발생생물학
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
발생생물학은 생명체의 발생 과정을 연구하는 학문으로, 고대 그리스 시대부터 닭 배아 연구를 시작으로 발전해왔다. 전성설과 후성설의 논쟁을 거쳐, 19세기 실험 발생학의 발달과 세포설의 정립을 통해 급격한 발전을 이루었다. 실험 발생학의 발전은 빌헬름 루, 한스 슈페만 등의 실험을 통해 이루어졌으며, 유전학과의 갈등을 겪으며 20세기 유전자형과 표현형의 관계를 연구하는 학문으로 자리 잡았다.
사람 발생에 대한 연구는 고대부터 종교적, 의학적인 이유로 이루어졌으며, 다양한 문화권에서 태아의 발생 과정을 기록하고 관찰했다.
발생 과정은 수정, 난할, 낭배 형성, 기관 형성, 변태, 생식세포 형성 등의 단계를 거치며, 세포 분화, 패턴 형성, 형태 형성, 성장, 그리고 타이밍의 조절이 주요 개념으로 다루어진다. 최근에는 진화발생생물학 분야에서 발생과 진화의 관계를 연구하며, 유전학, 해부학 등 다양한 분야와 관련된다.
더 읽어볼만한 페이지
- 발생생물학 - 내배엽
내배엽은 동물 배아 발생 시 가장 안쪽에 위치한 층으로 소화관, 간, 췌장, 폐 등 주요 기관을 형성하며, 낭배 형성 과정에서 다양한 신호 전달 경로를 통해 분화되고 줄기세포 연구에 활용되며, 식물에서는 뿌리의 피질 가장 안쪽 세포층을 의미한다. - 발생생물학 - 배엽
배엽은 동물의 초기 발생 과정에서 나타나는 외배엽, 중배엽, 내배엽의 세 가지 세포층으로, 각 층은 특정 기관과 조직으로 분화하며 동물의 발생과 진화를 이해하는 데 중요하다. - 생물학에 관한 - 해부학
해부학은 생물체의 구조와 구성 요소를 연구하는 학문으로, 육안 해부학과 현미경 해부학으로 나뉘며, 인체 해부학 외에도 동물, 식물, 미술 해부학 등 다양한 분야가 존재한다. - 생물학에 관한 - 죽음
죽음은 생명 활동의 영구적 종식으로 의학, 법, 사회, 종교, 심리, 생물학 등 다양한 관점에서 해석되며, 전통적인 심폐사 외에 뇌사도 죽음으로 인정되고, 개발도상국은 전염병, 선진국은 노화 관련 질병이 주요 사망 원인이며, 문화와 종교에 따라 다양한 이해와 관습이 존재하고, 수명 연장, 냉동 보존술, 존엄사, 안락사 등에 대한 논의가 이루어지고 있다.
| 발생생물학 | |
|---|---|
| 개요 | |
| 학문 분야 | 생물학 |
| 연구 대상 | 동물 식물 |
| 관련 학문 | 유전학 세포생물학 분자생물학 진화생물학 생화학 생리학 해부학 생태학 기형학 |
| 정의 | |
| 정의 | 발생 과정과 메커니즘을 연구하는 생물학 분야 |
| 연구 내용 | |
| 주요 연구 내용 | 세포 분화 형태 형성 성장 생식 재생 진화 환경 |
| 역사 | |
| 초기 발생생물학 | 아리스토텔레스의 발생 관찰 |
| 19세기 발생생물학 | 현미경 기술 발전 세포 이론 등장 진화론 등장 |
| 20세기 발생생물학 | 유전학과 통합 분자생물학과 통합 |
| 주요 개념 | |
| 세포 분화 | 세포가 특정 기능을 수행하도록 특화되는 과정 |
| 형태 형성 | 세포와 조직이 특정 형태를 갖추도록 배열되는 과정 |
| 유도 | 한 세포 또는 조직이 다른 세포 또는 조직의 발달에 영향을 미치는 과정 |
| 패턴 형성 | 공간적으로 조직된 세포 활동을 만드는 과정 |
| 연구 방법 | |
| 실험적 방법 | 유전자 조작 세포 이식 조직 배양 |
| 관찰적 방법 | 현미경 관찰 영상 기법 |
| 응용 분야 | |
| 의학 | 선천성 기형 연구 암 연구 재생 의학 |
| 농업 | 작물 개량 동물 생산성 향상 |
| 참고 문헌 | |
| 도서 | 제목: Developmental Biology 성: Gilbert 이름: Scott F. 성2: Barresi 이름2: Michael J. F. 날짜: 2016 판: 11th ed 출판사: Sinauer associates, Inc., publishers 위치: Sunderland (Mass.) 쪽: 1-2, 9-11, 785 isbn: 978-1-60535-470-5 제목: Principles of development 날짜: 2019 판: 6th ed 출판사: Oxford university press 위치: Oxford, United Kingdom 쪽: 8-11 isbn: 978-0-19-880056-9 |
2. 역사
고대 그리스 시대부터 히포크라테스나 아리스토텔레스 등에 의해 닭의 배아를 사용한 연구가 이루어졌다. 아리스토텔레스는 다른 동물에 대해서도 관찰했으며, 《동물지》, 《동물부분론》, 《동물 발생론》 등에 그 고찰을 볼 수 있다. 이 시기에 발생의 원리에 대해 전성설과 후성설이 일어났다. 아리스토텔레스는 후성설을 지지했지만, 이는 소수파에 속했고 오랫동안 전성설이 더 힘을 가졌다. 이는 난자의 개념에 혼란이 있었고, 곤충의 번데기나 식물의 씨앗도 난자와 동일시되었던 점도 크다.
윌리엄 하비는 포유류의 발생 초기를 관찰하고 난자를 발견할 수는 없었지만, 그 존재를 확신하고 '모든 것은 난자에서'라는 말을 남겼다.[56][57] 하지만 생물 연구에 현미경이 사용되게 된 후에도 오랫동안 난자가 발견되지 않았다. 정자의 발견은 전성설 옹호자를 난자론자와 정충론자로 나누게 되었다.
2. 1. 사람 발생에 대한 관찰
사람의 발생은 고대 시기부터 문화권을 막론하고 관심의 대상이 되었는데, 종교적인 이유가 많았다. 태반과 탯줄을 신성시하는 문화가 고대 이집트를 비롯한 여러 문명에서 발견되며[63], 한반도에도 삼국 시대부터 태실 등을 만들어 태반을 보관하는 안태(安胎) 풍습이 있었다.[64] 인도 문명의 경우 일찍부터 힌두교 및 불교 경전에 사람 발생 과정을 상세히 기록하였다. 기원전 1천년경에 저술된 것으로 추정되는 우파니샤드의 하나인 《가르브하 우파니샤드(Garbha Upaniṣadsa)》에는 사람 태아의 발생 과정이 “하룻밤이 지났을 때에는 물과 같고, 이렛날에는 거품과 같으며, 보름이 지나면 덩어리가 된다.” 이와 같이 대략적으로 기술되어 있으며[65], 바가바드 기타에는 양막과 같은 구조가 언급되어 있다.[67] 기원후에 저술된 것으로 보이는 《불설포태경(佛說胞胎經, Garbhāvakrāntinirdeśasa, Garbhāvakrāntisūtrasa)》은 부처가 아난타에게 임신 38주에 걸친 태아의 변화를 상세히 설명하는 내용을 담고 있는데[66], 특히 임신 5-8주째에 사지가 발생하는 시기와 순서를 오늘날의 기준으로 볼 때에도 정확하게 기술하였다.[65]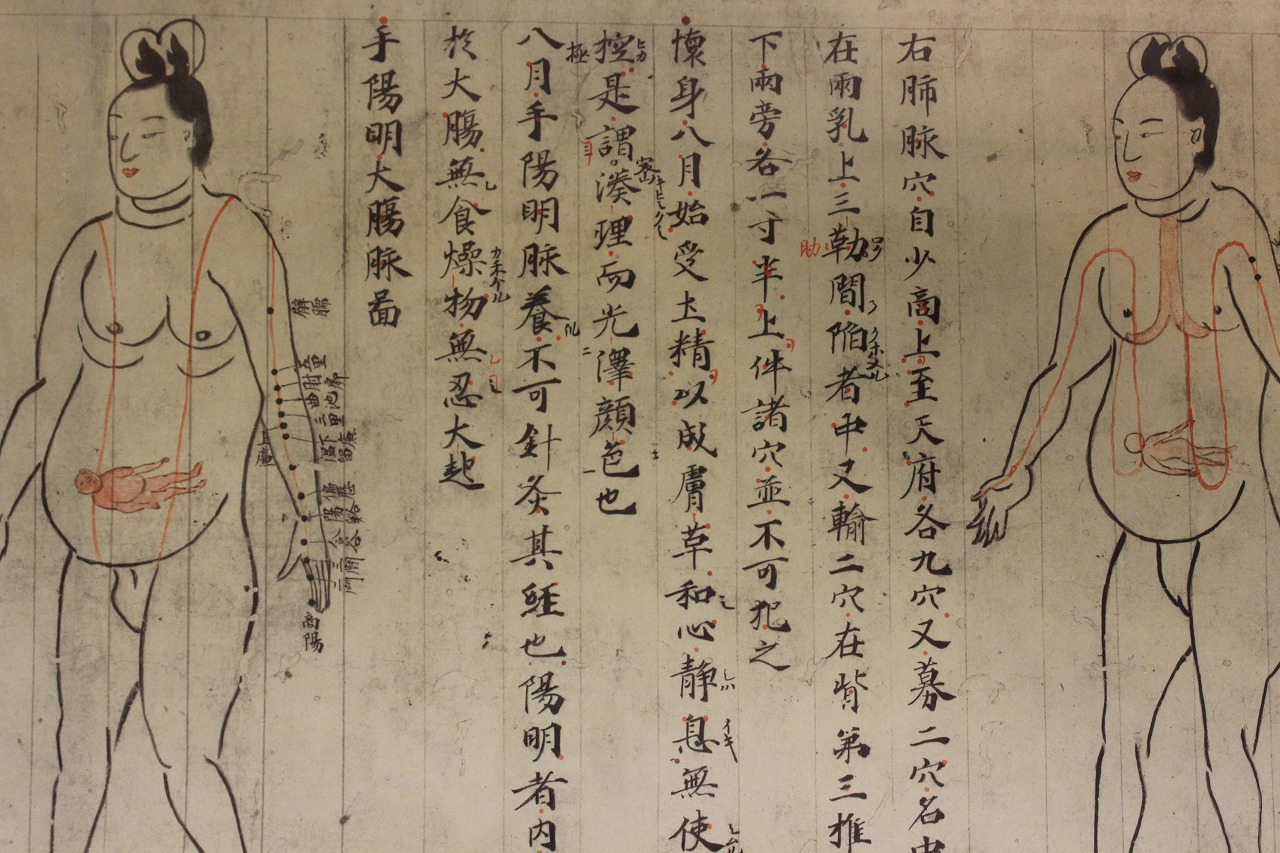
사람 발생에 대한 연구는 산과적인 이유로도 이루어졌다. 인도 아유르베다의 핵심 문헌인 《수슈루타 삼히타(Suśrutasaṃhitāsa)》와 《차라카 삼히타(Carakasaṃhitāsa)》는 임산부의 건강 유지를 도울 목적으로 임신·발생 과정에 대한 기술과 설명을 소개하였다.[65][67] 특히 동아시아에서 주로 종교보다는 의학적인 동기로 발생에 대한 기록을 남겼는데, 그림을 함께 싣곤 했다는 점이 특징이었다. 전한 시대의 무덤 마왕퇴에서 발견된 백서 중 하나인 《태산서(胎産書)》에는 열 달 동안 태아의 변화가 기술되어 있으며, 임산부를 위한 건강 지침과 더불어 임산부의 몸 형태 그림이 실려 있다.[65] 10세기에 중국 의서를 종합하여 저술된 일본 의서 《의심방(醫心方)》은 임산부에게 침술을 시행할 때 도움을 줄 목적으로 매달 임산부의 변화를 나타낸 그림을 수록하였는데, 여기에 열 달 동안 배아가 차츰 태아로 자라나는 모습을 함께 나타내었다.[65]

티베트에서는 9세기경 《가르브하 우파니샤드》가 번역된 이래 사람 발생 순서에 대해 다양한 의견이 제시되었다. 14세기 롱첸파(Longchenpa)는 사지가 임신 8주경에 발생한다고 정확하게 서술하였다. 티베트의 섭정이었던 상예갸초(Sangye Gyatso)는 1670년 티베트 전통의학의 핵심 문헌인 《사부의전(四部醫典, rGyud-bZhibo, the Four Tantras)》을 개정할 때 내용을 한결 효과적으로 전달하기 위해 77개의 그림을 수록하였다. 그 중 다섯 번째 삽화에서 두 화가(Lhobrag Norbu Gyatso, Lhasawa Genyen)는 사람 발생의 전체 과정을 순서대로 나타내었다.[65]
고대 그리스의 소크라테스 이전 철학자들도 사람 배아의 형성 과정을 탐구했다. 엠페도클레스는 여러 인체 부위의 조성을 사원소설을 바탕으로 설명했으며, 아낙사고라스, 알크마이온, 디오게네스 등은 여러 인체 부위가 생겨나는 순서에 대해 다양한 의견을 제기했다.[68] 기원전 5세기경 히포크라테스 학파는 주로 산과적인 문제와 연관 지어 사람 발생을 논의했으나, 사람 발생에서 다른 동물의 발생으로까지 시선을 넓혔다. 탯줄의 구조와 출산의 원인 등 다양한 주제에 대한 의견을 개진하는 문헌인 《발생론(Περὶ γονῆςgrc, On generation)》에는 달걀을 깨서 관찰한 내용을 근거로 제시하는 부분이 있다.[69] 아리스토텔레스 역시 사람뿐만 아니라 여러 동물의 발생에 관심을 가졌다. 그는 《동물발생론(Περὶ ζῴων γενέσεωςgrc, On the generation of animals)》에서 발생 과정을 동물 분류의 기준으로 삼을 요량으로 여러 동물의 생활사를 비교하여 서술했으며, 난생, 태생, 난태생을 구분하였다. 난할 방식을 완전(holoblastic) 난할과 불완전(meroblastic) 난할로 구분하기도 했다.[59]
2. 2. 전성설-후성설 논쟁
히포크라테스 전집에는 남자의 정액과 여자의 월경혈이 만나 발효와 유사한 과정을 거쳐 개체가 만들어진다는 주장이 실려 있다.[69][70] 아리스토텔레스는 성숙한 개체에서 보이는 구조들이 눈에 보이지 않을 뿐 처음부터 미리 만들어져 있다는 전성설과, 발생 과정에서 새로운 구조가 차츰 생겨난다는 후성설을 대조했으며 후성설을 옹호했다.[71] 이후 아리스토텔레스주의자들은 배아에 대한 세심한 관찰보다는 아리스토텔레스가 남긴 저작을 근거로 후성설을 계속 받아들였다.[72]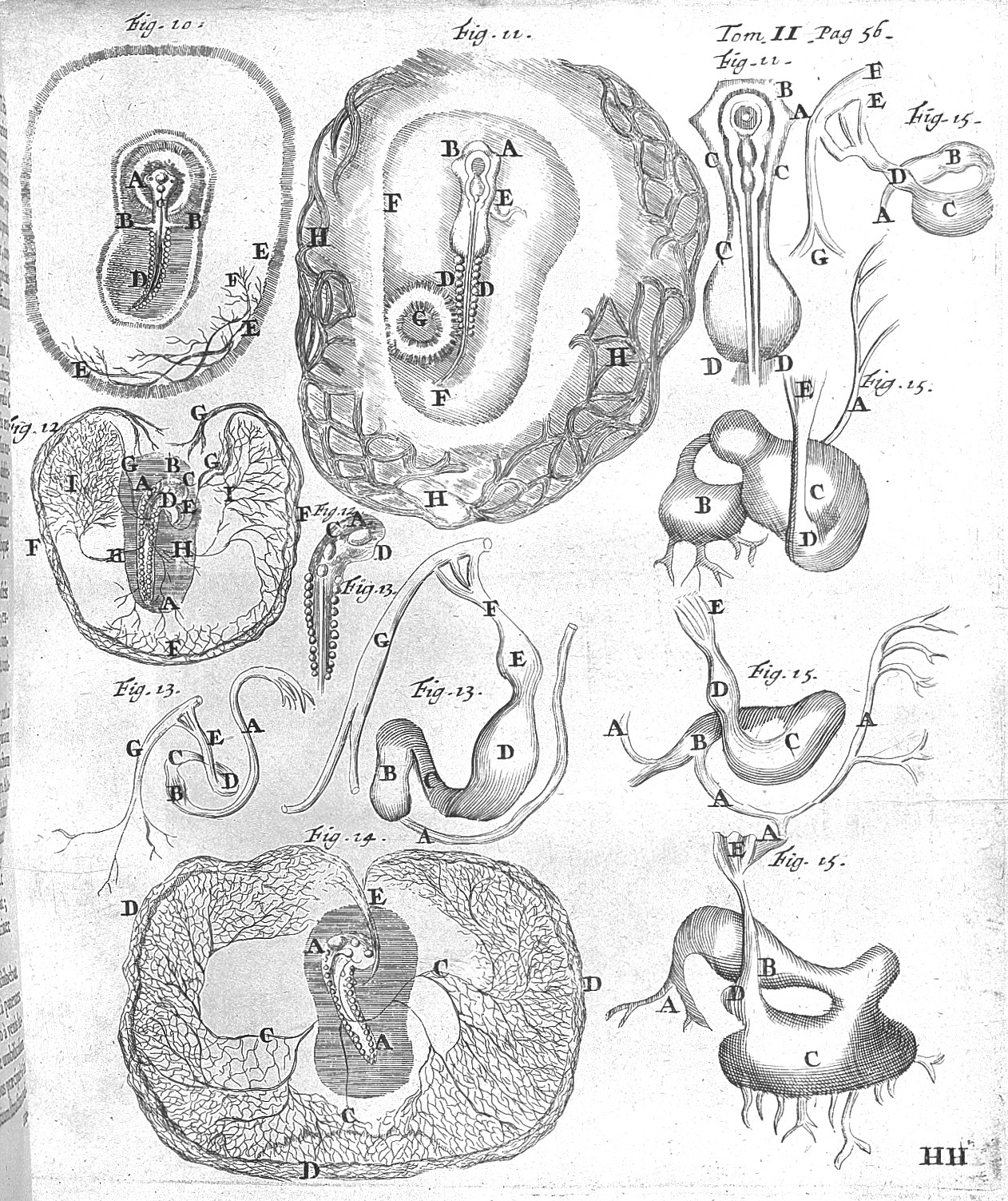
1651년 윌리엄 하비는 모든 생물이 알, 즉 난자로부터 생겨난다고 주장했으며, 닭의 배아에서 최초로 배반엽을 관찰하였다. 1672년 마르첼로 말피기는 최초로 닭의 발생 과정을 현미경으로 관찰한 기록을 출판하였다. 말피기는 발생이 진행되기 이전 상태의 달걀에 제법 발달된 구조들이 이미 존재하는 것을 보고 전성설을 주장했다.[59] 전성설-후성설 논쟁은 18세기 내내 이어졌다.[71] 카스파르 프리드리히 볼프 등은 발생 도중에 나타나는 구조가 성체에서 관찰되는 구조에 직접 대응하지 않는다는 것을 근거로 후성설을 주장했으나, 그 기제를 설명하기 위해서는 생기론에 의지해야만 했다.[59] 대체로 유물론을 근거로 사고한 학자들이 전성설을, 경험과 관찰에 근거해서 결론을 도출해야 한다고 믿었던 학자들이 후성설을 지지하는 경향이 있었다.[73]
고대 그리스 시대부터 이미 히포크라테스나 아리스토텔레스 등에 의해 닭의 배아를 사용한 연구가 이루어졌다. 아리스토텔레스는 다른 동물에 대해서도 관찰했으며, 《동물지》, 《동물부분론》, 《동물 발생론》 등에 그 고찰을 볼 수 있다. 이 시기에 발생의 원리에 대해 전성설과 후성설이 일어났다. 아리스토텔레스는 후성설을 지지했지만, 이는 소수파에 속했고 오랫동안 전성설이 더 힘을 가졌다. 이는 난자의 개념에 혼란이 있었고, 곤충의 번데기나 식물의 씨앗도 난자와 동일시되었던 점도 크다.
윌리엄 하비는 포유류의 발생 초기를 관찰하고 난자를 발견할 수는 없었지만, 그 존재를 확신하고 '모든 것은 난자에서'라는 말을 남겼다.[56][57] 하지만 생물 연구에 현미경이 사용되게 된 후에도 오랫동안 난자가 발견되지 않았다. 정자의 발견은 전성설 옹호자를 난자론자와 정충론자로 나누게 되었다.
2. 3. 진화론과의 만남
찰스 다윈 이전에도 여러 생물종의 성체 사이에 어떤 관계가 있는지에 대한 설명 방식이 있었으며, 서로 다른 생물종의 배아발생 과정의 차이점을 해명하는 것은 많은 학자들의 관심 주제였다. 이에 대한 한 가지 대답은 고등동물의 발생이 하등한 동물의 성체 단계를 거쳐 가는 과정이라는 발생반복설이었다.[76] 카를 폰 베어는 이를 비판하며 다음 네 가지 법칙을 제시했다.[77]# 배아에서 동물군의 일반적인 특징이 특수한 특징보다 더 일찍 나타난다.
# 가장 일반적인 특징으로부터 덜 일반적인 특징이 발달하며, 이같이 되풀이한 끝에 가장 특수한 특징이 나타난다.
# 특정한 종의 배아는 다른 동물들을 거쳐 가는 것이 아니라 오히려 그로부터 점점 더 멀어진다.
# 그러므로 고등한 동물의 배아는 절대 그보다 하등한 동물과 같지 않고, 다만 그 배아와 같을 뿐이다.
폰 베어의 법칙은 주의 깊은 관찰을 근거로 한 것이었지만, 그러한 경향이 성립하는 이유에 대한 설명은 다소 형이상학적이었다.[77] 다윈의 진화론은 생물종 사이의 관계를 이해하는 새로운 관점을 제공해 주었으며, 배아발생의 차이점을 설명하는 문제는 '개체발생(ontogeny)과 계통발생(phylogeny)'의 관계를 규명하는 문제로 재해석되었다. 특히 적응으로 말미암아 형태가 다양하게 변화하기 마련인 성체에 비해 배아는 선택압에 덜 노출되어 있을 것이므로, 배아 형태 비교가 계통 사이의 연관성을 밝히기에 유용할 것으로 기대되었다. 다윈 자신도 《종의 기원》에서 따개비의 기원을 조사할 때 발생 초기 형태의 유사성을 단서로 삼았다.[78]
비교해부학과 발생학을 동원하여 생명의 계통수를 추적한다는 구상을 실행에 옮긴 것은 진화론의 열렬한 지지자였던 에른스트 헤켈이었다.[79] 다만 개체발생과 계통발생의 관계에 대한 입장에는 차이가 있었다. 다윈은 폰 베어에 동의했던 반면,[78] 헤켈은 발생반복설을 재해석하여 “개체발생은 계통발생을 되풀이한다”고 주장하였고 이를 '생물발생법칙(biogenetic law)'이라 명명했다. 헤켈은 진화론에 근거한 생물발생법칙이야말로 기독교 철학이나 생기론에 바탕을 두었던 기존의 설명과 달리 발생의 원인에 대한 기계론적·역학적(mechanical) 이해를 가능하게 해 준다고 믿었다. “계통발생은 개체발생의 역학적 원인이다.” 하고 그는 주장하였다.[80] 다윈 이후 적어도 두 세대의 동물학자들이 헤켈의 연구 프로그램을 받아들여 발전시켰다.[81]
발생 과정에 대한 연구는 현미경 관찰이 이루어지면서부터 발전했다. 특히 무척추동물 각 군의 발생에 관한 지식 축적으로부터, 동물의 발생에는 기본적인 유형이 있으며, 많은 동물의 발생에는 공통된 특징이 있다는 것을 알게 되었다. 이 분야는 비교 발생학이라고 하며, 19세기에 성립하여 비교 해부학과 함께 비교 형태학이라고 불렸다. 에른스트 헤켈은 그것들을 종합하여 진화적인 의미를 발견했다. 그는 각 군의 동물의 발생이, 그 동물이 거쳐온 진화의 과정을 간략화한 것이라고 생각했다. 이 사실은 '개체 발생은 계통 발생을 반복한다'는 표현으로 알려져 있으며, 이를 '반복설'이라고 한다.
2. 4. 실험적 발생학의 발전
헤켈의 주장에 반대하며, 젊은 발생학자들은 발생의 "어떻게?"에 더 주목했다. 이들은 물리학처럼 실험을 통해 생물학을 정밀과학으로 만들고자 했다.[83] 에티엔 조프루아 생틸레르 등의 선구자도 있었지만, 헤켈의 연구에 대한 비판과 함께 발생학에 실험적 방법론이 본격적으로 도입되었다.[84]빌헬름 히스는 발생 과정의 복잡한 형태 변화가 배아 각 부분의 성장률 차이로 인한 물리적 압박 때문이라고 보았다. 그는 탄력적인 고무관을 변형했을 때 나타나는 모양이 배아 기관과 유사하다는 관찰 결과를 제시했다.[82]
빌헬름 루와 한스 드리슈는 헤켈의 제자였지만, 발생학에 실험적 방법론을 도입하였다. 루는 발생 과정을 실험적으로 조작하여 근접원인을 밝히는 "Entwicklungsmechanik"(발생역학)을 제안했다. 이는 '생리학적 발생학' 또는 '인과적 발생학'으로도 불렸다.[84][85]
루는 개구리 수정란의 한쪽 할구를 손상시켜 반쪽짜리 유생이 발생하는 것을 관찰했다. 이는 바이스만의 '모자이크 발생' 가설을 뒷받침하는 듯했다. 그러나 드리슈는 성게 알을 이용해 할구를 분리하는 실험을 하였고, 작지만 온전한 유생이 발생하는 결과를 얻었다. 토머스 헌트 모건도 개구리 알에서 같은 결과를 보고했다. 루의 실험에서는 손상된 할구의 잔해가 남아 발생에 영향을 미쳤던 것이다.[75]
드리슈의 실험은 배아의 여러 부위가 상호작용하는 '조절발생'의 증거였다. 1924년 한스 슈페만과 힐데 만골트는 영원 배아의 포공 등쪽 입술 조직을 다른 배아에 이식하면 새로운 배아 구조가 생겨남을 발견했다. 이들은 이 부위를 형성체(organizer)라 명명했고, 이는 오늘날 슈페만-만골트 형성체로 불린다. 슈페만은 이 업적으로 1935년 노벨 생리학·의학상을 수상했다.[75]
2. 5. 유전학과의 갈등
20세기 유전학과 분자생물학의 발전에 힘입어 발생생물학은 차츰 유전자형이 표현형으로 드러나는 과정을 연구하는 학문으로 규정되기에 이르렀다.[73] 토머스 헌트 모건을 비롯한 미국 유전학자들은 슈페만의 연구가 유전자의 역할을 간과했다며 비판하였고, 조지프 니덤과 콘래드 워딩턴(Conrad Waddington)은 슈페만-만골트 형성체의 유도 작용을 매개하는 분자를 찾아내고자 했다.[84]유전자가 발생생물학의 주인공 자리를 차지하는 것을 모든 학자가 반긴 것은 아니었다. 대표적으로 슈페만 자신은 “형성체 물질”을 찾으려는 시도에 회의적이었다. 형성체의 작용을 매개하는 분자의 정체를 밝혀내더라도 “어떻게 해서 배아의 앞뒤축이 그토록 정확하게 정해지는가?” 하는 큰 질문에 대답하는 데에는 도움이 되지 못한다고 생각했던 것이다.[83] 또한 워딩턴은 유전자의 중요성을 누구보다 일찍 인식했으면서도, 유전자가 발생 양상을 결정한다는 관점을 일종의 전성설이라고 보고 반발하였다. 그는 후성발생(epigenesis) 과정에 주목하는 '후성적 풍경(epigenetic landscape)'의 비유를 제안했다. 세포를 언덕 꼭대기에 놓인 공에, 세포의 운명이 나아갈 수 있는 가능성들을 여러 갈래로 나뉘는 골짜기에 비유할 때, 발생이란 공이 골짜기를 따라 언덕을 굴러 내려오는 과정으로 이해할 수 있다는 것이다. 이때 유전자의 역할은 마치 언덕 표면을 아래로 잡아당기는 닻과 같아서, 닻의 위치와 끌어당기는 힘에 따라 언덕이 다양하게 굴곡지는 것으로 상정하였다.[72]
형성체 작용의 생화학적 기제를 밝히려는 니덤과 워딩턴의 시도는 기술의 한계로 말미암아 당대에는 결실을 거두지 못했다. 하지만 이러한 시도의 과정에서 많은 종류의 유기분자가 비슷한 활성을 나타낸다는 사실이 밝혀졌다.[84] 발생에 영향을 미치는 유전자를 처음 밝혀낸 것은 자발적 돌연변이 덕택이었다. 오늘날 유전자 제거 등의 기법으로 유전자 변형 생물을 제작한 후 배아에 이상이 있는지 관찰하는 것은 발생생물학의 핵심 연구 기법으로 자리 잡았다.[73]
3. 발생 과정
여러 생물은 공통적으로 거쳐 가는 발생 단계를 가진다. 이러한 생활사 개념을 밝힌 것은 기술발생학(descriptive embryology)의 성과이다. 현대 발생생물학은 생물종의 생활사 단계마다 유전자 발현과 해부학적 구조 변화를 연구한다.[86]
3. 1. 동물의 발생

동물의 배아발생 과정은 대체로 수정, 난할, 낭배 형성, 기관 형성 단계로 구분할 수 있다.[86]
# '''수정'''(fertilization)은 정자와 난자가 융합하여 수정란(zygote)을 만드는 단계이다.
# '''난할'''(cleavage)은 수정란이 빠르게 분열하는 단계이다. 세포분열이 성장 없이 일어나므로, 처음에 부피가 컸던 수정란은 차츰 더 작은 할구(blastomere)들로 쪼개진다. 난할이 끝나고 할구들이 덩어리를 이룬 상태를 주머니배 또는 포배라고 부른다.
# '''낭배형성'''(gastrulation)은 포배의 세포들이 대규모로 이동하면서 배엽(germ layer)을 만드는 단계이다. 공 모양이었던 배아는 낭배형성 과정을 거치면서 외배엽(ectoderm), 중배엽(mesoderm), 내배엽(endoderm)이라는 세 층으로 이루어진 낭배(gastrula)가 된다.
# '''기관 형성'''(organogenesis)은 배엽이 만들어지고 나서 세포들이 움직이기도 하고 서로 신호를 주고받기도 하면서 조직과 기관을 만드는 단계이다.
위의 과정과 별개로, 배우자의 전구세포인 생식세포(germ cell)가 개체의 나머지 부분을 이루는 체세포(somatic cell)와 분리되는 현상이 많은 동물의 발생 과정에서 가장 먼저 일어난다. 이후 생식세포는 생식샘(gonad)으로 이동하여 배우자로 분화하는데, 이 과정을 배우자형성(gametogenesis)이라고 부른다. 배우자형성은 보통 개체가 성적으로 성숙하기 전까지 완료되지 않는다. 성숙한 개체에서 방출된 배우자가 다른 배우자를 만나 수정함으로써 생활사가 다음 세대로 이어진다.[86]
생활사의 자세한 사항은 동물 종에 따라 다르다. 예를 들어 진수하강에 속하는 포유류는 배아발생 동안 모체로부터 영양을 공급받아야 하므로, 태반과 같은 배아 바깥 구조(extraembryonic structure)를 만드는 영양외배엽이 구분되는 과정이 낭배형성 이전에 먼저 일어난다.
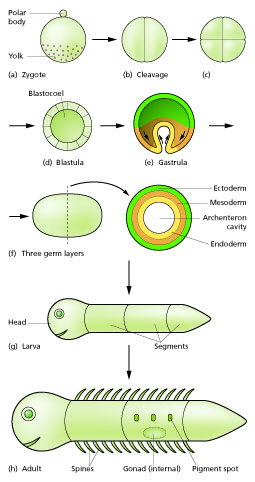
정자와 난자는 수정 과정을 통해 융합되어 수정란(접합자)을 형성한다.[15] 이 수정란은 세포 분열을 거쳐 배반포 또는 배반엽이라고 하는 유사한 세포 덩어리를 형성한다. 이러한 세포 분열은 일반적으로 성장 없이 빠르게 진행되므로 딸세포는 모세포의 절반 크기이며 전체 배아는 거의 동일한 크기를 유지한다. 이를 난할 분열이라고 한다.
생쥐 상배엽 원시 생식 세포 (그림: "인간 배아 발생의 초기 단계")는 광범위한 후생적 재프로그래밍을 거친다.[16] 이 과정에는 게놈 전체의 DNA 탈메틸화, 염색질 재구조화 및 후생적 각인 삭제가 포함되어 전능성으로 이어진다.[16] DNA 탈메틸화는 DNA 염기 절제 복구 경로를 활용하는 과정을 통해 수행된다.[17]
형태 형성 운동은 세포 덩어리를 외배엽, 중배엽, 내배엽의 다층 시트로 구성된 삼층 구조로 변환한다. 이러한 시트를 배엽이라고 한다. 이것이 장 형성 과정이다. 난할과 장 형성 동안 첫 번째 지역 지정 이벤트가 발생한다. 세 개의 배엽 자체의 형성 외에도, 이러한 사건들은 포유류의 태반과 같이 배아의 지지 및 영양 공급에 필요한 배아 외 구조를 생성하며,[18] 전후 축(머리, 몸통 및 꼬리)을 따라 헌신의 차이를 설정한다.[19]
지역 지정은 접합자의 한 부분에 세포질 결정 인자의 존재에 의해 시작된다. 결정 인자를 포함하는 세포는 신호 센터가 되어 유도 인자를 방출한다. 유도 인자는 한 곳에서 생성되어 확산되고 분해되기 때문에 농도 구배를 형성하며, 신호원 세포 근처에서는 높고 멀리 떨어져 있을수록 낮다.[20][21] 결정 인자를 포함하지 않는 배아의 나머지 세포는 특정 발달 조절 유전자를 상향 조절함으로써 다양한 농도에 반응할 수 있다. 그 결과 신호 센터에서 점점 더 멀리 떨어진 일련의 구역이 설정된다. 각 구역에서는 서로 다른 조합의 발달 조절 유전자가 상향 조절된다.[22] 이러한 유전자들은 각 영역에서 새로운 조합의 유전자 활성을 상향 조절하는 전사 인자를 암호화한다. 이러한 전사 인자들은 다른 기능과 함께 활성화된 세포에 특정 부착 및 운동성 특성을 부여하는 유전자 발현을 조절한다. 이러한 서로 다른 형태 발생 특성으로 인해 각 배엽의 세포는 이동하여 외배엽이 바깥쪽에 위치하고, 중배엽이 중간에 위치하며, 내배엽이 안쪽에 위치하는 시트를 형성한다.[23][24]

형태 형성 운동은 배아의 모양과 구조를 변경할 뿐만 아니라, 세포 시트를 새로운 공간적 관계로 가져옴으로써 그들 사이의 새로운 신호 전달 및 반응 단계를 가능하게 한다. 또한, 장 형성, 상피화 및 비틀림과 같은 배아 발생의 첫 번째 형태 형성 운동은 기계적 변환 과정을 통해 내중배엽 특성화와 관련된 경로를 직접 활성화한다.[25][26] 이 특성은 최초의 동물 유기체(최초의 다세포 동물)에서 해양 환경의 유체 역학적 흐름에 의해 기계적으로 자극된 내중배엽 특성화로부터 진화적으로 상속된 것으로 추정된다.[27] 좌선성 키랄성에 의한 신체 축을 따라 비틀림은 모든 척삭동물(척추동물 포함)에서 발견되며 축 비틀림 이론에 의해 설명된다.[28]
배아의 성장은 대부분 자율적이다.[29] 각 세포 영역의 성장 속도는 활성화된 유전자 조합에 의해 조절된다. 자유 생활 배아는 외부 음식 공급이 없기 때문에 질량으로 성장하지 않는다. 그러나 태반 또는 배아 외 난황 공급에 의해 영양을 공급받는 배아는 매우 빠르게 성장할 수 있으며, 이러한 유기체의 부분 간 상대적 성장 속도의 변화는 최종적인 전체 해부학을 생성하는 데 도움이 된다.
전체 과정은 시간 내에 조정되어야 하며, 이것이 어떻게 조절되는지는 아직 밝혀지지 않았다. 배아의 모든 부분과 통신하여 사건의 과정을 제어할 수 있는 마스터 시계가 있을 수 있으며, 또는 타이밍은 단순히 사건의 국소적 인과 관계에 따라 달라질 수 있다.[30]
많은 동물은 출생 직후에는 성적으로 미성숙하며, 유생(larva)에서 성체가 되려면 변태(metamorphosis) 과정을 거쳐야 한다.[86] 올챙이가 자라서 개구리로 탈바꿈하거나, 애벌레가 번데기가 되었다가 나비로 탈바꿈하는 것이 대표적인 예이다. 변태에 의해 개체의 모습은 극적으로 변화한다. 예컨대 올챙이가 개구리가 되면서 꼬리와 아가미는 퇴화하는 반면 사지와 허파는 발달하며, 식물을 뜯어먹기에 알맞은 치아는 사라지는 반면 파리를 잡아먹는 데에 필요한 혀 근육은 발달한다. 이처럼 변태는 신체의 거의 모든 부위를 포괄하는 과정이다.[87]
변태 과정 동안 발달 과정이 매우 뚜렷하게 나타난다. 이는 다양한 종류의 동물에서 발생한다. 잘 알려진 예로는 올챙이로 부화하여 성체 개구리로 변태하는 개구리, 애벌레로 부화하여 번데기 단계를 거쳐 성체 형태로 변태하는 특정 곤충 등이 있다.
위에 나열된 모든 발달 과정은 변태 동안 발생한다. 특히 잘 연구된 예로는 개구리 ''Xenopus''의 올챙이에서 꼬리 소실 및 기타 변화,[31][32] 그리고 파리 ''Drosophila melanogaster''의 성체 신체 부위를 생성하는 상상원반의 생물학이 있다.[33][34]
동물의 배아 발달에 관련된 주요 과정은 다음과 같다. 조직 패턴 형성(지역 특성화 및 패턴화된 세포 분화), 조직 성장, 그리고 조직 형태 형성.
- 지역 특성화는 처음에는 유사한 세포 덩어리나 시트에서 공간적 패턴을 만드는 과정을 말한다. 이것은 일반적으로 수정란의 일부에 위치한 세포질 결정 인자의 작용과 배아의 신호 중심에서 방출되는 유도 신호를 포함한다. 지역 특성화의 초기 단계는 기능적으로 분화된 세포를 생성하지 않지만, 유기체의 특정 영역 또는 부분으로 발달하도록 헌신하는 세포 집단을 생성한다. 이들은 특정 조합의 전사 인자의 발현에 의해 정의된다.
- 세포 분화는 신경, 근육, 분비 상피 등과 같은 기능적 세포 유형의 형성과 관련이 있다. 분화된 세포는 세포 기능과 관련된 많은 양의 특정 단백질을 함유한다.
- 형태 형성은 3차원 형태의 형성과 관련이 있다. 그것은 주로 세포 시트와 개별 세포의 조율된 움직임을 포함한다. 형태 형성은 초기 배아의 세 가지 배엽(외배엽, 중배엽, 내배엽)을 만들고 기관 발달 동안 복잡한 구조를 구축하는 데 중요하다.
- 조직 성장은 조직 크기의 전반적인 증가와 형태 형성에 기여하는 부분의 차등적 성장(얼로메트리)을 모두 포함한다. 성장은 주로 세포 증식을 통해 발생하지만, 세포 크기의 변화 또는 세포 외 물질의 축적을 통해서도 발생한다.
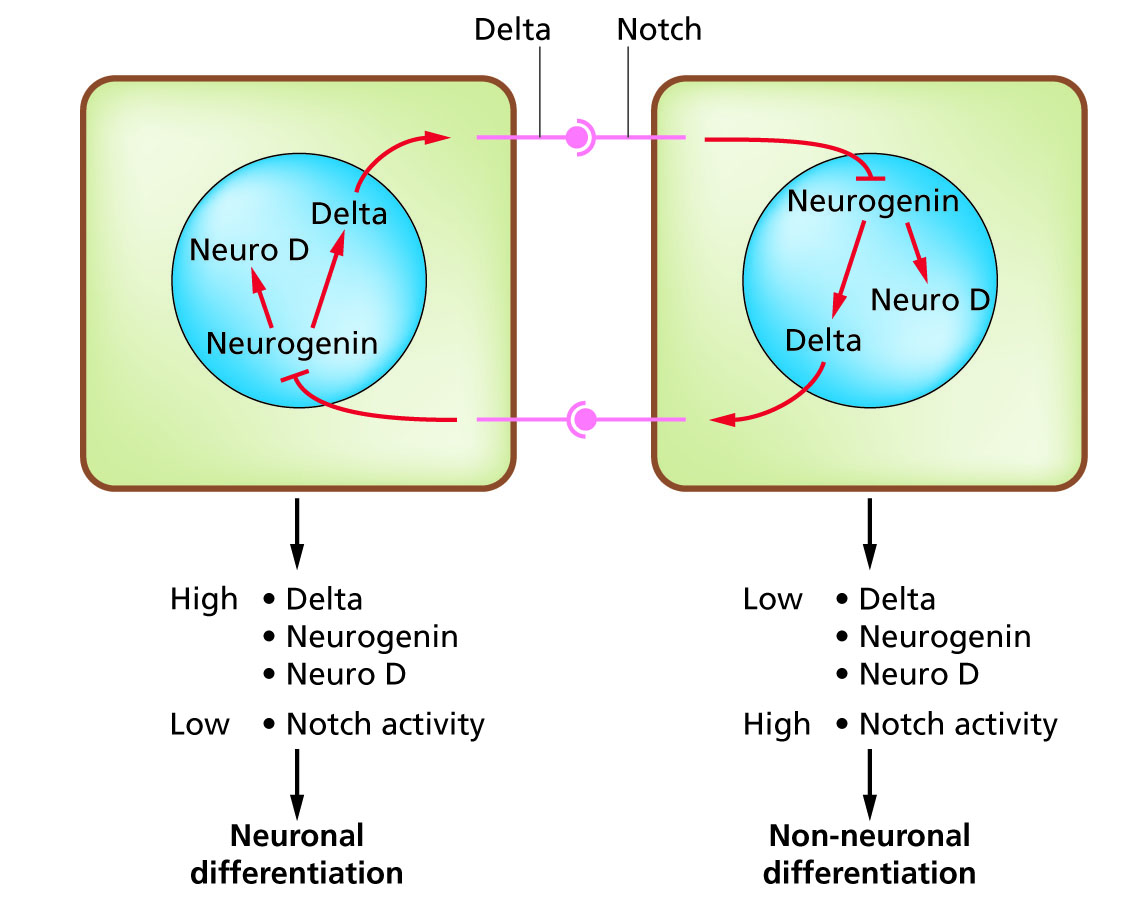
세포 분화는 발생 과정에서 서로 다른 기능적 세포 유형이 발생하는 과정이다. 예를 들어, 뉴런, 근육 섬유 및 간세포(간 세포)는 잘 알려진 분화된 세포 유형이다. 분화된 세포는 일반적으로 특정 기능에 필요한 소량의 단백질을 대량으로 생성하며, 이는 광학 현미경으로 인식할 수 있는 특징적인 외관을 부여한다. 이러한 단백질을 암호화하는 유전자는 매우 활성화되어 있다. 일반적으로 이들의 염색질 구조는 매우 열려 있어 전사 효소에 접근할 수 있으며, 특정 전사 인자가 유전자 발현을 활성화하기 위해 DNA의 조절 서열에 결합한다.[4][5] 예를 들어, NeuroD는 뉴런 분화를 위한 주요 전사 인자이고, 마이오제닌은 근육 분화를 위한 전사 인자이며, HNF4는 간세포 분화를 위한 전사 인자이다.
세포 분화는 일반적으로 발생의 최종 단계이며, 눈에 띄게 분화되지 않은 여러 단계의 결정이 선행된다. 단일 유형의 전구 세포 또는 줄기 세포로 형성된 단일 조직은 종종 여러 분화된 세포 유형으로 구성된다. 이들의 형성을 제어하는 것은 노치 신호 전달 경로의 특성에 기반한 측면 억제[6] 과정을 포함한다.[7] 예를 들어, 배아의 신경판에서 이 시스템은 NeuroD가 고도로 발현되는 신경 전구 세포 집단을 생성하기 위해 작동한다.
재생은 잃어버린 부분을 다시 자라게 하는 능력을 나타낸다.[8] 이는 지속적인 성장을 보이는 식물과 히드라(hydroids)나 멍게류(ascidians)와 같은 군체 동물에게서 매우 흔하게 나타난다. 그러나 발생 생물학자들은 자유 생활 동물에서의 부분 재생에 큰 관심을 보여 왔다. 특히 4가지 모델이 많은 연구의 대상이 되었다. 이 중 2가지 모델은 전체 몸체를 재생하는 능력을 가지고 있다. 작은 조각에서 폴립의 모든 부분을 재생할 수 있는 ''히드라''[9]와 머리와 꼬리를 모두 재생할 수 있는 플라나리아 벌레[10]가 그것이다. 이 두 가지 예시는 줄기 세포에 의해 지속적인 세포 교체가 일어나며, 적어도 플라나리아에서는 줄기 세포 중 일부가 다능성을 띄는 것으로 밝혀졌다.[11] 나머지 두 모델은 부속지의 원위부 재생만을 보인다. 이들은 곤충 부속지, 보통 귀뚜라미와 같은 불완전변태 곤충의 다리,[12] 및 도롱뇽목 양서류의 사지이다.[13] 양서류 사지 재생에 관해 상당한 정보가 있으며, 연골, 진피 및 힘줄 사이의 상당한 상호 전환이 있는 결합 조직을 제외하고 각 세포 유형이 자체적으로 재생된다는 것을 알고 있다. 구조의 패턴 측면에서, 이는 배아에서 활성화된 신호의 재활성화에 의해 제어된다.
재생이 "원시적" 특성인지 "적응적" 특성인지에 대한 오래된 질문에 대한 논쟁이 여전히 존재한다.[14] 만약 전자의 경우라면, 지식이 향상됨에 따라 인간의 재생 능력을 향상시킬 수 있을 것으로 예상할 수 있다. 후자의 경우라면, 각 재생 사례는 종에 특정한 상황에서 자연 선택에 의해 발생한 것으로 추정되므로 일반적인 규칙은 기대할 수 없을 것이다.
발생 과정에 대한 연구는 현미경 관찰이 이루어지면서부터 발전했다. 발생 초기의 관찰에는 세포 수준의 관찰이 필수적이기 때문이다. 특히 무척추동물 각 군의 발생에 관한 지식 축적으로부터, 동물의 발생에는 기본적인 유형이 있으며, 많은 동물의 발생에는 공통된 특징이 있다는 것을 알게 되었다. 이 분야, 또는 그 흐름을 비교 발생학이라고 하며, 19세기에 성립하여 비교 해부학과 함께 비교 형태학이라고 불렸다. 그것들을 종합하여, 거기에 진화적인 의미를 발견한 것이 에른스트 헤켈이다. 그는 각 군의 동물의 발생이, 그 동물이 거쳐온 진화의 과정을 간략화한 것이라고 생각했다. 이 사실은 '개체 발생은 계통 발생을 반복한다'는 표현으로 알려져 있으며, 이를 '반복설'이라고 한다.
| 모델 동물 | 특징 |
|---|---|
| 개구리: 제노푸스[43] (X. laevis 및 X. tropicalis).[44][45] | 배아 공급이 용이함. 특히 미세 수술에 적합함. |
| 제브라피쉬: 다니오 레리오.[46] | 배아 공급이 용이함. 유전학이 잘 발달됨. |
| 닭: 갈루스 갈루스.[47] | 초기 단계가 포유류와 유사하지만, 미세 수술이 더 쉬움. 저렴한 비용. |
| 생쥐: 무스 무스쿨루스.[48] | 유전학이 잘 발달된 포유류[43] |
| 초파리: Drosophila melanogaster.[49] | 배아 공급이 용이함. 유전학적으로 잘 발달됨. |
| 선충: Caenorhabditis elegans.[50] | 배아 공급이 용이함. 유전학적으로 잘 발달됨. 저렴함. |
3. 2. 식물의 발생
식물에서 발생은 식물이 자라면서 구조가 발생하고 성숙해지는 과정이다. 식물 해부학, 식물 생리학, 식물 형태학에서 연구된다. 식물 발생은 태아기에, 식물 생식에서, 그리고 뿌리, 싹, 꽃에서 뻗어나온 것에서 일어나며, 동물 발생과 비슷한 과정을 수반한다.[35]식물은 기관의 끝에 위치한 분열조직이나 성숙한 조직 사이에서 평생 동안 새로운 조직과 구조를 만들어낸다. 따라서 살아있는 식물은 항상 배아 조직을 가지고 있다. 반면, 동물 배아는 매우 초기에 평생 동안 가지게 될 모든 신체 부위를 생성한다. 동물이 태어날 때 모든 신체 부위를 가지며, 그 시점부터는 더 커지고 성숙해질 뿐이다.[35]
관다발 식물은 정자 세포가 난자를 수정시켜 형성된 단일 세포 접합자에서 시작된다. 배 발생 과정을 통해 식물 배를 형성하기 위해 분열한다. 세포는 한쪽 끝이 첫 번째 뿌리가 되고, 다른 쪽 끝이 줄기 끝을 형성하도록 정렬된다. 종자식물에서 배는 하나 이상의 "씨앗 잎"(자엽)을 발달시킨다. 배 발생이 끝나면 어린 식물은 생명을 시작하는 데 필요한 모든 부분을 갖추게 된다.[37]
배가 씨앗이나 모체에서 발아하면 기관 형성 과정을 통해 추가 기관(잎, 줄기, 뿌리)을 생성하기 시작한다. 새로운 뿌리는 뿌리 끝에 위치한 뿌리 분열조직에서 자라고, 새로운 줄기와 잎은 줄기 끝에 위치한 줄기 분열조직에서 자란다.[37] 가지는 분열조직에 의해 남겨진 세포 덩어리가 아직 세포 분화를 거쳐 특수 조직을 형성하지 않은 상태에서 새로운 뿌리나 줄기 끝으로 자라기 시작할 때 발생한다. 뿌리나 줄기 끝의 분열조직에서 자라는 것은 1차 생장이라고 하며, 그 뿌리나 줄기가 길어지는 결과를 낳는다. 2차 생장은 형성층의 세포 분열로 뿌리나 줄기가 넓어지는 결과를 낳는다.[38]
식물 성장과 발생은 특정 식물 호르몬과 식물 성장 조절제(PGR)에 의해 매개된다.[40] 내인성 호르몬 수치는 식물 나이, 내한성, 휴면 및 기타 대사 조건, 광주기, 가뭄, 온도 및 기타 외부 환경 조건, 외부에서 적용되고 근권 기원의 PGR 등 외인성 PGR 공급원에 의해 영향을 받는다.[40]
식물의 발달은 동물과 유사한 과정을 포함한다. 그러나 식물 세포는 대부분 움직이지 않으므로 형태 형성은 세포 이동 없이 차등적 성장을 통해 달성된다. 또한 유도 신호와 관련된 유전자도 동물의 발달을 제어하는 것과 다르다.
세포 분열에 의한 생장 외에도 식물은 '''세포 신장'''을 통해 생장할 수 있다.[39] 이는 개별 세포 또는 세포 그룹이 길어질 때 발생한다. 모든 식물 세포가 같은 길이로 자라는 것은 아니다. 줄기 한쪽의 세포가 다른 쪽의 세포보다 더 길고 빠르게 자라면, 줄기는 더 느리게 자라는 세포 쪽으로 구부러진다. 이러한 방향성 생장은 빛(광굴성), 중력(굴지성), 물(수분굴성), 물리적 접촉(촉성굴성)과 같은 특정 자극에 대한 식물의 반응을 통해 발생할 수 있다.
식물은 형태와 구조에서 자연적인 변이를 나타낸다. 하나의 개체 내에서, 형태와 구조가 다른 유사한 부분과 다를 수 있는 부분이 반복된다.
전사 인자 및 전사 조절 네트워크는 식물 형태 발생과 그 진화에 중요한 역할을 한다. 식물 육상화 과정에서 많은 새로운 전사 인자 계열이 등장했으며 다세포 발달, 생식 및 기관 발달 네트워크에 우선적으로 연결되어 육상 식물의 더 복잡한 형태 발생에 기여했다.[41]
대부분의 육상 식물은 다세포 조류라는 공통 조상을 공유한다. 식물 형태 진화의 한 예는 차로조류에서 볼 수 있다. 연구에 따르면 차로조류는 육상 식물과 상동성을 갖는 특징을 가지고 있다. 식물 형태 진화에 대한 두 가지 주요 이론은 상동 이론과 대립 이론이다. 일반적으로 받아들여지는 이론은 대립 이론이다. 대립 이론은 감수 분열 전에 일어나는 여러 번의 유사 분열이 포자체의 발달을 유발한다고 말한다. 그런 다음 포자체는 독립적인 유기체로 발달한다.[42]
- 쇠비름(''Arabidopsis thaliana'')[43]
4. 주요 개념
발생에 관여하는 주요 과정은 세포 분화, 패턴 형성, 형태형성, 성장, 그리고 진화 발생 생물학에서 탐구되는 시기의 종합적인 조절이다.
- 세포 분화(cell differentiation)는 신경 세포, 근세포, 각종 샘을 이루는 상피세포 등 특정한 기능을 수행하는 세포들이 만들어지는 과정을 가리킨다. 분화된 세포들은 세포 기능과 관련된 많은 양의 특정 단백질을 포함한다. 이때 처음부터 기능적으로 분화된 세포들이 만들어지는 것이 아니라, 장차 개체의 특정 부분으로 발달하기로 예정된 세포 집단이 정의되는 과정이 먼저 일어난다. 이를 예정화(specification)라고 한다. 이것들은 전사 인자들의 특정한 조합의 표현으로 정의된다.
- 패턴 형성(pattern formation)은 처음에 서로 비슷했던 세포들끼리 점차 달라져서 공간적인 패턴이 만들어지는 과정을 가리킨다. 여기에는 수정란의 여러 부분에 포함된 세포질 결정 요인(cytoplasmic determinant)의 활동과 배아의 신호 중추에서 방출되는 유도 신호가 관여한다.
- 형태형성(morphogenesis)은 삼차원 모양이 형성되는 과정을 가리킨다. 세포 집단이나 개별 세포가 조직적으로 움직임으로써 일어난다. 형태 발생은 초기 태아의 세 개의 배엽을 만들고 기관 발생 동안의 복잡한 구조를 만드는 데 중요하다.
- 성장은 개체 크기가 전체적으로 증가하는 현상과 개체의 각 부분이 서로 다른 비율로 자라나는 현상을 아울러 가리킨다. 성장은 주로 세포 분열을 통해 일어나지만 세포 크기의 변화나 세포외기질의 축적으로도 일어난다.
- 각 발생 단계의 시점이 어떻게 조절되는지, 그리고 다양한 과정이 서로 어떻게 통합되는지는 발생생물학에서 가장 적게 이해된 부분이다. 동물 배아에 시계와 같은 기제가 있는지는 아직 불명확하다.
동물의 발생은 세포를 단위로 생각하면, 난할부터 배 발생 동안 개별 세포가 어떻게 분화해 가는 과정으로 볼 수 있다. 앞서 언급한 유도 현상 등은 그 과정이 구체적으로 드러난 것이라고 할 수 있지만, 당시에는 더 이상 탐구할 방법이 없었다. 세포 분화의 비교적 단순한 예로, 이른바 세포성 점균류인 키이로타마호코리카비가 모델 생물로 선택되어 세계적으로 집중적인 연구가 이루어진 시기도 있었지만, 이는 일반화할 수 있는 성과를 거두지 못했다. 그 후, 동물 중 가장 분화가 단순해 보이는 종으로 예쁜꼬마선충(Caenorhabditis elegans)이 선택되었다.
5. 발생과 진화
공간적 예정화는 수정란의 한 부분에 존재하는 세포질 결정 요인으로부터 시작된다. 결정 요인을 포함하는 세포는 신호 중추가 되어 유도 인자를 방출한다. 결정 요인을 포함하지 않는 태아의 나머지 세포들은 특정 발생 조절 유전자를 양성 피드백함으로써 다양한 농도에 반응한다. 이는 일련의 구역이 형성되고 신호 중추로부터 거리가 멀어지는 순서로 배열되는 결과를 낳는다. 각 구역에서는 서로 다른 조합의 발생 조절 유전자들이 양성 피드백된다. 이 유전자들은 각 지역에서 새로운 조합의 유전자 활동을 양성 피드백하는 전사 인자를 암호화한다. 이러한 전사 인자들은 세포 내에서 구체적인 부착 및 운동성을 부여하는 유전자의 발현을 조절하는 등의 기능을 한다. 각 배엽의 세포들은 이러한 다양한 형태 형성 성질로 인해 층을 형성하기 위해 이동하며, 외배엽은 바깥, 중배엽은 중간, 내배엽은 안쪽에 위치하게 된다. 형태 형성 운동은 태아의 모양과 구조를 변화시킬 뿐만 아니라, 배엽을 새로운 공간적 관계로 만들어 서로 간의 새로운 신호와 반응을 가능하게 한다.
발생, 즉 개체발생과 진화, 즉 계통발생의 관계는 오랫동안 많은 학자들을 매료시킨 주제였다. '진화(evolution)'라는 용어는 찰스 다윈 이전부터 발생까지 포괄하는 의미로 사용되었다. 찰스 다윈이 여러 생물 종의 공통 기원을 조사할 때 발생 초기 형태의 유사성을 단서로 삼은 이후, 발생 과정의 차이와 유사성을 계통발생과 연관 지어 설명하려는 노력이 계속되었다. 에른스트 헤켈의 발생반복설이 대표적인 예이다. 그러나 집단유전학을 중심으로 한 현대 종합설의 성립 과정에서 발생생물학적 관점은 배제되었다.[89] 오늘날 발생을 진화생물학적 관점에서, 그리고 진화를 발생생물학적 관점에서 설명하려는 시도는 진화발생생물학 분야에서 활발히 이루어지고 있다.[61][62]
6. 관련 분야
이 분야는 과거 '발생학(embryology)'이라고 불렸지만, 현재는 더 넓은 의미를 가진 발생생물학이라는 명칭으로 불리고 있다. 발생학에서는 성게 등의 배(embryo)의 발생을 관찰하고 기술하는 것을 주로 했다. "이것은 기술적인 한계로 인해 연구 대상이 크고 투명한 알로 한정되었기 때문이다."[1] 또한, 다종 생물 간의 비교를 주로 하는 경우에는 비교 발생학이라고 불린다. 이 분야는 19세기에 비교 해부학과 함께 진화론을 뒷받침하는 근거가 되었다. 그 후, 이식 등의 조작을 하는 실험 발생학이라고 불리는 분야가 발달해 왔다.
최근에는 분자 생물학, 유전학, 세포 생물학의 수법·지견을 도입하면서 발전해 왔으며, 연구 대상은 다양한 생물 종·발생 과정에 미치고 있다. 다양한 생물의 발생 생물학적 지견이 축적되고, 그것들을 비교함으로써 진화를 탐구하려는 진화 발생 생물학(영어: Evolutionary developmental biology, 통칭: evo-devo〈에보데보〉라고도 함)도 활발해지고 있다.
현재의 발생 생물학 연구에서는 주로 모델 생물을 이용하여 연구가 이루어진다. 동물 전반의 모델로는 초파리와 선충이, 척추동물 레벨로는 닭, 아프리카발톱개구리, 제브라피쉬, 송사리 등이, 포유류의 모델로는 마우스가 종종 사용된다. 식물에서는 애기장대가 가장 유명하다.
발생 생물학의 지견은 의료나 농업 분야에서 '발생 공학'으로 응용된다.
참조
[1]
서적
Form and Transformation: Generative and Relational Principles in Biology
Cambridge University Press
1996-11-13
[2]
뉴스
Generative Biology: Designing Biologic Medicines with Greater Speed and Success
https://www.amgen.co[...]
2024-04-05
[3]
웹사이트
Generative Biology: Learning to Program Cellular Machines
https://www.youtube.[...]
NIH
2024-04-05
[4]
논문
The role of chromatin during transcription
2007-02
[5]
논문
Distinct and predictive chromatin signatures of transcriptional promoters and enhancers in the human genome
2007-03
[6]
논문
Pattern formation by local self-activation and lateral inhibition
http://www.me.ucsb.e[...]
[7]
논문
Cis-interactions between Notch and Delta generate mutually exclusive signalling states
2010-05
[8]
서적
Principles of Regenerative Biology.
Academic Press
2007
[9]
논문
Why polyps regenerate and we don't: towards a cellular and molecular framework for Hydra regeneration
2007-03
[10]
논문
Fundamentals of planarian regeneration
[11]
논문
Clonogenic neoblasts are pluripotent adult stem cells that underlie planarian regeneration
2011-05
[12]
논문
Dissecting insect leg regeneration through RNA interference
2008-01
[13]
논문
Limb regeneration
[14]
서적
Essential Developmental Biology
Wiley-Blackwell
2013
[15]
논문
In the beginning: lessons from fertilization in mice and worms
2003-08
[16]
논문
Germline DNA demethylation dynamics and imprint erasure through 5-hydroxymethylcytosine
2013-01
[17]
논문
Genome-wide reprogramming in the mouse germ line entails the base excision repair pathway
2010-07
[18]
서적
Comparative Placentation.
Academic Press
1975
[19]
논문
Anterior-posterior patterning in early development: three strategies
[20]
논문
Morphogenetic gradients - past and present
[21]
논문
Morphogen gradients: from generation to interpretation
[22]
논문
Boundary formation and maintenance in tissue development
2011-01
[23]
논문
Models of morphogenesis: the mechanisms and mechanics of cell rearrangement
2004-08
[24]
논문
Regulated adhesion as a driving force of gastrulation movements
2008-11
[25]
논문
Mechanical induction of twist in the Drosophila foregut/stomodeal primordium
[26]
논문
Evolutionary conservation of early mesoderm specification by mechanotransduction in Bilateria
[27]
논문
Mechano-biochemical marine stimulation of inversion, gastrulation, and endomesoderm specification in multicellular Eukaryota
[28]
논문
An ancestral axial twist explains the contralateral forebain and the optic chiasm in vertebrates
[29]
서적
Cell Growth: Control of Cell Size
Cold Spring Harbor Laboratory Press
[30]
논문
Cell-intrinsic timing in animal development
[31]
논문
Amphibian metamorphosis: an exquisite model for hormonal regulation of postembryonic development in vertebrates
[32]
논문
Amphibian metamorphosis
2007-06
[33]
서적
The Development of Drosophila melanogaster
Cold Spring Harbor Press
1993
[34]
논문
Transdetermination in Drosophila imaginal discs: a model for understanding pluripotency and selector gene maintenance
2003-10
[35]
논문
Apical meristems: the plant's fountain of youth
2003-10
[36]
서적
Plant Growth and Development
https://archive.org/[...]
McGraw-Hill
1964
[37]
논문
Functional domains in plant shoot meristems
2001-02
[38]
논문
Patterned cell determination in a plant tissue: the secondary phloem of trees
2005-05
[39]
논문
Arabidopsis root
2018-08
[40]
서적
Plant growth regulating chemicals
CRC Press
1983
[41]
논문
An Arabidopsis Transcriptional Regulatory Map Reveals Distinct Functional and Evolutionary Features of Novel Transcription Factors
http://mbe.oxfordjou[...]
2015-07
[42]
논문
Morphological evolution in land plants: new designs with old genes
2012-02
[43]
논문
Expression of the cell cycle in sperm of ''Arabidopsis'': implications for understanding patterns of gametogenesis and fertilization in plants and other eukaryotes
The Company of Biologists
[44]
서적
Normal table of ''Xenopus laevis'' (Daudin)
[45]
논문
Xenopus research: metamorphosed by genetics and genomics
2011-12
[46]
논문
Forward and reverse genetic approaches for the analysis of vertebrate development in the zebrafish
2011-07
[47]
논문
The chick embryo: hatching a model for contemporary biomedical research
2009-04
[48]
서적
Manipulating the Mouse Embryo. A Laboratory Manual
Cold Spring Harbor Laboratory Press
2014
[49]
논문
The art and design of genetic screens: Drosophila melanogaster
2002-03
[50]
서적
C.elegans II.
Cold Spring Harbor Laboratory Press
[51]
서적
Current Topics in Developmental Biology Volume 50
Academic Press
[52]
논문
Evolutionary crossroads in developmental biology: the tunicates
2011-06
[53]
논문
Limb regeneration: a new development?
[54]
논문
Modeling human development in 3D culture
2014-12
[55]
서적
Arabidopsis. A Laboratory Manual.
Cold Spring Harbor Laboratory Press
2002
[56]
문서
松村、P316。
[57]
문서
中村、P156 - P175、シャケルフォード、P102 - P104。
[58]
서적
Developmental biology
2016
[59]
서적
Developmental biology
2016
[60]
서적
Principles of development
2019
[61]
간행물
분자생물학 만능 시대 극복하는 통합 생물학 - 진화와 발생으로 아우르는 이보-디보의 첫걸음
동아사이언스
2003-04
[62]
서적
Developmental biology
2016
[63]
서적
A history of embryology
2014
[64]
웹인용
서삼릉 학술이야기
https://royaltombs.c[...]
2023-10-19
[65]
저널
Aristotle, Buddhist scripture and embryology in ancient Mexico: building inclusion by re-thinking what counts as the history of developmental biology
https://journals.bio[...]
2021-02-01
[66]
웹인용
불설포태경
https://kabc.dongguk[...]
2023-10-19
[67]
서적
A history of embryology
2014
[68]
서적
A history of embryology
2014
[69]
서적
A history of embryology
2014
[70]
서적
A history of biology
2021
[71]
서적
Principles of development
Oxford university press
2019
[72]
서적
Theories of Biological Development
https://plato.stanfo[...]
Metaphysics Research Lab, Stanford University
2022
[73]
서적
Principles of development
Oxford university press
2019
[74]
서적
A history of biology
Princeton University Press
2021
[75]
서적
Principles of development
Oxford university press
2019
[76]
서적
Ontogeny and Phylogeny
https://archive.org/[...]
Harvard University Press
1977
[77]
서적
Ontogeny and Phylogeny
https://archive.org/[...]
Harvard University Press
1977
[78]
서적
Ontogeny and Phylogeny
https://archive.org/[...]
Harvard University Press
1977
[79]
저널
Ernst Haeckel in the history of biology
https://linkinghub.e[...]
2019-12
[80]
서적
Ontogeny and Phylogeny
https://archive.org/[...]
Harvard University Press
1977
[81]
서적
From Embryology to Evo-Devo: A History of Developmental Evolution
https://direct.mit.e[...]
The MIT Press
2007-03-02
[82]
서적
Ontogeny and Phylogeny
https://archive.org/[...]
Harvard University Press
1977
[83]
서적
From Embryology to Evo-Devo: A History of Developmental Evolution
https://direct.mit.e[...]
The MIT Press
2007-03-02
[84]
서적
A history of biology
Princeton University Press
2021
[85]
저널
Philosophy of Developmental Biology
https://www.cambridg[...]
2022-03
[86]
서적
Developmental biology
Sinauer associates, Inc., publishers
2016
[87]
서적
Developmental biology
Sinauer associates, Inc., publishers
2016
[88]
웹인용
Whence “Arrival of the Fittest”?
https://ncse.ngo/whe[...]
2023-12-03
[89]
서적
Evolution and Development
https://plato.stanfo[...]
Metaphysics Research Lab, Stanford University
2021
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com