 4.3배나 크다.[224]
소화는 빠르며 장 통과 시간은 30분 이하이다.[226] 소화기관 구조는 초식동물과 유사하며, 부드러운 과일이나 꽃꿀 섭취에 적합하다. 소화 기관 길이는 초식 동물치고는 짧고, 곤충을 먹는 박쥐보다도 짧다. 이는 식이섬유가 구개, 혀, 이빨의 작용으로 분리되어 배출되기 때문이다. 많은 종이 U자형 위를 가지고 있으며, 소장과 대장 사이에 뚜렷한 구분이 없고 직장도 명확하게 시작되지 않는다. 장의 융모 밀도는 매우 높아 영양소 흡수를 위한 표면적을 넓힌다.[227]
2. 4. 감각
큰박쥐류는 극히 예외적인 경우를 제외하고 반향정위를 하지 않으며, 시각과 후각에 의존하여 이동한다.[64] 머리 앞쪽에 큰 눈을 가지고 있으며,[63] 이 눈은 모든 박쥐의 공통 조상보다 크다.[61] 18종의 대형박쥐류 눈을 조사한 연구에 따르면, 일반꽃박쥐의 눈이 지름 5.03mm로 가장 작았고, 큰과일박쥐의 눈이 지름 12.34mm로 가장 컸다.[61] 대형박쥐류의 홍채는 보통 갈색이지만, Desmalopex, Mirimiri, Pteralopex, 그리고 일부 ''Pteropus'' 종처럼 붉은색이나 주황색일 수 있다.[62]
높은 밝기 수준에서 대형박쥐류의 시력은 인간보다 떨어지지만, 낮은 밝기에서는 더 우수하다.[63] 일부 ''Rousettus'', ''Epomophorus'', ''Eidolon'', 그리고 ''Pteropus'' 종의 눈을 조사한 한 연구에 따르면, 처음 세 속은 눈에 반사판을 가지고 있어 낮은 조명 수준에서 시력을 향상시키는 반면, ''Pteropus'' 종은 그렇지 않다.[64] 조사된 모든 종은 망막에 간상세포와 추상세포가 모두 있었지만, ''Pteropus'' 종만이 가장 짧은 파장의 빛을 감지하는 S-추상세포를 가지고 있었다. 옵신의 스펙트럼 조정을 구분할 수 없었기 때문에, ''Pteropus'' 종의 S-추상세포가 청색광 또는 자외선을 감지하는지 여부는 불분명하다. ''Pteropus'' 박쥐는 두 종류의 추상세포를 가지고 있는 이색성이다. S-추상세포가 없는 다른 세 속은 일색성으로 색을 볼 수 없다. 모든 속은 매우 높은 간상세포 밀도를 가지고 있어 빛에 대한 감도가 높았는데, 이는 야행성 활동 패턴과 일치한다. ''Pteropus''와 ''Rousettus''에서 측정된 간상세포 밀도는 제곱밀리미터당 35만~80만 개로, 집쥐, 고양이, 토끼와 같은 다른 야행성 또는 박명성 동물과 같거나 능가한다.[64]
큰박쥐류는 과일이나 꿀과 같은 먹이를 찾는 데 후각을 이용한다.[66] 이들의 후각은 개의 후각과 맞먹을 정도로 예민하다.[65] 동부관코박쥐와 같은 관코과일박쥐는 입체 후각을 가지고 있어 냄새의 흐름을 3차원적으로 추적할 수 있다.[65]
대부분(혹은 모든) 다른 박쥐 종과 마찬가지로, 큰박쥐류 어미와 새끼는 서로를 알아보고 개체를 인식하는 데에도 냄새를 이용한다.[66] 과일박쥐의 경우 수컷은 어깨에 확대된 안드로겐에 민감한 피지선을 가지고 있으며, 특히 번식기 동안 자신의 영역을 표시하는 데 사용한다. 이 분비물은 종에 따라 다르다.[67] 수컷은 또한 소변 씻기를 하거나, 자신의 소변을 몸에 바른다.[67][68]
대형박쥐류는 ''TAS1R2'' 유전자를 가지고 있어 음식의 단맛을 감지할 수 있다. 다른 모든 박쥐와 마찬가지로, 대형박쥐류는 ''TAS1R1'' 유전자가 없기 때문에 감칠맛을 맛볼 수 없다.[66] 대형박쥐류는 또한 여러 개의 ''TAS2R'' 유전자를 가지고 있어 쓴맛을 맛볼 수 있음을 나타낸다.[69]
3. 생태
큰박쥐류는 대부분 과일을 먹거나 꽃꿀을 먹으며, 종자살포를 통해 숲의 생태에 중요한 역할을 한다.
대부분의 큰박쥐류는 과일을 먹는다.[94] 이들은 거의 188개에 달하는 식물 속의 다양한 과일을 섭취한다.[96] 일부 종은 꽃꿀을 먹기도 하며,[94] 특히 호주에서는 유칼립투스 꽃이 중요한 먹이원이다.[58] 그 외에도 잎, 싹, 꽃가루, 씨앗 꼬투리, 수액, 구과, 나무껍질, 가지 등도 먹는다.[95] 큰박쥐류는 식욕이 왕성하여 하룻밤에 자기 체중의 최대 2.5배에 달하는 과일을 먹기도 한다.[96]
큰박쥐류는 먹이를 찾아 서식지 사이를 이동한다. 박쥐 중에서는 비교적 빠르게 직선으로 비행하지만, 일부 종은 더 느리고 기동성이 뛰어나다. 종에 따라 하룻밤에 20km를 이동할 수 있으며, ''Eidolon'', ''Pteropus'', ''Epomophorus'', ''Rousettus'', ''Myonycteris'', ''Nanonycteris'' 속의 이동성 종은 최대 750km까지 이동할 수 있다. 대부분의 큰박쥐류는 평균 이하의 종횡비를 가지며,[97] 날개 하중은 평균 또는 평균 이상이다.[97]
큰박쥐류는 종자살포에 중요한 역할을 한다. 오랜 진화의 결과로, 일부 식물은 박쥐의 감각에 맞는 특징, 즉 강한 향이 나고, 밝은 색을 띠며, 잎사귀에서 눈에 잘 띄는 위치에 있는 열매를 진화시켰다. 열매의 밝은 색깔과 위치는 큰박쥐류가 시각적 단서에 의존하고, 덤불 속을 이동하는 데 어려움을 겪는다는 것을 반영할 수 있다. 40종이 넘는 무화과 종의 열매를 조사한 연구에 따르면, 단 한 종의 무화과만이 조류와 큰박쥐류 모두에게 섭취되었고, 대부분의 종은 조류 또는 큰박쥐류 중 하나에게만 섭취되었다.[98] 조류가 섭취하는 무화과는 붉은색이나 주황색인 경우가 많지만, 큰박쥐류가 섭취하는 무화과는 종종 노란색이나 녹색이다.[98] 빠른 소화 시간으로 인해 대부분의 종자는 섭취 후 곧 배설되지만, 일부 종자는 12시간 이상 장내에 머물 수 있다. 이는 큰박쥐류가 모체 식물로부터 멀리 종자를 퍼뜨릴 수 있는 능력을 높인다.[99] 고도로 이동성이 높은 과일 섭취 동물로서 큰박쥐류는 벌채된 지역에 나무 종자를 퍼뜨림으로써 고립된 산림 파편 사이의 산림을 복원할 수 있다.[100] 이러한 종자 살포 능력은 길이가 4mm 미만인 작은 종자를 가진 식물로 제한되는데, 이보다 큰 종자는 섭취되지 않기 때문이다.[101]
큰박쥐는 여러 기생충 분류군의 숙주이다. 알려진 기생충에는 박쥐파리과와 흡혈파리과 ("박쥐파리"),[106][107] 그리고 ''모낭충'' 속의 진드기가 있다.[108] 혈구충과의 혈액 기생충과 톡소카리과의 장내 선충도 큰박쥐 종에 영향을 미친다.[58][109]
3. 1. 서식지
큰박쥐류는 구세계 열대 지방에 널리 분포하며, 아프리카, 아시아, 오스트레일리아와 인도양 및 오세아니아 제도 전역에서 서식한다.[25]아프리카2013년 기준으로 아프리카에는 14개 속, 28종의 큰박쥐류가 있다. 이 중 24종은 열대 또는 아열대 기후에서만 발견되며, 나머지 4종은 주로 열대 지방에 분포하지만 온대 지역에도 서식한다. 서식지별로 보면, 8종은 주로 삼림에 서식하고, 9종은 삼림과 사바나 모두에 서식하며, 9종은 주로 사바나에 서식하고, 2종은 섬에 서식한다. 아프리카 종 중 긴털큰박쥐는 주로 산악 지역에 서식하지만, 13종은 산악 지역과 다른 지역에도 서식한다.[110]
아시아동남아시아를 제외하고 아시아에서는 큰박쥐류의 종 다양성이 비교적 낮다. 이집트과일박쥐는 분포 지역의 대부분이 구북구에 있는 유일한 큰박쥐류이다.[111] 이집트과일박쥐와 옅은황색과일박쥐만이 중동에서 발견된다.[111][112] 이집트과일박쥐의 분포 지역 최북단은 지중해 북동부이다.[111] 동아시아에서는 중국과 일본에만 분포한다. 중국에서는 6종이 연중 분포하고, 다른 7종은 분포 지역의 끝자락에 있거나, 오인될 가능성이 있거나, 우연한 기록으로 간주된다.[113] 일본에서는 류큐 열도와 오가사와라 제도에 큰박쥐속 4종이 분포한다.[114][115][116][117] 남아시아에서는 몰디브의 2종부터 인도의 13종까지 다양성이 다양하다.[118] 동남아시아에서는 싱가포르에서 5종, 인도네시아에서는 76종이 알려져 있다.[118] 아시아에 분포하는 98종의 큰박쥐류 중 95종이 삼림에 서식한다. 그 외 서식지로는 인간이 개조한 토지(66종), 동굴(23종), 사바나(7종), 관목지(4종), 암석 지대(3종), 초원(2종), 사막(1종)이 있다.[118]
오스트레일리아오스트레일리아에는 큰박쥐속, 꽃꿀먹이박쥐속, 민등큰박쥐속, 코큰박쥐속, 긴혀박쥐속의 5속 8종이 분포한다.[58] 오스트레일리아의 큰박쥐속은 맹그로브 숲, 열대 우림, 관목 지대의 습윤 경엽수림 등 다양한 서식지에서 발견된다.[58] 또한 도시 지역에 대규모 군집을 형성하기 때문에 인간과 공존하는 경우가 많으며, 특히 5월과 6월에 도시 지역 개체군이 가장 많이 발견된다.[119]
오세아니아오세아니아에서는 팔라우와 통가 2개국에 각각 1종씩으로 종 수가 가장 적다. 종 수가 가장 많은 곳은 파푸아뉴기니로 36종이 알려져 있다.[120] 오세아니아의 65종 중 58종이 삼림에 서식한다. 그 외에는 인간이 개조한 토지에 42종, 동굴에 9종, 사바나에 5종, 관목지에 3종, 암석 지대에 3종이 서식한다.[120] 큰박쥐류 중 약 19%는 단일 섬의 고유종으로 추정된다. 박쥐류 중 고유종 비율이 높은 분류군은 2종 모두 마다가스카르섬의 고유종인 흡착발박쥐과이다.[121]
3. 2. 행동
큰박쥐류는 사회적인 동물로, 서로 소통하기 위해 다양한 소리를 낸다. 이러한 소리에는 "떨리는 듯한 소리의 폭발",[83] 경적 소리,[84] 큰 울음소리[85] 등이 있다. 이집트과일박쥐는 음성 학습 능력이 있어, 다른 개체들과의 상호작용을 통해 소리를 배우고 방언을 습득한다.[86][87] 이러한 방언은 무리마다 다른 주파수로 소통하게 만들기도 한다.[88][89]큰박쥐류의 사회적 행동에는 번식 외의 목적으로 성적 행동을 하는 것도 포함된다. 암컷 이집트과일박쥐는 수컷에게서 먹이를 얻는 대가로 성관계를 맺기도 하는데, 친자 확인 결과 먹이를 준 수컷이 새끼의 아버지가 될 확률이 더 높았다.[90] 보닌큰박쥐는 동성 간 펠라티오가 관찰되기도 했는데,[91][92] 이는 추운 환경에서 수컷들이 무리를 짓도록 돕는 행동으로 추정된다.[91][92]
대부분의 큰박쥐류는 야행성 또는 박명박모성 동물이지만, 낮에 활동하는 종도 있다.[93] 특히 포식자가 없는 섬 지역에서는 주행성인 경우가 있는데, 검은귀큰박쥐의 아종,[160] 모리셔스큰박쥐,[160] 캐롤라인큰박쥐,[160] ''Pteropus pelagicus''의 아종,[160] 세이셸과일박쥐[160] 등이 이에 해당한다.
큰박쥐류는 서식지에 따라 나무나 동굴 등 다양한 곳에서 생활한다. 1992년 연구에 따르면, 41개 속 중 29개 속은 나무에, 11개 속은 동굴에 서식하며, 나머지 6개 속은 인공 구조물, 광산, 틈새 등에 서식한다.[73] 나무에 사는 종들은 단독 생활을 하거나 최대 100만 마리까지 모이는 군집을 이루는 반면, 동굴에 사는 종들은 10마리에서 수천 마리까지 무리를 짓는다. 군집성이 강한 종들은 같은 서식지를 오랫동안 사용하는 경향이 있지만, 단독 생활을 하거나 작은 무리를 짓는 종들은 서식지를 자주 옮긴다.[73]

대부분의 큰박쥐는 주로 과일을 먹는 과식성(Frugivore)이다.[262] 이들은 거의 188개에 달하는 식물 속의 다양한 과일을 섭취한다.[264] 일부 종은 꽃꿀을 먹기도 하며,[262] 특히 호주에서는 유칼립투스 꽃이 중요한 먹이원이 된다.[226] 그 외에도 잎, 싹, 꽃가루, 꼬투리, 수액, 구과, 나무껍질, 가지 등도 먹는다.[263] 큰박쥐는 식욕이 왕성하여 하룻밤에 자기 체중의 최대 2.5배에 달하는 과일을 먹기도 한다.[264]
큰박쥐는 일반적으로 직선으로 비교적 빠르게 날지만, 일부 종은 더 느리고 기동성이 뛰어나다. 종에 따라 하룻밤에 20km를 이동할 수 있으며, ''Eidolon'', ''Pteropus'', ''Epomophorus'', ''Rousettus'', ''Myonycteris'' 및 ''Nanonycteris'' 속의 이동성 종은 최대 750km까지 이동할 수 있다. 대부분의 큰박쥐는 평균 이하의 종횡비를 가지며,[265] 날개 하중은 평균 또는 평균 이상이다.[265]
3. 3. 번식
큰박쥐류는 다른 모든 박쥐류와 마찬가지로 포유류 중에서 크기에 비해 수명이 긴 편이다. 일부 사육 중인 큰박쥐류는 30년 이상 생존한 경우도 있다.[53] 큰박쥐류는 생식률이 낮고 성적 성숙이 늦어 대부분 종의 암컷은 1~2살이 되어야 출산을 한다.[73] 일부 큰박쥐류는 일년 내내 번식이 가능해 보이지만, 대부분의 종은 계절 번식 동물일 가능성이 높다.[53] 교미는 서식지에서 이루어진다.[70]임신 기간은 다양하지만,[71] 대부분의 종은 4~6개월이다. 여러 종류의 큰박쥐류는 교미와 출산 사이의 기간을 늘리는 생식적 적응을 가지고 있다. 엷은 황갈색 과일박쥐와 같은 일부 종은 착상 지연이라는 생식 적응을 가지고 있는데, 이는 6월이나 7월에 교미가 일어나지만, 접합자가 몇 달 후인 11월까지 자궁 벽에 착상되지 않는다는 것을 의미한다.[73] 피셔의 왜소과일박쥐는 착상 후 지연 적응을 통해 최대 11.5개월로 박쥐 종 중 가장 긴 임신 기간을 가지고 있다.[71] 착상 후 지연은 배아의 발달이 자궁벽에 착상된 후 최대 8개월 동안 중단됨을 의미하며, 이것이 매우 긴 임신 기간의 원인이다.[73] 큰 짧은코 과일박쥐는 3개월로 짧은 임신 기간을 가지고 있다.[72]
모든 큰박쥐류의 한배 새끼 수는 일반적으로 한 마리이다.[73] 다음 종에서 쌍둥이에 대한 기록이 거의 없다.
| 종 |
|---|
| 마다가스카르 큰박쥐 |
| 돕슨의 어깨털 과일박쥐 |
| 회색머리 큰박쥐 |
| 검은 큰박쥐 |
| 안경 큰박쥐[74] |
| 큰 짧은코 과일박쥐[75] |
| 피터스의 어깨털 과일박쥐 |
| 망치머리 박쥐 |
| 엷은 황갈색 과일박쥐 |
| 작은 칼라 과일박쥐 |
| 이집트 과일박쥐 |
| 레슈노의 붉은박쥐[80] |
쌍둥이의 경우, 두 마리의 새끼가 모두 생존하는 것은 드물다.[74] 모든 박쥐류와 마찬가지로 큰박쥐류는 생식률이 낮기 때문에 개체 수가 감소에서 회복되는 속도가 느리다.[76]
출생 시 큰박쥐류 새끼의 몸무게는 평균적으로 어미의 분만 후 몸무게의 17.5%이다. 이것은 모든 박쥐과 중에서 새끼와 어미의 몸무게 비율이 가장 작은 것이다. 모든 박쥐류에서 신생아의 몸무게는 어미의 분만 후 몸무게의 22.3%이다. 큰박쥐류 새끼는 전통적인 미숙아(출생 시 무력함) 또는 조숙아(출생 시 능력 있음) 범주로 쉽게 분류되지 않는다. 큰 짧은코 과일박쥐와 같은 종은 눈을 뜬 채 태어나지만(조숙아의 징후), 이집트 과일박쥐 새끼의 눈은 출생 후 9일이 지나야 뜬다(미숙아의 징후).[77]
거의 모든 박쥐 종과 마찬가지로 수컷은 암컷의 육아를 돕지 않는다.[78] 새끼는 이유될 때까지 어미와 함께 지낸다. 이유 기간은 과에 따라 다르다. 큰박쥐류는 다른 모든 박쥐류와 마찬가지로 수유 기간이 비교적 길다. 새끼는 성체 몸무게의 약 71%에 달할 때까지 수유를 하는데, 이는 박쥐가 아닌 포유류의 경우 성체 몸무게의 40%와 비교된다.[79] ''Micropteropus'' 속의 종들은 7~8주령에 새끼의 이유를 끝내지만, 인도 큰박쥐는 5개월령이 될 때까지 새끼의 이유를 끝내지 않는다.[80] 매우 이례적으로, 비스마르크 가면 큰박쥐와 다야크 과일박쥐의 두 종의 수컷 개체에서 젖을 생산하는 것이 관찰되었지만, 수컷이 새끼에게 젖을 먹이는 것은 관찰된 적이 없다.[81] 이러한 젖 분비가 기능적이며 수컷이 실제로 새끼에게 젖을 먹이는지, 아니면 스트레스 또는 영양실조의 결과인지는 불분명하다.[82]
3. 4. 천적
큰박쥐류는 특히 섬에 서식하는 종의 경우 토착 포식자가 거의 없다. 외래 포식자로는 들고양이와 쥐가 있다. 맹그로브왕도마뱀은 일부 큰박쥐 종에게는 토착 포식자이지만, 다른 종에게는 도입종 포식자인데, 나무를 잘 타는 능력을 이용해 큰박쥐를 기회적으로 잡아먹는다.[270] 남방큰머리뱀은 괌에서 외래 포식자로, 마리아나큰박쥐 새끼를 너무 많이 잡아먹어 개체군 모집을 거의 0으로 줄였다. 괌은 이제 마리아나과일박쥐의 싱크로 간주되며, 괌의 개체군은 번식 성공보다는 인근 로타섬에서 이주해 온 박쥐에 의존한다.[271]큰박쥐와 자연적으로 동소적인 포식자에는 악어, 뱀, 큰 도마뱀과 같은 파충류와 매, 독수리, 올빼미와 같은 조류가 있다.[241] 바다악어는 호주 북부에서 악어 위 내용물 분석을 통해 큰박쥐의 포식자로 알려져 있다.[272] 극심한 더위 속에서 회색큰박쥐와 같은 큰박쥐는 물가에서 물을 마시며 더위를 식혀야 하므로 민물악어의 기회적인 포식에 취약해진다.[273]
큰박쥐는 여러 기생충 분류군의 숙주이다. 알려진 기생충에는 박쥐파리과와 흡혈파리과 ("박쥐파리"),[274][275] 그리고 모낭충 속의 진드기가 있다.[276] 혈구충과과의 혈액 기생충과 톡소카리과의 장 선충도 큰박쥐 종에 영향을 미친다.[226][277]
3. 5. 종자 살포
큰박쥐류는 종자살포에 중요한 역할을 한다. 오랜 진화의 결과로, 일부 식물은 박쥐의 감각에 맞는 특징, 즉 강한 향이 나고, 밝은 색을 띠며, 잎사귀에서 눈에 잘 띄는 위치에 있는 열매를 진화시켰다. 열매의 밝은 색깔과 위치는 큰박쥐류가 시각적 단서에 의존하고, 덤불 속을 이동하는 데 어려움을 겪는다는 것을 반영할 수 있다.[98] 40종이 넘는 무화과 종의 열매를 조사한 연구에 따르면, 단 한 종의 무화과만이 조류와 큰박쥐류 모두에게 섭취되었고, 대부분의 종은 조류 또는 큰박쥐류 중 하나에게만 섭취되었다.[98] 조류가 섭취하는 무화과는 붉은색이나 주황색인 경우가 많지만, 큰박쥐류가 섭취하는 무화과는 종종 노란색이나 녹색이다.[98] 빠른 장 통과 시간으로 인해 대부분의 종자는 섭취 후 곧 배설되지만, 일부 종자는 12시간 이상 장내에 머물 수 있다. 이는 큰박쥐류가 모수로부터 멀리 종자를 퍼뜨릴 수 있는 능력을 높인다.[99] 고도로 이동성이 높은 과일 섭취 동물로서 큰박쥐류는 벌채된 지역에 나무 종자를 퍼뜨림으로써 고립된 산림 파편 사이의 산림을 복원할 수 있다.[100] 이러한 종자 살포 능력은 길이가 4mm 미만인 작은 종자를 가진 식물로 제한되는데, 이보다 큰 종자는 섭취되지 않기 때문이다.[101]4. 진화와 분류
2002년 박쥐목의 계통 분류는 다음과 같다.[334]
{| class="wikitable"
|-
! 박쥐목
|-
|
{| class="wikitable"
|-
| 큰박쥐류
|
{| class="wikitable"
|-
| 대꼬리박쥐과
|-
|
|-
|
{| class="wikitable"
|-
| 위흡혈박쥐과
|-
|
{| class="wikitable"
|-
| 틈새얼굴박쥐
|-
|
|}
|}
|}
|}
|-
|
{| class="wikitable"
|-
| 흡반발박쥐속
|-
|
{| class="wikitable"
|-
| 원반날개박쥐류
|-
|
|}
|}
|-
|
{| class="wikitable"
|-
| 짧은꼬리박쥐
|-
|
{| class="wikitable"
|-
| 불독박쥐
|-
|
|}
|}
|}
|-
|
|}
|}
미토콘드리아 DNA와 핵 DNA에 기반한 아프리카 큰박쥐류의 계통수는 다음과 같다.[185]
{| class="wikitable"
|-
! 큰박쥐과
|-
|
{| class="wikitable"
|-
| 큰박쥐아과
|-
|
{| class="wikitable"
|-
| 관코박쥐아과
|-
|
| Cynopterinaela |
| Eidolinaela |
|}
|-
! Rousettinaela
|-
| Scotonycterinila
|-
|
{| class="wikitable"
|-
| Eonycterinila
|-
|
{| class="wikitable"
|-
| Rousettinila
|-
|
{| class="wikitable"
|-
| Stenonycterinila
|-
|
{| class="wikitable"
|-
|
| Plerotinila |
| Myonycterinila |
|-
| Epomophorinila
|}
|}
|}
|}
|}
|}
큰박쥐과는 1821년 영국의 동물학자 존 에드워드 그레이가 큰박쥐속(''Pteropus'')을 모식종으로 하여 설립하고, 학명을 Pteropidaela로 하였다.[164] 이 과는 현재 사용되지 않는 과일박쥐아목(Fructivorae)에 분류되었으며, 과일박쥐아목에는 Cephalotidaela 과(Cephalotesla 속 포함)가 포함되어 있었다.[164][165] Cephalotesla 속은 현재 검은날개큰박쥐속(''Dobsonia'')의 동종이명으로 간주된다.[167] 1838년 프랑스의 생물학자 샤를 뤼시앵 보나파르트가 수정된 학명 Pteropodidaela를 처음 사용했다.[172]
1875년 동물학자 조지 에드워드 돕슨은 박쥐목을 대박쥐아목(Megachiropterala)과 소박쥐아목(Microchiropterala)으로 나누고, 큰박쥐아목에 큰박쥐과만을 분류했다.[168][173]
칼 폰 린네 시대 이후, 큰박쥐가 영장류에 가깝다는 의견과 "작은박쥐는 땃쥐목에서 진화하고, 큰박쥐는 영장목에서 진화했다"라는 박쥐류 2계통설이 1986년에 제창되었다.[174] 그러나 이후 박쥐는 단계통군임이 명확해졌다.
2001년 연구에서 양익수아목(Yangochiropterala)과 음익수아목(Yinpterochiropterala)이 제창되었고,[180] 이 분류는 널리 지지되고 있다.[176][177][178][179] 2005년 이후 음익수아목은 "Pteropodiformes"라고도 불린다.[172] 음익수아목에는 큰박쥐과, 긴코박쥐과, 긴꼬리박쥐과, 주름잎코박쥐과, 돼지코박쥐과, 발굽박쥐과 등이 포함되며,[180] 발굽박쥐상과(Rhinolophoideala)와 큰박쥐상과(Pteropodoideala)로 구성된다.[181]
덴마크의 포유류학자 크누트 안데르센은 큰박쥐과를 Macroglossinaela, Pteropinaela(Pteropodinaela로 수정됨), Harpyionycterinaela의 3아과로 분류했다.[182] 1995년 연구에서 Macroglossinae아과는 측계통군임이 판명되었고,[183] 이후 Pteropodinaela 아과 내의 Macroglossinila족에 분류되어 긴혀박쥐속과 꽃박쥐속만을 포함하는 것으로 여겨진다.[184][194]
1917년 안데르센의 분류 이후, 1997년에 형태적 특징에 기반하여 6아과와 9족으로 분류되었지만, 2011년 유전학 연구에서 일부 아과는 측계통군으로 밝혀졌다. 2016년 연구에서는 Epomophorinaela 아과가 Rousettinaela 아과에 포함되고, Rousettinaela 아과의 족이 세분화되었으며, 대나무박쥐속은 Eidolinaela 아과로 옮겨졌다.[185]
1984년 아프리카에서 발견된 화석 속인 Propottola에 대해 Propottininaela 아과가 제창되었으나,[186] 이 속은 여우원숭이상과로 분류되었다.[187] 2018년 시점에서 큰박쥐과에는 197종이 분류되어 있으며,[188] 약 3분의 1은 큰박쥐속이 차지한다.[189] 2024년 현재는 202종이 분류되어 있다.[190]
4. 1. 진화 역사
큰박쥐류의 화석 기록은 다른 어떤 박쥐과보다도 가장 불완전하다. 박쥐 골격은 약하기 때문에 화석으로 남기 어렵지만, 큰박쥐류는 일반적으로 가장 크고 튼튼한 골격을 가지고 있음에도 불구하고 가장 불완전한 기록을 가지고 있다. 또한 큰박쥐류가 가장 먼저 갈라져 나온 주요 그룹임에도 불구하고 가장 적게 나타나는 것은 놀라운 일이다.[22] 소수의 큰박쥐류 화석만 발견된 이유를 설명할 수 있는 몇 가지 요인이 있다. 화석이 발견될 수 있는 열대 지역은 유럽과 북미에 비해 표본 채취가 부족하고, 열대 지방에서는 화석화 조건이 좋지 않아 전반적으로 화석 수가 적을 수 있으며, 화석이 형성되더라도 이후의 지질 활동으로 파괴될 수 있다.[23] 큰박쥐류 화석 기록의 98% 이상이 누락된 것으로 추정된다.[24] 화석이 없더라도 계통 발생학적 계산을 이용하여 과의 연대와 분기 시기를 추정할 수 있다. 큰박쥐류는 약 5800만 년 전(Mya)에 상과인 주름박쥐상과(소목인 익수박쥐아목의 다른 모든 과를 포함함)에서 분리되었다.[24] 현존하는 모든 종을 포함하는 큰박쥐류의 현생종군 조상은 약 3100만 년 전에 살았다.[26]4. 2. 생물지리학
큰박쥐과(Pteropodidae)는 생물지리학적 재구성에 근거하여 오스트랄라시아에서 기원했을 가능성이 높다.[17] 다른 생물지리학적 분석에 따르면, 신기니아를 포함한 멜라네시아 제도가 킨옵테리나아과(Cynopterinae)를 제외한 대부분의 큰박쥐 아과의 기원지 후보로 제시되고 있다.[25] 킨옵테리나아과는 6개의 핵 및 미토콘드리아 유전자에 대한 가중 조상 지역 분석 결과에 따르면 순다랜드에서 기원했을 가능성이 높다.[26] 이 지역에서 큰박쥐들은 아시아 대륙과 아프리카를 포함한 다른 지역으로 확산되었다.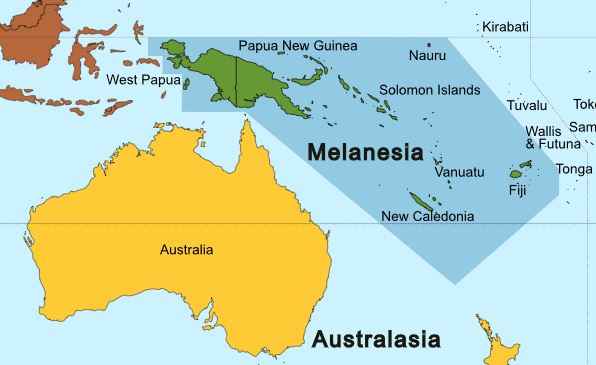
큰박쥐는 적어도 네 번의 별개의 사건을 통해 아프리카에 도착했다. 2016년 연구에 따르면 제안된 네 가지 사건은 (1) *스코토닉테리스속(Scotonycteris)*, (2) *루세투스속(Rousettus)*, (3) 스코토닉테리니족(Scotonycterini), 그리고 (4) 스테노닉테리니족(Stenonycterini), 플레로티니족(Plerotini), 미오닉테리니족(Myonycterini), 에포모포리니족(Epomophorini)을 포함하는 "아프리카 고유 분지"로 나타난다. 큰박쥐가 아프리카에 도착한 시기는 알 수 없지만, 몇몇 족(스코토닉테리니족, 스테노닉테리니족, 플레로티니족, 미오닉테리니족, 에포모포리니족)은 중신세 후기에 존재했다.
큰박쥐가 아프리카에 도착한 경로 또한 알려지지 않았다. 중신세 말에 건조해지기 전에 중동을 통해 도착했을 것이라는 가설이 제기되었다. 반대로, 아프리카와 아라비아 반도를 유라시아에 연결하는 곰포테리움 육교를 통해 도착했을 가능성도 있다. 아프리카 본토에는 서식하지 않는 *큰박쥐속(Pteropus)* (큰박쥐)은 멜라네시아에서 인도양을 가로질러 섬 홉핑을 통해 퍼져나갔을 것이라고 추정된다.[27] 다른 큰박쥐 속들은 몸집이 작고 비행 능력이 제한적이기 때문에 이러한 확산 경로가 적용될 가능성은 낮다.[17]
4. 3. 반향정위
큰박쥐류는 후두를 이용한 반향정위를 할 수 없는 유일한 박쥐과이다. 모든 박쥐의 공통 조상이 반향정위 능력을 가지고 있었는지, 아니면 여러 박쥐 계통이 독립적으로 반향정위 능력을 진화시켰는지는 불분명하며, 이는 "생물학의 큰 과제"라고 불린다.[28] 2017년 박쥐 개체 발생(배아 발달) 연구는 큰박쥐류 배아가 처음에는 반향정위를 하는 작은박쥐류와 유사하게 크고 발달된 달팽이관을 가지고 있지만, 출생 시에는 반향정위를 하지 않는 포유류와 유사한 작은 달팽이관을 가지고 있다는 증거를 제시했다. 이 증거는 후두를 이용한 반향정위가 박쥐 중에서 한 번 진화했고, 큰박쥐류에서 상실되었다는 것을 뒷받침한다.[29]- Rousettus* 속의 큰박쥐류는 혀를 찰칵거리는 소리를 내어 원시적인 반향정위를 할 수 있다.[30] 동굴꿀먹이박쥐(*Eonycteris spelaea*), 작은짧은코과일박쥐(*Cynopterus brachyotis*), 긴혀과일박쥐(*Macroglossus sobrinus*)와 같은 일부 종은 날개를 이용하여 반향정위를 하는 박쥐와 유사한 찰칵 소리를 만드는 것으로 나타났다.[31]
반향정위와 비행은 모두 에너지 소모가 많은 과정이다. 반향정위를 하는 박쥐는 비행에 사용되는 기전과 소리 생성을 결합하여 반향정위의 추가적인 에너지 부담을 줄일 수 있다. 후두를 이용하여 반향정위를 하는 박쥐는 소리 생성을 위해 공기 덩어리를 가압하는 대신, 날갯짓의 아래 방향 힘을 이용하여 공기를 가압하는 것으로 추정되며, 날갯짓과 반향정위를 동기화함으로써 에너지 비용을 절감한다.[33] 큰박쥐류에서 반향정위의 상실은 비행과 반향정위의 분리가 원인일 수 있다.[34] 반향정위를 하는 박쥐에 비해 큰박쥐류의 평균 체구가 더 크다는 점[38]은 더 큰 체구가 비행과 반향정위의 결합을 방해하고, 반향정위를 큰박쥐류에서 유지하기에는 너무 에너지 소모가 많게 만들었음을 시사한다.[34]
대부분의 큰박쥐류는 반향정위를 하지 않고 시각과 후각에 의존하여 이동한다.[64] 큰박쥐류는 머리 앞쪽에 큰 눈을 가지고 있다.[63] 18종의 큰박쥐류 눈을 조사한 연구에 따르면, 일반꽃박쥐 (*Syconycteris australis*)의 눈이 지름 5.03mm로 가장 작았고, 큰과일박쥐 (*Pteropus vampyrus*)의 눈이 지름 12.34mm로 가장 컸다.[61] 큰박쥐류의 홍채는 보통 갈색이지만, *Desmalopex*, *Mirimiri*, *Pteralopex*, 그리고 일부 *Pteropus* 종처럼 붉은색이나 주황색일 수 있다.[62]
밝은 곳에서는 큰박쥐류의 시력이 인간보다 떨어지지만, 어두운 곳에서는 더 우수하다.[63] 일부 *Rousettus*, *Epomophorus*, *Eidolon*, 그리고 *Pteropus* 종의 눈을 조사한 한 연구에 따르면, 처음 세 속은 눈에 낮은 조명 수준에서 시력을 향상시키는 반사 구조인 반사판을 가지고 있는 반면, *Pteropus* 종은 그렇지 않다.[64] 조사된 모든 종은 망막에 간상세포와 추상세포가 모두 있었지만, *Pteropus* 종만이 가장 짧은 파장의 빛을 감지하는 S-추상세포를 가지고 있었다. 옵신의 스펙트럼 조정을 구분할 수 없었기 때문에, *Pteropus* 종의 S-추상세포가 청색광 또는 자외선을 감지하는지 여부는 불분명하다. *Pteropus* 박쥐는 두 종류의 추상세포를 가지고 있는 이색성이다. S-추상세포가 없는 다른 세 속은 일색성으로 색을 볼 수 없다. 모든 속은 매우 높은 간상세포 밀도를 가지고 있어 빛에 대한 감도가 높았는데, 이는 야행성 활동 패턴과 일치한다. *Pteropus*와 *Rousettus*에서 측정된 간상세포 밀도는 제곱밀리미터당 35만~80만 개로, 집쥐, 고양이, 토끼와 같은 다른 야행성 또는 박명성 동물과 같거나 능가한다.[64]
4. 4. 하위 분류
과일박쥐과(Pteropodidae)는 6개의 아과와 46개의 속으로 구성된다.[17][25]'''과일박쥐과 (Pteropodidae)'''
- 애기과일박쥐아과 (Cynopterinae)[25]
- * 난쟁이과일박쥐속 (''Aethalops'')
- * ''Alionycteris''
- * ''Balionycteris''
- * ''Chironax''
- * 개코과일박쥐속 또는 짧은코과일박쥐속 (''Cynopterus'')
- * 다약과일박쥐속 (''Dyacopterus'')
- * ''Haplonycteris''
- * ''Latidens''
- * ''Megaerops''
- * ''Otopteropus''
- * ''Penthetor''
- * 사향과일박쥐속 (''Ptenochirus'')
- * ''Sphaerias''
- * ''Thoopterus''
- 황갈색과일박쥐아과 (Eidolinae)[17]
- * 황갈색과일박쥐속 (''Eidolon'')
- 하피과일박쥐아과 (Harpyionycterinae)[17]
- * ''Aproteles''
- * ''Boneia''
- * 벌거벗은등과일박쥐속 (''Dobsonia'')
- * ''Harpyionycteris''
- 관코박쥐아과 (Nyctimeninae)[25]
- * 관코과일박쥐속 (''Nyctimene'')
- * ''Paranyctimene''
- 큰과일박쥐아과 (Pteropodinae)
- ** ''Melonycteris''[25]
- * 큰과일박쥐족 (Pteropodini)[25]
- ** ''Acerodon''
- ** ''Pteralopex''
- ** 큰박쥐속 (''Pteropus'')
- ** ''Styloctenium''
- 작은과일박쥐아과 (Rousettinae)
- * 새벽과일박쥐족 (Eonycterini)[17]
- ** 새벽과일박쥐속 (''Eonycteris'')
- * 어깨띠과일박쥐족 (Epomophorini)[17][25]
- ** 어깨띠과일박쥐속 (''Epomophorus'')
- ** 어깨띠박쥐속 (''Epomops'')
- ** ''Hypsignathus''
- ** 난쟁이어깨띠과일박쥐속 (''Micropteropus'')
- ** ''Nanonycteris''
- * 족 ''incertae sedis''[35]
- ** ''Pilonycteris''
- * 작은목도리과일박쥐족 (Myonycterini)[17]
- ** ''Megaloglossus''
- ** 작은목도리과일박쥐속 (''Myonycteris'')
- * Plerotini[17]
- ** ''Plerotes''
- * 루셋과일박쥐족 (Rousettini)[17]
- ** 루셋과일박쥐속 (''Rousettus'')
- * Scotonycterini[17]
- ** ''Casinycteris''
- ** ''Scotonycteris''
- * Stenonycterini[17]
- ** ''Stenonycteris''
- ''Incertae sedis''
- * 긴꼬리과일박쥐속 (''Notopteris'')[25]
- * ''Mirimiri''[25]
- * ''Neopteryx''[25]
- * ''Desmalopex''[25]
- * ''Turkanycteris''[36]
- * 긴혀과일박쥐족 (Macroglossini)[25]
- ** 긴혀과일박쥐속 (''Macroglossus'')
- ** 꽃과일박쥐속 (''Syconycteris'')




5. 인간과의 관계
큰박쥐류는 사람에게 여러 가지 영향을 미친다. 식용으로 섭취되기도 하고, 질병을 옮기기도 하며, 문화적인 의미를 지니기도 한다.
큰박쥐류는 마르부르크바이러스, 에볼라 바이러스, 오스트레일리아박쥐광견병바이러스, 헨드라 바이러스, 니파 바이러스 등 다양한 바이러스를 옮길 수 있다.
큰박쥐는 원주민 문화와 전통에서 중요한 역할을 한다. 오스트레일리아와 파푸아뉴기니의 민속 이야기에 등장하며, 오스트레일리아 원주민 동굴 벽화에도 묘사되어 있다.[143][144][145]
오세아니아 원주민 사회에서는 큰박쥐의 뼈나 털을 무기나 장식품으로 사용했다. 솔로몬 제도에서는 큰박쥐 뼈로 창의 갈고리를 만들었고,[148] 뉴칼레도니아에서는 옥으로 만든 의례용 도끼에 큰박쥐 털로 만든 끈을 장식했다.[146] 인도네시아 아스마트족은 큰박쥐 날개를 전쟁 방패에 그려 넣어 전사들을 보호한다고 믿었다.[147]
큰박쥐의 털이나 이빨은 화폐로 사용되기도 했다. 뉴칼레도니아에서는 큰박쥐 털로 만든 끈이 화폐로 사용되었고,[148] 솔로몬 제도의 마키라 섬에서는 큰박쥐 이빨을 목걸이에 꿰어 화폐로 사용한다.[149]
5. 1. 식용
큰박쥐류는 서식지 전역에서 야생고기로 사냥되어 식용된다. 박쥐는 아시아 전역에서 널리 섭취되며, 인도양 서부와 태평양의 섬들에서도 ''Pteropus'' 종이 집중적으로 사냥된다. ''Pteropus'' 종이 서식하지 않는 아프리카 대륙에서는 황갈색과일박쥐가 그 지역에서 가장 큰 큰박쥐류이기 때문에 선호되는 사냥 대상이다.[122]괌에서는 마리아나과일박쥐를 섭취하면 지역 주민들이 신경독소인 베타-메틸아미노-L-알라닌(BMAA)에 노출되어 나중에 신경변성 질환으로 이어질 수 있다. BMAA는 과일박쥐를 섭취하는 사람들에게 특히 생물농축될 수 있으며, 과일박쥐는 소철 열매를 먹으면서 BMAA에 노출된다.[123][124][125]
5. 2. 질병 매개체
과일박쥐류는 사람과 가축에게 치명적일 수 있는 여러 질병의 숙주가 될 수 있지만, 과일박쥐 자신은 질병에 감염된 징후를 보이지 않는 것으로 알려져 있다.[332]2001년부터 2년간 아프리카 중·서부의 3종(망치머리박쥐, 프란켓어깨장식과일박쥐, 작은목걸이과일박쥐) 과일박쥐류가 에볼라 바이러스를 가지고 있는지 테스트했고, 양성 반응을 보였으나 증상은 나타내지 않았다.[332] 이는 이들 박쥐가 바이러스의 숙주가 될 수 있음을 의미한다. 2007년에는 전염성이 있는 마르부르크 바이러스가 이집트과일박쥐 표본에서 발견되어, 이 종이 위험한 바이러스의 숙주가 될 수 있다는 의심을 확정시켰다.[333]

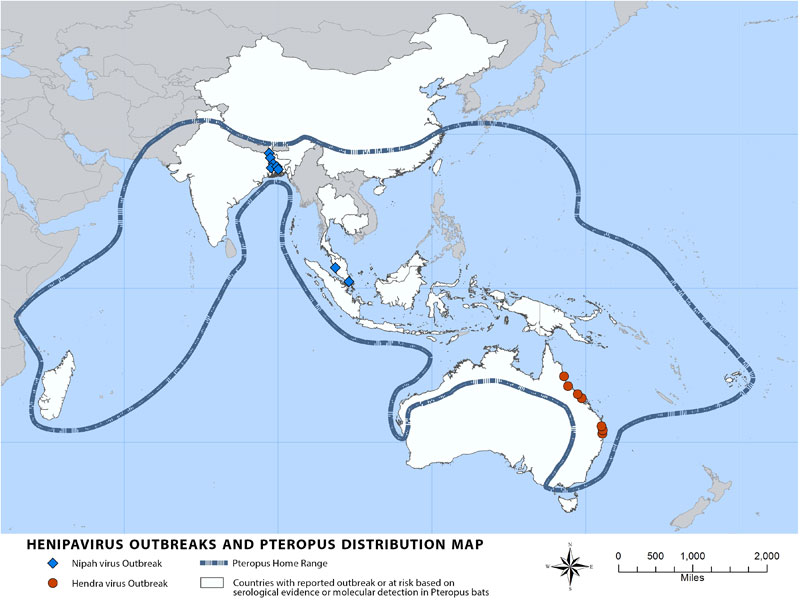
큰박쥐류는 에볼라 바이러스(EBOV)와 ''마르부르크바이러스''를 포함한 필로바이러스를 보유할 수 있으며,[128] 마르부르크 바이러스 질환을 일으키는 ''마르부르크바이러스''는 이집트과일박쥐 한 종에서 확인되었다. 이 질병은 드물지만, 발생 시 치사율은 최대 88%에 달할 수 있다.[128][126] 1967년 독일 마르부르크, 프랑크푸르트, 세르비아 벨그라드에서 처음 인지되었으며,[126] 31명이 감염되어 7명이 사망했다.[127] 이 발병은 우간다산 베르벳원숭이를 이용한 실험실 작업으로 추적되었다.[126] 이 바이러스는 박쥐 숙주(주로 이집트과일박쥐가 서식하는 광산이나 동굴)에서 사람에게 전염될 수 있으며, 감염된 체액(혈액, 정액 등)과의 접촉을 통해 사람 간에 전염될 수 있다.[126] 미국 질병통제예방센터는 1967년부터 2014년까지 총 601건의 마르부르크 바이러스 질환 확진 사례를 보고했으며, 그중 373명이 사망했다(전체 사망률 62%).[127]
EBOV 양성 반응을 보인 종에는 프랑케 칼라과일박쥐, 망치머리과일박쥐, 작은 칼라과일박쥐가 있다. 감비아칼라과일박쥐, 피터스난쟁이칼라과일박쥐, 벨트캄프난쟁이칼라과일박쥐, 레슈노과일박쥐, 이집트과일박쥐에서 EBOV에 대한 항체가 발견되었다.[128] 인간이 에볼라 바이러스에 감염되는 경로의 대부분은 알려져 있지 않지만, 과학자들은 감염된 동물(큰박쥐, 비인간 영장류 등)과의 접촉을 통해 감염된다고 추측한다.[129] 큰박쥐류는 에볼라 바이러스의 자연 숙주로 추정되지만, 확실하게 밝혀지지 않았다.[130] 긴손가락큰발톱박쥐는 2019년에 바이러스 게놈의 5분의 1을 보유하고 있는 것으로 밝혀졌지만, 실제 바이러스에 대해서는 양성 반응을 보이지 않았다.[131] 2013~2016년 유행 기간 동안 여러 서아프리카 국가들은 부시미트(큰박쥐류 포함)를 금지하거나 경고를 발령했으며, 많은 금지 조치는 이후 해제되었다.[132]
''Pteropus'' 종 큰박쥐류는 오스트레일리아박쥐광견병바이러스를 전염시킬 수 있는데, 이 바이러스는 광견병 바이러스와 함께 광견병을 유발한다. 1996년에 처음 확인되었으며, 인간에게 전염되는 경우는 매우 드물다. 감염된 동물의 물림, 할큄, 타액이 점막이나 열린 상처에 닿아 전염될 수 있지만, 혈액, 소변, 배설물에 노출되어서는 감염되지 않는다. 1994년 이후 퀸즐랜드에서 3건의 감염 사례가 보고되었으며, 모두 치명적이었다.[133]
큰박쥐류는 헨드라 바이러스, 니파 바이러스와 같은 헨파바이러스의 숙주이기도 하다. 헨드라 바이러스는 1994년에 처음 확인되었으며, 인간에게 발생하는 경우는 드물다. 1994년부터 2013년까지 7건의 인간 헨드라 바이러스 감염 사례가 보고되었으며, 그중 4건은 치명적이었다. 인간 감염의 주요 경로는 큰박쥐의 소변과 접촉한 말과의 접촉을 통해 이루어진다.[134] 큰박쥐와 인간 간의 직접 전염 사례는 보고된 바 없다.[135] 2012년부터 말에 대한 백신이 사용 가능해져 감염 및 전염 가능성을 줄였다.[136]
니파 바이러스는 1998년 말레이시아에서 처음 확인되었다. 1998년 이후 말레이시아, 싱가포르, 인도, 방글라데시에서 여러 차례 발생이 있었으며, 100명 이상의 사망자가 발생했다. 인도 케랄라에서 발생한 2018년 발생은 19명의 인간 감염자를 발생시켰고, 그중 17명이 사망했다.[137] 전체 치사율은 40~75%이다. 인간은 큰박쥐 또는 그 체액과의 직접 접촉, 돼지와 같은 중간 숙주에 대한 노출, 감염된 사람과의 접촉을 통해 감염될 수 있다.[138] 2014년 연구에 따르면, 니파 바이러스 발생은 큰박쥐가 선호하는 지역에서 더 많이 발생할 가능성이 높지만, "큰박쥐 자체의 존재는 니파 바이러스 감염의 위험 요소로 간주되지 않는다". 대추야자 수액 섭취가 중요한 전염 경로이며, 채취 용기에 대나무 치마를 사용하면 박쥐 소변으로 인한 오염 위험을 줄일 수 있다.[139]
큰박쥐류는 메냉글바이러스[140]와 넬슨베이바이러스[141]와 같이 치명적이지 않은 질병을 전염시킬 수 있다. 이러한 바이러스는 인간에게 거의 영향을 미치지 않으며, 보고된 사례는 거의 없다.[140][141] 큰박쥐류는 코로나바이러스의 매개체로 의심되지 않는다.[142]
5. 3. 문화
큰박쥐류, 특히 큰박쥐는 원주민 문화와 전통에 등장한다. 오스트레일리아와 파푸아뉴기니의 민속 이야기에도 큰박쥐가 등장한다.[143][144] 오스트레일리아 원주민 동굴 벽화에도 큰박쥐가 그려져 있으며, 몇몇 벽화는 현재까지 남아 있다.[145]오세아니아의 원주민 사회에서는 큰박쥐의 일부를 기능적이고 의례적인 무기로 사용했다. 솔로몬 제도에서는 큰박쥐의 뼈로 창에 사용할 갈고리를 만들었다.[148] 뉴칼레도니아에서는 옥으로 만든 의례용 도끼에 큰박쥐 털로 만든 끈을 장식했다.[146] 인도네시아 아스마트족의 전쟁 방패에는 큰박쥐 날개가 그려져 있었는데, 그들은 날개가 전사들을 보호해 준다고 믿었다.[147]
큰박쥐 부산물을 화폐로 사용했다는 현대적이고 역사적인 언급이 있다. 뉴칼레도니아에서는 한때 큰박쥐 털로 만든 끈이 화폐로 사용되었다.[148] 솔로몬 제도 마키라 섬에서는 원주민들이 여전히 큰박쥐를 사냥하여 이빨과 고기를 얻는다. 견치는 목걸이에 꿰어 화폐로 사용된다.[149] 큰박쥐속 큰박쥐(''Pteropus tonganus'')의 이빨은 구멍을 뚫을 만큼 충분히 크기 때문에 특히 귀하게 여겨진다. 마키라 큰박쥐(''Pteropus cognatus'')도 이빨이 더 작음에도 불구하고 사냥된다.
큰박쥐 이빨을 화폐로 사용하는 것을 막는 것은 종에게 해로울 수 있으며, Lavery와 Fasi는 "중요한 문화적 자원을 제공하는 종은 매우 귀중하게 여겨질 수 있다"고 언급했다. 문화적 화폐를 보존하기 위해 큰박쥐의 지속 가능한 사냥을 강조하는 것이 문화적 화폐를 포기하도록 권장하는 것보다 더 효과적일 수 있다. 큰박쥐를 더 이상 이빨 때문에 사냥하지 않더라도, 여전히 고기 때문에 죽일 것이기 때문에, 문화적 가치를 유지하는 것이 지속 가능한 사냥 관행을 장려할 수 있다. Lavery는 "그들의 이빨이 문화적으로 매우 귀중하다는 것은 부정적이 아니라 긍정적이다. 박쥐 사냥 관행을 반드시 중단해야 하는 것은 아니며, 지속 가능하게 관리되어야 한다"고 말했다.[149]
6. 보존
2014년 기준으로, 세계자연보전연맹(IUCN)은 모든 큰박쥐 종의 4분의 1을 위협받는 종으로 평가했는데, 여기에는 멸종위기종, 멸종 위기에 처한 종, 취약종이 포함된다.[160] 큰박쥐는 식량과 약용으로 사냥되기 때문에 인간에 의해 상당히 위협받고 있다. 또한, 특히 과일 생산에 대한 실제 또는 인식된 농업 피해를 줄이기 위해 도태되기도 한다.[151]
2019년 기준으로 IUCN이 평가한 187종의 큰박쥐 상태별 분류는 다음과 같다.[152]

큰박쥐류는 인간에 의한 서식지 파괴로 위협받고 있다. 산림 벌채는 중요한 휴식처 서식지를 잃게 만든다. 또한 토착 과일 나무가 베어지는 것으로 인해 먹이 자원의 손실로 이어진다. 서식지 손실과 그에 따른 도시화는 새로운 도로 건설로 이어져 과도한 포획을 위해 큰박쥐 군집에 더 쉽게 접근할 수 있게 한다. 산림 벌채를 통한 서식지 손실은 산림이 파편화되어 태풍 강풍의 피해에 더 취약해짐으로써 자연적인 위협을 더욱 악화시킨다.[73] 동굴에 서식하는 큰박쥐류는 구아노 채굴, 광물 채굴, 동굴 관광 등 인간의 방해로 위협받고 있다.[73]
큰박쥐류는 의도적이든 우연적이든 인간에 의해 죽임을 당하기도 한다. 모든 큰박쥐류 종의 절반이 식용으로 사냥되는 반면, 곤충을 먹는 종은 8%에 불과하다.[153] 작물에 대한 피해로 인식되는 인간의 박해 또한 큰 사망 원인이다. 일부 큰박쥐류는 과일 작물보다 토착 과일 나무를 선호하는 것으로 기록되었지만, 산림 벌채는 먹이 공급을 줄여 과일 작물에 의존하게 만들 수 있다.[73] 이들은 개체 수 감소를 위해 사살되거나, 구타당하거나, 독살된다. 큰박쥐류가 과일을 먹는 것을 막기 위해 사용되는 그물에 우연히 걸려 죽는 경우도 있다.[154] 모리셔스에서는 2014년부터 2016년까지 4만 마리가 넘는 모리셔스큰박쥐가 살처분되어 개체 수가 약 45% 감소한 것으로 추산된다.[155] 큰박쥐류는 감전으로도 죽는다. 호주 과수원 한 곳에서는 8주 만에 2만 1천 마리가 넘는 박쥐가 감전사한 것으로 추산된다.[156] 농부들은 큰박쥐류가 작물을 먹기 전에 죽이기 위해 과일 나무 위에 감전 그물을 설치한다. 그러나 이러한 그물은 작물 손실을 막는 데 효과가 의심스러우며, 그물을 설치한 한 농부는 여전히 1년에 약 45.36조kg의 과일을 큰박쥐류에게 잃었다고 추산했다.[157] 박쥐가 고압선에 부딪히는 등의 우연한 감전사도 발생한다.[158]
기후변화는 큰박쥐류의 사망을 야기하고 종의 지속성에 대한 우려의 원인이 되고 있다. 호주에서 발생한 극심한 폭염은 1994년부터 2008년까지 3만 마리가 넘는 큰박쥐류의 사망을 초래했다. 암컷과 어린 박쥐는 극심한 더위에 가장 취약하며, 이는 개체군의 회복 능력에 영향을 미친다.[159] 여러 종이 저지대 환초에 서식하기 때문에 큰박쥐류는 기후변화와 관련된 해수면 상승으로 위협받고 있다.[102]
많은 종이 단일 섬에 고유하기 때문에 태풍과 같은 불규칙적인 사건에 취약하다. 1979년 태풍으로 인해 로드리게스큰박쥐의 남은 개체수가 절반으로 줄었다. 태풍은 간접적인 사망률도 야기한다. 태풍으로 인해 나무의 잎이 떨어지면, 큰박쥐가 더 눈에 잘 띄게 되어 사람들에게 더 쉽게 사냥당하게 된다. 큰 폭풍 후에는 박쥐의 먹이 자원이 부족해지고, 큰박쥐는 땅에서 떨어진 과일을 먹는 것과 같이 더 위험한 먹이 탐색 전략을 사용하게 된다. 그곳에서 그들은 집고양이, 개, 돼지에게 더 취약해진다.[160] 많은 큰박쥐 종이 지진 활동이 활발한 불의 고리에 위치해 있기 때문에 화산 폭발의 위협도 받고 있다. 멸종 위기에 처한 마리아나큰박쥐를 포함한 큰박쥐[117][161]는 2003년부터 시작된 일련의 폭발 이후 아나타한 섬에서 거의 멸종되었다.[162]
참조
[1]
서적
Classification of mammals: above the species level
Columbia University Press
[2]
저널
On the natural arrangement of vertebrose animals
http://www.rhinoreso[...]
[3]
저널
The Families and Genera of Bats
https://biodiversity[...]
[4]
저널
A moveable face: deconstructing the Microchiroptera and a new classification of extant bats
[5]
웹사이트
Definition of PTEROPUS
https://www.merriam-[...]
2019-05-22
[6]
서적
Taxonomy of Australian Mammals
CSIRO Publishing
[7]
저널
Conspectus of the suborders, families, and genera of Chiroptera arranged according to their natural affinities
https://biodiversity[...]
[8]
저널
Phylogenomic analyses of bat subordinal relationships based on transcriptome data
[9]
저널
Phylogenomic Analyses Elucidate the Evolutionary Relationships of Bats
[10]
저널
Characterization of the mitochondrial genome of ''Rousettus'' leschenaulti
[11]
저널
A Molecular Phylogeny for Bats Illuminates Biogeography and the Fossil Record
[12]
저널
Integrated fossil and molecular data reconstruct bat echolocation
[13]
서적
Mammal Teeth: Origin, Evolution, and Diversity
JHU Press
[14]
저널
A phylogeny of megachiropteran bats (Mammalia: Chiroptera: Pteropodidae) based on direct optimization analysis of one nuclear and four mitochondrial genes
[15]
저널
A Phylogeny of Indo-West Pacific Megachiroptera Based on Ribosomal DNA
[16]
저널
Taxonomy and biogeography of African fruit bats (Mammalia, Megachiroptera). 5. The genera Lissonycteris Andersen, 1912, Myonycteris Matschie, 1899 and Megaloglossus Pagenstecher, 1885; general remarks and conclusions; annex: key to all species
[17]
저널
The Evolutionary History of the African Fruit Bats (Chiroptera: Pteropodidae)
https://www.research[...]
[18]
저널
Macroscelidea, Insectivora and Chiroptera from the Miocene of east Africa
https://palaeoverteb[...]
[19]
저널
Fossil lemurs from Egypt and Kenya suggest an African origin for Madagascar's aye-aye
[20]
저널
How many species of mammals are there?
[21]
웹사이트
Taxonomy=Pteropus
https://www.iucnredl[...]
2019-05-19
[22]
저널
Quantifying the completeness of the bat fossil record
https://onlinelibrar[...]
Paleobiology Database
2019-03-25
[23]
저널
Global Completeness of the Bat Fossil Record
[24]
저널
A Molecular Phylogeny for Bats Illuminates Biogeography and the Fossil Record
https://courses.cit.[...]
[25]
저널
Evolutionary relationships of the old world fruit bats (Chiroptera, Pteropodidae): Another star phylogeny?
[26]
저널
The phylogenetic relationships of cynopterine fruit bats (Chiroptera: Pteropodidae: Cynopterinae)
[27]
저널
Multiple colonisations of the western Indian Ocean by Pteropus fruit bats (Megachiroptera: Pteropodidae): The furthest islands were colonised first
[28]
서적
Bat Bioacoustics
Springer
[29]
저널
Prenatal development supports a single origin of laryngeal echolocation in bats
[30]
저널
Echolocation signal structure in the Megachiropteran bat Rousettus aegyptiacus Geoffroy 1810
2004-12
[31]
저널
Nonecholocating fruit bats produce biosonar clicks with their wings
2014-12
[32]
저널
No cost of echolocation for bats in flight
1991-04
[33]
저널
Respiratory muscle activity in relation to vocalization in flying bats
http://jeb.biologist[...]
1995-01
[34]
서적
Bats: From Evolution to Conservation
Oxford University Press
[35]
저널
Interrogating Phylogenetic Discordance Resolves Deep Splits in the Rapid Radiation of Old World Fruit Bats (Chiroptera: Pteropodidae)
http://dx.doi.org/10[...]
2021-03-04
[36]
논문
Pliocene bats (Chiroptera) from Kanapoi, Turkana Basin, Kenya
2018-04-00
[37]
서적
Mammals of the South-West Pacific & Moluccan Islands
Cornell University Press
[38]
논문
Are Megabats Big?
[39]
서적
Grzimek's Animal Life Encyclopedia Mammals II
Gale
[40]
논문
Roosting Ecology and the Evolution of Pelage Markings in Bats
[41]
논문
Electrophoretic studies of the systematic and biogeographic relationships of the Fijian bat genera Pteropus, Pteralopex, Chaerephon and Notopteris
[42]
서적
Mammalogy
https://books.google[...]
Jones & Bartlett Publishers
2013-12-11
[43]
논문
Phylogenetic Relationships of Mormoopid Bats (Chiroptera: Mormoopidae) Based on Morphological Data
https://zenodo.org/r[...]
[44]
논문
Middle Pleistocene bats (Mammalia: Chiroptera) from the Yarimburgaz Cave in Turkish Thrace (Turkey)
[45]
논문
Results of the Archbold Expeditions No. 48: Pteropodidae (Chiroptera) of the Archbold Collections
http://digitallibrar[...]
[46]
서적
The teeth of mammalian vertebrates
Academic Press
[47]
논문
On the cranial osteology of chiroptera. I. Pteropus (Megachiroptera: Pteropodidae)
[48]
논문
An asymmetric dental formula in a mammal, the Sao Tomé Island fruit bat Myonycteris brachycephala (Mammalia: Megachiroptera)
[49]
논문
Element homology and the evolution of dental formulae in megachiropteran bats (Mammalia: Chiroptera: Pteropodidae)
http://digitallibrar[...]
[50]
논문
Evolution of dental replacement in mammals
https://web.archive.[...]
[51]
서적
Flying Foxes: Fruit and Blossom Bats of Australia
UNSW Press
[52]
서적
Biology of Bats
Academic Press
[53]
서적
Walker's Mammals of the World
JHU Press
[54]
논문
Morphology and Homology of the Chiropteran Calca, with Comments on the Phylogenetic Relationships of ''Archaeopteropus''
[55]
논문
Structural modifications involved in the fore- and hind limb grip of some flying foxes (Chiroptera: Pteropodidae)
[56]
논문
Correlations between structure and function in the design of the bat lung: a morphometric study
http://jeb.biologist[...]
[57]
논문
Flight Physiology of Intermediate-Sized Fruit Bats (Pteropodidae)
http://jeb.biologist[...]
[58]
보고서
Fauna of Australia
https://www.environm[...]
Australian Government Department of the Environment and Energy
[59]
서적
Comparative Anatomy of the Gastrointestinal Tract in Eutheria II
Walter de Gruyter GmbH & Co KG
[60]
논문
The genome sizes of megabats (Chiroptera: Pteropodidae) are remarkably constrained
[61]
논문
Auditory opportunity and visual constraint enabled the evolution of echolocation in bats
[62]
논문
The systematic position of Pteropus leucopterus and its bearing on the monophyly and relationships of Pteropus (Chiroptera: Pteropodidae)
[63]
논문
Vision in Flying-Foxes (Chiroptera:Pteropodidae)
https://books.google[...]
[64]
논문
Cone Photoreceptor Diversity in the Retinas of Fruit Bats (Megachiroptera)
[65]
논문
A choroidal sleight of hand
[66]
논문
From the ultrasonic to the infrared: Molecular evolution and the sensory biology of bats
[67]
논문
Volatile Compounds in Shoulder Gland Secretions of Male Flying Foxes, Genus Pteropus (Pteropodidae, Chiroptera)
https://www.research[...]
[68]
논문
Glandular secretions of male Pteropus (Flying foxes): preliminary chemical comparisons among species
http://digitalcollec[...]
[69]
논문
Diet Shapes the Evolution of the Vertebrate Bitter Taste Receptor Gene Repertoire
[70]
논문
Diurnal behavior and activity budget of the golden-crowned flying fox (''Acerodon jubatus'') in the Subic bay forest reserve area, the Philippines
2017-00-00
[71]
논문
The timing of reproduction in the fruit bat ''Haplonycteris fischeri'' (Pteropodidae): Geographic variation and delayed development
[72]
서적
Walker's Mammals of the World
JHU Press
[73]
보고서
Old World fruit bats: An action plan for their conservation
https://portals.iucn[...]
IUCN
[74]
논문
Analysis of twinning in flying-foxes (Megachiroptera) reveals superfoetation and multiple-paternity
2008
[75]
논문
Observations on the Reproductive Cycle of Cynopterus sphinx sphinx Vahl, 1797 (Chiroptera: Pteropidae)
1974-03-30
[76]
서적
Bats: from evolution to conservation
Oxford University Press
2011
[77]
논문
Size of bats at birth and maternal investment during pregnancy
http://www.bu.edu/ce[...]
[78]
논문
Social Bats: The Males' Perspective
[79]
서적
Reproductive Biology of Bats
https://archive.org/[...]
Academic Press
[80]
서적
Asdell's Patterns of Mammalian Reproduction: A Compendium of Species-specific Data
https://books.google[...]
Cornell University Press
[81]
논문
Galactorrhoea is not lactation
[82]
논문
Male lactation: Why, why not and is it care?
[83]
논문
Vocalizations in the Malagasy Cave-Dwelling Fruit Bat, Eidolon dupreanum: Possible Evidence of Incipient Echolocation?
[84]
잡지
Hammer-headed Fruit Bat
http://www.batcon.or[...]
2019-05-30
[85]
논문
Nyctimene robinsoni (Chiroptera: Pteropodidae)
[86]
논문
Vocal learning in a social mammal: Demonstrated by isolation and playback experiments in bats
2015
[87]
논문
What bats have to say about speech and language
[88]
논문
Crowd vocal learning induces vocal dialects in bats: Playback of conspecifics shapes fundamental frequency usage by pups
2017
[89]
웹사이트
What Bat Quarrels Tell Us About Vocal Learning
https://www.the-scie[...]
2018-01-01
[90]
논문
Food for Sex in Bats Revealed as Producer Males Reproduce with Scrounging Females
[91]
논문
Homosexual Fellatio: Erect Penis Licking between Male Bonin Flying Foxes Pteropus pselaphon
[92]
논문
Fellatio by Fruit Bats Prolongs Copulation Time
[93]
서적
Walker's bats of the world
https://archive.org/[...]
JHU Press
[94]
논문
Food Hardness and Feeding Behavior in Old World Fruit Bats (Pteropodidae)
[95]
논문
Dietary strategies of Old World Fruit Bats (Megachiroptera, Pteropodidae): How do they obtain sufficient protein?
[96]
논문
Molecular Evolution of the Nuclear Factor (Erythroid-Derived 2)-Like 2 Gene Nrf2 in Old World Fruit Bats (Chiroptera: Pteropodidae)
[97]
논문
Ecological morphology and flight in bats (Mammalia: Chiroptera): wing adaptations, flight performance, foraging strategy and echolocation
1987
[98]
논문
Fruit Bats (Chiroptera: Pteropodidae) as Seed Dispersers and Pollinators in a Lowland Malaysian Rain Forest1
[99]
논문
Old World fruit bats can be long-distance seed dispersers through extended retention of viable seeds in the gut
[100]
논문
High-resolution GPS tracking reveals habitat selection and the potential for long-distance seed dispersal by Madagascan flying foxes Pteropus rufus
[101]
논문
Frugivory and seed dispersal by vertebrates in tropical and subtropical Asia: An update
[102]
논문
Taxonomy, distribution, and natural history of flying foxes (Chiroptera, Pteropodidae) in the Mortlock Islands and Chuuk State, Caroline Islands
https://archive.org/[...]
[103]
논문
Impact of Posttyphoon Hunting on Mariana Fruit Bats (Pteropus mariannus)
[104]
논문
Estuarine crocodiles in a tropical coastal floodplain obtain nutrition from terrestrial prey
[105]
비디오
Flying Foxes Vs Freshwater Crocodile
https://www.youtube.[...]
BBC Earth
2015-04-10
[106]
논문
Hidden diversity of Nycteribiidae (Diptera) bat flies from the Malagasy region and insights on host-parasite interactions
[107]
논문
Rates of hematophagous ectoparasite consumption during grooming by an endemic Madagascar fruit bat
[108]
논문
A new species of demodicid mite (Acari: Prostigmata) from Western Australia parasitic on Macroglossus minimus (Chiroptera: Pteropodidae)
http://museum.wa.gov[...]
[109]
논문
The haemosporidian parasites of bats with description of ''Sprattiella'' alectogen. Nov., sp. Nov
[110]
서적
Mammals of Africa
https://www.research[...]
A&C Black
[111]
논문
The Egyptian fruit bat Rousettus aegyptiacus (Chiroptera: Pteropodidae) in the Palaearctic: Geographical variation and taxonomic status
https://digitalcommo[...]
[112]
IUCN
Eidolon helvum
[113]
논문
Recent Surveys of Bats (Mammalia: Chiroptera) from China II. Pteropodidae
[114]
IUCN
Pteropus dasymallus
2017
[115]
IUCN
Pteropus loochoensis
[116]
IUCN
Pteropus pselaphon
2017
[117]
IUCN
Pteropus mariannus
2008
[118]
웹사이트
Taxonomy=Pteropodidae, Land Regions=West and Central Asia, South and Southeast Asia
https://www.iucnredl[...]
2019-08-24
[119]
논문
Are Flying-Foxes Coming to Town? Urbanisation of the Spectacled Flying-Fox (Pteropus conspicillatus) in Australia
[120]
웹사이트
Taxonomy=Pteropodidae, Land Regions=Oceania
https://www.iucnredl[...]
2019-08-24
[121]
서적
Evolution ecology and conservation of Island bats
University of Chicago Press
[122]
논문
Bats as bushmeat: A global review
[123]
논문
Neurotoxic flying foxes as dietary items for the Chamorro people, Marianas Islands
[124]
논문
Dietary exposure to an environmental toxin triggers neurofibrillary tangles and amyloid deposits in the brain
[125]
논문
The emerging science of BMAA: do cyanobacteria contribute to neurodegenerative disease?
[126]
웹사이트
Marburg Haemorrhagic Fever
https://www.afro.who[...]
2019-05-30
[127]
웹사이트
Chronology of Marburg Hemorrhagic Fever Outbreaks
https://www.cdc.gov/[...]
2014-10-09
[128]
논문
Comparative phylogeography of African fruit bats (Chiroptera, Pteropodidae) provide new insights into the outbreak of Ebola virus disease in West Africa, 2014–2016
https://hal.sorbonne[...]
[129]
웹사이트
Ebola (Ebola Virus Disease): Transmission
https://www.cdc.gov/[...]
2019-05-17
[130]
웹사이트
Ebola Reservoir Study
https://www.cdc.gov/[...]
2018-07-09
[131]
웹사이트
This bat species may be the source of the Ebola epidemic that killed more than 11,000 people in West Africa
https://www.science.[...]
2019-01-24
[132]
웹사이트
Post-Ebola, West Africans flock back to bushmeat, with risk
https://www.apnews.c[...]
2016-09-21
[133]
웹사이트
Rabies and Australian bat lyssavirus infection fact sheet
http://www.health.ns[...]
State of New South Wales NSW Ministry of Health 2015
2015-11-30
[134]
웹사이트
Hendra Virus Disease(HeV)
https://www.cdc.gov/[...]
U.S. Department of Health & Human Services
[135]
논문
Disease Risk Perception and Safety Practices: A Survey of Australian Flying Fox Rehabilitators
[136]
웹사이트
The Hendra vaccine
http://www.ava.com.a[...]
The Australian Veterinary Association Ltd (AVA)
2018
[137]
웹사이트
Nipah virus 'under control' in India – but Britain and the world must be alert for signs of infected travellers
https://www.telegrap[...]
Telegraph Media Group Limited 2018
2018-06-12
[138]
웹사이트
Nipah virus
https://www.who.int/[...]
WHO
2018-05-30
[139]
논문
Roosting behaviour and habitat selection of Pteropus giganteus reveal potential links to Nipah virus epidemiology
[140]
논문
Molecular Characterization of Menangle Virus, a novel Paramyxovirus which Infects Pigs, Fruit Bats, and Humans
[141]
논문
Imported Case of Acute Respiratory Tract Infection Associated with a Member of Species Nelson Bay Orthoreovirus
[142]
논문
Bats and their virome: An important source of emerging viruses capable of infecting humans
[143]
서적
Traditional Storytelling Today: An International Sourcebook
Routledge
[144]
서적
One Thousand One Papua New Guinean Nights: Tales from 1972–1985
Masalai Press
[145]
서적
A natural history of Australian Bats: working the night shift
Csiro Publishing
[146]
논문
Strange Kinds of Money
https://books.google[...]
[147]
서적
Continuum Encyclopedia of Native Art: Worldview, Symbolism, and Culture in Africa, Oceania, and North America
A&C Black
[148]
서적
Handbook to the Ethnographical Collections
https://archive.org/[...]
Printed by order of the Trustees
[149]
웹사이트
In Makira, Flying Fox Teeth Are Currency…And That Could Save the Species
http://blogs.discove[...]
Kalmbach Media
2018-06-02
[150]
논문
Buying through your teeth: Traditional currency and conservation of flying foxes Pteropus spp. In Solomon Islands
[151]
서적
Bats in the Anthropocene: Conservation of Bats in a Changing World
Springer
[152]
웹사이트
Taxonomy=Pteropodidae
https://www.iucnredl[...]
2019-05-19
[153]
서적
Bats in the Anthropocene: Conservation of Bats in a Changing World
Springer
[154]
논문
Near threatened? First report of unsuspected human-driven decline factors in the Ryukyu flying fox (Pteropus dasymallus) in Japan
[155]
논문
Can we protect island flying foxes?
[156]
논문
On the intrinsic capacity for increase of Australian flying-foxes (Pteropus spp., Megachiroptera)
[157]
서적
The Biology and Conservation of Australasian Bats
Royal Zoological Society of New South Wales
[158]
웹사이트
Electric wires threaten flying foxes and their new babies
https://www.dailytel[...]
2018-06-26
[159]
논문
Climate change and the effects of temperature extremes on Australian flying-foxes
[160]
논문
The biology of flying foxes of the genus Pteropus: a review
https://www.research[...]
[161]
보고서
Population Assessment of the Mariana Fruit Bat (Pteropus mariannus mariannus) on Anatahan, Sarigan, Guguan, Alamagan, Pagan, Agrihan, Asuncion, and Maug
http://www.cnmijoint[...]
USGS
2019-08-24
[162]
서적
Island Bats: Evolution, Ecology, and Conservation
https://books.google[...]
University of Chicago Press
[163]
서적
Classification of mammals: above the species level
Columbia University Press
[164]
논문
On the natural arrangement of vertebrose animals
http://www.rhinoreso[...]
[165]
논문
翼手類の類縁に関する綜説
[166]
웹사이트
琥珀の中のアリと現代アリ
https://www.oist.jp/[...]
2024-11-13
[167]
논문
The Families and Genera of Bats
https://biodiversity[...]
[168]
논문
A moveable face: deconstructing the Microchiroptera and a new classification of extant bats
[169]
서적
生物学名命名法辞典
平凡社
[170]
웹사이트
Definition of PTEROPUS
https://www.merriam-[...]
2024-11-02
[171]
서적
生物学名命名法辞典
平凡社
[172]
서적
Taxonomy of Australian Mammals
Csiro Publishing
[173]
논문
Conspectus of the suborders, families, and genera of Chiroptera arranged according to their natural affinities
https://biodiversity[...]
[174]
논문
Flying primates? Megabats have the advanced pathway from eye to midbrain
[175]
논문
哺乳類の高次分類群および分類階級の日本語名称の提案について
[176]
논문
Phylogenomic analyses of bat subordinal relationships based on transcriptome data
[177]
논문
Phylogenomic Analyses Elucidate the Evolutionary Relationships of Bats
[178]
논문
Characterization of the mitochondrial genome of ''Rousettus'' leschenaulti
[179]
논문
A Molecular Phylogeny for Bats Illuminates Biogeography and the Fossil Record
[180]
논문
Integrated fossil and molecular data reconstruct bat echolocation
[181]
서적
Mammal Teeth: Origin, Evolution, and Diversity
JHU Press
[182]
논문
A phylogeny of megachiropteran bats (Mammalia: Chiroptera: Pteropodidae) based on direct optimization analysis of one nuclear and four mitochondrial genes
[183]
논문
A Phylogeny of Indo-West Pacific Megachiroptera Based on Ribosomal DNA
[184]
논문
Taxonomy and biogeography of African fruit bats (Mammalia, Megachiroptera). 5. The genera Lissonycteris Andersen, 1912, Myonycteris Matschie, 1899 and Megaloglossus Pagenstecher, 1885; general remarks and conclusions; annex: key to all species
[185]
논문
The Evolutionary History of the African Fruit Bats (Chiroptera: Pteropodidae)
https://www.research[...]
[186]
논문
Macroscelidea, Insectivora and Chiroptera from the Miocene of east Africa
https://palaeoverteb[...]
[187]
논문
Fossil lemurs from Egypt and Kenya suggest an African origin for Madagascar's aye-aye
[188]
논문
How many species of mammals are there?
[189]
웹사이트
Taxonomy=Pteropus
https://www.iucnredl[...]
2024-11-03
[190]
웹사이트
Batnames.org Species List Version 1.6
https://zenodo.org/r[...]
2024-07-20
[191]
논문
Quantifying the completeness of the bat fossil record
https://onlinelibrar[...]
Paleobiology Database
2019-03-25
[192]
논문
Global Completeness of the Bat Fossil Record
[193]
논문
A Molecular Phylogeny for Bats Illuminates Biogeography and the Fossil Record
https://courses.cit.[...]
[194]
논문
Evolutionary relationships of the old world fruit bats (Chiroptera, Pteropodidae): Another star phylogeny?
[195]
논문
The phylogenetic relationships of cynopterine fruit bats (Chiroptera: Pteropodidae: Cynopterinae)
[196]
논문
Multiple colonisations of the western Indian Ocean by Pteropus fruit bats (Megachiroptera: Pteropodidae): The furthest islands were colonised first
[197]
서적
Bat Bioacoustics
Springer
[198]
논문
Prenatal development supports a single origin of laryngeal echolocation in bats
[199]
논문
Echolocation signal structure in the Megachiropteran bat Rousettus aegyptiacus Geoffroy 1810
2004-12
[200]
논문
Nonecholocating fruit bats produce biosonar clicks with their wings
2014-12
[201]
논문
No cost of echolocation for bats in flight
1991-04
[202]
논문
Respiratory muscle activity in relation to vocalization in flying bats
http://jeb.biologist[...]
1995-01
[203]
서적
Bats: From Evolution to Conservation
Oxford University Press
[204]
간행물
세계 哺乳類標準和名目録
https://doi.org/10.1[...]
日本哺乳類学会
[205]
논문
Interrogating Phylogenetic Discordance Resolves Deep Splits in the Rapid Radiation of Old World Fruit Bats (Chiroptera: Pteropodidae)
https://doi.org/10.1[...]
2021-03-04
[206]
논문
Pliocene bats (Chiroptera) from Kanapoi, Turkana Basin, Kenya
2018-04
[207]
서적
Mammals of the South-West Pacific & Moluccan Islands
Cornell University Press
[208]
논문
Are Megabats Big?
[209]
서적
Grzimek's Animal Life Encyclopedia Mammals II
Gale
[210]
논문
Roosting Ecology and the Evolution of Pelage Markings in Bats
[211]
논문
Electrophoretic studies of the systematic and biogeographic relationships of the Fijian bat genera Pteropus, Pteralopex, Chaerephon and Notopteris
[212]
서적
Mammalogy
https://books.google[...]
Jones & Bartlett Publishers
2013-12-11
[213]
논문
Phylogenetic Relationships of Mormoopid Bats (Chiroptera: Mormoopidae) Based on Morphological Data
https://zenodo.org/r[...]
[214]
논문
Middle Pleistocene bats (Mammalia: Chiroptera) from the Yarimburgaz Cave in Turkish Thrace (Turkey)
[215]
논문
Results of the Archbold Expeditions No. 48: Pteropodidae (Chiroptera) of the Archbold Collections
http://digitallibrar[...]
[216]
논문
An asymmetric dental formula in a mammal, the Sao Tomé Island fruit bat Myonycteris brachycephala (Mammalia: Megachiroptera)
[217]
논문
Element homology and the evolution of dental formulae in megachiropteran bats (Mammalia: Chiroptera: Pteropodidae)
http://digitallibrar[...]
[218]
논문
Evolution of dental replacement in mammals
https://pdfs.semanti[...]
[219]
서적
Flying Foxes: Fruit and Blossom Bats of Australia
UNSW Press
[220]
서적
Biology of Bats
Academic Press
[221]
서적
Walker's Mammals of the World
JHU Press
[222]
논문
Morphology and Homology of the Chiropteran Calca, with Comments on the Phylogenetic Relationships of ''Archaeopteropus''
[223]
논문
Structural modifications involved in the fore- and hind limb grip of some flying foxes (Chiroptera: Pteropodidae)
[224]
논문
Correlations between structure and function in the design of the bat lung: a morphometric study
http://jeb.biologist[...]
[225]
논문
Flight Physiology of Intermediate-Sized Fruit Bats (Pteropodidae)
http://jeb.biologist[...]
[226]
보고서
Fauna of Australia
https://www.environm[...]
Australian Government Department of the Environment and Energy
[227]
서적
Comparative Anatomy of the Gastrointestinal Tract in Eutheria II
Walter de Gruyter GmbH & Co KG
[228]
논문
The genome sizes of megabats (Chiroptera: Pteropodidae) are remarkably constrained
[229]
논문
Auditory opportunity and visual constraint enabled the evolution of echolocation in bats
[230]
논문
The systematic position of Pteropus leucopterus and its bearing on the monophyly and relationships of Pteropus (Chiroptera: Pteropodidae)
[231]
논문
Vision in Flying-Foxes (Chiroptera:Pteropodidae)
https://books.google[...]
[232]
논문
Cone Photoreceptor Diversity in the Retinas of Fruit Bats (Megachiroptera)
[233]
논문
A choroidal sleight of hand
[234]
논문
From the ultrasonic to the infrared: Molecular evolution and the sensory biology of bats
[235]
논문
Volatile Compounds in Shoulder Gland Secretions of Male Flying Foxes, Genus Pteropus (Pteropodidae, Chiroptera)
https://www.research[...]
[236]
논문
Glandular secretions of male Pteropus (Flying foxes): preliminary chemical comparisons among species
http://digitalcollec[...]
[237]
논문
Diet Shapes the Evolution of the Vertebrate Bitter Taste Receptor Gene Repertoire
[238]
논문
Diurnal behavior and activity budget of the golden-crowned flying fox (''Acerodon jubatus'') in the Subic bay forest reserve area, the Philippines
2017-00-00
[239]
논문
The timing of reproduction in the fruit bat ''Haplonycteris fischeri'' (Pteropodidae): Geographic variation and delayed development
[240]
서적
Walker's Mammals of the World
JHU Press
[241]
보고서
Old World fruit bats: An action plan for their conservation
https://portals.iucn[...]
IUCN
[242]
논문
Analysis of twinning in flying-foxes (Megachiroptera) reveals superfoetation and multiple-paternity
2008-00-00
[243]
논문
Observations on the Reproductive Cycle of Cynopterus sphinx sphinx Vahl, 1797 (Chiroptera: Pteropidae)
1974-03-30
[244]
서적
Bats: from evolution to conservation
Oxford University Press
2011-00-00
[245]
논문
Size of bats at birth and maternal investment during pregnancy
http://www.bu.edu/ce[...]
[246]
논문
Social Bats: The Males' Perspective
[247]
서적
Reproductive Biology of Bats
https://archive.org/[...]
Academic Press
[248]
서적
Asdell's Patterns of Mammalian Reproduction: A Compendium of Species-specific Data
https://books.google[...]
Cornell University Press
[249]
논문
Galactorrhoea is not lactation
[250]
논문
Male lactation: Why, why not and is it care?
[251]
논문
Vocalizations in the Malagasy Cave-Dwelling Fruit Bat, Eidolon dupreanum: Possible Evidence of Incipient Echolocation?
[252]
잡지
Hammer-headed Fruit Bat
http://www.batcon.or[...]
2019-05-30
[253]
논문
Nyctimene robinsoni (Chiroptera: Pteropodidae)
[254]
논문
Vocal learning in a social mammal: Demonstrated by isolation and playback experiments in bats
2015
[255]
논문
What bats have to say about speech and language
[256]
논문
Crowd vocal learning induces vocal dialects in bats: Playback of conspecifics shapes fundamental frequency usage by pups
2017
[257]
웹사이트
What Bat Quarrels Tell Us About Vocal Learning
https://www.the-scie[...]
2018-01-01
[258]
논문
Food for Sex in Bats Revealed as Producer Males Reproduce with Scrounging Females
[259]
논문
Homosexual Fellatio: Erect Penis Licking between Male Bonin Flying Foxes Pteropus pselaphon
[260]
논문
Fellatio by Fruit Bats Prolongs Copulation Time
[261]
서적
Walker's bats of the world
https://archive.org/[...]
JHU Press
[262]
논문
Food Hardness and Feeding Behavior in Old World Fruit Bats (Pteropodidae)
[263]
논문
Dietary strategies of Old World Fruit Bats (Megachiroptera, Pteropodidae): How do they obtain sufficient protein?
1998
[264]
논문
Molecular Evolution of the Nuclear Factor (Erythroid-Derived 2)-Like 2 Gene Nrf2 in Old World Fruit Bats (Chiroptera: Pteropodidae)
[265]
논문
Ecological morphology and flight in bats (Mammalia: Chiroptera): wing adaptations, flight performance, foraging strategy and echolocation
1987
[266]
논문
Fruit Bats (Chiroptera: Pteropodidae) as Seed Dispersers and Pollinators in a Lowland Malaysian Rain Forest1
[267]
논문
Old World fruit bats can be long-distance seed dispersers through extended retention of viable seeds in the gut
[268]
논문
High-resolution GPS tracking reveals habitat selection and the potential for long-distance seed dispersal by Madagascan flying foxes Pteropus rufus
[269]
논문
Frugivory and seed dispersal by vertebrates in tropical and subtropical Asia: An update
[270]
논문
Taxonomy, distribution, and natural history of flying foxes (Chiroptera, Pteropodidae) in the Mortlock Islands and Chuuk State, Caroline Islands
https://archive.org/[...]
2013
[271]
논문
Impact of Posttyphoon Hunting on Mariana Fruit Bats (Pteropus mariannus)
2006
[272]
논문
Estuarine crocodiles in a tropical coastal floodplain obtain nutrition from terrestrial prey
[273]
영상미디어
Flying Foxes Vs Freshwater Crocodile
https://www.youtube.[...]
BBC Earth
2015-04-10
[274]
논문
Hidden diversity of Nycteribiidae (Diptera) bat flies from the Malagasy region and insights on host-parasite interactions
[275]
논문
Rates of hematophagous ectoparasite consumption during grooming by an endemic Madagascar fruit bat
[276]
논문
A new species of demodicid mite (Acari: Prostigmata) from Western Australia parasitic on Macroglossus minimus (Chiroptera: Pteropodidae)
http://museum.wa.gov[...]
1981
[277]
논문
The haemosporidian parasites of bats with description of ''Sprattiella'' alectogen. Nov., sp. Nov
[278]
서적
Mammals of Africa
https://www.research[...]
A&C Black
2013
[279]
논문
The Egyptian fruit bat Rousettus aegyptiacus (Chiroptera: Pteropodidae) in the Palaearctic: Geographical variation and taxonomic status
https://digitalcommo[...]
[280]
IUCN
Eidolon helvum
[281]
논문
Recent Surveys of Bats (Mammalia: Chiroptera) from China II. Pteropodidae
[282]
IUCN
Pteropus dasymallus
2017
[283]
IUCN
Pteropus loochoensis
[284]
IUCN
Pteropus pselaphon
2017
[285]
IUCN
Pteropus mariannus
2008
[286]
웹사이트
Taxonomy=Pteropodidae, Land Regions=West and Central Asia, South and Southeast Asia
https://www.iucnredl[...]
2019-08-24
[287]
논문
Are Flying-Foxes Coming to Town? Urbanisation of the Spectacled Flying-Fox (Pteropus conspicillatus) in Australia
[288]
웹사이트
Taxonomy=Pteropodidae, Land Regions=Oceania
https://www.iucnredl[...]
2019-08-24
[289]
서적
Evolution ecology and conservation of Island bats
University of Chicago Press
[290]
논문
Bats as bushmeat: A global review
[291]
논문
Neurotoxic flying foxes as dietary items for the Chamorro people, Marianas Islands
[292]
논문
Dietary exposure to an environmental toxin triggers neurofibrillary tangles and amyloid deposits in the brain
[293]
논문
The emerging science of BMAA: do cyanobacteria contribute to neurodegenerative disease?
[294]
웹사이트
Marburg Haemorrhagic Fever
https://www.afro.who[...]
2024-11-05
[295]
웹사이트
Marburg virus disease
https://www.cdc.gov/[...]
2024-11-05
[296]
논문
Comparative phylogeography of African fruit bats (Chiroptera, Pteropodidae) provide new insights into the outbreak of Ebola virus disease in West Africa, 2014–2016
https://hal.sorbonne[...]
[297]
웹사이트
How Ebola Disease Spreads
https://www.cdc.gov/[...]
2024-11-05
[298]
웹사이트
Ebola Reservoir Study
https://archive.cdc.[...]
2024-11-05
[299]
웹사이트
This bat species may be the source of the Ebola epidemic that killed more than 11,000 people in West Africa
https://www.science.[...]
2024-11-05
[300]
웹사이트
Post-Ebola, West Africans flock back to bushmeat, with risk
https://www.apnews.c[...]
2016-09-21
[301]
웹사이트
Rabies and Australian bat lyssavirus infection fact sheet
http://www.health.ns[...]
State of New South Wales NSW Ministry of Health 2015
2024-11-05
[302]
웹사이트
About Hendra Disease
https://www.cdc.gov/[...]
U.S. Department of Health & Human Services
2024-11-05
[303]
논문
Disease Risk Perception and Safety Practices: A Survey of Australian Flying Fox Rehabilitators
[304]
웹사이트
The Hendra vaccine
http://www.ava.com.a[...]
The Australian Veterinary Association Ltd (AVA)
2018
[305]
웹사이트
Nipah virus 'under control' in India – but Britain and the world must be alert for signs of infected travellers
https://www.telegrap[...]
Telegraph Media Group Limited 2018
2024-11-05
[306]
웹사이트
Nipah virus
https://www.who.int/[...]
WHO
2024-11-05
[307]
논문
Roosting behaviour and habitat selection of Pteropus giganteus reveal potential links to Nipah virus epidemiology
2014
[308]
논문
Molecular Characterization of Menangle Virus, a Novel Paramyxovirus which Infects Pigs, Fruit Bats, and Humans
[309]
논문
Imported Case of Acute Respiratory Tract Infection Associated with a Member of Species Nelson Bay Orthoreovirus
[310]
논문
Bats and their virome: An important source of emerging viruses capable of infecting humans
[311]
서적
Traditional Storytelling Today: An International Sourcebook
Routledge
[312]
서적
One Thousand One Papua New Guinean Nights: Tales from 1972–1985
Masalai Press
[313]
서적
A natural history of Australian Bats: working the night shift
Csiro Publishing
[314]
논문
Strange Kinds of Money
https://books.google[...]
[315]
서적
Continuum Encyclopedia of Native Art: Worldview, Symbolism, and Culture in Africa, Oceania, and North America
A&C Black
[316]
서적
Handbook to the Ethnographical Collections
https://archive.org/[...]
Printed by order of the Trustees
[317]
웹사이트
In Makira, Flying Fox Teeth Are Currency…And That Could Save the Species
http://blogs.discove[...]
Kalmbach Media
2018-06-02
[318]
저널
Buying through your teeth: Traditional currency and conservation of flying foxes Pteropus spp. In Solomon Islands
[319]
서적
Bats in the Anthropocene: Conservation of Bats in a Changing World
Springer
[320]
웹사이트
Taxonomy=Pteropodidae
https://www.iucnredl[...]
2024-11-05
[321]
서적
Bats in the Anthropocene: Conservation of Bats in a Changing World
Springer
[322]
저널
Near threatened? First report of unsuspected human-driven decline factors in the Ryukyu flying fox (Pteropus dasymallus) in Japan
[323]
저널
Can we protect island flying foxes?
[324]
저널
On the intrinsic capacity for increase of Australian flying-foxes (Pteropus spp., Megachiroptera)
[325]
서적
The Biology and Conservation of Australasian Bats
Royal Zoological Society of New South Wales
[326]
웹사이트
Electric wires threaten flying foxes and their new babies
https://www.dailytel[...]
2024-11-05
[327]
저널
Climate change and the effects of temperature extremes on Australian flying-foxes
[328]
저널
The biology of flying foxes of the genus Pteropus: a review
https://www.research[...]
[329]
보고서
Population Assessment of the Mariana Fruit Bat (Pteropus mariannus mariannus) on Anatahan, Sarigan, Guguan, Alamagan, Pagan, Agrihan, Asuncion, and Maug
http://www.cnmijoint[...]
USGS
2019-08-24
[330]
서적
Island Bats: Evolution, Ecology, and Conservation
https://books.google[...]
University of Chicago Press
[331]
문서
[332]
뉴스
Deadly Contact
National Geographic
2007-10
[333]
웹인용
Deadly Marburg virus discovered in fruit bats
http://www.msnbc.msn[...]
msnbc
2008-03-11
[334]
저널
A phylogenetic supertree of the bats (Mammalia: Chiroptera)
http://www.molekular[...]
2014-04-03
[335]
저널
The Evolutionary History of the African Fruit Bats (Chiroptera: Pteropodidae)
https://www.research[...]
[336]
저널
Comparative phylogeography of African fruit bats (Chiroptera, Pteropodidae) provide new insights into the outbreak of Ebola virus disease in West Africa, 2014–2016
https://hal.sorbonne[...]
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com