2025. 4. 14. 오후 8:02:49
뇌 1㎣에 5억개 시냅스…‘불가능’의 뇌 지도 나왔다
출처: 한겨레 ( 한국 / 한국어 )
축삭돌기는 신경계의 주요 전달선으로, 신경 세포체에서 뻗어 나와 다른 뉴런이나 표적 세포로 신호를 전달하는 역할을 한다. 축삭돌기는 말초 신경계에서는 신경을, 중추 신경계에서는 신경로를 형성하며, 길이는 수 마이크로미터에서 수십 cm까지 다양하다. 축삭돌기는 수초의 유무, 직경, 전도 속도 등에 따라 분류되며, 손상 시 왈러 변성과 같은 병변을 유발할 수 있다.
| 축삭돌기 | |
|---|---|
| 개요 | |
| 명칭 | 축삭, 축색돌기 |
| 영어 명칭 | axon, nerve fiber |
| 구조 및 기능 | |
| 정의 | 신경 세포에서 다른 신경 세포로 신호를 전달하는 긴 돌기 |
| 역할 | 신경 전달 물질 전달 |
| 특징 | 하나의 뉴런에 한 개의 축삭 존재 길이는 다양함 |
| 위치 | 뇌 척수 |
| 관련 구조 | 세포체 수상돌기 신경 섬유 말이집 (미엘린 수초) 랑비에르 좁은틈 축삭 말단 |
| 설명 | 축삭은 신경 세포의 주요 구성 요소 중 하나임 축삭은 전기 신호를 다른 신경 세포로 전달하는 역할을 수행함 축삭의 손상은 신경 질환을 유발할 수 있음 |
| 추가 정보 | |
| 관련 연구 | 축삭의 기능과 구조에 대한 연구는 지속적으로 진행 중임 |
| 참고 자료 | 신경 과학 관련 서적 및 논문 |
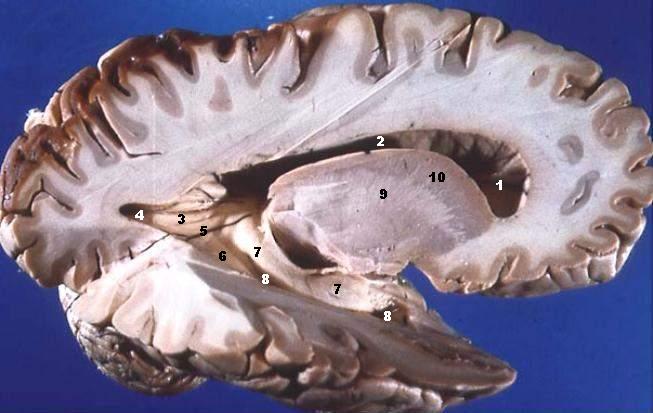
축삭돌기는 신경계의 주요 전달선으로, 다발을 이루어 말초 신경계에서는 신경을 형성하고, 중추 신경계에서는 신경로를 형성한다. 피질 뉴런에서 나온 수초가 있는 축삭돌기는 뇌의 백질을 구성하며, 신경 세포체가 포함된 대뇌 피질의 회백질과 대조적으로 흰색 외관을 띈다. 소뇌에서도 유사한 배열을 보인다. 수초가 있는 축삭돌기 다발은 중추 신경계의 신경로를 구성하며, 뇌의 정중선을 가로질러 반대쪽 영역을 연결하는 곳을 교련 섬유라고 한다. 이 중 가장 큰 것은 두 개의 대뇌 반구를 연결하는 뇌량이며, 약 2천만 개의 축삭돌기를 가지고 있다.[4]

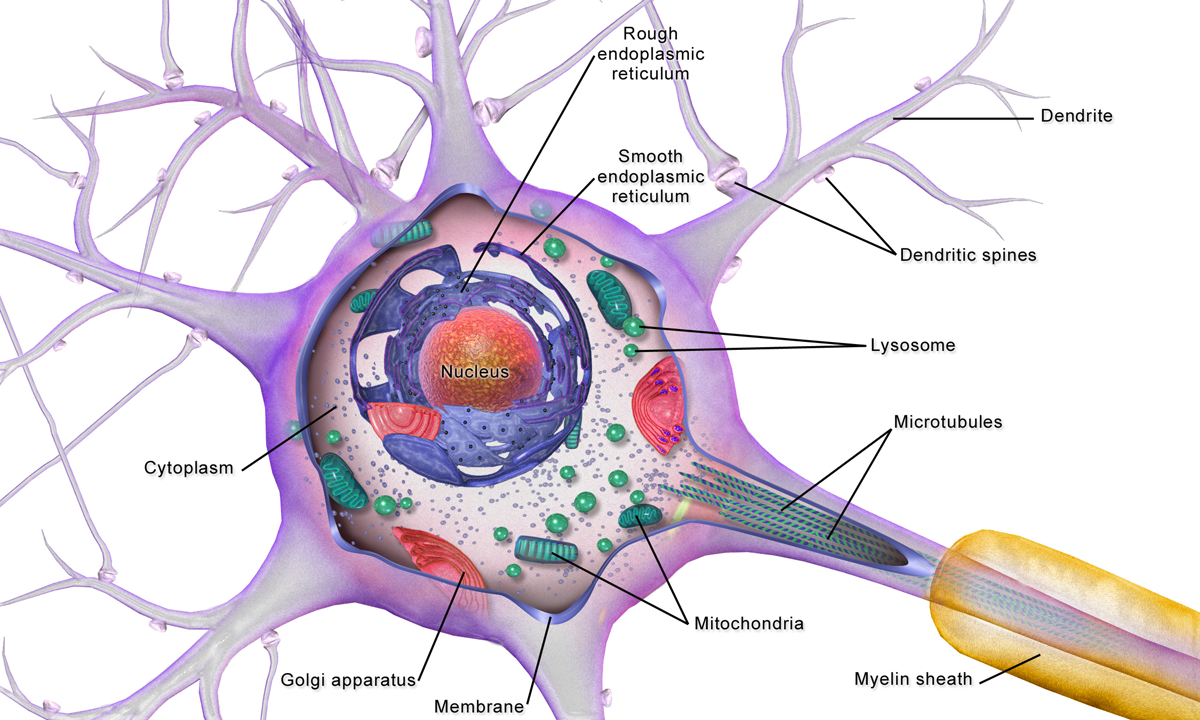

대부분의 축삭돌기는 세포체에서 시작하여 축삭돌기를 따라 빠르게 이동하는 개별적인 전기화학적 임펄스인 활동 전위 형태로 신호를 전달하며, 시냅스 접촉을 하는 지점에서 끝난다.[5]
축삭돌기에서 활동 전위의 전파에 대한 세포 외 기록은 자유롭게 움직이는 동물에서 입증되었다. 장소 세포와 같이 자유롭게 움직이는 동물에서 세포 활동을 연구하기 위해 세포 외 체성 활동 전위가 사용되는 동안, 백색질과 회백질 모두에서 축삭 활동을 기록할 수도 있다. 축삭 활동 전위 전파에 대한 세포 외 기록은 체성 활동 전위와 다음 세 가지 면에서 구별된다.
# 신호는 피라미드 세포 (~500μs) 또는 사이신경세포 (~250μs)보다 짧은 피크-트러프 지속 시간(~150μs)을 갖는다.
# 전압 변화는 3상이다.
# 테트로드에서 기록된 활동은 4개의 기록 와이어 중 하나에서만 보인다.
자유롭게 움직이는 쥐의 기록에서 축삭돌기 신호는 해마 회색질뿐만 아니라 알부스(alvus)와 뇌량과 같은 백색질 트랙에서 분리되었다.[24]
생체 내 활동 전위의 생성은 순차적이며, 이러한 순차적 스파이크는 뉴런의 디지털 코드를 구성한다.[25][26]
축삭돌기는 신경계의 주요 전달선으로, 다발을 이루어 말초 신경계에서는 신경을 형성하고, 중추 신경계 (CNS)에서는 신경로를 형성한다. 일부 축삭돌기는 1미터 이상 뻗을 수 있는 반면, 다른 축삭돌기는 1밀리미터 정도로 짧다. 인체에서 가장 긴 축삭돌기는 각 발의 엄지발가락에서 척수 기저부까지 뻗어 있는 좌골 신경의 축삭돌기이다.
축삭돌기의 직경은 다양하다. 대부분의 개별 축삭돌기는 직경이 현미경적(일반적으로 약 1 마이크로미터 (μm) 정도)이다. 가장 큰 포유류의 축삭돌기는 직경이 최대 20 μm에 달할 수 있다. 신호를 매우 빠르게 전달하도록 특화된 오징어 거대 축삭은 직경이 거의 1밀리미터로, 작은 연필심 크기이다.
축삭돌기의 종말 가지(축삭돌기 끝에 있는 분기 구조)의 수도 신경 섬유마다 다를 수 있다. 중추 신경계의 축삭돌기는 일반적으로 여러 개의 종말 가지를 나타내며, 많은 시냅스 종말점을 갖는다. 이에 비해 소뇌 과립 세포 축삭돌기는 두 개의 병렬 섬유가 뻗어 나오는 단일 T자형 분기 노드가 특징이다. 정교한 분기를 통해 뇌의 단일 영역 내에서 다수의 표적 뉴런에 메시지를 동시에 전달할 수 있다.
신경계에는 수초가 있는 축삭돌기와 수초가 없는 축삭돌기의 두 가지 유형이 있다.[5] 수초는 지방 절연 물질의 층으로, 말초 신경계에서는 슈반 세포가, 중추 신경계에서는 희소돌기아교세포가 형성한다. 수초가 있는 신경 섬유를 따라 랑비에 결절이 균등하게 간격을 두고 발생하며, 이는 도약 전도라고 하는 빠른 전기적 임펄스 전파를 가능하게 한다.
피질 뉴런에서 나온 수초가 있는 축삭돌기는 뇌의 백질을 구성한다. 수초는 신경 세포체가 포함된 대뇌 피질의 회백질과 대조적으로 조직에 흰색 외관을 부여한다. 수초가 있는 축삭돌기 다발은 중추 신경계의 신경로를 구성하며, 반대쪽 영역을 연결하기 위해 뇌의 정중선을 가로지르는 곳을 교련 섬유라고 한다. 이 중 가장 큰 것은 두 개의 대뇌 반구를 연결하는 뇌량이며, 약 2천만 개의 축삭돌기를 가지고 있다.[4]
인간 말초 신경계의 뉴런 축삭은 물리적 특징과 신호 전달 특성에 따라 분류할 수 있다. 조셉 얼랭어와 허버트 스펜서 개서는 축삭의 직경과 신경 전도 속도 사이의 관계를 확립하고, 여러 유형의 신경 섬유를 식별하여 1941년에 축삭의 첫 번째 분류를 제시했다.[3]
축삭은 수초의 유무에 따라 유수 신경 섬유(myelinated axon)와 무수 신경 섬유(unmyelinated axon)로 나뉜다.[5] 수초는 지방 절연 물질 층으로, 슈반 세포와 희소돌기아교세포라는 두 가지 유형의 신경교 세포에 의해 형성된다. 말초 신경계에서는 슈반 세포가, 중추 신경계에서는 희소돌기아교세포가 수초를 형성한다. 수초화는 도약 전도를 가능하게 하여 전기적 신호 전달 속도를 높인다.
말초 신경의 신경 섬유는 수초 유무, 직경, 전도 속도 등에 따라 분류된다. 유수 신경 섬유가 무수 신경 섬유보다 전도 속도가 빠르며, 같은 종류의 신경 섬유 간에서는 직경이 클수록 전도 속도가 빠르다. 이는 유수 신경 섬유의 도약 전도와 전기 긴장 전위의 확산 차이 때문이다.
일반적으로 골격근 운동, 고유 감각, 부위가 확실한 피부 감각은 전도 속도가 빠른 신경 섬유를 이용하고, 교감 신경 활동이나 둔통 등은 전도 속도가 느린 신경 섬유를 이용하여 전달된다.
| 분류 | 수초 | 평균 직경(μm) | 평균 전도 속도(m/s) | 역할 |
|---|---|---|---|---|
| Aα | 유 | 15 | 100 | 골격근 및 힘줄로부터의 감각, 골격근 운동 |
| Aβ | 유 | 8 | 50 | 피부의 촉압각 |
| Aγ | 유 | 8 | 20 | 근방추의 추내근 운동 |
| Aδ | 유 | 3 | 15 | 부위가 비교적 명확한 피부의 온통각 |
| B | 유 | 3 | 7 | 교감 신경의 절전 섬유 |
| C | 무 | 0.5 | 1 | 교감 신경의 절후 섬유, 피부의 온통각 |
감각 신경(구심성 신경)에서는 다른 분류가 사용되기도 한다.
| 분류 | 수초 | 평균 직경(μm) | 평균 전도 속도(m/s) | 감각 |
|---|---|---|---|---|
| Ia | 유 | 15 | 100 | 근방추 |
| Ib | 유 | 15 | 100 | 건 기관 |
| II | 유 | 9 | 50 | 근방추, 피부 촉압각 |
| III | 유 | 3 | 20 | 부위가 비교적 명확한 피부의 온통각 |
| IV | 무 | 0.5 | 1 | 둔통, 내장통 |
| 분류 | 수초 | 평균 직경(μm) | 평균 전도 속도(m/s) | 감각 |
|---|---|---|---|---|
| Ia | 유 | 15 | 100 | 근방추 |
| Ib | 유 | 15 | 100 | 건 기관 |
| II | 유 | 9 | 50 | 근방추, 피부 촉압각 |
| III | 유 | 3 | 20 | 부위가 비교적 명확한 피부의 온통각 |
| IV | 무 | 0.5 | 1 | 둔통, 내장통 |
| 유형 | Erlanger-Gasser 분류 | 직경 (μm) | 수초 | 전도 속도 (m/s) | 관련 감각 수용체 | 고유 수용기 | 기계 수용체 | 통각 수용체 및 온도 수용체 |
|---|---|---|---|---|---|---|---|---|
| Ia | Aα | 13–20 | 예 | 80–120 | 근방추의 1차 수용체 (환나선 종말) | ✔ | rowspan="2" | | rowspan="3" | |
| Ib | Aα | 13–20 | 예 | 80–120 | 골지 힘줄 기관 | |||
| II | Aβ | 6–12 | 예 | 33–75 | 근방추의 2차 수용체 (꽃다발 종말). 모든 피부 기계 수용체 | ✔ | ||
| III | Aδ | 1–5 | 얇음 | 3–30 | 촉각 및 압력의 자유 신경 종말 가쪽 척수 시상로의 통각 수용체 차가운 온도 수용체 | rowspan=2| | ✔ | |
| IV | C | 0.2–1.5 | 아니요 | 0.5–2.0 | 앞 척수 시상로의 통각 수용체 온열 수용체 |
| 유형 | 에를랑거-가서 분류 | 직경 (μm) | 수초 | 전도 속도 (m/s) |
|---|---|---|---|---|
| 신경절 이전 섬유 | B | 1–5 | 예 | 3–15 |
| 신경절 이후 섬유 | C | 0.2–1.5 | 아니요 | 0.5–2.0 |
말초 신경계의 신경 손상은 그 심각도에 따라 신경 차단증, 축삭 절단증, 신경 절단증으로 나눌 수 있다. 뇌진탕은 미만성 축삭 손상의 가벼운 형태로 간주된다.[54] 축삭 손상은 중심성 색소변성을 일으킬 수도 있다. 신경계의 축삭 기능 장애는 말초 및 중추 신경 모두에 영향을 미치는 많은 유전적, 후천적 신경 질환의 주요 원인 중 하나이다.[5]
축삭이 으스러지면, 세포체에서 가장 멀리 떨어진 축삭 부분에서 축삭 변성이라는 활성 과정이 일어난다. 이 변성은 손상 후 빠르게 진행되며, 축삭의 일부는 막에서 밀봉되고 대식세포에 의해 분해된다. 이를 왈러 변성이라고 한다.[55] 축삭의 사멸은 신경 퇴행성 질환에서도 발생할 수 있는데, 특히 축삭 수송이 손상되었을 때 발생하며, 이를 왈러 유사 변성이라고 한다.[56] 연구에 따르면 이 변성은 축삭 단백질인 NMNAT2가 축삭 전체에 도달하지 못해 발생한다고 한다.[57]
축삭의 탈수초화는 다발성 경화증과 같은 질병에서 나타나는 여러 신경학적 증상을 유발한다.
이상 수초 형성은 수초의 비정상적인 형성이다. 이는 여러 백질 이영양증과 조현병과 관련이 있다.[58][59][60]
심각한 외상성 뇌 손상은 신경 경로에 광범위한 병변을 일으켜 미만성 축삭 손상이라는 상태에서 축삭을 손상시킬 수 있다. 이는 지속적 식물 상태로 이어질 수 있다.[61] 쥐를 대상으로 한 연구에서는 단 한 번의 가벼운 외상성 뇌 손상으로 인한 축삭 손상이 반복적인 가벼운 외상성 뇌 손상 후 추가 손상에 대한 민감성을 높일 수 있다는 사실이 밝혀졌다.[62]
신경 유도 도관은 축삭 성장을 유도하여 신경 재생을 가능하게 하는 인공적인 방법이며, 다양한 종류의 신경 손상 치료에 사용된다.
"신경 섬유"라는 용어는 일반적으로 축삭만을 지칭하는 데 사용된다.
오토 프리드리히 카를 다이터스(Otto Friedrich Karl Deiters)는 가지돌기와 구별하여 축삭을 발견한 것으로 알려져 있다.[5] 루돌프 알베르트 폰 쾰리커(Rudolf Albert von Kölliker)와 로베르트 레마크(Robert Remak)는 축삭 초기 분절을 처음으로 확인하고 특징을 규명했다. 쾰리커는 1896년에 축삭이라는 이름을 붙였다.[67] 루이앙투안 랑비에(Louis-Antoine Ranvier)는 축삭에서 발견되는 틈이나 마디를 처음으로 묘사했으며, 이 기여로 인해 이러한 축삭 특징은 현재 랑비에 결절이라고 불린다. 산티아고 라몬 이 카할(Santiago Ramón y Cajal)은 축삭이 뉴런의 출력 구성 요소이며, 그 기능을 설명한다고 제안했다.[5]
조지프 얼랭어(Joseph Erlanger)와 허버트 스펜서 개서(Herbert Spencer Gasser)는 축삭 전도 속도, 미엘린화, 섬유 크기 등을 기반으로 말초 신경 섬유에 대한 분류 시스템을 개발했다.[68] 그들은 1941년에 연구 결과를 발표하여 축삭의 첫 번째 분류를 제시했다. 이후 다른 연구자들의 연구 결과에 따르면 Aa 섬유에는 두 개의 감각 섬유 그룹이 확인되었다.
앨런 호지킨(Alan Hodgkin)과 앤드루 헉슬리(Andrew Huxley)는 오징어 거대 축삭(1939)을 사용했으며, 1952년까지 활동 전위의 이온 기반에 대한 완전한 정량적 설명을 얻어 호지킨-헉슬리 모델을 공식화했다. 호지킨과 헉슬리는 이 연구로 1963년 노벨 생리학·의학상을 공동으로 수상했다.
모델 생물로 자주 사용되는 롱핀 근해 오징어는 가장 긴 축삭을 가진 것으로 알려져 있다.[69] 대왕 오징어는 알려진 것 중 가장 큰 축삭을 가지고 있다. 그 크기는 지름이 0.5mm에서 1mm에 이르며 제트 추진 시스템 제어에 사용된다. 가장 빠른 전도 속도인 210m/s는 일부 원양 보리새우의 피복된 축삭에서 발견되며,[70] 일반적인 범위는 초당 90m에서 200m 사이이다.[71]
[1]
논문
Axons emanating from dendrites: phylogenetic repercussions with Cajalian hues
2014
[2]
논문
Receptive fields, geometry and conduction block of sensory neurones in the central nervous system of the leech
1976-12
[3]
서적
Fundamental neuroscience
Elsevier/Academic Press
2013
[4]
논문
The development of the corpus callosum in the healthy human brain
2010-08
[5]
논문
Axon physiology
https://hal-amu.arch[...]
2011-04
[6]
논문
Axonal Membranes and Their Domains: Assembly and Function of the Axon Initial Segment and Node of Ranvier
2017
[7]
논문
Ankyrin G Membrane Partners Drive the Establishment and Maintenance of the Axon Initial Segment
2017
[8]
논문
The Axon Initial Segment: An Updated Viewpoint
2018-02
[9]
논문
The axon initial segment and the maintenance of neuronal polarity
2010-08
[10]
논문
Axon Initial Segment Cytoskeleton: Architecture, Development, and Role in Neuron Polarity
2016
[11]
논문
Electrogenic tuning of the axon initial segment
2009-12
[12]
논문
Structural and Functional Plasticity at the Axon Initial Segment
2016
[13]
논문
Activity-dependent regulation of excitable axonal domains
2016-03
[14]
논문
Heterogeneity of the Axon Initial Segment in Interneurons and Pyramidal Cells of Rodent Visual Cortex
2017
[15]
서적
Essential cell biology: an introduction to the molecular biology of the cell
https://archive.org/[...]
Garland
2004
[16]
서적
Molecular biology of the cell
Garland
2002
[17]
논문
Oligodendroglial membrane dynamics in relation to myelin biogenesis.
2016-09
[18]
서적
Langman's medical embryology
https://archive.org/[...]
Lippincott William & Wilkins
2010
[19]
논문
The nodes of Ranvier
1952-11
[20]
웹사이트
Medical Definition of bouton
https://www.merriam-[...]
2024-09-21
[21]
서적
Fundamental neuroscience
Elsevier/Academic Press
2013
[22]
논문
Rapid and Reversible Development of Axonal Varicosities: A New Form of Neural Plasticity
2021
[23]
논문
CLARITY reveals a more protracted temporal course of axon swelling and disconnection than previously described following traumatic brain injury
2019-05
[24]
논문
Short duration waveforms recorded extracellularly from freely moving rats are representative of axonal activity
2013-11
[25]
논문
Molecular Brain 4(19), 1~11
2011
[26]
논문
Molecular Brain 7(26):1-16
2014
[27]
논문
Axons amplify somatic incomplete spikes into uniform amplitudes in mouse cortical pyramidal neurons
2010-07
[28]
서적
Principles of development
Oxford University Press
2015
[29]
논문
The establishment of polarity by hippocampal neurons: the relationship between the stage of a cell's development in situ and its subsequent development in culture
1989-12
[30]
논문
Axon formation: fate versus growth
2005-05
[31]
논문
Experimental observations on the development of polarity by hippocampal neurons in culture
1989-04
[32]
논문
Mechanical tension can specify axonal fate in hippocampal neurons
2002-11
[33]
논문
Neuronal polarity: from extracellular signals to intracellular mechanisms
2007-03
[34]
문서
Neuroglia and pioneer neurons express UNC-6 to provide global and local netrin cues for guiding migrations in C. elegans
[35]
논문
The netrins define a family of axon outgrowth-promoting proteins homologous to C. elegans UNC-6
1994-08
[36]
논문
A ligand-gated association between cytoplasmic domains of UNC5 and DCC family receptors converts netrin-induced growth cone attraction to repulsion
1999-06
[37]
논문
The unc-5, unc-6, and unc-40 genes guide circumferential migrations of pioneer axons and mesodermal cells on the epidermis in C. elegans
1990-01
[38]
논문
Trk receptors: roles in neuronal signal transduction
[39]
논문
Asymmetric membrane ganglioside sialidase activity specifies axonal fate
2005-05
[40]
논문
The role of local actin instability in axon formation
1999-03
[41]
논문
The axonal glycoprotein TAG-1 is an immunoglobulin superfamily member with neurite outgrowth-promoting activity
1990-04
[42]
서적
Molecular biology of the cell
2015
[43]
논문
Laser-based single-axon transection for high-content axon injury and regeneration studies
[44]
논문
Nogo and axon regeneration
2004-02
[45]
논문
Macrophages promote axon regeneration with concurrent neurotoxicity
2009-03
[46]
논문
Kinesin-5 regulates the growth of the axon by acting as a brake on its microtubule array
2007-09
[47]
논문
A motor-driven mechanism for cell-length sensing
2012-06
[48]
논문
Delayed feedback model of axonal length sensing
2015-05
[49]
논문
A frequency-dependent decoding mechanism for axonal length sensing
[50]
논문
Sound of an axon's growth
2019-05
[51]
논문
Properties of fast and slow motor units in hind limb and tail muscles of the rat
1972-04
[52]
논문
Axonal conduction velocity changes following muscle tenotomy or deafferentation during development in the rat
1980-01
[53]
서적
Human Physiology
Oxford University Press
[54]
웹사이트
Traumatic Brain Injury (TBI) - Definition, Epidemiology, Pathophysiology
https://emedicine.me[...]
2017-08-16
[55]
웹사이트
Trauma and Wallerian Degeneration
http://missinglink.u[...]
[56]
논문
Wallerian degeneration, wld(s), and nmnat
2010-06-01
[57]
논문
Endogenous Nmnat2 is an essential survival factor for maintenance of healthy axons
2010-01
[58]
논문
Perturbed interactions of mutant proteolipid protein/DM20 with cholesterol and lipid rafts in oligodendroglia: implications for dysmyelination in spastic paraplegia
2006-11
[59]
서적
N-Acetylaspartate
[60]
논문
Further evidence for altered myelin biosynthesis and glutamatergic dysfunction in schizophrenia
2007-08
[61]
웹사이트
Brain Injury, Traumatic
http://www.medcyclop[...]
General Electric
2018-06-20
[62]
논문
Repeated mild traumatic brain injuries induce persistent changes in plasma protein and magnetic resonance imaging biomarkers in the rat
2019-10
[63]
웹사이트
nerve fiber
https://www.merriam-[...]
2023-04-21
[64]
웹사이트
nerve fibre
https://www.oed.com/[...]
2023-04-21
[65]
웹사이트
nerve fiber
https://www.tabers.c[...]
2023-04-21
[66]
웹사이트
nerve fiber
https://dictionary.a[...]
American Psychological Association
2023-04-21
[67]
서적
Origins of neuroscience: a history of explorations into brain function
Oxford University Press
[68]
논문
The 1932 and 1944 Nobel Prizes in physiology or medicine: rewards for ground-breaking studies in neurophysiology
2006-12
[69]
서적
The Brain, the Nervous System, and Their Diseases [3 volumes]
https://books.google[...]
ABC-CLIO
2014-12-16
[70]
논문
Fenestration in the myelin sheath of nerve fibers of the shrimp: a novel node of excitation for saltatory conduction
1996-07
[71]
학술지
Myelination
2016-10
[72]
웹사이트
신경섬유vs.신경섬유――세계대전하의 신경전(상) - 말씀 Magazine:아사히 신문 디지털
http://www.asahi.com[...]
2023-02-01
[73]
웹사이트
DNA From The Beginning, section 6: Genes are real things., "Amination" section, final slide
http://www.dnaftb.or[...]
( 최근 20개의 뉴스만 표기 됩니다. )
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com