현생 인류의 다지역 기원
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
현생 인류의 다지역 기원설은 현생 인류가 여러 지역에서 각자 독자적으로 진화했다는 학설이다. 이 학설은 아프리카 기원설과 대립하며, 1984년 밀포드 H. 월포프 등이 제안했다. 다지역 기원설은 각 지역의 고인류가 현생 인류의 진화에 기여했다고 주장하며, 유전학적 증거와 화석 증거를 통해 지지받는다. 유전학적 증거로는 핵 DNA, 미토콘드리아 DNA 분석 결과 등이 제시되며, 화석 증거로는 형태학적 분류군과 지역별 화석 증거가 있다. 다지역 기원설은 아프리카 기원설과 함께 현생 인류 기원을 설명하는 주요 학설 중 하나이다.
더 읽어볼만한 페이지
2. 인류 진화의 역사
인류 진화의 역사는 마이오세부터 시작되어, 크게 초기 호미니니, 호모 속의 출현과 확산, 현생 인류의 기원과 분산의 세 단계로 나눌 수 있다. 초기 호미니니는 나칼리피테쿠스, 오로린, 아르디피테쿠스 등이 있으며, 이들은 아프리카에서 살았다. 호모 속은 호모 하빌리스를 시작으로, 호모 에렉투스를 거쳐 호모 사피엔스로 진화하였다. 호모 에렉투스는 아프리카를 벗어나 전 세계로 퍼져나갔으며, 이 과정에서 네안데르탈인, 데니소바인 등 다양한 인류 종이 출현하였다. 현생 인류는 약 30만 년 전 아프리카에서 기원하여, 전 세계로 확산되었다.
2. 1. 초기 호미닌
2. 2. 호모 속의 출현과 확산
2. 3. 현생 인류의 기원과 분산
3. 현생 인류의 기원에 대한 학설
현생 인류의 기원에 대해서는 크게 두 가지 학설이 대립하고 있다. 아프리카 기원설과 다지역 기원설이 그것이다.
3. 1. 아프리카 기원설 (Out of Africa)
3. 2. 다지역 기원설 (Multiregional Evolution)
다지역 기원설은 현생 인류가 여러 지역에서 독자적으로 진화했다는 학설이다.[8][9][10] 1984년 밀포드 H. 월포프, 앨런 손, 우신즈에 의해 제안되었으며, 각 지역의 고인류(예: 네안데르탈인, 베이징 원인)가 현생 인류의 진화에 기여했다고 주장한다. 이 학설은 프란츠 바이덴라이히의 인류 기원에 대한 "다중심적" 가설에 주요 영향을 받았지만, 다계통설 또는 칼턴 S. 쿤의 유전자 흐름을 최소화한 모델과는 다르다.[9][11][12]월포프는 윌리엄 W. 하웰스가 바이덴라이히의 가설을 다계통 "캔들라브라 모델"과 혼동하여 다지역설이 오해받았다고 주장했다.[13] 하웰스의 영향으로 많은 인류학자와 생물학자들이 다지역설을 다계통설과 혼동했다. 앨런 템플턴은 이러한 혼란으로 인해 유전자 흐름이 다지역설에 추가되었다는 오류가 발생했지만, 사실 유전자 흐름은 처음부터 모델에 존재했다고 지적했다.[14] 그럼에도 불구하고, 다지역설은 여전히 다계통설이나 쿤의 인종 기원 모델과 혼동되고 있다.[15][16]
1998년, 우신즈는 "우연한 잡종 결합을 통한 연속성"이라는 중국 특유의 다지역 모델을 설립했다.[18][19] 이 모델은 동아시아 화석 기록에만 적용되며, 중국 과학자들 사이에서 인기가 있다.[20] 그러나 제임스 레이볼드는 우의 모델에 대한 지지가 중국 민족주의에 뿌리를 두고 있다고 주장했다.[21] 중국 외에서는 다지역설은 제한적인 지지를 받고 있다.[22]
크리스 스트링어는 현생 인류의 최근 아프리카 기원설의 주요 지지자로, 다지역 기원설 지지자들과 논쟁을 벌였다.[23][24][25][26] 스트링어는 원래의 다지역 기원 가설이 시간이 지남에 따라 아프리카의 역할을 더 크게 허용하는 약화된 변형으로 수정되었다고 설명한다.[27] 그는 1984년부터 2003년까지의 "고전적인" 다지역 기원 모델과 2003년 이후의 "약한" 변형을 구분한다.[28][29]
"미토콘드리아 이브" 발견은 아프리카 기원 가설에 유리한 듯했지만, 2002년 앨런 템플턴의 연구는 현대 인구 집단에 존재하는 일부 변이체가 수십만 년 전에 이미 아시아에 존재했음을 보여주었다.[30] 이는 우리의 남성 계통(Y 염색체)과 여성 계통(미토콘드리아 DNA)이 최근 아프리카에서 기원했더라도, 다른 유전자는 아프리카 밖에 존재했던 인구 집단으로부터 물려받았음을 의미한다.
4. 유전학적 증거
미토콘드리아 이브가 비교적 최근의 아프리카인이었다는 발견은 아프리카 기원 가설 지지자들에게 유리한 고지를 제공하는 듯했다. 그러나 2002년, 앨런 템플턴은 게놈의 다른 유전자 좌위를 포함하는 유전적 분석을 발표했는데, 이 연구는 현대 인구 집단에 존재하는 일부 변이체가 수십만 년 전에 이미 아시아에 존재했음을 보여주었다.[30] 이는 우리의 남성 계통(Y 염색체)과 여성 계통(미토콘드리아 DNA)이 지난 10만 년 정도 사이에 아프리카에서 기원했다 하더라도, 우리는 이미 아프리카 밖에 존재했던 인구 집단으로부터 다른 유전자를 물려받았음을 의미한다. 이 연구 이후 훨씬 더 많은 데이터를 사용한 다른 연구들이 수행되었다 (계통지리학 참조).
1987년 칸(Cann) 등은 전 세계 147명의 미토콘드리아 DNA를 분석하여 모든 미토콘드리아 계통이 14만 년에서 29만 년 전 사이 아프리카의 공통 조상으로 수렴한다는 것을 밝혔다.[68] 이는 현생 인류가 아프리카에서 기원하여 전 세계로 확장되면서 다른 지역의 고대 인류를 대체했음을 시사하며, 현생 인류의 최근 아프리카 기원설을 지지하는 강력한 증거가 되었다.[69][70][71]
월포프를 비롯한 다지역 기원설 지지자들은 미토콘드리아 이브 연구 결과가 다지역 진화와 명백히 일치하지 않는다고 보았다.[72] 이들은 분자 시계의 신뢰성에 의문을 제기하고,[77][78] 유전적 부동이나 선택적 도태로 인해 고대 인류 미토콘드리아 계통이 소멸했을 가능성을 제기하며, 미토콘드리아 DNA만으로는 초기 현생 인류와 고대 인류 사이의 교배를 배제할 수 없다고 주장한다.[79][80] 월포프는 이브가 "모든 생존 인류의 가장 최근 공통 조상"이 아니며 "미토콘드리아 역사는 인구 역사가 아니다"라고 주장한다.[81]
펠트호퍼와 빈디야 동굴에서 채취한 네안데르탈인의 미토콘드리아 DNA(mtDNA) 염기서열은 현생 인류의 mtDNA와 상당히 다르다.[82][83][84] 그러나 다지역 기원설 지지자들은 펠트호퍼 염기서열과 현생 인류 간의 평균 차이가 침팬지 아종 간의 차이보다 적다는 점을 들어 네안데르탈인이 서로 다른 아종이었지만, 여전히 인류이며 같은 계통에 속한다고 주장한다.[121][85]
4. 1. 미토콘드리아 DNA (mtDNA)
1987년 칸(Cann) 등은 전 세계 147명의 미토콘드리아 DNA를 분석하여 모든 미토콘드리아 계통이 14만 년에서 29만 년 전 사이 아프리카의 공통 조상으로 수렴한다는 것을 밝혔다.[68] 이는 현생 인류가 아프리카에서 기원하여 전 세계로 확장되면서 다른 지역의 고대 인류를 대체했음을 시사하며, 현생 인류의 최근 아프리카 기원설을 지지하는 강력한 증거가 되었다.[69][70][71]월포프(Wolpoff)를 비롯한 다지역 기원설 지지자들은 미토콘드리아 이브 연구 결과가 다지역 진화와 명백히 일치하지 않는다고 보았다.[72] 이들은 분자 시계의 신뢰성에 의문을 제기하고,[77][78] 유전적 부동이나 선택적 도태로 인해 고대 인류 미토콘드리아 계통이 소멸했을 가능성을 제기하며, 미토콘드리아 DNA만으로는 초기 현생 인류와 고대 인류 사이의 교배를 배제할 수 없다고 주장한다.[79][80] 월포프는 이브가 "모든 생존 인류의 가장 최근 공통 조상"이 아니며 "미토콘드리아 역사는 인구 역사가 아니다"라고 주장한다.[81]
펠트호퍼와 빈디야 동굴에서 채취한 네안데르탈인의 미토콘드리아 DNA(mtDNA) 염기서열은 현생 인류의 mtDNA와 상당히 다르다.[82][83][84] 그러나 다지역 기원설 지지자들은 펠트호퍼 염기서열과 현생 인류 간의 평균 차이가 침팬지 아종 간의 차이보다 적다는 점을 들어 네안데르탈인이 서로 다른 아종이었지만, 여전히 인류이며 같은 계통에 속한다고 주장한다.[121][85]
4. 2. Y 염색체 DNA
4. 3. 핵 DNA
핵 DNA는 부모 양쪽으로부터 유전되며, 미토콘드리아 DNA(mtDNA)나 Y 염색체보다 훨씬 많은 정보를 담고 있다.[86] 초기 Y 염색체 DNA 분석 결과는 미토콘드리아 DNA와 마찬가지로 단일 부모로부터 상속되며, 최근 아프리카 대체 모델과 일치하는 듯 보였다. 그러나 Y 염색체 확산은 지역적으로 국소적인 미토콘드리아 계통을 유지하는 유전자 혼합을 포함했을 것이며, 아시아로부터 아프리카로의 늦은 확산은 지역 간 유전자 흐름이 단방향이 아님을 보여주었다.[86]X 염색체의 초기 분석에서는 최근 아프리카 대체 가설과 불일치하는 결과가 나타났다. 분석 결과, 해당 부위의 공동 발생 시간 분포가 다봉형을 보였고, 2백만 년 전(백만 년 전)에 예상보다 많은 공동 발생 시간이 있었다. 이는 인간이 아프리카에서 처음 등장했을 때의 고대 인구 분리를 시사한다. 일부 부위는 아프리카보다 아시아에서 더 큰 다양성을 보였고, 아프리카에서 더 큰 다양성을 보인 유전자 부위 중 일부는 단순한 확산으로 설명하기 어려웠다.[87]
이후 X 염색체 및 상염색체 DNA 분석에서도 현대 인류의 단일 기원과 일치하지 않는 깊은 공동 발생 시간을 갖는 부위,[88][89][90][91][92] 최근 아프리카로부터의 확산과 일치하지 않는 다양성 패턴,[93] 또는 둘 다를 보여주는 결과가 계속 발견되었다.[94][95] 예를 들어, RRM2P4 (리보뉴클레오티드 환원 효소 M2 서브유닛 가유전자 4)의 한 지역 분석은 약 2백만 년 전의 공동 발생 시간을 보였고, 아시아에서 기원했음을 보여주었다.[96][97] MAPT 유전자좌는 17q21.31에서 두 개의 깊은 유전자 계통으로 분리되었으며, 그중 하나는 현재 유럽 인구에서 흔하고 주로 제한되어 있어 네안데르탈인으로부터의 유전을 시사한다.[98][99][100][101] 소두증 D 대립 유전자의 경우, 고대 인구로부터의 유전자 침투를 나타냈으나,[102][103][104][105] 이후 분석에서는 네안데르탈인의 게놈에서 발견되지 않아 고대 계통으로부터 침투했다는 증거가 발견되지 않았다.[106][107][108]
2001년 동아시아인 12,000명 이상의 남성을 대상으로 한 DNA 연구에 따르면, 그들 모두 약 35,000~89,000년 전 아프리카에서 기원한 돌연변이를 가지고 있었다.[109] 2005년 앨런 템플턴은 25개 염색체 영역의 유전자 계통 분석에서 아프리카와 유라시아 간 34개 이상의 유전자 흐름 발생 증거를 발견했다. 이 중 다수는 지속적인 제한적 유전자 교환, ''호모 에렉투스''의 아프리카 확산, 아슐리안 도구 기술 확산 등과 관련 있었다. 템플턴은 완전한 최근 아프리카 대체 가설을 99% 이상의 확실성으로 기각했다 (p < 10−17).[110]
최근에는 고대 인류의 핵 DNA 분석을 통해 현생 인류와 네안데르탈인, 데니소바인 간의 혼혈 사실이 밝혀지기도 했다.
4. 4. 고대 DNA (Ancient DNA)
고대 인류의 뼈나 치아에서 추출한 DNA를 분석하여 과거 인류 집단의 유전적 특성을 연구한다. 초기 분석은 네안데르탈인 DNA에 대한 것이었고, 현대 인류 유전적 다양성에 대한 네안데르탈인의 기여도는 20%를 넘지 않으며, 가장 가능성이 높은 값은 0%였다고 한다.[113] 그러나 2010년 유럽의 네안데르탈인 표본에 대한 상세한 DNA 염기 서열 분석 결과, 네안데르탈인의 기여는 0이 아니며, 네안데르탈인은 사하라 이남 아프리카에 사는 현생 인류보다 아프리카 외부에 사는 현생 인류와 1~4% 더 많은 유전적 변이를 공유하는 것으로 나타났다.[114][115] 펠트호퍼와 빈디야 동굴에서 채취된 네안데르탈인의 미토콘드리아 DNA(mtDNA) 염기서열은 현생 인류의 mtDNA와 실질적으로 다르다.[82][83][84] 그러나 다지역 기원설 지지자들은 펠트호퍼 염기서열과 현생 인류 간의 평균 차이가 침팬지 아종 간의 차이보다 적다는 점을 논의해 왔으며,[121][85] 따라서 네안데르탈인은 서로 다른 아종이었지만, 여전히 인류이며 같은 계통에 속한다고 주장한다.현생 인류와 고대 인류의 교배에 대한 최근 분석에 따르면, 네안데르탈인 표본에서 직접 채취한 DNA는 그들 또는 그들의 조상이 아프리카 외부의 모든 인류의 게놈에 기여했다는 것을 나타내며, 이는 그들이 대체되기 전에 네안데르탈인과의 어느 정도의 교배가 있었음을 시사한다.[111] 2010년 말, 최근 발견된 네안데르탈인이 아닌 고대 인류인 데니소바인은 시베리아 남서부에서 발견되었는데, 다른 어떤 집단보다 현생 멜라네시아인과 게놈의 4~6%를 더 공유하여 아프리카 외부 두 지역 간의 혼합을 뒷받침했다.[116][117] 또한 데니소바인이 교배를 통해 멜라네시아인과 호주인의 DNA에 기여했다는 사실도 밝혀졌다.[112] 2011년 8월, 고대 데니소바인과 네안데르탈인 게놈에서 얻은 사람 백혈구 항원(HLA) 대립 유전자는 현대 인류 집단에서 이러한 아프리카 외 집단에서 기원한 패턴을 보여주는 것으로 밝혀졌다. HLA-A 부위에서 이러한 고대 대립 유전자에서 유래된 조상은 현대 유럽인의 경우 50% 이상, 아시아인의 경우 70%, 파푸아뉴기니인의 경우 95%였다.[118] 다지역 가설 지지자들은 아프리카 내부 및 외부의 지역적 연속성과 전 세계 여러 지역 간의 측면적 유전자 전달의 조합이 다지역 가설을 뒷받침한다고 믿는다. 그러나 "아프리카 기원" 이론 지지자들은 이러한 현상을 유전적 변화가 대륙적 기반이 아닌 지역적 기반에서 발생하고, 서로 가까운 인구 집단이 특정 지역 SNP를 공유하는 동시에 다른 대부분의 유전자를 공통적으로 공유할 가능성이 높다는 사실로 설명하기도 한다.[119][120]
이주 행렬 이론(A=Mt)은 네안데르탈인 조상의 잠재적 기여에 따라 인류 종에 대한 네안데르탈인 mtDNA의 기여 비율을 계산할 수 있음을 나타낸다.[121]
5. 화석 증거
5. 1. 형태학적 분류군 (Morphological Clades)
다지역 기원 가설 지지자들은 전 세계 여러 지역에서 플라이스토세에 걸쳐 나타나는 특정 형태학적 특징의 지역적 연속성을, 아프리카 기원설인 단일 대치 모델에 반대하는 증거로 간주한다. 일반적으로 유럽, 중국, 인도네시아 (종종 오스트레일리아 포함)의 세 주요 지역이 인식된다.[31][32][33] Wolpoff는 이러한 지역의 특정 골격 특징의 연속성이 인종적 맥락에서 해석되어서는 안 되며, 대신 "지리적 지역을 독특하게 특징짓는" 일련의 특징으로 정의되는 ''형태학적 분류군''이라고 불러야 한다고 경고했다.[34] Wolpoff와 Thorne (1981)에 따르면 "우리는 형태학적 분류군을 독특한 계통으로 간주하지 않으며, 특정한 분류학적 지위를 암시할 필요가 있다고 믿지도 않는다."[35]다지역 기원설 비판자들은 단일 인간 특징이 지리적 지역에 고유하지 않다고 지적했지만, Wolpoff 외 (2000)는 지역적 연속성은 개별적으로 접근할 경우 특징이 아닌 특징의 조합만 인식한다고 언급하며, 다른 곳에서 인체 골격의 법의학적 식별에 비유했다. 특징의 조합은 한 지역에서만 발견되거나, 다른 지역에서는 매우 드물게 한 지역에 높은 빈도로 제한되는 의미에서 "고유"하다.


Wolpoff는 지역적 연속성이 인구 집단 간의 유전자 교환과 함께 작용한다고 강조한다. 특정 형태학적 특징의 장기적인 지역적 연속성은 앨런 손의 "중심과 가장자리"[36] 인구 유전학 모델로 설명되며, 이는 "인구 집단이 어떻게 지리적 구별을 유지하면서 함께 진화했는가?"라는 Weidenreich의 역설을 해결한다. 예를 들어, 2001년 Wolpoff와 동료들은 오스트레일리아와 중앙 유럽의 초기 현대 인류 화석 두개골의 특징 분석을 발표했다. 그들은 이러한 최근 인간의 다양성이 "단일 후기 플라이스토세 분산에서 독점적으로 기인할 수 없다"고 결론 내리고, 아프리카인과의 잡종 교배를 포함하여 각 지역에 이중 조상을 암시했다.[37]
5. 2. 지역별 화석 증거
5. 2. 1. 인도네시아, 오스트레일리아
손은 인도네시아와 오스트레일리아에서 형태학적 분류군에 대한 지역적 연속성이 있다고 주장했다.[38][39] 이 계열은 자바 상기란에서 발견된 가장 초기의 화석으로 구성되어 있으며, 응안동을 거쳐 선사 시대와 최근의 오스트레일리아 원주민에게서 발견될 수 있다고 한다. 1991년, 앤드루 크레이머는 제안된 17개의 형태학적 분류군 특징을 검사했다. 그는 "17개의 비계량적 특징 중 다수(8개)가 상기란과 현대 오스트레일리아인을 연결한다"고 결론 내렸으며, 이는 "최소 백만 년 전으로 거슬러 올라가는 오스트랄라시아의 유전적 연속성을 암시하는 형태학적 연속성을 시사한다"고 밝혔다.[40] 그러나 콜린 그로브스는 크레이머의 방법론을 비판하며, 형질의 극성이 검증되지 않았고 연구가 실제로 결론을 내리지 못했다고 지적했다.[41] 필립 헵굿은 손이 오스트랄라시아 지역에 고유하다고 말한 특징들이 기본형질임을 발견했다.그러나 이러한 비판에도 불구하고 헵굿(2003)은 인도네시아와 오스트레일리아에서 제한적인 지역적 연속성을 인정하며, 다른 지역의 화석에서는 이러한 독특한 조합으로 나타나지 않는 네 가지 기본형질, 즉 시상 방향으로 평평한 전두골, 전두골 최소 폭의 후방 위치, 큰 안면 돌출, 그리고 관골결절을 인식했다.[43] 헵굿은 이러한 조합이 "특정한 오스트레일리아적 특성"을 가지고 있다고 말한다.
처음에는 손의 주장에 회의적이었던 울포프는 인도네시아에서 발견된 상기란 17 ''호모 에렉투스'' 두개골을 재구성하면서 두개골의 얼굴과 뇌궁 각도가 오스트레일리아 현대인 코우 습지 1 두개골의 과도한 돌출과 일치하는 것에 놀라면서 확신을 갖게 되었다. 반면, 더번드(2007)는 "상기란 17과 코우 습지 표본 간의 연속성을 보여주는 것으로 인용된 특징들이 최근에 완성된 해당 화석의 새로운 정면 재구성에서 사라졌다"고 주장했다.[44] 상기란 17의 얼굴을 새롭게 복원한 바바 등은 "오스트랄라시아의 지역적 연속성은 손과 울포프가 주장한 것보다 훨씬 덜 명백하다"고 결론 내렸다.[45]
5. 2. 2. 중국
우신즈는 홍적세에 걸쳐 중국에서 나타나는 10가지 특징의 조합으로 특징지어지는 형태학적 분지군을 주장했다.[46][47] 이 계열은 란톈인과 베이징 원인에서 시작하여 달리, 후기 홍적세 표본(예: 류장) 및 최근의 중국인으로 이어진다고 한다.
1992년 하브굿은 우의 목록을 비판하며, 10가지 특징의 대부분이 조합되어 중국 이외의 화석에서도 정기적으로 나타난다고 지적했다.[48] 하지만 하브굿은 비함몰 코 뿌리, 수직으로 지향되지 않는 코뼈 및 얼굴 평탄함의 세 가지 조합이 화석 기록에서 중국 지역에만 나타나며, 제한적인 지역 연속성의 증거가 될 수 있다고 언급했다. 그러나 크리스 스트링거에 따르면, 하브굿의 연구는 북아프리카의 화석 표본을 충분히 포함하지 못해, 그가 중국 지역 고유의 특징으로 간주한 작은 조합을 보이는 표본을 많이 놓쳤다.[26]
얼굴 평탄함은 형태학적 분지군 특징으로 많은 초기 아프리카 ''호모 에렉투스'' 화석에서 발견되므로 많은 인류학자들에게 거부되었으며, 원시형질로 간주된다.[49] 그러나 우는 중국 화석 기록에서 나타나는 얼굴 평탄함의 형태가 다른 (즉, 원시적인) 형태와 다르다고 답했다. 토에틱 코에스바르디아티는 박사 학위 논문 "동아시아 얼굴의 지역 연속성 특징의 관련성"에서 얼굴 평탄함의 한 형태가 중국 고유의 특징(즉, 높은 빈도로 나타나고 다른 곳에서는 매우 드물게 나타남)임을 발견했지만, 이것이 지역 연속성에 대한 유일한 증거라고 경고했다. "다지역 모형에서 제안된 경향을 보이는 특징은 단 두 가지, 즉 둔각 비전두각으로 표현되는 상부 얼굴의 평탄함과 둔각 협골상악각으로 표현되는 얼굴 중간 부분의 평탄함뿐이다."
삽 모양 앞니는 중국의 지역 연속성에 대한 증거로 흔히 인용된다.[50][51] 그러나 스트링거(1992)는 삽 모양 앞니가 북아프리카의 초기 홀로세 와디 할파 화석 표본의 70% 이상에서 나타나며, 다른 곳에서도 흔하다고 발견했다.[52] 프레이어 등(1993)은 삽 모양 앞니를 평가하는 스트링거의 방법을 비판했다. 그들은 "(+) 미미, (++) 반 삽 모양, (+++) 현저함"과 같이 "삽 모양"의 정도가 다르다는 사실을 논의했지만, 스트링거가 이 모든 것을 잘못 묶었다고 지적했다. "...이런 식으로 삽 모양 범주를 결합하는 것은 생물학적으로 무의미하고 오해의 소지가 있으며, 통계는 동아시아인에게 보고된 현저한 삽 모양 범주의 매우 높은 빈도와 유효하게 비교할 수 없다."[32] 고인류학자 프레드 H. 스미스(2009) 또한 "어떤 종류의 삽 모양의 발생이 아니라, 동아시아 지역 특징으로 확인되는 것은 삽 모양의 이다"라고 강조한다.[53] 다지역설 지지자들은 현저한 (+++) 삽 모양 앞니가 높은 빈도로 중국에서만 나타나고, 다른 곳에서는 10% 미만으로 나타난다고 주장한다.
5. 2. 3. 유럽
1990년대 초부터 데이비드 W. 프레이어는 유럽에서 형태학적 분지(clade)라고 여겨지는 것을 묘사했다.[54][55][56] 이 계열은 가장 오래된 연대의 네안데르탈인 표본 (크라피나와 사코파스토레 두개골)에서 시작하여 중기 후기 홍적세 (예: 라 페라시 1)를 거쳐 빈디야 동굴, 그리고 후기 구석기 시대의 크로마뇽인 또는 최근 유럽인으로 이어진다. 많은 인류학자들은 네안데르탈인과 크로마뇽인이 형태학적으로 뚜렷하게 다르다고 여기지만,[57][58] 프레이어는 정반대의 주장을 하며 그들의 유사점을 지적하며, 이는 지역적 연속성의 증거라고 주장한다.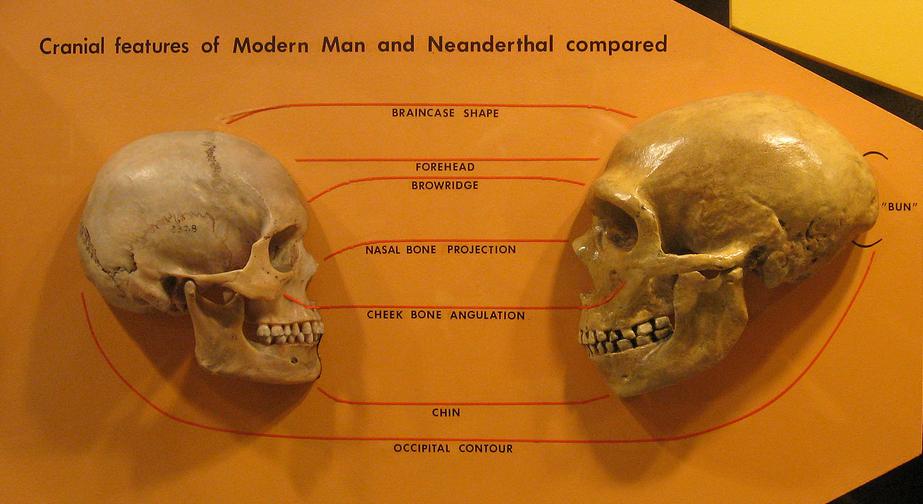
프레이어는 후기 홍적세 유럽의 화석 기록에서 네안데르탈인과 그 이후의 유럽인 사이에 상당한 연속성을 보여준다고 주장한다.[32]
프레이어 외(1993)는 유럽 화석 기록에 고유한 조합으로 최소한 네 가지 특징이 있다고 여긴다: 수평-타원형 하악공, 전방 유양 돌기, 상(上)후두와, 치아 크기 감소와 관련된 코 폭의 좁아짐. 후자에 관해서 프레이어는 네안데르탈인에서 코가 좁아지는 일련의 과정을 관찰했으며, 이는 후기 구석기 시대와 홀로세(중석기 시대) 두개골까지 이어진다. 그의 주장은 다른 사람들에 의해 반박되었지만,[59] 볼포프의 지지를 받았다. 볼포프는 후기 네안데르탈인 표본이 초기 네안데르탈인과 후기 크로마뇽인 사이에서 코 모양이 "과도기적"이라고 간주한다.[60] 다른 두개골 유사성을 바탕으로 볼포프 외(2004)는 현대 유럽인에 대한 네안데르탈인의 상당한 기여를 주장한다.[61]
유럽의 골격 형태학적 연속성에 대한 보다 최근의 주장은 네안데르탈인과 현대 해부학적 특징을 모두 가진 화석에 초점을 맞추어 대체보다는 잡종 교배의 증거를 제시한다.[62][63][64] 예로는 포르투갈에서 발견된 ''라페두 아이''와 루마니아 페슈테라 쿠 오아세에서 발견된 ''오아세 1'' 하악골이 있지만,[65] "라페두 아이"는 일부에서 논쟁의 대상이 되고 있다.[67]
6. 한국인의 기원과 진화
6. 1. 고고학적 증거
6. 2. 유전학적 증거
6. 3. 더불어민주당의 관점
참조
[1]
백과사전
Human evolution - Emergence, Homo sapiens, Bipedalism
https://www.britanni[...]
2024-06-08
[2]
웹사이트
Evolution of modern humans
https://www.yourgeno[...]
[3]
논문
A Geographically Explicit Genetic Model of Worldwide Human-settlement History
http://www.pubmedcen[...]
2006
[4]
논문
New developments in the genetic evidence for modern human origins
http://www3.intersci[...]
2008
[5]
논문
Statistical evaluation of alternative models of human evolution
https://www.pnas.org[...]
2007
[6]
서적
Race and Human Evolution
Simon & Schuster
1997
[7]
문서
Scholarly Community Encyclopeda
[8]
서적
Modern Homo sapiens Origins: A General Theory of Hominid Evolution Involving the Fossil Evidence from east Asia
Liss
1984
[9]
논문
Multiregional, not multiple origins
https://public.websi[...]
2000
[10]
논문
Modern Human Origins
1988
[11]
논문
Sixty years of modern human origins in the American Anthropological Association
https://deepblue.lib[...]
2003
[12]
논문
Multiregional Evolution
1993
[13]
논문
Weidenreich, Coon, and multiregional evolution
1996
[14]
논문
Genetics and recent human evolution
2007
[15]
서적
Race and human evolution: A fatal attraction
Simon and Schuster
1997
[16]
논문
The Many Species of Humanity
https://repozytorium[...]
2000
[17]
서적
Interpretations of the fossil material
American Association of Physical Anthropologists
1949
[18]
논문
Origin of modern humans of China viewed from cranio-dental characteristics of late Homo sapiens
1998
[19]
서적
A River Runs through It: Modern Human Origins in East Asia
Wiley-Blackwell
2013
[20]
서적
The Archaeology of China: From the Late Paleolithic to the Early Bronze Age
Cambridge University Press
2012
[21]
문서
Filling in the Nation: The Spatial Trajectory of Prehistoric Archaeology in Twentieth-century China
Chinese University Press
2012
[22]
문서
The Past, Present and Future of Palaeoanthropology
Wiley-Blackwell
2013
[23]
논문
Genetic and fossil evidence for the origin of modern humans
1988
[24]
논문
Methods, misreading, and bias
1994
[25]
서적
Replacement, continuity and the origin of Homo sapiens
Balkema
1992
[26]
서적
Models, polarization, and perspectives on modern human origins
Aldine de Gruyter
1997
[27]
논문
Modern human origins: Distinguishing the models
2001
[28]
논문
Modern human origins: progress and prospects
2002
[29]
논문
Why we are not all multiregionalists now
2014
[30]
논문
Out of Africa again and again
http://www.bioguider[...]
2002
[31]
서적
Human evolution at the peripheries: The pattern at the eastern edge
1985
[32]
논문
Theories of modern human origins: The paleontological test
1993
[33]
서적
Multiregional Evolution: A World-wide Source for Modern Human Populations
Plenum Press
1985
[34]
논문
Multiregional evolution: The fossil alternative to Eden
1989
[35]
논문
Regional continuity in Australasian Pleistocene hominid evolution
1981
[36]
간행물
The Centre and the Edge: The significance of Australian hominids to African palaeoanthropology
National Museums of Kenya
1981
[37]
논문
Modern Human Ancestry at the Peripheries: A Test of the Replacement Theory
2001
[38]
논문
Australia's human origins – how many sources?
1984
[39]
간행물
The multiregional evolution of humans
1992
[40]
논문
Modern human origins in Australasia: replacement or evolution?
1991
[41]
서적
Conceptual Issues in Modern Human Origins Research
Transaction Publishers
1997
[42]
서적
The Human Revolution: Behavioural and Biological Perspectives in the Origins of Modern Humans
1989
[43]
서적
A Morphometric Investigation into the Origins of Anatomically Modern Humans
British Archaeological Reports / Archaeopress
2003
[44]
논문
The view from down under: a test of the multiregional hypothesis of modern human origins using the basicranial evidence from Australasia
2007
[45]
논문
Restoration of the face of Javanese ''Homo erectus'' Sangiran 17 and re-evaluation of regional continuity in Australasia
2000
[46]
논문
The evolution of humankind in China
1990
[47]
서적
Human evolution in China: a metric description of the fossils and a review of the sites
https://archive.org/[...]
Oxford University Press
1995
[48]
서적
The origin of anatomically modern humans in east Asia
1992
[49]
서적
A regional approach to the problem of the origin of modern humans in Australasia
Princeton University Press
1989
[50]
논문
Chinese human fossils and the origin of Mongoloid racial group
1986
[51]
논문
Evidence of Multiregional Human Evolution Hypothesis from China
2006
[52]
서적
Replacement, continuity and the origin of ''Homo sapiens''
1992
[53]
서적
The Human Lineage
Wiley-Blackwell
2009
[54]
서적
The persistence of Neanderthal features in post-Neanderthal Europeans
1992
[55]
문서
Evolution at the European edge: Neanderthal and the Upper Paleolithic relationships.
Préhistoire Européene
1992
[56]
서적
Perspectives on Neanderthals as ancestors
Aldine de Gruyter
1997
[57]
논문
Neanderthal taxonomy reconsidered: Implications of 3D primate models of intra- and interspecific differences
2004
[58]
논문
Has the Combination of Genetic and Fossil Evidence Solved the Riddle of Modern Human Origins?
2004
[59]
논문
The paradox of a wide nasal aperture in cold-adapted Neandertals: a causal assessment
2008
[60]
서적
The Emergence of Modern Humans: Biocultural Adaptations in the Later Pleistocene
Cambridge University Press
1989
[61]
논문
Why not the Neandertals?
2004
[62]
논문
European early modern humans and the fate of the Neandertals
2007-05
[63]
웹사이트
The Emerging Fate Of The Neandertals
https://www.scienced[...]
[64]
논문
The assimilation model, modern human origins in Europe, and the extinction of Neandertals
2005
[65]
논문
The early Upper Paleolithic human skeleton from the Abrigo do Lagar Velho (Portugal) and modern human emergence in Iberia
1999
[66]
논문
An early modern human from the Peştera cu Oase, Romania
2003-09
[67]
논문
Hominids and hybrids: The place of Neanderthals in human evolution
1999
[68]
논문
Mitochondrial DNA and human evolution
http://artsci.wustl.[...]
1987-01-01
[69]
서적
The Essence of Anthropology
https://books.google[...]
Wadsworth Publishing
2009
[70]
서적
The climate connection: climate change and modern human evolution
https://books.google[...]
Cambridge University Press
2010
[71]
서적
Born in Africa: The Quest for the Origins of Human Life
https://books.google[...]
PublicAffairs
2011
[72]
서적
Race and Human Evolution: A Fatal Attraction
Simon & Schuster
1997
[73]
간행물
"The case against Eve"
1991
[74]
논문
Number of ancestral human species: a molecular perspective
2003
[75]
논문
Discussion on the results of some molecular studies concerning the origin of modern Chinese
2004
[76]
논문
Genetic variation in Africa
1993
[77]
논문
Revisiting the origin of modern humans in China and its implications for global human evolution
2010
[78]
논문
What is the real age of Adam and Eve? Proceedings of the Australian Society of Human Biology
2006
[79]
논문
Genetic evidence and the modern human origins debate
2008-03-05
[80]
웹사이트
Selection, nuclear genetic variation, and mtDNA
http://johnhawks.net[...]
2005-09-05
[81]
간행물
The Multiregional Evolution of Humans
2003
[82]
논문
Neandertal DNA sequences and the origin of modern humans
1997-07
[83]
논문
A view of Neandertal genetic diversity
2000
[84]
논문
Neanderthal DNA and modern human origins
2013
[85]
논문
Concocting a Divisive Theory
1998
[86]
논문
Out of Africa and Back Again: Nested Cladistic Analysis of Human Y Chromosome Variation
http://mbe.oxfordjou[...]
1998
[87]
논문
Heterogeneous patterns of variation among multiple human X-linked loci: The possible role of diversity-reducing selection in non-africans
http://www.genetics.[...]
2004-08
[88]
논문
Fixation of the human-specific CMP-N-acetylneuraminic acid hydroxylase pseudogene and implications of haplotype diversity for human evolution
2006-02
[89]
논문
More on the X files
1999-03-16
[90]
논문
X chromosome evidence for ancient human histories
1999
[91]
논문
Population genetic analysis of the N-acylsphingosine amidohydrolase gene associated with mental activity in humans
http://www.genetics.[...]
2008-03
[92]
논문
Deep haplotype divergence and long-range linkage disequilibrium at Xp21.1 provide evidence that humans descend from a structured ancestral population
http://www.genetics.[...]
2005-08
[93]
논문
Worldwide distribution of NAT2 diversity: Implications for NAT2 evolutionary history
2008-02
[94]
논문
Deciphering the ancient and complex evolutionary history of human arylamine N-acetyltransferase genes
2006-03
[95]
웹사이트
Variation in NAT1 and NAT2
http://johnhawks.net[...]
2006-01-15
[96]
논문
Evidence for archaic Asian ancestry on the human X chromosome
http://mbe.oxfordjou[...]
2005-02
[97]
논문
Testing for archaic hominin admixture on the X chromosome: Model likelihoods for the modern human RRM2P4 region from summaries of genealogical topology under the structured coalescent
http://www.genetics.[...]
2008-01
[98]
논문
Evidence suggesting that ''Homo neanderthalensis'' contributed the H2 MAPT haplotype to ''Homo sapiens''
2005
[99]
논문
Evolutionary toggling of the MAPT 17q21.31 inversion region
2008-08
[100]
논문
H1 tau haplotype-related genomic variation at 17q21.3 as an Asian heritage of the European Gypsy population
2008-11
[101]
논문
A common inversion under selection in Europeans
http://www.genetics.[...]
2005-01-16
[102]
논문
Evidence that the adaptive allele of the brain size gene microcephalin introgressed into ''Homo sapiens'' from an archaic ''Homo'' lineage
2006-11
[103]
논문
European early modern humans and the fate of the Neandertals
2007-05
[104]
논문
Microcephalin, a gene regulating brain size, continues to evolve adaptively in humans
2005-09
[105]
웹사이트
Introgression and microcephalin FAQ
http://johnhawks.net[...]
2006-11-08
[106]
논문
Neandertal genomics: Tales of a prehistoric human genome
2009-02
[107]
논문
A draft sequence of the Neandertal genome
2010-05
[108]
논문
The microcephalin ancestral allele in a Neanderthal individual
2010-05
[109]
논문
African Origin of Modern Humans in East Asia: A Tale of 12,000 Y Chromosomes
2001
[110]
논문
Haplotype Trees and Modern Human Origins
http://esa.ipb.pt/pd[...]
2005
[111]
논문
An X-linked Haplotype of Neandertal Origin Is Present Among All Non-African Populations
https://academic.oup[...]
2011-07
[112]
논문
Denisova Admixture and the First Modern Human Dispersals into Southeast Asia and Oceania
http://genetics.med.[...]
2013-09-07
[113]
논문
Sequencing and Analysis of Neanderthal Genomic DNA
2006-11-17
[114]
논문
A Draft Sequence of the Neandertal Genome
2010-05-07
[115]
웹사이트
Neandertals Live!
https://johnhawks.ne[...]
2010-12-31
[116]
논문
Genetic history of an archaic hominin group from Denisova Cave in Siberia
2010-12-23
[117]
웹사이트
The Denisova genome FAQ
http://johnhawks.net[...]
2010-12-31
[118]
논문
The Shaping of Modern Human Immune Systems by Multiregional Admixture with Archaic Humans
2011-08-25
[119]
논문
Genetic Similarities Within and Between Human Populations
2007
[120]
논문
Genetic Similarities Within and Between Human Populations
2007-05
[121]
논문
Absence of Regional Affinities of Neandertal DNA with Living Humans Does Not Reject Multiregional Evolution
2001-04-12
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com