근수축
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
근수축은 힘과 길이라는 두 가지 변수를 기준으로 설명되며, 등척성 수축, 등장성 수축, 동심성 수축, 편심성 수축으로 분류된다. 근수축은 근육의 종류에 따라 평활근, 골격근, 심근으로 나뉘며, 골격근은 운동 뉴런의 신호에 따라 수축하며, 연축, 가중, 강축 수축의 강도를 나타낸다. 근수축은 근원섬유, 미오신, 액틴, 근원섬유마디 등의 단위로 구성되며, 활주 필라멘트 이론에 의해 설명된다. 또한 길이-장력, 부하-속도 관계를 통해 역학적으로 접근할 수 있으며, 근육과 일의 관계를 보여준다. 무척추동물에서는 환형근과 종주근, 사선 무늬 근육, 비동기 근육 등이 존재한다. 근수축 연구는 1780년 루이지 갈바니의 전기 자극 실험을 시작으로, 1954년 활주 필라멘트 이론, 1952년 흥분-수축 연관 용어의 등장과 같은 역사적 발전을 거쳤다.
더 읽어볼만한 페이지
- 근육뼈대계 - 지느러미
지느러미는 물고기의 몸에 붙어 추진, 조향, 균형 유지 등 다양한 기능을 하는 부속지로, 쌍을 이루는 것과 그렇지 않은 것으로 나뉘며 종에 따라 형태와 위치가 다양하고 번식, 체온 조절, 먹이 유인 등에도 사용되는 사지동물 사지 진화와 관련된 구조이지만, 상어 지느러미 채취와 같은 위협도 존재한다. - 근육뼈대계 - 볼프의 법칙
뼈는 가해지는 하중에 반응하여 재형성되는 볼프의 법칙에 따라 기계적 변환을 통해 하중을 감지하고, 골세포와 골전구 세포를 이용하여 재형성이 조절되며, 테니스 선수 팔 뼈 강화, 역도 선수 골밀도 증가, 우주비행사 골밀도 감소 등이 그 예시이다. - 운동생리학 - 피로
피로는 신체적, 정신적 에너지 고갈 상태로, 다양한 원인으로 인해 발생하며, 약물 치료, 정신 건강 도구 활용, 생활 습관 개선 등 다양한 방식으로 관리될 수 있다. - 운동생리학 - 아데노신 삼인산
아데노신 삼인산(ATP)은 아데닌, 리보스, 세 개의 인산기로 구성된 뉴클레오티드로서, 고에너지 인산 결합의 가수분해를 통해 에너지를 방출하여 세포 과정에 사용되는 생명체의 주요 에너지 저장 및 전달 분자이며, 다양한 대사 경로를 통해 생성 및 재생될 뿐만 아니라 세포 신호 전달과 DNA 및 RNA 합성에도 필수적인 역할을 한다. - 근육계 - 힘줄
힘줄은 건세포로 이루어진 밀집규칙결합조직으로, 근육과 뼈를 연결하여 힘을 전달하며, I형 콜라겐 섬유와 프로테오글리칸으로 구성되어 탄성과 강도를 가지며, 위치 힘줄과 에너지 저장 힘줄로 기능이 나뉘고, 손상 시 염증, 수복/증식, 개조 단계를 거쳐 치유되며, 과거에는 섬유로 활용되고 일부 문화권에서는 식용으로도 사용되었다. - 근육계 - 단백질 보충제
단백질 보충제는 근력 훈련과 함께 섭취 시 근육량 증가 및 근력 향상에 도움을 줄 수 있으며, 과체중인 사람의 체중 및 지방 감소, 노인의 단백질 섭취 요구량 충족에도 기여할 수 있다.
| 근수축 | |
|---|---|
| 개요 | |
| 정의 | 근육 내에서 장력 생성 부위의 활성화 |
| 참고 문헌 | |
| 관련 항목 | |
| 영어 | muscle contraction |
| 다른 이름 | 근육 수축 |
2. 근수축의 종류
근수축은 힘과 길이라는 두 가지 변수를 기준으로 구분할 수 있다. 힘은 근육이 물체에 가하는 힘인 장력과 물체가 근육에 가하는 힘인 부하로 나뉜다.[1] 근육의 길이 변화 여부에 따라 다음과 같이 근수축의 종류를 구분할 수 있다.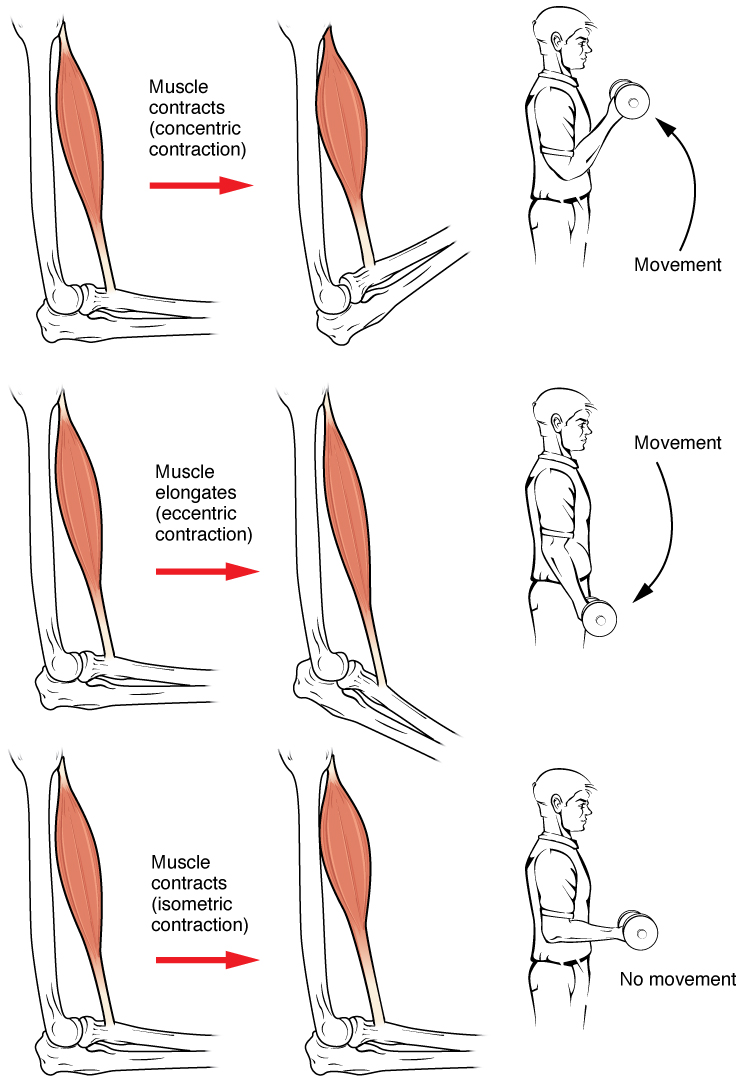
- 등척성 수축: 근육 길이가 변하지 않고 근육 장력만 변하는 경우이다.[1][4][5][6]
- 등장성 수축: 근육 장력이 동일하게 유지되면서 근육 길이가 변하는 경우이다.[1][4][5][6]
- 구심성 수축: 근육이 짧아지는 수축이다.
- 원심성 수축: 근육이 늘어나는 수축이다.[1][7]
하지만 실제 움직임에서는 근육의 길이와 장력이 시간에 따라 다양하게 변하므로, 한 가지 유형으로만 근수축을 설명하기는 어렵다.[3]
척추동물의 근육 조직은 골격근, 평활근, 심근 세 가지로 나뉜다. 골격근은 운동을 담당하고, 평활근은 혈관, 위장관 등 지속적인 수축이 필요한 부위에 분포한다. 심근은 심장 박동을 일으킨다. 골격근과 심근은 규칙적인 배열 때문에 가로무늬근으로 불린다.
2. 1. 등장성 수축 (Isotonic contraction)
등장성 수축은 근육의 장력이 근육 길이의 변화에도 불구하고 일정하게 유지되는 수축이다.[1][4][5][6] 이는 근육의 수축력이 근육에 가해지는 총 부하와 일치할 때 발생하며, 물건을 들어 올리거나 내려놓는 경우와 같이 손발을 움직이는 경우가 이에 해당한다. 등장성 수축에는 단축성 수축과 신장성 수축이 있다.
자연스러운 움직임에서 근수축은 길이와 장력의 변화를 시간에 따라 다양하게 생성하기 때문에 다면적이다.[3] 따라서 운동 중 근육이 활성화될 때 길이와 장력이 모두 일정하게 유지되지는 않는다.
2. 1. 1. 동심성 수축 (Concentric contraction)
근육이 부하보다 큰 장력을 일으켜 근육이 수축하면서 짧아지는 경우이다. 따라서 장력이 발생하는 방향과 같은 방향으로 근수축이 일어나게 된다.[57] 근수축에서 근육의 장력은 부하를 극복하기에 충분하며, 근육은 수축하면서 짧아진다.[8] 이는 근육이 생성하는 힘이 수축에 반대하는 부하를 초과할 때 발생한다.동심성 수축 동안, 근육은 활주 필라멘트 이론에 따라 수축하도록 자극을 받는다. 이는 근육 전체 길이에서 발생하여 기시점과 종착점에서 힘을 생성하여 근육을 짧게 만들고 관절의 각도를 변화시킨다. 팔꿈치 관절과 관련하여, 이두근의 동심성 수축은 손이 다리에서 어깨로 이동하면서 팔꿈치에서 팔을 구부리게 한다(바이셉스 컬). 위팔세갈래근의 동심성 수축은 관절의 각도를 반대 방향으로 변화시켜 팔을 펴고 손을 다리 쪽으로 이동시킨다.
2. 1. 2. 편심성 수축 (Eccentric contraction)
근육이 늘어나 외부 힘의 작용에 저항하는 수축이다. 물건을 내려놓는 경우와 같이 근육을 잡아당기면서 장력을 내는 경우이다. 이 경우 이두근에 있는 근섬유는 늘어나더라도 늘어나는 방향과 반대로 수축작용이 일어난다.[57] 생성된 장력이 근육의 외부 부하를 극복하기에 불충분하여 근육 섬유가 수축하면서 길어지는 현상이다.[9] 근육이 관절을 수축 방향으로 당기는 대신, 움직임의 끝에서 관절의 속도를 늦추거나 부하의 위치를 제어하는 역할을 한다.이는 비자발적으로 발생할 수 있으며 (예: 근육이 들어올릴 수 없을 정도로 무거운 무게를 들어올리려 할 때) 자발적으로 발생할 수도 있다 (예: 근육이 움직임을 '부드럽게' 하거나 내리막길을 걸을 때처럼 중력에 저항할 때). 단기적으로, 원심성 수축과 단축성 수축을 모두 포함하는 근력 운동은 단축성 수축만으로 운동하는 것보다 근력을 더 많이 증가시키는 것으로 보인다.[11] 그러나 운동으로 인한 근육 손상 또한 근육이 길어지는 수축 동안 더 크다.[10]
상완이두근의 원심성 수축 동안, 팔꿈치는 구부러진 상태에서 움직임을 시작한 다음 손이 어깨에서 멀어짐에 따라 펴진다. 상완삼두근의 원심성 수축 동안, 팔꿈치는 펴진 상태에서 움직임을 시작한 다음 손이 어깨쪽으로 움직임에 따라 구부러진다. 데스민, 타이틴, 그리고 다른 Z-선 단백질은 원심성 수축에 관여하지만, 그 메커니즘은 동심성 수축에서의 교차 다리 순환에 비해 잘 이해되지 않고 있다.[12]
근육이 부(-)의 역학적 일을 하고 있지만 (일이 근육 ''에'' 수행됨), 화학 에너지 (지방 또는 포도당, 또는 ATP에 일시적으로 저장됨)는 그럼에도 불구하고 소모되는데, 이는 동일한 힘의 동심성 수축 동안 소모되는 것보다 적다. 예를 들어, 계단을 내려가는 것보다 올라가는 데 더 많은 에너지를 소비한다.
과도한 원심성 부하를 받는 근육은 과부하 시 동심성 부하에 비해 더 큰 손상을 입는다 (예: 근비대 또는 근력 운동 시). 원심성 수축이 웨이트 트레이닝에 사용될 때, 이는 일반적으로 ''네거티브''라고 불린다. 동심성 수축 동안, 미오신과 액틴의 수축성 근육 마이오필라멘트가 서로 미끄러지면서 Z선을 함께 당긴다. 원심성 수축 동안, 마이오필라멘트는 반대 방향으로 서로 미끄러지지만, 원심성 수축 동안 미오신 헤드의 실제 움직임은 알려져 있지 않다. 무거운 원심성 부하를 특징으로 하는 운동은 실제로 더 큰 무게를 지탱할 수 있으며 (원심성 수축 동안 근육은 동심성 수축 동안보다 약 40% 더 강하다) 훈련 후 1~2일 후에 더 큰 근육 손상과 지연성 근육통을 유발한다. 원심성 및 동심성 근육 수축을 모두 포함하는 운동 (예: 강한 수축과 무게를 제어하여 낮추는 것)은 동심성 수축만으로는 얻을 수 없는 더 큰 근력 증가를 가져올 수 있다.[11][12] 익숙하지 않은 무거운 원심성 수축은 쉽게 과훈련으로 이어질 수 있지만, 적절한 훈련은 부상에 대한 보호를 제공할 수 있다.[12]
2. 2. 등척성 수축 (Isometric contraction)
등척성 수축은 장력은 증가하더라도 근육의 길이가 변하지 않아 수행된 역학적 일은 없다. 하지만 근육내에서 생리학적일은 수행되었으며, 이 때 발생되는 열은 등장성 열생산이라고 한다. 손발이 움직이지 않는 힘, 사람이 부동자세로 서있으면서 체중을 지탱하는 경우, 물체의 무게가 무거워서 들어올리지 못할 때에 적용될 수 있다.근육의 길이가 변하지 않고 근육 장력만 변하는 경우, 근수축은 등척성 수축으로 묘사된다.[1][4][5][6]
근육의 등척성 수축은 길이를 변화시키지 않으면서 장력을 발생시킨다.[1][4][5][6] 예를 들어 손과 전완의 근육이 물체를 잡을 때, 손의 관절은 움직이지 않지만 근육은 물체를 떨어뜨리지 않기에 충분한 힘을 생성한다.
3. 근수축의 단위
골격근은 다음과 같이 구성된다.[55]
척추동물에는 골격근, 평활근, 심근의 세 가지 유형의 근육 조직이 있다. 골격근은 신체 근육량의 대부분을 차지하며 운동을 담당한다. 평활근은 혈관, 위장관 등 지속적인 수축이 필요한 신체 부위를 구성한다. 심근은 심장을 구성하여 혈액을 펌프질한다. 골격근과 심근은 A 밴드와 I 밴드가 번갈아 나타나는 규칙적인 배열 때문에 현미경으로 관찰하면 가로무늬가 보여 가로무늬근이라고 불린다.
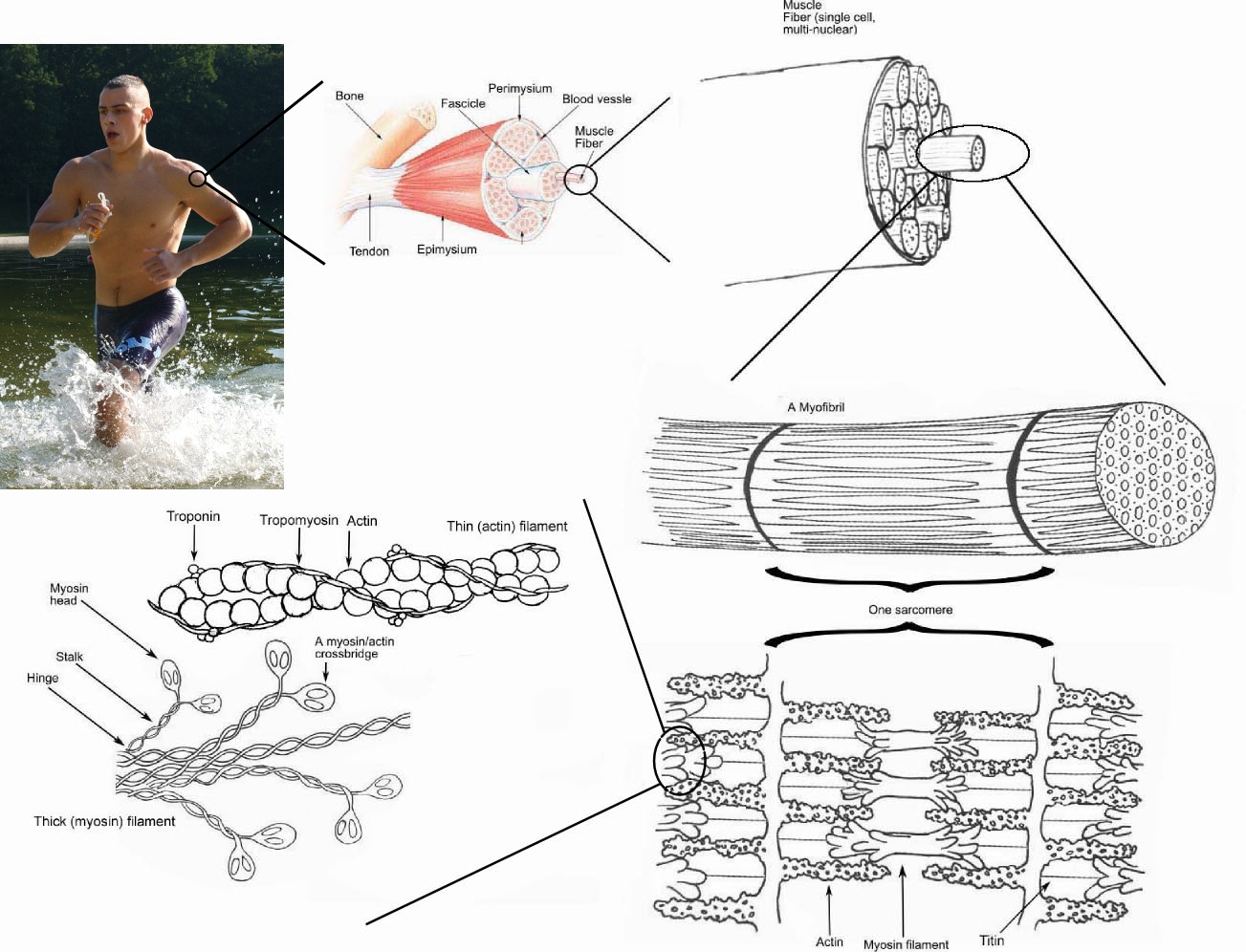
근육은 여러 단계의 구조로 구성되며, 가장 작은 단위는 미오신과 액틴 단백질이다. 이들은 각각 굵은 필라멘트와 가는 필라멘트를 구성하며, 이 필라멘트들이 모여 근원섬유를 이룬다. 근원섬유는 근섬유 내에서 수축 기능을 담당하는 단위이다.
3. 1. 근원섬유 (Myofibril)
근육을 이루는 단일 골격근 세포, 즉 근섬유는 지름이 10~100㎛ 정도로 특수화된 세포소기관인 다량의 근원섬유로 이루어지며 근원섬유는 근섬유의 수축 단위가 된다.[56] 각각의 근원섬유들은 지름이 1um 정도로 세포골격 요소인 굵은 필라멘트와 가는 필라멘트의 구조로 이루어져 있다. 굵은 필라멘트는 미오신이라는 단백질로 이루어져 있으며, 가는 필라멘트는 둥근 모양의 액틴 단백질로 구성되어 있다.3. 2. 굵은 필라멘트: 미오신 (Myosin)
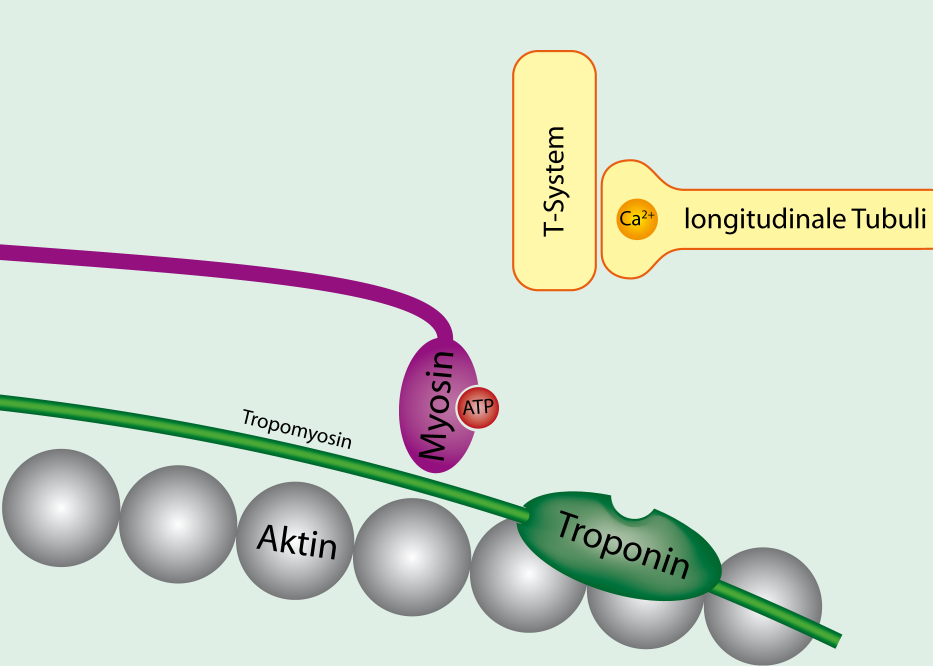
미오신 단백질은 2개의 소단위가 붙어있는 형태로, 각 소단위는 꼬리 부분과 머리 부분으로 구성되어 있다. 꼬리 부분은 중앙쪽을 향하며, 머리 부분은 액틴과 결합할 수 있는 부분과 ATP와 결합할 수 있는 부분을 가지고 있어 교차결합을 하는데 중요한 역할을 한다. 결합된 ATP가 분해되면서 방출되는 에너지에 의해 미오신 머리가 움직이고 이에 의해 액틴을 근원섬유마디의 가운데 쪽으로 끌어오면 근원섬유마디의 간격이 좁아지게 된다. 이 때, 각각의 근원섬유마디에서 발생하는 힘이 합쳐져 근력으로 작용하게 된다.[1]
3. 3. 가는 필라멘트: 액틴 (Actin)
액틴 분자는 미오신 분자에 비해 1/10보다도 작은 분자량을 가지며, 두 나선형의 꼬인 사슬처럼 연결되어 있다.
트로포마이오신(tropomyosin)은 액틴 필라멘트를 따라 띠처럼 감겨있는 구조를 하며 교차 결합하는 액틴 부위를 덮고 있어서 액틴과 마이오신의 결합에서 중요한 역할을 하는 단백질이다.
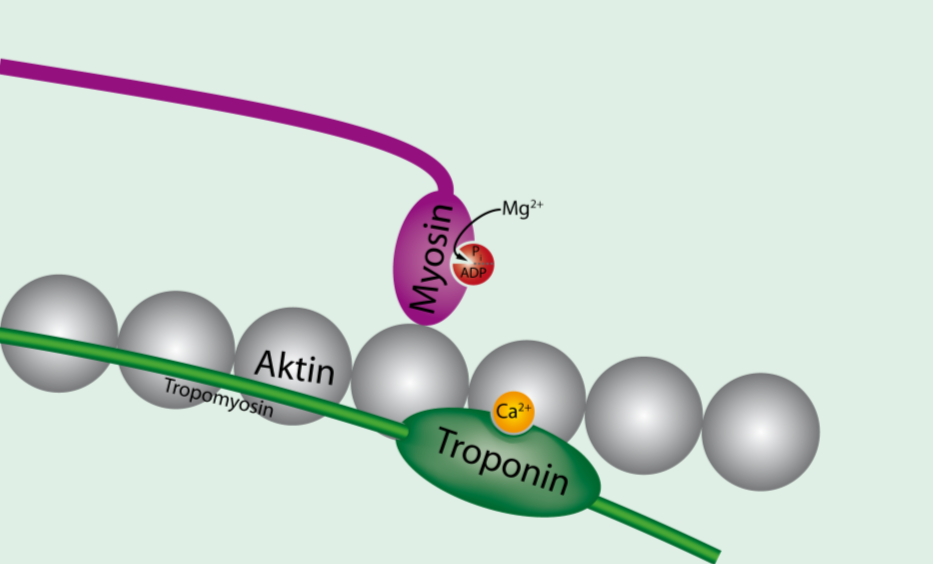
트로포닌은 칼슘 이온과의 결합력이 높은 단백질로 액틴 필라멘트 위에 존재하여 트로포마이오신을 안정화시키고 액틴과 마이오신의 결합을 돕는다. 소포체에서 빠져나온 칼슘 이온이 트로포닌과 결합하면 불안정해진 트로포마이오신이 변형되면서 액틴과 마이오신은 결합을 하게 된다. 반면에 칼슘 이온 없이 트로포마이오신과 트로포닌이 액틴에 붙으면 미오신이 결합하지 못한다.
3. 4. 근원섬유마디 (Sarcomere)
근육을 이루는 단일 골격근세포, 즉 근섬유는 지름이 10~100㎛ 정도로 특수화된 세포소기관인 다량의 근원섬유로 이루어지며 근원섬유는 근섬유의 수축 단위가 된다.[56] 각각의 근원섬유들은 지름이 1㎛ 정도로 세포골격 요소인 굵은 필라멘트와 가는 필라멘트의 구조로 이루어져 있다. 굵은 필라멘트는 미오신이라는 단백질로 이루어져 있으며, 가는 필라멘트는 둥근 모양의 액틴 단백질로 구성되어 있다.근원섬유를 옆에서 보면 각각의 근원섬유는 밝고 어두운 띠의 연속적인 배열로 구성되어 있는데, 굵은 필라멘트와 가는 필라멘트의 겹쳐진 배열이 이러한 구조로 보이게끔 한다. 밝은 부분의 중앙에는 수직으로 선이 나타나는데 이를 Z선이라고 한다. Z선을 기준으로 나누어지는 각각의 단위를 근원섬유마디 혹은 근절(sarcomere)이라고 부른다. 골격근의 기능적 단위가 되는 근절 길이는 수축과 이완에 따라 달라져서, 휴식기에는 2.5um지만 최대 수축기에는 약 1.5um, 최대 이완기에는 약 정도 된다.

4. 골격근 수축의 과정
골격근 수축은 신경근 접합부에서 시작하여 흥분-수축 연접을 거쳐 활주 필라멘트 이론에 따라 일어난다.
신경근 접합부는 운동 신경과 근육 섬유 사이의 화학적 시냅스로, 운동 신경이 근육 섬유에 신호를 전달하여 근육 수축을 시작하는 곳이다. 운동 신경의 활동 전위가 신경근 접합부에 도달하면, 칼슘 이온(Ca2+)이 유입되고, 이는 신경전달물질인 아세틸콜린의 방출을 유도한다. 아세틸콜린은 신경근 접합부의 니코틴성 아세틸콜린 수용체에 결합하여 나트륨/칼륨 통로를 열어 근육 세포막의 탈분극을 일으킨다. 이로 인해 발생하는 종판 전위는 근육 세포막의 전압 개폐 이온 통로를 열어 활동 전위를 생성하고, 이는 모든 방향으로 퍼져 나간다. 활동 전위가 멈추면 아세틸콜린은 더 이상 방출되지 않고, 시냅스 틈새에 남아있는 아세틸콜린은 아세틸콜린 에스테라아제에 의해 분해되거나 시냅스 단추에 의해 재흡수된다.[18]
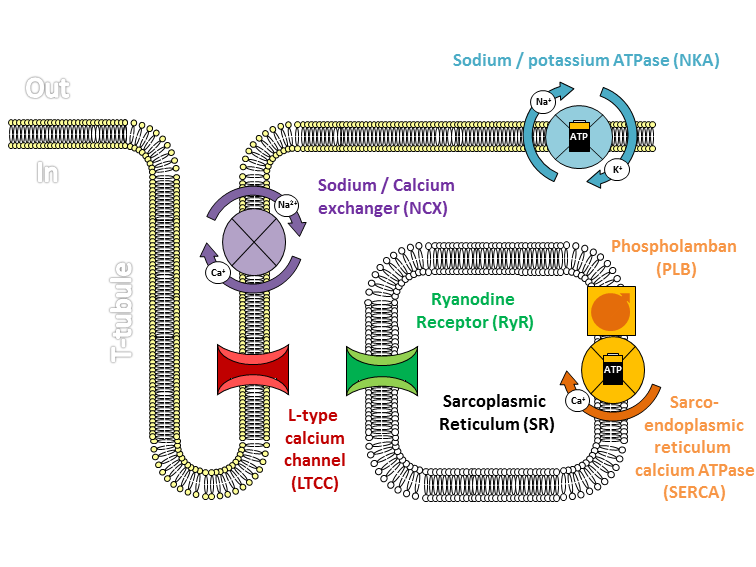
흥분-수축 연접(ECC)은 골격근의 탈분극이 근육 활동 전위를 발생시키는 과정이다. 활동 전위는 근육 섬유의 T-세관 네트워크를 통해 퍼져 근육 섬유 내부를 탈분극시킨다. 이는 종말 수조의 디히드로피리딘 수용체를 활성화시키고, 이는 근소포체의 리아노딘 수용체와 상호 작용하여 칼슘 이온(Ca2+)을 방출시킨다. 방출된 칼슘 이온은 액틴 필라멘트의 트로포닌 C에 결합하여 교차 브리지 사이클을 가능하게 한다.[19]
원하는 움직임이 달성되면 이완이 일어난다. 칼슘 이온 완충제는 칼슘 이온에 결합하여 빠른 이완을 돕고, 근/소포체 칼슘-ATPase (SERCA)는 칼슘 이온을 근소포체로 능동적으로 펌핑하여 영구적인 이완을 유발한다.[19][20] 미토콘드리아 또한 칼슘 이온 재흡수에 참여하여 근소포체에 전달한다. 칼슘 이온 농도가 휴지 상태로 감소하면, 칼슘 이온이 트로포닌 C에서 방출되어 교차 브리지 사이클이 중단되고 이완이 발생한다.[21]
미끄럼 필라멘트 이론은 얇은 필라멘트가 두꺼운 필라멘트 위로 미끄러져 근육에 장력을 발생시키는 반복적인 과정을 설명한다.[22] 이 이론은 1954년에 앤드루 헉슬리와 롤프 니더르게르케, 휴 헉슬리와 진 핸슨에 의해 독립적으로 개발되었다.[23][24] 생리학적으로 이러한 수축은 근절 전체에서 균일하지 않지만, 티틴의 탄성 근세사 작용에 의해 두꺼운 필라멘트가 중앙 위치로 당겨져 근절 전체에 걸쳐 균일한 장력이 유지된다.[25]
4. 1. 신경근 접합부 (Neuromuscular junction)
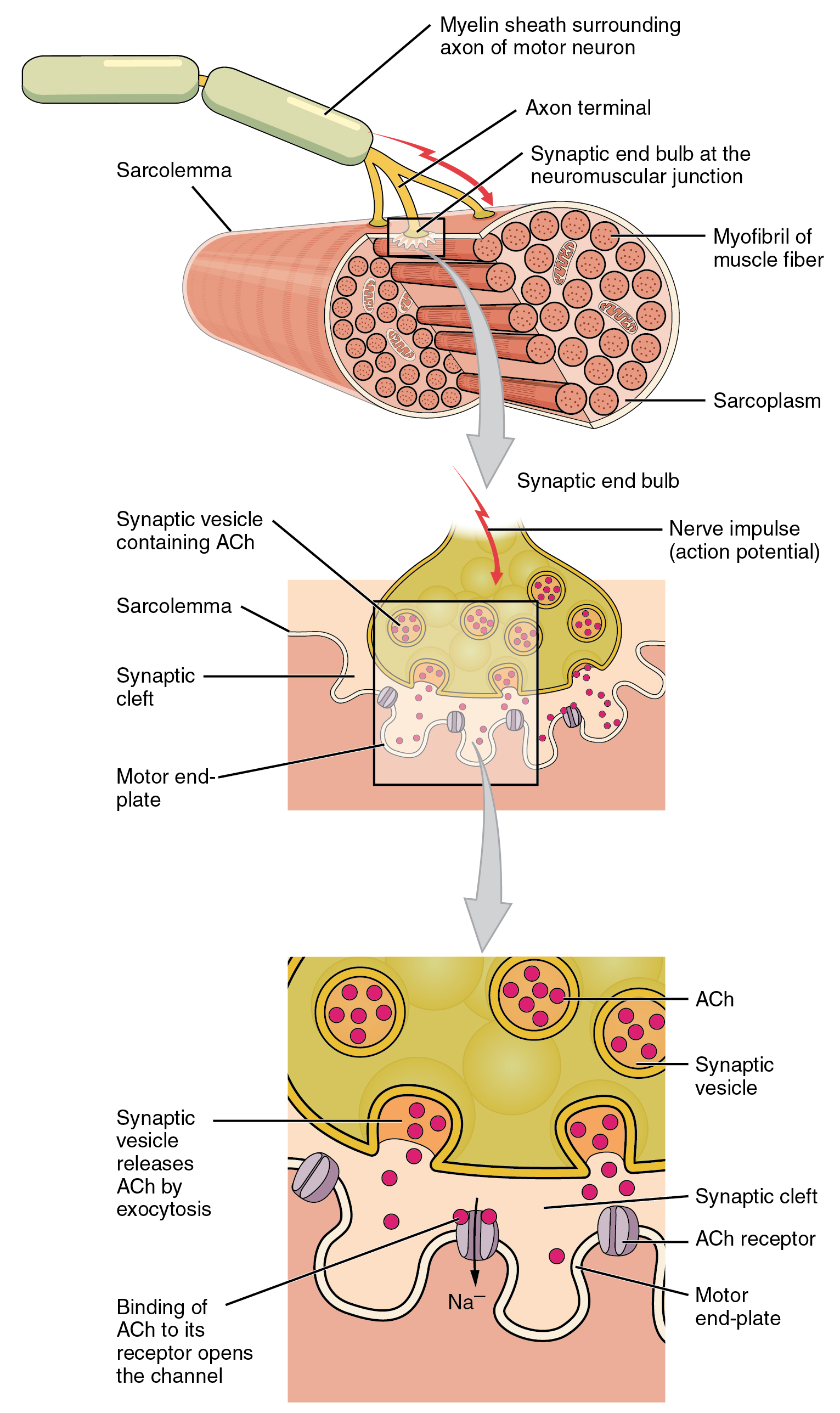
신경근 접합부는 운동 신경과 근육 섬유 사이의 접촉으로 형성되는 화학 시냅스이다.[17] 이곳은 운동 신경이 근육 섬유로 신호를 전달하여 근육 수축을 시작하는 부위이다. 신경근 접합부에서 근육 섬유의 탈분극을 일으키는 일련의 과정은 운동 신경의 세포체에서 활동 전위가 시작될 때 시작되며, 이는 신경근 접합부를 향해 축삭을 따라 도약 전도로 전파된다. 종말 단추에 도달하면 활동 전위로 인해 전압 개폐 칼슘 통로를 통해 칼슘 이온( )이 종말 부위로 유입된다. 칼슘 이온 유입은 신경전달물질 아세틸콜린을 포함하는 시냅스 소포가 혈장막과 융합되어 아세틸콜린을 운동 신경 종말과 골격근 섬유의 신경근 접합부 사이의 시냅스 틈새로 방출하게 한다. 아세틸콜린은 시냅스를 가로질러 확산되어 신경근 접합부의 니코틴성 아세틸콜린 수용체에 결합하여 활성화시킨다. 니코틴성 수용체가 활성화되면 내재된 나트륨/칼륨 통로가 열리고 나트륨이 유입되고 칼륨이 유출된다. 그 결과, 근육 세포막의 극성이 반전되고 나트륨이 유입되면서 전압이 휴지막 전위인 -90mV에서 최대 +75mV까지 빠르게 상승한다. 그런 다음 칼륨이 유출되면 막 전위가 과분극되고 다시 휴지막 전위로 조정된다. 이러한 급격한 변동을 종판 전위라고 한다.[18] 종판 옆의 근육 세포막의 전압 개폐 이온 통로는 종판 전위에 반응하여 열린다. 이들은 나트륨과 칼륨에 특이적이며 하나만 통과시킨다. 이러한 이온 이동의 파동은 모든 방향으로 운동 종판에서 퍼지는 활동 전위를 생성한다.[18] 활동 전위의 도달이 멈추면 아세틸콜린이 종말 단추에서 더 이상 방출되지 않는다. 시냅스 틈새에 남아있는 아세틸콜린은 활성 아세틸콜린 에스테라아제에 의해 분해되거나 시냅스 단추에 의해 재흡수되며 분해된 아세틸콜린을 대체할 아세틸콜린은 남지 않는다.
4. 2. 흥분-수축 연접 (Excitation-contraction coupling)
운동뉴런의 말단에서 분비된 아세틸콜린에 의하여 근세포막이 탈분극된다.[19] 세포막을 따라 온 활동전위는 T세관따라 근섬유로 전도된다.[19] 근소포체에 저장되어있던 칼슘이온이 방출된다.[19] 방출된 칼슘이온과 트로포닌이 결합하면서 액틴의 결합부위가 노출된다.[19] ATP분해효소의 활성 후 ADP + P 로 분해시키면서 방출된 에너지는 미오신 머리가 다시 곧추서게 한다.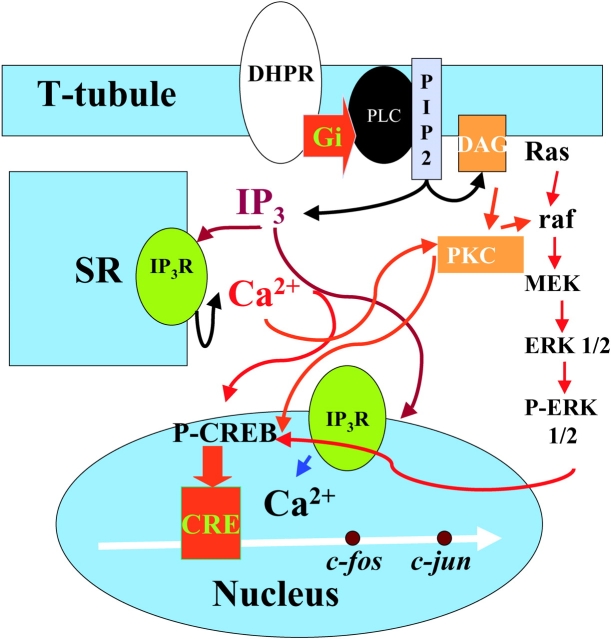
흥분-수축 연계(ECC)는 골격근의 탈분극(보통 신경 지배를 통해)이 근육 활동 전위를 발생시킬 때 발생한다. 이 활동 전위는 근육 표면을 가로질러 근육 섬유의 T-세관 네트워크로 퍼져 근육 섬유의 내부 부분을 탈분극시킨다. 이는 종말 수조의 디히드로피리딘 수용체를 활성화시키며, 종말 수조는 인접한 근소포체의 리아노딘 수용체와 가깝게 위치한다. 활성화된 디히드로피리딘 수용체는 발판 과정(알로스테릭하게 리아노딘 수용체를 활성화하는 형태 변화 포함)을 통해 리아노딘 수용체와 물리적으로 상호 작용하여 활성화시킨다. 리아노딘 수용체가 열리면 Ca2+이 근소포체에서 국소 접합 공간으로 방출되어 벌크 세포질로 확산되어 칼슘 스파크를 유발한다.[19] 활동 전위는 수천 개의 칼슘 스파크를 거의 동시적으로 활성화시키고 세포 전체의 칼슘 증가를 유발하여 칼슘 과도 현상의 상승을 일으킨다. 세포질로 방출된 Ca2+는 액틴 필라멘트의 트로포닌 C에 결합한다.[19] 이 결합은 액틴 필라멘트가 교차 브리지 사이클을 수행하여 힘을 생성하고, 경우에 따라 움직임을 생성할 수 있도록 한다.
원하는 움직임이 달성되면, 다양한 경로를 통해 빠르게 이완을 달성할 수 있다. 이완은 매우 높은 친화도로 Ca2+에 결합하는 다양한 세포질 단백질을 포함하는 Ca2+ 완충제를 통해 신속하게 달성된다.[20] 이러한 세포질 단백질은 속근 섬유에서 빠른 이완을 가능하게 한다. 느리기는 하지만, 근/소포체 칼슘-ATPase (SERCA)는 Ca2+을 근소포체로 능동적으로 펌핑하여 다음 활동 전위가 도착할 때까지 영구적인 이완을 유발한다.[19]
미토콘드리아는 또한 Ca2+ 재흡수에 참여하여 궁극적으로 수집된 Ca2+을 근소포체의 저장고인 SERCA에 전달한다. 몇 가지 이완 기전(NCX, Ca2+ 펌프 및 Ca2+ 누출 채널)은 Ca2+을 세포 밖으로 완전히 이동시킨다.[21] Ca2+ 농도가 휴지 상태로 감소하면, Ca2+이 트로포닌 C에서 방출되어 교차 브리지 사이클을 허용하지 않으므로 힘이 감소하고 이완이 발생한다. 이완이 완전히 완료되면, 근육은 다시 수축할 수 있으며, 따라서 사이클이 완전히 재설정된다.
4. 3. 활주 필라멘트 이론 (Sliding filament theory)
미오신 단백질은 2개의 소단위가 붙어있는 형태로, 각 소단위는 꼬리부분과 머리부분으로 구성되어 있다. 꼬리부분은 중앙쪽을 향하며, 머리부분은 액틴과 결합할 수 있는 부분과 ATP와 결합할 수 있는 부분을 가지고 있어 교차결합을 하는데 중요한 역할을 한다. 결합된 ATP가 분해되면서 방출되는 에너지에 의해 미오신 머리가 움직이고 이에 의해 액틴을 근원섬유마디의 가운데 쪽으로 끌어오면 근원섬유마디의 간격이 좁아지게 된다. 이 때, 각각의 근원섬유마디에서 발생하는 힘이 합쳐져 근력으로 작용하게 된다.- 운동뉴런의 말단에서 분비된 아세틸콜린에 의하여 근세포막이 탈분극된다.
- 세포막을 따라 온 활동전위는 T세관따라 근섬유로 전도된다.
- 근소포체에 저장되어있던 칼슘이온이 방출된다.
- 방출된 칼슘이온과 트로포닌이 결합하면서 액틴의 결합부위가 노출된다.
- ATP분해효소의 활성 후 ADP + P 로 분해시키면서 방출된 에너지는 미오신 머리가 다시 곧추서게 한다.

미끄럼 필라멘트 이론은 근육이 수축하는 과정을 설명한다. 이는 얇은 필라멘트가 두꺼운 필라멘트 위로 미끄러져 근육에 장력을 발생시키는 반복적인 사건의 순환이다.[22] 이 이론은 1954년에 앤드루 헉슬리와 롤프 니더르게르케 그리고 휴 헉슬리와 진 핸슨에 의해 독립적으로 개발되었다.[23][24] 생리학적으로 이러한 수축은 근절 전체에서 균일하지 않다. 두꺼운 필라멘트의 중심 위치는 불안정해지고 수축 중에 이동할 수 있지만, 이는 티틴의 탄성 근세사 작용에 의해 상쇄된다. 이 미세 근세사는 두꺼운 필라멘트를 중앙 위치로 당겨 근절 전체에 걸쳐 균일한 장력을 유지한다.[25]
5. 근수축에 대한 역학적 접근
근육이 생산해내는 힘은 근수축에 대한 생리학적 기작뿐만 아니라 역학적 특성에 의하여 영향을 받을 수 있으며, 이에 대한 연구가 이루어져 왔다.
근육에 가해지는 부하가 증가하면 수축 속도는 감소하고, 결국 속도가 0이 되는 지점에서 최대 등척성 장력에 도달한다.[58] 근수축 속도는 최대 장력에 도달하는 데 필요한 시간으로 계산할 수 있다.[58]
등장성 수축에서는 하중이 커질수록 근육의 수축 속도가 감소한다. 하중이 0일 때 근육의 수축 속도는 최대가 되며, 근육의 최대 장력보다 하중이 커지면 수축 속도는 0이 된다. 즉, 무거운 물체는 천천히 들어 올려지는데, 이는 큰 하중의 경우 교차 결합이 더 오랫동안 스트로크를 하기 때문이다.[59]
힘-속도 관계는 근육이 길이를 변화시키는 속도와 생성하는 힘의 양 사이의 관계를 나타낸다. 단축 속도가 증가함에 따라 힘은 쌍곡선 형태로 감소하며, 결국 최대 속도에서 0에 도달한다. 근육이 늘어날 때는 반대로 힘이 등척성 최대 힘 이상으로 증가하다가 최대 힘에 도달한다.
펜(Fenn)과 마시(March)는 1935년 논문에서 힘-속도 관계를 다음 식으로 나타냈다.[60]
:
:(P: 주어진 힘, V: 수축 속도, : 부하가 없을 때의 수축속도, B와 K는 상수)
아치볼드 힐(Archibald Hill)은 1938년 근육의 역학적 성질에 대해 연구하여 근육의 수축 속도와 부하 간 관계는 직각쌍곡선이 된다는 것을 밝혀내었고 이를 방정식으로 표현하였다. P는 근육이 늘어나 줄어들지 않고 지탱할 수 있는 최대 부하이고, 보다 작은 P만큼의 부하가 가해지면 근육은 수축하며 열이 발생하는데, 이때 수축하는 속도를 V로 정의한다. a[g] 와 b[cm/sec]는 상수이다. 위 관계를 정리한 기본 방정식은 다음과 같다.[61]
: ----------식①
변수는 수축 속도 V와 외부 힘 P이다. 위 식을 전개 후 정리하면 다음과 같다(c는 상수).
:
힐(Hill)의 기본 방정식을 근육의 생리학적 기작에 적용하면, 전체 근육의 최대 부하는 Z선을 이루는 각 교차결합의 힘과 결합 수의 곱이다. 가해진 힘 P는 각 결합에 전해지는 외부 힘과 결합 수의 곱으로 표현할 수 있다.
: ,
그러나 모든 결합이 작동하는 것이 아니고 일부 결합만 작동하므로, 전체 결합 중 작동하는 결합의 수를 로 정의한다. 결합과 분리 과정은 마찰력의 존재를 의미하며, 마찰력은 수축 속도에 비례하여 마찰계수 와 의 곱으로 표현한다. 따라서 전체 다리에 의해 발생된 힘은 외부 힘에서 마찰력 를 뺀 값이다. 뉴턴의 제2법칙에 의해 정리하면 다음과 같다.
:
결합에 의해 발생된 힘과 마찰력은 결합수에 비례하므로, 를 한 결합에 의한 마찰계수로 정의하면 이다. 식을 다음과 같이 변형할 수 있다.
: ----------식②
값은 힘에 선형적으로 의존한다고 보면 다음 식을 세울 수 있다.[62]
:
이면 이고 이다. 이고 수축속도가 최대일 때는 이며, 이때 를 라고 한다. 두 경우를 식으로 표현하면 다음과 같다.
:인 경우 :
: 인 경우 :
두 식을 식②에 대입하면 다음 식을 얻는다.
:
이 결과를 식①에 대입하면 다음 식을 얻는다.
:--------식③
이는 힐(Hill)의 방정식에서 , 로 놓았을 때와 유사하다. 한 결합에 가해지는 힘이 가장 적을 때, 즉 인 경우 근육은 최대로 수축하는데
:
이 식은 근육의 최대 수축속도가 마찰계수와 최대장력으로 표현됨을 보여준다. 이는 힐(Hill)의 방정식이 마찰이 있는 단백질 띠의 움직임, 즉 소성흐름을 묘사함을 보여준다.[63]
근육이 장력을 발휘하여 물체를 움직이면 일을 했다고 정의한다. 그러나 근육 수축 동안 마찰력과 열이 발생한다. 근육에 걸리는 부하가 클수록 더 큰 장력을 발휘해야 하므로 더 많은 열이 발생한다. 일반적으로 에너지의 약 25%가 일에 사용되고, 나머지 75%는 열로 전환되어[59] 체온 유지 등에 사용된다.
5. 1. 길이-장력 곡선 관계
장력은 근섬유에서 가는 필라멘트와 굵은 필라멘트 간 교차결합 수에 의존한다.장력과 근육의 길이 변화 간 관계 연구는 1966년 Gordon에 의해 이루어졌다. 생체 내에서 근육은 최대의 수축력을 갖도록 하기 위하여 길이-장력 관계에서의 적정 길이 부근으로 가게 되지만, 근육에 부착된 골격에 의해 제한을 받기 때문에 적정 길이로부터 30% 범위 내에서 길이 변화가 이루어진다.
- '''근육의 길이가 적정 길이일 때''': 근절이 2.0~2.5㎛ 정도의 적정 길이를 보이는 휴지기일 때 액틴 필라멘트와 미오신 필라멘트의 교차결합 수는 최대가 되고, 이때에 최대 장력이 발휘된다.
- '''근육의 길이가 적정 길이보다 클 때''': 근육의 길이가 늘어날 경우 두 개의 필라멘트는 반대 방향으로 당겨져 서로 겹치지 않는 부위가 생기게 된다. 따라서 교차결합의 수는 점차 줄어들게 되면서 장력도 약해지게 된다. 4㎛ 부근에 이르러서는 가는 필라멘트가 굵은 필라멘트와 이루는 중첩이 없어지면서 더 이상 장력을 발휘하지 못하게 된다.
- '''근육의 길이가 적정 길이보다 작을 때''': 근육이 수축하게 되면 교차결합끼리 중첩이 일어나 교차결합의 수는 줄어들게 되고, 장력도 감소한다. 장력이 감소하는 데에는 여러 가지 물리적 원인이 있는데, 굵은 필라멘트가 Z선 이상으로 더 이상 수축이 불가능하다는 점, 양쪽에서의 가는 필라멘트가 서로 겹쳐서 교차결합 수가 줄어들 수밖에 없다는 점을 들 수 있다.
이렇게 근육이 안정 길이를 벗어나 신장되는 경우, 수동 장력은 커지며 능동 장력은 감소하면서 총 장력은 증가하게 된다.
근육의 길이-장력 관계는 등척성 수축의 강도와 수축이 일어나는 근육의 길이를 연관시킨다. 근육은 이상적인 길이(종종 휴식 길이)에 가까울 때 가장 큰 능동적 장력으로 작동한다. 이 길이를 벗어나 늘어나거나 짧아지면(근육 자체의 작용 또는 외부 힘에 의해), 생성되는 최대 능동적 장력이 감소한다.[31] 이러한 감소는 작은 편차에서는 미미하지만, 길이가 이상적인 길이에서 더 멀어질수록 장력은 급격히 떨어진다. 근육 세포 내의 탄성 단백질(예: 타이틴)과 세포 외 기질로 인해, 근육이 특정 길이를 넘어 늘어나면 신장을 반대하는 완전히 수동적인 장력이 발생한다. 이를 합하면, 능동적 장력의 최고점을 훨씬 넘어 활성 근육을 늘리는 것에 대한 강한 저항이 존재한다.
5. 2. 부하-속도 관계
근육에 가해지는 부하가 증가하면 수축 속도는 감소하고, 결국 속도가 0이 되는 지점에서 최대 등척성 장력에 도달한다.[58] 근수축 속도는 최대 장력에 도달하는 데 필요한 시간으로 계산할 수 있다.[58]등장성 수축에서는 하중이 커질수록 근육의 수축 속도가 감소한다. 하중이 0일 때 근육의 수축 속도는 최대가 되며, 근육의 최대 장력보다 하중이 커지면 수축 속도는 0이 된다. 즉, 무거운 물체는 천천히 들어 올려지는데, 이는 큰 하중의 경우 교차 결합이 더 오랫동안 스트로크를 하기 때문이다.[59]
힘-속도 관계는 근육이 길이를 변화시키는 속도와 생성하는 힘의 양 사이의 관계를 나타낸다. 단축 속도가 증가함에 따라 힘은 쌍곡선 형태로 감소하며, 결국 최대 속도에서 0에 도달한다. 근육이 늘어날 때는 반대로 힘이 등척성 최대 힘 이상으로 증가하다가 최대 힘에 도달한다.
5. 3. 근육의 수축 속도와 부하 간 기본 방정식
펜(Fenn)과 마시(March)는 1935년 발표한 논문에서 힘-속도 관계를 다음 식으로 나타냈다.[60]:
:(P: 주어진 힘, V: 수축 속도, : 부하가 없을 때의 수축속도, B와 K는 상수)
아치볼드 힐(Archibald Hill)은 1938년 근육의 역학적 성질에 대해 연구하여 근육의 수축 속도와 부하 간 관계는 직각쌍곡선이 된다는 것을 밝혀내었고 이를 방정식으로 표현하였다. 이때, P는 근육이 늘어나 줄어들지 않고 지탱할 수 있는 최대 부하이다. 보다 작은 P만큼의 부하가 가해지면, 그에 의해 근육은 수축하게 되며, 열이 발생하게 되는데, 이때 수축하는 속도를 V로 정의한다. a[g] 와 b[cm/sec]는 상수이다. 위와 같은 관계를 정리한 기본 방정식은 다음과 같다.[61]
: ----------식①
이 식에서 변수는 수축 속도 V와 외부에서 가해진 힘 P이다. 위의 식을 전개한 뒤 정리하면 다음과 같이 나타낼 수 있다(c는 상수).
:
힐(Hill)의 기본 방정식을 근육의 생리학적 기작에 적용시켜보면, 전체 근육이 발휘할 수 있는 최대 부하는 Z선을 이루는 각각의 교차결합이 낼 수 있는 힘에 결합의 수를 곱한 것과 같다. 또한, 이때 가해진 힘 P는 각각의 결합에 전해지는 외부의 힘에 결합의 수를 곱한 것으로 표현해볼 수 있다.
: ,
그러나 이때, 모든 결합이 다 작동하는 것이 아니고, 일부의 결합만 작동한다고 할 때, 전체 결합 중 작동하는 결합의 수를 로 정의한다. 결합과 분리의 과정은 마찰력의 존재를 의미하며, 이때 마찰력은 수축 속도에 비례하므로 마찰계수 와 의 곱으로 마찰력을 표현할 수 있다. 따라서, 전체 다리에 의해 발생된 힘은 전해지는 외부의 힘와 마찰력 만큼을 빼고 고려해야 한다. 뉴턴의 제2법칙에 의해 다시 정리를 해보면 다음과 같다.
:
결합에 의해 발생된 힘과 마찰력은 결합수에 비례하므로, 를 한 결합에 의한 마찰계수로 정의할 때 이다. 식을 다음과 같이 변형해 볼 수 있다.
: ----------식②
값은 힘에 선형적으로 의존한다고 보면 다음과 같은 식을 세울 수 있다.[62]
:
이때 인 경우 가 되면서 이 된다. 그러나 이고 수축속도가 최대일 때에는 이 되고 이때의 를 라고하자. 위의 두가지 경우를 식으로 표현해보면 다음과 같다.
:인 경우 :
: 인 경우 :
이 두 식을 다시 식②에 대입하면 다음과 같은 식을 얻을 수 있다.
:
이 결과를 다시 식①에 대입하면 다음과 같은 식을 얻을 수 있다.
:--------식③
그런데 이는 힐(Hill)의 방정식에서 , 로 놓았을 때와 유사한 식이다. 마찬가지로 한 결합에 가해지는 힘이 가장 적을 때, 즉 인 경우 근육은 최대로 수축하게 되는데
:
이 식을 보면 근육에서의 최대 수축속도가 마찰계수와 최대장력으로 표현된 것을 알 수 있다. 이는 힐(Hill)의 방정식이 마찰이 있는 단백질 띠의 움직임, 즉 소성흐름을 묘사한다는 것을 보여준다.[63]
5. 4. 근육과 일
근육이 장력을 발휘하여 물체를 움직이면 일을 했다고 정의한다. 그러나 근육이 수축하는 동안 마찰력이 발생하고 열도 발생한다. 근육에 걸리는 부하가 클수록 더 큰 장력을 발휘해야 하므로, 더 많은 열이 발생한다. 일반적으로 에너지의 약 25%가 일에 사용되고, 나머지 75%는 열로 전환되어[59] 체온 유지 등에 사용된다.6. 평활근
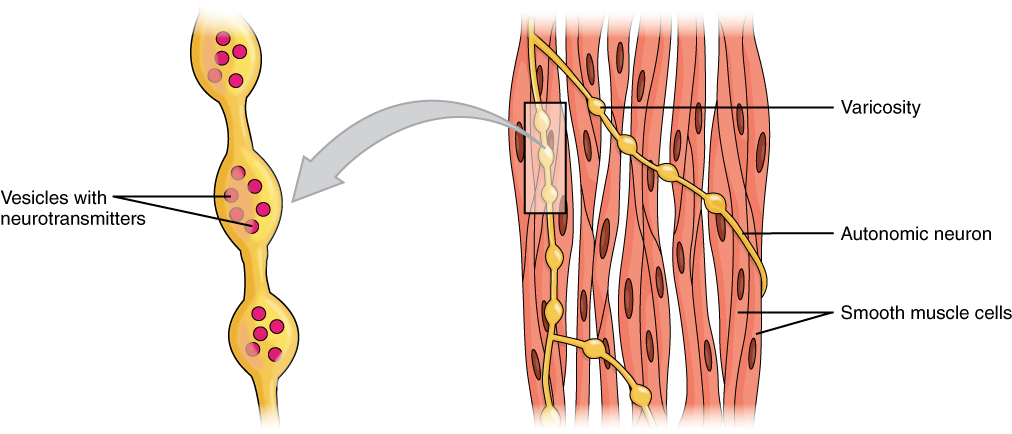
민무늬근(평활근)은 단위형 민무늬근과 다단위형 민무늬근 두 가지 하위 그룹으로 나뉜다. 단위형 민무늬근 세포는 창자와 혈관에서 발견되며, 틈새 연접으로 연결되어 기능적 합포체로 수축한다. 단위형 민무늬근 세포는 자가성으로 수축하며, 자율 신경계에 의해 조절될 수 있다.
단위형 민무늬근 세포와 달리, 다단위형 민무늬근 세포는 눈 근육과 모낭 기저부에서 발견된다. 다단위형 민무늬근 세포는 자율 신경계의 신경에 의해 개별적으로 자극되어 수축하므로, 골격근의 운동 단위 동원과 마찬가지로 섬세한 제어와 점진적인 반응이 가능하다.[1]
6. 1. 평활근의 종류
민무늬근은 단위형 민무늬근과 다단위형 민무늬근의 두 가지 하위 그룹으로 나눌 수 있다. 단위형 민무늬근 세포는 창자와 혈관에서 발견되며, 틈새 연접으로 연결되어 기능적 합포체로 수축할 수 있다. 단위형 민무늬근 세포는 자가성으로 수축하며, 자율 신경계에 의해 조절될 수 있다.
단위형 민무늬근 세포와 달리, 다단위형 민무늬근 세포는 눈 근육과 모낭 기저부에서 발견된다. 다단위형 민무늬근 세포는 자율 신경계의 신경에 의해 개별적으로 자극되어 수축하므로, 골격근의 운동 단위 동원과 마찬가지로 섬세한 제어와 점진적인 반응이 가능하다.
6. 2. 평활근 수축 기전
민무늬근 세포의 수축은 긴장성(지속적) 또는 위상성(일시적)일 수 있으며, 자발적인 전기적 활동, 신경 및 호르몬 입력, 화학적 구성의 국소적 변화 및 신장과 같은 여러 입력의 영향을 받는다.[1] 이는 단일 신경 입력을 사용하는 골격근 세포의 수축 활동과는 대조적이다. 일부 민무늬근 세포는 페이스메이커 전위 또는 느린 파동 전위에 따라 자체적으로 활동 전위를 자발적으로 생성할 수 있다. 이러한 활동 전위는 세포 외 의 유입에 의해 생성되며 에 의해 생성되지 않는다. 골격근과 마찬가지로 세포질 이온도 민무늬근 세포에서 가교 순환에 필요하다.민무늬근 세포에서 세포질 의 두 가지 출처는 칼슘 통로를 통해 들어오는 세포 외 와 근소포체에서 방출되는 이온이다. 세포질 의 증가는 더 많은 가 칼모듈린에 결합하게 하고, 이는 다시 미오신 경쇄 키나아제에 결합하여 활성화시킨다. 칼슘-칼모듈린-미오신 경쇄 키나아제 복합체는 20 킬로달톤(kDa) 미오신 경쇄의 아미노산 잔기-세린 19에서 미오신을 인산화하여 미오신과 액틴의 분자적 상호 작용을 가능하게 하고 수축을 시작하며 미오신 ATP가수분해효소를 활성화한다. 골격근 세포와 달리 민무늬근 세포에는 트로포닌이 없지만, 가는 필라멘트 단백질인 트로포미오신과 칼데스몬 및 칼포닌과 같은 다른 주목할 만한 단백질을 포함한다. 따라서 민무늬근 수축은 골격근과 심근에서처럼 액틴의 미오신 결합 부위를 조절하는 트로포닌 복합체에 대한 결합이 아닌, 미오신의 활성화 인산화에 의해 시작된다.
가교 순환의 종료(및 래치 상태에서 근육 유지)는 미오신 경쇄 포스파타제가 미오신 헤드에서 인산기를 제거할 때 발생한다. 20 kDa 미오신 경쇄의 인산화는 민무늬근의 단축 속도와 잘 관련된다. 이 기간 동안 산소 소비량으로 측정된 에너지 사용량이 급증한다. 시작 후 몇 분 이내에 칼슘 수치가 현저히 감소하고 20 kDa 미오신 경쇄의 인산화가 감소하며 에너지 사용량이 감소한다. 그러나 긴장성 민무늬근의 힘은 유지된다. 근육의 수축 동안 빠르게 순환하는 가교가 활성화된 액틴과 인산화된 미오신 사이에 형성되어 힘을 생성한다. 힘의 유지는 천천히 순환하고 힘을 유지하는 탈인산화된 "래치-브리지"의 결과라고 가정한다. 로 키나아제, DAPK3, 단백질 키나아제 C와 같은 여러 키나아제가 수축의 지속적인 단계에 관여하는 것으로 여겨지며 플럭스는 중요할 수 있다.[33]
6. 3. 신경 조절
민무늬근은 단위형 민무늬근과 다단위형 민무늬근의 두 가지 하위 그룹으로 나눌 수 있다. 단위형 민무늬근 세포는 창자와 혈관에서 발견되며, 틈새 연접으로 연결되어 기능적 합포체로 수축할 수 있다. 단위형 민무늬근 세포는 자가성으로 수축하며, 자율 신경계에 의해 조절될 수 있다.
다단위형 민무늬근 세포는 단위형 민무늬근 세포와 달리 눈 근육과 모낭 기저부에서 발견된다. 다단위형 민무늬근 세포는 자율 신경계의 신경에 의해 개별적으로 자극되어 수축한다. 이처럼, 골격근의 운동 단위 동원과 마찬가지로 섬세한 제어와 점진적인 반응이 가능하다.
민무늬근(평활근) 수축은 근원성이지만, 수축의 속도와 강도는 자율 신경계에 의해 조절될 수 있다. 부교감 신경계의 가지 신경 섬유는 신경 전달 물질 아세틸콜린을 분비하며, 이는 민무늬근 세포의 무스카린성 아세틸콜린 수용체(mAChRs)에 결합한다. 이러한 수용체는 대사성 수용체 또는 G 단백질 연결 수용체로, 2차 전달자 연쇄 반응을 시작한다. 반대로, 교감 신경계의 가지 신경 섬유는 에피네프린과 노르에피네프린 신경 전달 물질을 분비하며, 이는 또한 대사성 수용체인 아드레날린성 수용체에 결합한다. 민무늬근에 미치는 정확한 영향은 활성화된 수용체의 특정 특성에 따라 다르며, 부교감 신경의 입력과 교감 신경의 입력 모두 흥분성(수축) 또는 억제성(이완)일 수 있다.
7. 심근
심근 세포에는 자율 박동성과 수축성의 두 가지 유형이 있다. 자율 박동성 세포는 수축하지 않고, 대신 자율 신경계에 의해 조절될 수 있는 다른 심근 세포의 수축 속도를 설정한다. 반대로, 수축성 근육 세포(심근 세포)는 심장 근육의 대부분을 구성하며 수축할 수 있다.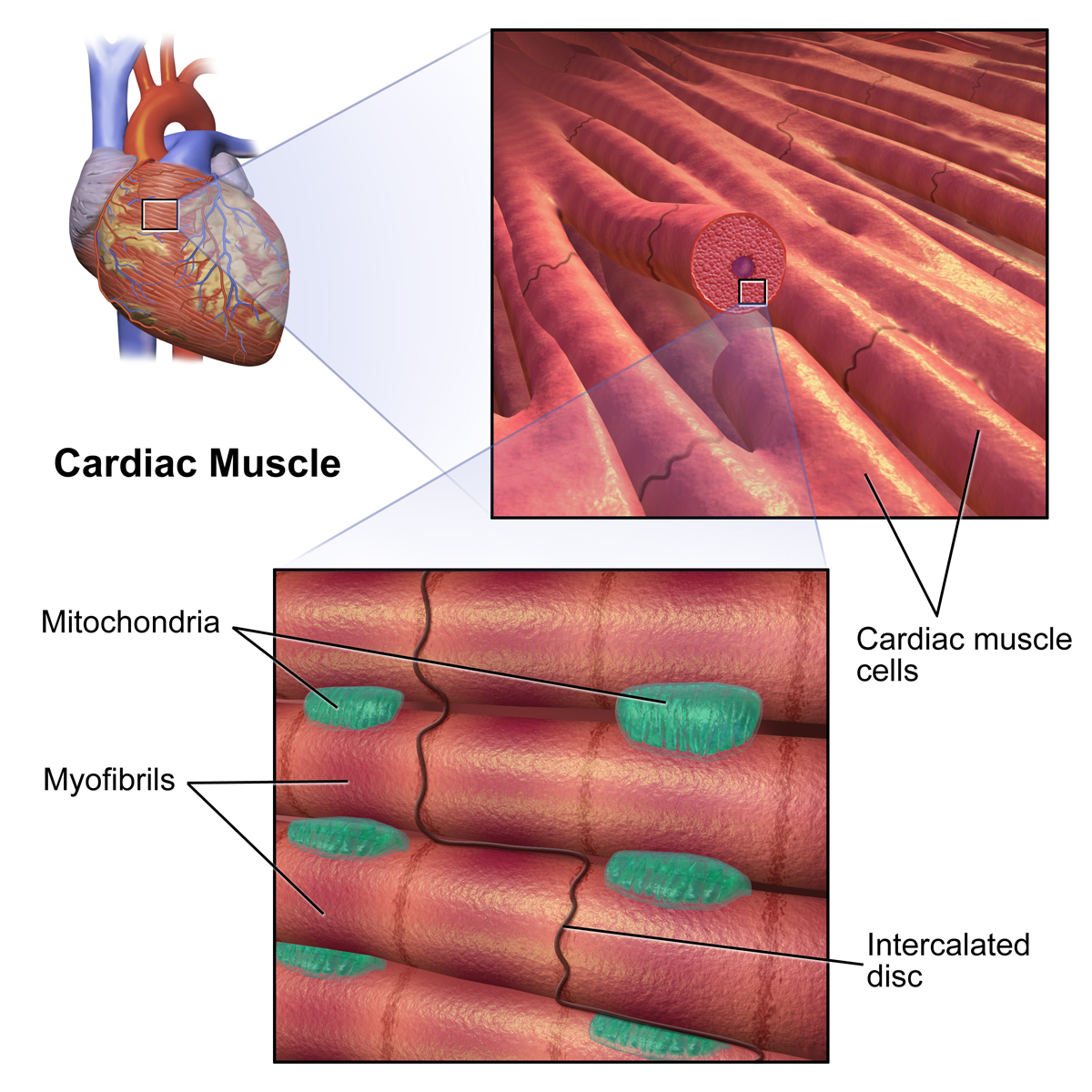
7. 1. 심근 세포의 종류
심근 세포에는 자율 박동성과 수축성의 두 가지 유형이 있다. 자율 박동성 세포는 수축하지 않고, 대신 자율 신경계에 의해 조절될 수 있는 다른 심근 세포의 수축 속도를 설정한다. 반대로, 수축성 근육 세포(심근 세포)는 심장 근육의 대부분을 구성하며 수축할 수 있다.7. 2. 흥분-수축 연접
골격근과 심근 모두 흥분-수축(E-C) 연접에서 탈분극 전도와 Ca2+ 방출 과정이 일어난다. 그러나 관여하는 단백질은 유사하지만 구조와 조절 방식은 다르다. 디히드로피리딘 수용체(DHPR)는 서로 다른 유전자에 의해 암호화되며, 리아노딘 수용체(RyR)는 별개의 이소형이다. 게다가, DHPR은 골격근에서 Ca2+ 방출을 조절하기 위해 RyR1(골격근의 주요 RyR 이소형)과 접촉하는 반면, L형 칼슘 통로(심근세포의 DHPR)와 RyR2(심근의 주요 RyR 이소형)는 심근에서 물리적으로 결합되어 있지 않지만 접합 연접에 의해 서로 마주보고 있다.[34]골격근과 달리, 심근의 E-C 연접은 주로 칼슘 유도 칼슘 방출이라는 메커니즘에 의존하는 것으로 여겨진다.[35] 이는 T-세관과 근소포체 사이의 접합 구조에 기반한다. 정크토필린-2(JPH2)는 이 구조를 유지하는 데 필수적이며, T-세관의 완전성 유지에도 중요하다.[36][37][38] 또 다른 단백질인 수용체 부속 단백질 5(REEP5)는 접합 SR의 정상적인 형태를 유지하는 기능을 한다.[39] 접합 연접의 결함은 이 두 단백질 중 하나의 결핍으로 인해 발생할 수 있다. 칼슘 유도 칼슘 방출 과정에서 RyR2는 L형 칼슘 통로를 통해 Ca2+가 흐르면서 유발되는 칼슘 방아쇠에 의해 활성화된다. 이후 심근은 삼합체보다는 이합체 구조를 나타내는 경향이 있다.
심근 세포의 흥분-수축 연접은 동방 결절 또는 방실 결절의 박동 조절 세포에 의해 활동 전위가 시작되어 갭 연접을 통해 심장의 모든 세포로 전도될 때 발생한다. 활동 전위는 표면 막을 따라 T-세관(후자는 모든 심장 세포 유형에서 보이지는 않음)으로 이동하고 탈분극은 세포외 Ca2+가 L형 칼슘 통로를 통해 세포로 유입되도록 하며, 나트륨-칼슘 교환체(NCX)를 통해 플라토 단계 초기에 유입될 수 있다. 이러한 Ca2+ 유입은 활성화에 필요한 Ca2+의 약 10%에 불과하지만, 골격근의 Ca2+ 유입보다 상대적으로 크다. 이 Ca2+ 유입은 세포내 Ca2+의 작은 국소적 증가를 유발한다. 세포내 Ca2+의 증가는 근소포체 막의 RyR2에 의해 감지되며, 이는 양성 피드백 생리적 반응으로 Ca2+를 방출한다. 이 양성 피드백은 칼슘 유도 칼슘 방출[35]으로 알려져 있으며, 칼슘 스파크(Ca2+ 스파크[40])를 발생시킨다. 약 30,000개의 Ca2+ 스파크의 공간적 및 시간적 합산은 세포 전체의 세포질 칼슘 농도 증가를 가져온다.[41] 세포 막과 근소포체를 통한 칼슘 흐름에 따른 세포질 칼슘의 증가는 칼슘 완충제에 의해 조절되며, 이는 세포내 칼슘의 많은 부분을 결합한다. 그 결과, 총 칼슘의 큰 증가는 자유 Ca2+의 비교적 작은 증가로 이어진다.[42]
세포질 칼슘은 트로포닌 C에 결합하여 액틴 결합 부위에서 트로포미오신 복합체를 이동시켜 미오신 머리가 액틴 필라멘트에 결합하도록 한다. 이 시점부터 수축 기전은 골격근과 본질적으로 동일하다. 간단히 말해, ATP 가수분해를 사용하여 미오신 머리는 액틴 필라멘트를 근절 중앙으로 당긴다.
수축기 이후, 세포내 칼슘은 근소포체/내형질 망 ATPase(SERCA) 펌프에 의해 다음 주기가 시작될 준비가 되도록 근소포체로 다시 흡수된다. 칼슘은 또한 주로 나트륨-칼슘 교환체(NCX)에 의해, 그리고 덜 중요한 정도로는 세포막 칼슘 ATPase에 의해 세포에서 배출된다. 또한 일부 칼슘은 미토콘드리아에 의해 흡수된다.[43] 효소 포스포람반은 SERCA의 브레이크 역할을 한다. 낮은 심박수에서는 포스포람반이 활성화되어 ATPase의 활성을 늦추어 Ca2+가 세포를 완전히 떠날 필요가 없도록 한다. 높은 심박수에서는 포스포람반이 인산화되어 비활성화되어 대부분의 Ca2+을 세포질에서 근소포체로 다시 가져온다. 다시 한 번, 칼슘 완충제는 이러한 Ca2+ 농도 감소를 조절하여 총 칼슘의 큰 변화에 대한 자유 Ca2+ 농도의 비교적 작은 감소를 허용한다. Ca2+ 농도가 감소하면 트로포닌 복합체가 액틴 필라멘트에서 해리되어 수축이 종료된다. 심장이 이완되어 심실이 혈액으로 채워지고 심장 주기가 다시 시작된다.
8. 무척추동물의 근육
무척추동물은 척추동물과 다른 다양한 근육 구조를 통해 근수축을 일으킨다. 환형동물의 환상근과 종주근, 연체동물, 선형동물의 사선 무늬 근육, 고등 곤충의 비동기 근육이 대표적이다.
8. 1. 환형근과 종주근
환형동물인 지렁이와 거머리에서, 환상근과 종주근 세포는 이들 동물의 몸통 벽을 형성하며 이들의 운동을 담당한다.[44] 예를 들어, 흙 속을 이동하는 지렁이의 경우, 환상근과 종주근의 수축이 상호적으로 일어나며, 체강액은 지렁이의 팽압을 유지하여 정수압 골격 역할을 한다.[45] 앞쪽 분절의 환상근이 수축하면 동물의 앞부분이 방사형으로 수축하기 시작하여, 비압축성 체강액을 앞으로 밀어내어 동물의 길이를 늘린다. 결과적으로 동물의 앞쪽 끝이 앞으로 이동한다. 지렁이의 앞쪽 끝이 고정되고 앞쪽 분절의 환상근이 이완되면 종주근 수축의 파동이 뒤쪽으로 지나가면서 동물의 나머지 몸통을 앞으로 당긴다.[44][45] 이러한 환상근과 종주근의 교대적인 수축 파동을 연동 운동이라고 하며, 이는 지렁이의 기어가는 운동의 기반이 된다.8. 2. 사선 무늬 근육 (Obliquely striated muscles)
환형동물, 연체동물, 선형동물과 같은 무척추동물은 사선 무늬 근육을 가지고 있는데, 이는 척추동물의 골격근이나 심장근처럼 가로로 배열되기보다는 두껍고 얇은 필라멘트 띠가 나선형으로 배열되어 있다.[46] 이매패류에서 사선 무늬 근육은 많은 에너지를 사용하지 않으면서 장기간 긴장을 유지할 수 있다. 이매패류는 이러한 근육을 사용하여 껍질을 닫아둔다.8. 3. 비동기 근육 (Asynchronous muscles)
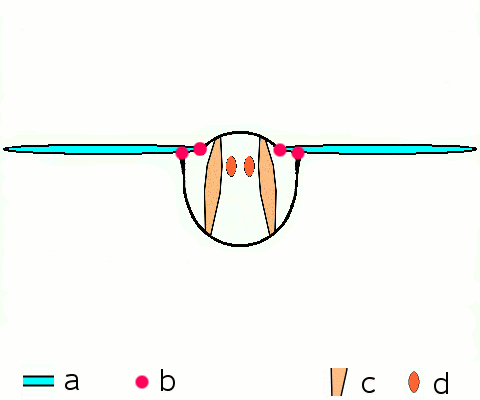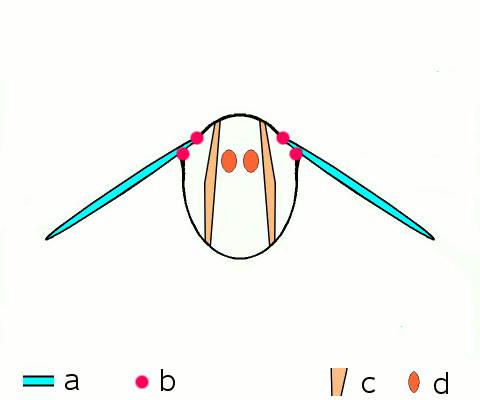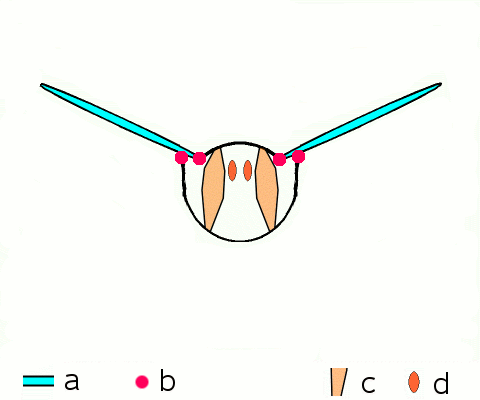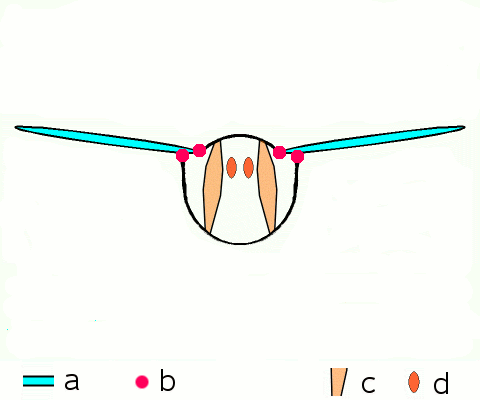
말벌, 파리, 벌, 딱정벌레와 같은 고등 곤충은 비행 근육을 구성하는 비동기 근육을 가지고 있다.[46] 이러한 비행 근육은 두껍고 눈에 띄는 근원 섬유를 포함하고 있기 때문에 종종 ''섬유 근육''이라고 불린다.[47] 비동기 근육의 놀라운 특징은 각 근육 수축에 대한 자극이 필요하지 않다는 것이다. 따라서 이러한 근육의 수축 횟수가 활동 전위의 횟수와 일치하지 않기 (또는 동기화되지 않기) 때문에 ''비동기 근육''이라고 불린다. 예를 들어, 묶인 파리의 날개 근육은 3Hz의 빈도로 활동 전위를 수신할 수 있지만 120Hz의 빈도로 움직일 수 있다.[46] 이러한 고주파수 움직임은 근육이 진동의 고유 주파수로 구동되는 공명 시스템에 연결되어 있기 때문에 가능하다.
9. 근육 연구의 역사
루이지 갈바니는 1780년에 전기 스파크가 죽은 개구리 다리의 근육을 경련시킨다는 것을 발견했다.[49] 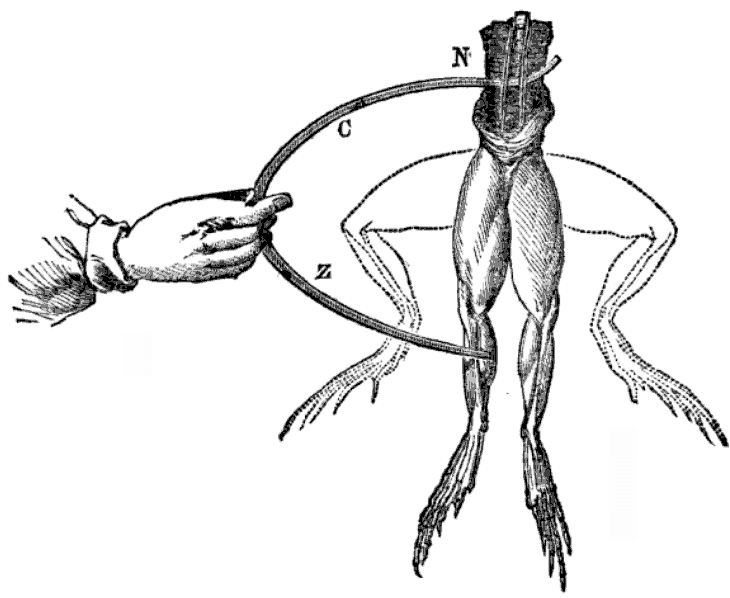
1952년, 흥분-수축 연관이라는 용어가 전기 자극을 기계적 반응으로 변환하는 생리적 과정을 설명하기 위해 만들어졌다.[50] 이 과정은 근육 생리에 기본적이며, 여전히 활발한 연구 분야이다.
9. 1. 19세기 이전
아리스토텔레스는 저서 《동물론(De motu animalium)》에서 근육에 대하여 "움직임의 기능은 미는것과 당기는 것이며, 따라서 움직임의 도구(근육)는 수축과 확장이 가능해야한다. 이것은 바로 생기(pneuma)의 특성이다.”라고 기술하였다.[52]16세기 안드레아스 베살리우스는 로마 시대 해부학자 갈레노스의 생기설을 깨트리고 인체 해부학 개론을 출판했다. 그는 근육을 "동물의 생기를 전달하는 신경의 작용으로 더 두껍고 짧아지며, 한데 모여 자기 자신을 끌어당기고, 또다시 풀어지고 늘어남으로써 근육이 붙어있는 부분을 움직여 원래 상태로 돌아가게 하는 주된 매개자"라고 주장하였다.[53]
17세기 보렐리는 여러 가지 행동에서 근육과 뼈가 어떻게 작용하는지 연구하였고, 이를 일반화하고자 노력하였다. 하지만 근육이 수축할 때 순간적으로 부피가 확장된다고 주장하였다.[54]
9. 2. 19세기: 현미경의 도입
편광현미경이 발달하면서 근육을 더 자세히 볼 수 있게 되었다. 르윈후크는 현재의 Z선을 관찰했고, 이어서 부르케는 현재의 A밴드라고 불리는 띠를 관찰하였다.9. 3. 20세기: 활발한 근수축 연구
아치볼드 힐은 근육에서 생기는 열을 연구하여 수행된 일과 수축 속도의 관련성을 밝히려 했다. 그는 근육 연구를 통해 수축 정도와 하중이 클수록 열이 많이 발생하지만, 열은 수축 속도와 관계가 없음을 발견했다. 이러한 업적으로 1922년 오토 마이어호프와 함께 노벨 생리학·의학상을 수상하였다. 비슷한 시기에 ATP와 근육에서 산소의 역할에 대한 연구가 이루어져 후속 이론의 바탕이 되었다.
1964년 앤드루 헉슬리와 휴 헉슬리가 제안한 활주필라멘트이론은 현재 근수축 이론 중 가장 널리 받아들여지고 있다. 이 이론은 액틴과 미오신 분자 간 미끄러짐으로 활주가 일어나며, 이때의 원동력은 이들 간 교차연결의 움직임에서 온다고 설명한다. 전자현미경의 발달은 근수축 단위의 구조를 명확히 하고, 필라멘트 자체가 수축하는 것이 아님을 직접 확인하는 계기가 되었다.
1952년에는 전기 자극을 기계적 반응으로 변환하는 생리적 과정을 설명하기 위해 흥분-수축 연관이라는 용어가 만들어졌다.[50] 이 과정은 근육 생리에 기본적이며, 전기적 자극(일반적으로 활동 전위)이 근육 수축(기계적 반응)을 유발한다. 흥분-수축 연관은 여러 질병에서 조절 장애가 발생할 수 있으며, 반세기 이상 알려졌음에도 여전히 활발한 연구 분야이다. 일반적인 방식은 활동 전위가 세포막을 탈분극시키고, 근육 유형에 특정한 메커니즘을 통해 세포질 내 칼슘 농도를 증가시킨다(칼슘 과도 현상). 이 칼슘 증가는 ATP를 사용하여 세포 단축을 유발하는 칼슘 민감성 수축성 단백질을 활성화한다.
근육 수축 메커니즘은 오랫동안 과학자들을 곤경에 빠뜨렸으며, 지속적인 연구와 업데이트가 필요하다.[51] 활주 필라멘트 이론은 앤드류 헉슬리와 롤프 니더거케, 휴 헉슬리와 진 한슨이 독립적으로 개발했다. 이들의 발견은 1954년 5월 22일 ''네이처''에 "수축 중 근육의 구조적 변화"라는 주제로 두 편의 연속 논문으로 게재되었다.[23][24]
참조
[1]
서적
Vander's Human Physiology: The Mechanisms of Body Function
McGraw-Hill
2010
[2]
서적
Human Physiology: An Integrated Approach
Pearson
2016
[3]
서적
Animal Locomotion
Oxford University Press
2003
[4]
서적
The Physiology of Excitable Cells
https://archive.org/[...]
Cambridge University Press
1998
[5]
서적
Principles of Medical Physiology
Thieme
2008
[6]
서적
NMS Physiology
Lippincott Williams and Wilkins
2001
[7]
서적
Muscle strength
CRC Press
2008
[8]
논문
Terminology for contractions of muscles during shortening, while isometric, and during lengthening
[9]
웹사이트
Types of contractions
http://muscle.ucsd.e[...]
2006-05-31
[10]
논문
Redox biology of exercise: an integrative and comparative consideration of some overlooked issues
[11]
논문
Effects of eccentric and concentric muscle actions in resistance training
[12]
서적
Exercise Physiology: Human Bioenergetics and Its Applications
Mayfield Publishing Co
[13]
논문
Heavy-load eccentric calf muscle training for the treatment of chronic Achilles tendinosis
http://yaroslavvb.co[...]
[14]
논문
Effectiveness of physical therapy for Achilles tendinopathy: An evidence based review of eccentric exercises
http://iospress.meta[...]
[15]
논문
A randomised clinical trial of the efficacy of drop squats or leg extension/leg curl exercises to treat clinically diagnosed jumper's knee in athletes: pilot study
[16]
서적
Handbook of Psychophysiology
Cambridge University Press
[17]
서적
The Neuron: Cell and Molecular Biology
Oxford University Press
2015-08-19
[18]
문서
Anatomy & Physiology: The Unity of Form and Function
McGraw-Hill Education
2015
[19]
논문
Ryanodine receptors: structure, expression, molecular details, and function in calcium release
https://pubmed.ncbi.[...]
2010-11
[20]
논문
The excitation-contraction coupling mechanism in skeletal muscle
https://pubmed.ncbi.[...]
2014-03
[21]
논문
SMOOTH MUSCLE CONTRACTION AND RELAXATION
https://www.physiolo[...]
2003-12
[22]
서적
Anatomy and Physiology: The Unity of Form and Function
McGraw Hill
[23]
논문
Structural Changes in Muscle During Contraction: Interference Microscopy of Living Muscle Fibres
[24]
논문
Changes in the cross-striations of muscle during contraction and stretch and their structural interpretation
[25]
논문
The positional stability of thick filaments in activated skeletal muscle depends on sarcomere length: evidence for the role of titin filaments.
http://jcb.rupress.o[...]
1987-11
[26]
서적
Principles of Neural Science
McGraw-Hill Medical
2013
[27]
서적
Quantitative Human Physiology: An Introduction
Academic Press
2012
[28]
서적
Textbook Of Medical Physiology
Elsevier
2006
[29]
논문
A brief contraction has complex effects on summation of twitch pairs in human adductor pollicis
https://physoc.onlin[...]
2022-04-05
[30]
논문
A nonstationary model for the Electromyogram
[31]
논문
The variation in isometric tension with sarcomere length in vertebrate muscle fibres
[32]
논문
Muscle co-contraction modulates damping and joint stability in a three-link biomechanical limb
[33]
논문
Tonic and phasic smooth muscle contraction is not regulated by the PKCα - CPI-17 pathway in swine stomach antrum and fundus.
[34]
논문
The network of calcium regulation in muscle
[35]
논문
Calcium-induced calcium release from the cardiac sarcoplasmic reticulum
[36]
논문
Overexpression of junctophilin-2 does not enhance baseline function but attenuates heart failure development after cardiac stress
2014-08-19
[37]
논문
T-tubule remodeling during transition from hypertrophy to heart failure
2010-08-20
[38]
논문
Junctophilins: a novel family of junctional membrane complex proteins
https://pubmed.ncbi.[...]
2000-07
[39]
논문
REEP5 (Receptor Accessory Protein 5) Acts as a Sarcoplasmic Reticulum Membrane Sculptor to Modulate Cardiac Function
2018-02-03
[40]
논문
Calcium sparks: elementary events underlying excitation-contraction coupling in heart muscle
1993-10
[41]
논문
Spatial non-uniformities in {{chem|Ca|2+}}i during excitation-contraction coupling in cardiac myocytes
1994-11
[42]
서적
Excitation-contraction coupling and cardiac contractile force
Kluwer Academic Publishers
2001
[43]
논문
Kinetics, stoichiometry and role of the Na-Ca exchange mechanism in isolated cardiac myocytes
1990-06
[44]
서적
Principles of Life
Sinauer Associates
2014
[45]
논문
The neuronal control of locomotion in the earthworm
[46]
서적
Principles of Animal Locomotion
Princeton University Press
2003
[47]
논문
Asynchronous muscle: a primer
http://jeb.biologist[...]
2000-09-15
[48]
서적
The Science of Common Things: A Familiar Explanation of the First Principles of Physical Science
Ivison & Phinney
1859
[49]
서적
A History of the Theories of Aether and Electricity. Vol 1
Nelson, London
[50]
논문
Excitation-Contraction Coupling in Muscular Response
[51]
논문
Past, Present and Future Experiments on Muscle
2000-04
[52]
서적
힘의원천,근육의 과학과 문화
전파과학사
[53]
서적
힘의원천,근육의 과학과 문화
전파과학사
[54]
서적
힘의원천,근육의 과학과 문화
전파과학사
[55]
서적
동물생리학
라이프사이언스
[56]
서적
운동생리학
금광
[57]
서적
동물생리학
라이프사이언스
[58]
서적
운동생리학
금광
[59]
서적
동물생리학
라이프사이언스
[60]
문서
Muscular force at different speeds of shortening.
[61]
문서
The heat of shortening and dynamic constants of muscle.
[62]
서적
생물물리학
을유문화사
[63]
서적
생물물리학
을유문화사
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com