척삭동물
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
척삭동물은 척삭, 등쪽 신경삭, 인두열, 항문 뒤 꼬리, 내주 등의 해부학적 특징을 공유하는 동물군이다. 척삭동물의 진화는 단일 조상에서 유두동물과 두삭동물로 분화되었다는 가설이 널리 받아들여지며, 척추동물, 멍게류, 두삭류가 단계통군을 이룬다는 증거가 있다. 척삭동물은 후구동물에 속하며, 좌우대칭의 몸 계획과 체강을 가지고 있다.
더 읽어볼만한 페이지
- 생물 외부 링크 - 목련
목련은 동아시아 온대·난대 지역에 분포하는 낙엽 활엽수로, 이른 봄 흰색의 향기로운 꽃을 피우고 가을에 붉은 씨앗을 가진 열매를 맺으며, 다양한 이름으로 불리고 한약재 및 여러 용도로 활용된다.
| 척삭동물 - [생물]에 관한 문서 | |
|---|---|
| 지도 | |
| 기본 정보 | |
| 이름 | 척삭동물 |
| 학명 | Chordata |
| 명명자 | Haeckel, 1874 |
| 화석 범위 | 캄브리아기 제3세–현세, }} (에디아카라기 기록 가능성, 555 Ma) |
 | |
| |
| 분류 | |
| 하위 분류군 | †고충동물 †베툴리콜리아? †윤나노존? †카타이미루스 †피카이아 두삭동물 후각동물 미삭동물 총척추동물군 †에몬사스피스 †메타스프리기나 †누우치크티스 †미로쿤밍기아 †코노돈트 척추동물 |
| 계통 분류 | |
| 상문 | 신구동물 |
| 음성 표기 | |
| IPA (영국식) | /ˈkɔːrdeɪt/ |
| 발음 (영국식) | KOR-dayt |
| IPA (미국식) | /kɔːrˈdeɪtə/ |
| 발음 (미국식) | kor-DAY-tə |
2. 정의
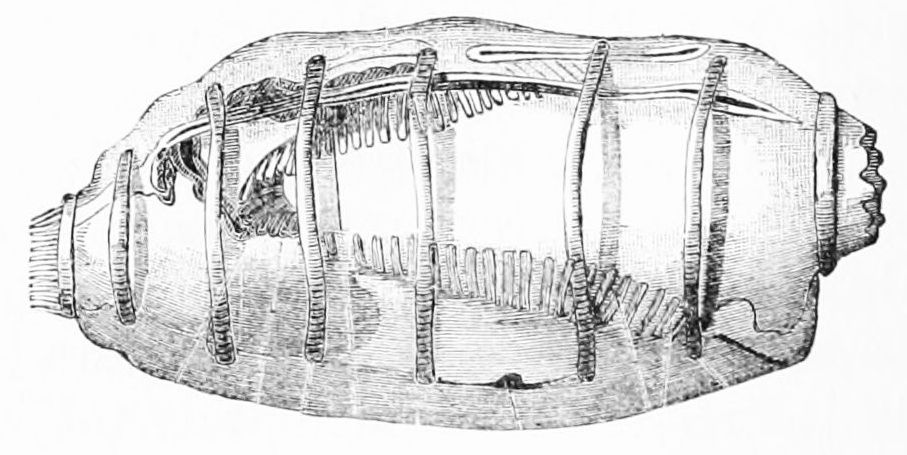
척삭동물은 동물의 문으로, 생애 어느 시점에 척삭, 등쪽 신경삭, 인두열, 항문 뒤 꼬리, 내주와 같은 해부학적 특징을 모두 공유하는 동물군이다.[13]
- 척삭: 신체 중심축을 따라 뻗어 있는, 두 개의 콜라겐 나선으로 감싸인 딱딱하지만 탄력 있는 당단백질 막대이다. 척추동물아문에서는 척삭이 연골 또는 골조직의 척주로 대체되고, 척삭의 잔여물은 추간판이 된다. 완전히 수생인 종의 경우, 척삭은 동물이 꼬리를 좌우로 움직여 효율적으로 헤엄치는 데 도움이 된다.
- 등쪽 신경다발: 신경관이라고도 하며, 신경계의 주요 통신 줄기인 척수가 된다. 척추동물에서 신경관의 뇌측 끝은 배 발생 동안 여러 소포로 커지며, 이것이 뇌를 형성한다.
- 인두열: 인두에 있는 틈으로, 어류에서는 아가미로 변형되지만, 다른 척삭동물에서는 섭취된 물에서 먹이 입자를 추출하는 여과 섭식 시스템의 일부이다. 사지동물에서는 발생의 배아 단계에서만 존재한다.
- 항문 뒤 꼬리: 항문 뒤쪽으로 뻗어 있는 근육질 꼬리이다. 영장류와 같은 일부 척삭동물에서는 배아 단계에서만 존재한다.
- 내주: 인두의 복측 벽에 있는 홈이다. 여과 섭식 종에서는 먹이 입자를 모으는 점액을 생성하여 식도로 먹이를 운반하는 데 도움이 된다.[14] 또한 요오드를 저장하며, 척추동물의 갑상샘의 전구체일 수 있다.[13]
척삭동물은 후구동물이며, 좌우대칭 몸 계획을 기반으로 한다.[15] 또한 체강동물이며, 중배엽에서 유래된 완전한 장막 내막인 중피로 둘러싸인 체액으로 채워진 체강(체강)을 가지고 있다.[16]


3. 진화
척삭동물의 진화에는 몇 가지 가설이 있다. 현재 널리 인정되는 이론은 척삭동물이 모두 하나의 공통 조상에서 진화한 단계통군(monophyletic)이며, 어느 시점에 유두동물과 두삭동물로 나뉘었다고 본다.[8] 척삭동물문, 반삭동물문, 극피동물문, 진와충동물문은 후구동물상문을 이룬다. 후구동물상문은 전체 동물계 종 수의 3분의 2를 차지한다.
초기 척삭동물 화석은 대부분 초기 캄브리아기 청장(Chengjiang) 동물군에서 발견되었으며, 어류로 간주되는 두 종을 포함한다. 이는 이들이 척추동물임을 시사한다. 초기 척삭동물의 화석 기록이 부족하기 때문에, 분자 계통유전학(molecular phylogenetics)만이 그들의 출현 시기를 추정할 수 있다. 그러나 진화적 전환 시기를 추정하는 데 분자 계통유전학을 사용하는 것은 논란의 여지가 있다.[57]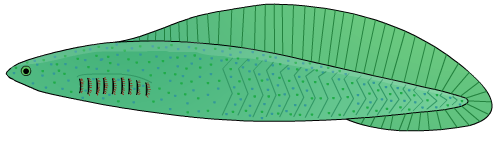
척삭동물 중 가장 단순한 형태를 구분하기 위한 차별적(DNA 염기서열 기반) 비교 연구가 아직도 많이 진행되고 있다.[40] 척추 또는 척삭이 없는 종의 90%에 해당하는 일부 계통에서 이러한 구조가 시간이 지남에 따라 상실되었을 가능성이 있기 때문에 척삭동물의 분류가 복잡해진다.[40]
척삭동물의 진화적 관계를 밝히려는 시도에서 여러 가설이 제시되었다. 현재의 합의는 척삭동물이 단계통군이라는 것이다.[8] 즉, 척삭동물문은 단일 공통 조상의 모든 자손만을 포함하며, 척추동물과 가장 가까운 친척은 멍게류라는 것이다. 사이클로필린 유사 단백질과 미토콘드리아 내막 단백질 분해효소 ATP23에서 모든 척추동물, 멍게류(tunicate) 및 두삭류(cephalochordate)가 독점적으로 공유하는 두 개의 보존된 시그니처 인델(conserved signature indels, CSI)(CSIs)이 최근 확인되어 척삭동물의 단계통성을 강력하게 뒷받침한다.[8]
살아있는 척삭동물 내에서 자세한 분류를 만드는 것은 어려운 것으로 입증되었다. 진화적 "계통도"를 만들려는 시도는 많은 전통적인 강이 측계통군(paraphyletic)임을 보여준다.
{{clade
|label1=후구동물(Deuterostome)
|1={{clade
|1={{clade
|label1=극피동물(Ambulacraria)
|1={{clade
|1=반삭동물(Hemichordate)
|2=극피동물(Echinoderm)

}}
|label2='''척삭동물'''
|2={{clade
|1=두삭류(Cephalochordate)
|label2=후각류(Olfactores)
|2={{clade
|1=멍게류(Tunicate)
|2=척추동물(Vertebrate)
}}
}}
}}
}}
해파리 및 기타 자포동물(cnidarians)보다 복잡한 대부분의 동물은 원구동물(protostome)과 후구동물(deuterostome)의 두 그룹으로 나뉘며, 후자에는 척삭동물이 포함된다.[42] 5억 5500만 년 전의 ''킴베렐라(Kimberella)''가 원구동물에 속했을 가능성이 매우 높다.[43][44] 그렇다면 원구동물과 후구동물 계통은 ''킴베렐라''가 출현하기 전, 적어도 5억 5800만 년 전, 즉 캄브리아기 시작 전에 분리되었어야 한다.[42]
극피동물(echinoderm)(현대의 구성원으로는 불가사리(starfish), 성게(sea urchin), 바다나리(crinoid) 등이 있음)이라는 한 주요 후구동물 그룹의 화석은 캄브리아기 시작, 5억 4200만 년 전부터 매우 흔하다.[47]
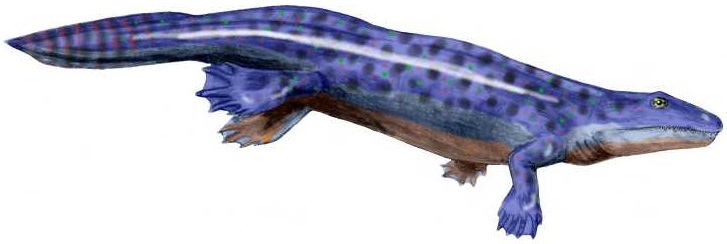
초기 캄브리아기의 청장(Chengjiang) 동물군 화석 ''윤나노조온(Yunnanozoon)''이 반삭동물인지 척삭동물인지에 대한 의견이 다르다.[49][50] 청장 동물군 출신의 또 다른 화석 ''하이쿠엘라 란세올라타(Haikouella lanceolata)''는 척삭동물이며 아마도 두개류일 가능성이 있다고 해석된다. 심장, 동맥, 아가미 필라멘트, 꼬리, 앞쪽에 뇌가 있는 신경 삭, 그리고 눈의 징후를 보이기 때문이다. 비록 입 주위에 짧은 촉수가 있었지만.[50] 청장 동물군 출신의 ''하이쿠이치스(Haikouichthys)''와 ''밀로쿠민기아(Myllokunmingia)''는 어류로 간주된다.[51][52]
중 캄브리아기 버제스 셰일(Burgess Shale)(5억 500만 년 전)에서 훨씬 이전(1911년)에 발견된 ''피카이아(Pikaia)''도 원시 척삭동물로 간주된다.[53] 반면, 초기 척삭동물의 화석은 매우 드물다. 무척추 척삭동물은 뼈나 이빨이 없고, 나머지 캄브리아기에 대해서는 하나만 보고되었기 때문이다.[54]
척삭동물 그룹 간의 진화적 관계와 전체 척삭동물과 그들의 가장 가까운 후구동물 친척 간의 진화적 관계는 1890년부터 논의되어 왔다. 해부학적, 배아 발생(embryology), 고생물학적 데이터를 기반으로 한 연구에서는 서로 다른 "계통도"가 생성되었다. 일부는 척삭동물과 반삭동물을 밀접하게 연결했지만, 그러한 아이디어는 이제 거부된다.[14] 소수의 리보솜 RNA 유전자 데이터와 이러한 분석을 결합하면 일부 이전의 아이디어가 제거되었지만, 멍게류(미삭류)가 "기저 후구동물"이며 극피동물, 반삭동물 및 척삭동물이 진화한 그룹에서 살아남은 구성원일 가능성이 열렸다.[56] 일부 연구원들은 척삭동물 내에서 두개류가 두삭류와 가장 밀접하게 관련되어 있다고 믿지만, 멍게류(미삭류)를 두개류와 가장 가까운 친척으로 간주할 이유도 있다.[14][57]
18세기에 발견되었을 당시, 나막지우오(Amphioxus)는 연체동물의 민달팽이의 일종으로 여겨졌다. 또한, 멍게도 마찬가지로 연체동물로 분류되었다. 1840년대에 들어서면서, 나막지우오는 척삭, 아가미틈, 배쪽신경삭을 가지고 있는 것이 밝혀지면서 척추동물과의 유사성이 지적되기 시작했다. 1866년, 알렉산더 코발레프스키(Alexander Kovalevsky)는 멍게류의 유생이 꼬리에 가지는 축삭상 기관이 척추동물과 나막지우오의 척삭과 상동임을 보였다. 이로 인해 척추동물, 나막지우오를 포함하는 두삭동물, 그리고 멍게를 포함하는 미삭동물이 단계통군임이 널리 받아들여지게 되었다.
4. 해부학적 특징
척삭동물은 생애 어느 시점에 척삭, 등쪽 신경삭, 인두열, 항문 뒤 꼬리, 내주를 포함한 여러 해부학적 특징을 가진다.[13] 척삭은 몸의 중심축을 따라 뻗어 있는 탄력 있는 막대 구조로, 척추동물아문에서는 척추로 대체된다.[13] 등쪽 신경다발은 신경계의 주요 통신 줄기로, 척추동물에서는 뇌와 척수로 분화한다. 인두열은 물을 여과하여 먹이를 섭취하거나 아가미를 통한 호흡에 사용되며, 육상 척추동물에서는 발생 과정에서 다른 기관으로 변형된다.
척삭동물은 좌우대칭 몸 계획을 기반으로 하는 후구동물이며, 체강과 폐쇄 순환계를 갖는다.[15][16] 척삭은 두 개의 콜라겐 나선으로 감싸인 당단백질 막대로, 추간판을 통해 척추뼈가 서로 구부러지고 비틀릴 수 있게 한다. 완전히 수생인 종의 경우, 꼬리를 좌우로 움직여 효율적으로 헤엄칠 수 있도록 돕는다. 신경관의 뇌측 끝은 배 발생 동안 여러 소포로 커져 뇌를 형성한다.[13]
사지동물에서는 인두열이 발생의 배아 단계에서만 존재한다. 항문 뒤 꼬리는 항문 뒤쪽으로 뻗어 있는 근육질 꼬리이며, 영장류와 같은 일부 척삭동물에서는 배아 단계에서만 존재한다. 내주는 인두 복측 벽에 있는 홈으로, 여과 섭식 종에서는 점액을 생성하여 먹이 입자를 모으고 식도로 운반하는 데 도움을 준다.[14] 또한 요오드를 저장하며, 척추동물의 갑상샘 전구체일 수 있다.[13]
5. 하위 분류
두삭동물은 뇌와 특수한 감각 기관이 없는 작은 동물군이다.[25] 몸 안에 연골로 된 척삭을 가지며, 등골 안에 신경관이 있어 중추신경계가 발달하였다. 창고기강 (Leptocardii)이 여기에 속한다.[26][27] 18세기에 발견되었을 당시, (Amphioxus)는 연체동물의 민달팽이의 일종으로 여겨졌다. 1840년대에 들어서면서, 창고기는 척삭, 아가미틈, 배쪽신경삭을 가지고 있는 것이 밝혀지면서 척추동물과의 유사성이 지적되기 시작했다.
'''미삭동물아문'''(Tunicata, 이전의 Urochordata)은 척삭동물의 한 아문으로, 멍게, 해초 등이 속한다. 유생 시기에는 척삭과 신경삭을 가지지만 성체가 되면서 사라지는 특징이 있다.[31] 몸은 피낭으로 덮여 있으며, 플랑크톤을 걸러 먹는 여과 섭식을 한다.
미삭동물은 세 가지 독립적인 성체 형태를 가지며, 이는 세 개의 단계통 분류군으로 나뉜다. 모든 피낭류 유생은 긴 올챙이 모양의 꼬리를 포함하여 표준 척색동물 특징을 가지고 있으며, 뇌, 감광체, 기울기 감지기 등의 기본적인 기관을 가진다.[31]
꼬리돌기류는 평생 올챙이와 같은 모양을 유지하며 능동적으로 헤엄친다.[28] 해삭과 살프는 척삭, 신경삭, 항문 후 꼬리를 잃는 성체 형태로 변태하며, 연체의 여과 섭식 동물로 여러 개의 아가미 구멍을 가지고 점액에 모아진 플랑크톤을 먹고 산다. 해삭은 고착성으로 바다 바닥에 부착하여 평생 한 곳에 머물고,[31] 살프는 중수층에 떠서 플랑크톤을 먹고 살며, 단독 생활과 사슬 모양의 군체를 형성하는 두 세대 주기를 가진다.[29]
미삭동물(Urochordata)이라는 용어는 꼬리에만 척삭이 존재하기 때문에 붙여진 이름이다.(Balfour 1881) oura/οὐράgrc, "꼬리" + 라틴어 chorda ("삭")[30] '''피낭류'''(Tunicata) (Lamarck 1816)라는 용어가 현재 더 일반적으로 사용된다.[31]
18세기에 발견되었을 당시, 멍게는 연체동물로 분류되었다. 1866년, 알렉산더 코발레프스키(Alexander Kovalevsky)는 멍게류 유생이 꼬리에 가지는 축삭상 기관이 척추동물과 나막지우오의 척삭과 상동임을 보였다. 이로 인해 척추동물, 두삭동물, 미삭동물이 단계통군임이 널리 받아들여지게 되었다.
이후 미삭동물과 두삭동물을 합쳐 "원삭동물문"(Protochordata)으로 취급하기도 했으나, 계통 간 연관 관계는 두삭동물과 척추동물 사이가 더 가깝다고 여겨짐에 따라 모두 척삭동물로 묶는 분류가 주류가 되었다. 최근 분자 진화적 분석에서는 미삭동물과 척추동물이 자매군이고, 두삭동물은 그들의 조상적인 계통 관계가 된다는 것이 제시되고 있다.
미삭동물아문은 다음 분류군을 포함한다.
- 해초강 (Ascidiacea)
- 탈리아강 (Thaliacea)
- 유형강 (Appendicularia)
- 소르베라강 (Sorberacea)
척삭이 척주로 대체된 동물군이다. 어류, 양서류, 파충류, 조류, 포유류 등이 척추동물아문에 속한다.[33] 척추동물은 뇌와 척수를 포함하는 발달된 중추신경계와 두개골을 가진다.[34] 칠성장어는 불완전한 뇌함과 척추가 없으므로 척추동물로 간주되지 않지만,[34] 척추동물이 진화했다고 생각되는 그룹 내에 있어 척추동물에서 칠성장어를 계통적으로 제외하는 것은 논란의 여지가 있다.[35][36]
분자 계통 유전학 이전에는 먹장어의 위치가 모호했다. 먹장어는 완전한 뇌함과 흔적 척추를 가지고 있어 척추동물이자 진정한 어류로 간주될 수 있었다.[37] 그러나 DNA를 사용하는 분자 계통 유전학은 먹장어를 척추동물과 함께 분류하는 결과와 칠성장어와 함께 분류하는 결과를 모두 산출했다.[38] 만약 먹장어가 다른 척추동물보다 칠성장어와 더 밀접한 관련이 있다면, 원구류라는 클레이드를 형성한다.[39]
†판피어강 (Placodermi)
연골어강 (Chondrichthyes)
†극어강 (Acanthodii)
- ** 경골어상강 (Osteichthyes)
조기어강 (Actinopterygii)
육기어강 (Sarcopterygii)
- ** 사지상강 (Tetrapoda)
양서강 (Amphibia)
파충강 (Reptilia)
†단궁강 (synapsidalia)
조강 (Aves)
포유강 (Mammalia)
무악상강은 턱이 없는 척추동물을 포함하며, 먹장어강 및 칠성장어강 등이 여기에 속한다. 이들은 척추가 없거나 불완전한 형태로 존재하며, 원시적인 형태의 척추동물로 간주된다.
유악하문은 턱이 있는 척추동물이다. 판피어강, 연골어강, 극어강, 경골어상강 등이 속한다.
경골어상강은 다시 조기어강과 육기어강으로 나뉘며, 육기어강은 사지상강으로 이어진다.
사지상강은 양서강, 파충강, 단궁강, 조강, 포유강을 포함한다.
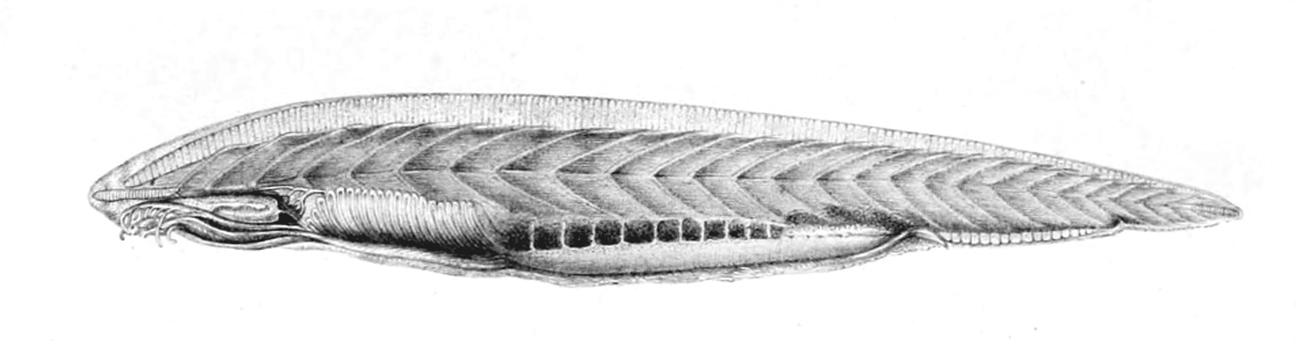
5. 1. 두삭동물아문 (Cephalochordata)
두삭동물은 뇌와 특수한 감각 기관이 없는 작은 동물군이다.[25] 몸 안에 연골로 된 척삭을 가지며, 등골 안에 신경관이 있어 중추신경계가 발달하였다. 창고기강 (Leptocardii)이 여기에 속한다.[26][27] 18세기에 발견되었을 당시, (Amphioxus)는 연체동물의 민달팽이의 일종으로 여겨졌다. 1840년대에 들어서면서, 창고기는 척삭, 아가미틈, 배쪽신경삭을 가지고 있는 것이 밝혀지면서 척추동물과의 유사성이 지적되기 시작했다.5. 2. 미삭동물아문 (Tunicata)
'''미삭동물아문'''(Tunicata, 이전의 Urochordata)은 척삭동물의 한 아문으로, 멍게, 해초 등이 속한다. 유생 시기에는 척삭과 신경삭을 가지지만 성체가 되면서 사라지는 특징이 있다.[31] 몸은 피낭으로 덮여 있으며, 플랑크톤을 걸러 먹는 여과 섭식을 한다.미삭동물은 세 가지 독립적인 성체 형태를 가지며, 이는 세 개의 단계통 분류군으로 나뉜다. 모든 피낭류 유생은 긴 올챙이 모양의 꼬리를 포함하여 표준 척색동물 특징을 가지고 있으며, 뇌, 감광체, 기울기 감지기 등의 기본적인 기관을 가진다.[31]
꼬리돌기류는 평생 올챙이와 같은 모양을 유지하며 능동적으로 헤엄친다.[28] 해삭과 살프는 척삭, 신경삭, 항문 후 꼬리를 잃는 성체 형태로 변태하며, 연체의 여과 섭식 동물로 여러 개의 아가미 구멍을 가지고 점액에 모아진 플랑크톤을 먹고 산다. 해삭은 고착성으로 바다 바닥에 부착하여 평생 한 곳에 머물고,[31] 살프는 중수층에 떠서 플랑크톤을 먹고 살며, 단독 생활과 사슬 모양의 군체를 형성하는 두 세대 주기를 가진다.[29]
미삭동물(Urochordata)이라는 용어는 꼬리에만 척삭이 존재하기 때문에 붙여진 이름이다.(Balfour 1881) οὐρά (oura, "꼬리") + 라틴어 chorda ("삭")[30] '''피낭류'''(Tunicata) (Lamarck 1816)라는 용어가 현재 더 일반적으로 사용된다.[31]
18세기에 발견되었을 당시, 멍게는 연체동물로 분류되었다. 1866년, 알렉산더 코발레프스키(Alexander Kovalevsky)는 멍게류 유생이 꼬리에 가지는 축삭상 기관이 척추동물과 나막지우오의 척삭과 상동임을 보였다. 이로 인해 척추동물, 두삭동물, 미삭동물이 단계통군임이 널리 받아들여지게 되었다.
이후 미삭동물과 두삭동물을 합쳐 "원삭동물문"(Protochordata)으로 취급하기도 했으나, 계통 간 연관 관계는 두삭동물과 척추동물 사이가 더 가깝다고 여겨짐에 따라 모두 척삭동물로 묶는 분류가 주류가 되었다. 최근 분자 진화적 분석에서는 미삭동물과 척추동물이 자매군이고, 두삭동물은 그들의 조상적인 계통 관계가 된다는 것이 제시되고 있다.
미삭동물아문은 다음 분류군을 포함한다.
- 해초강 (Ascidiacea)
- 탈리아강 (Thaliacea)
- 유형강 (Appendicularia)
- 소르베라강 (Sorberacea)
5. 3. 척추동물아문 (Vertebrata)
척삭이 척주로 대체된 동물군이다. 어류, 양서류, 파충류, 조류, 포유류 등이 척추동물아문에 속한다.[33] 척추동물은 뇌와 척수를 포함하는 발달된 중추신경계와 두개골을 가진다.[34] 칠성장어는 불완전한 뇌함과 척추가 없으므로 척추동물로 간주되지 않지만,[34] 척추동물이 진화했다고 생각되는 그룹 내에 있어 척추동물에서 칠성장어를 계통적으로 제외하는 것은 논란의 여지가 있다.[35][36]분자 계통 유전학 이전에는 먹장어의 위치가 모호했다. 먹장어는 완전한 뇌함과 흔적 척추를 가지고 있어 척추동물이자 진정한 어류로 간주될 수 있었다.[37] 그러나 DNA를 사용하는 분자 계통 유전학은 먹장어를 척추동물과 함께 분류하는 결과와 칠성장어와 함께 분류하는 결과를 모두 산출했다.[38] 만약 먹장어가 다른 척추동물보다 칠성장어와 더 밀접한 관련이 있다면, 원구류라는 클레이드를 형성한다.[39]
†판피어강 (Placodermi)
연골어강 (Chondrichthyes)
†극어강 (Acanthodii)
- ** 경골어상강 (Osteichthyes)
조기어강 (Actinopterygii)
육기어강 (Sarcopterygii)
- ** 사지상강 (Tetrapoda)
양서강 (Amphibia)
파충강 (Reptilia)
†단궁강 (synapsidalia)
조강 (Aves)
포유강 (Mammalia)
5. 3. 1. 무악상강 (Agnatha)
무악상강은 턱이 없는 척추동물을 포함하며, 먹장어강 및 칠성장어강 등이 여기에 속한다. 이들은 척추가 없거나 불완전한 형태로 존재하며, 원시적인 형태의 척추동물로 간주된다.5. 3. 2. 유악하문 (Gnathostomata)
유악하문은 턱이 있는 척추동물이다. 판피어강, 연골어강, 극어강, 경골어상강 등이 속한다.경골어상강은 다시 조기어강과 육기어강으로 나뉘며, 육기어강은 네발동물상강으로 이어진다.
네발동물상강은 양서강, 파충강, 단궁강, 조강, 포유강을 포함한다.
6. 계통 분류
척삭동물은 후구동물상문에 속하며, 반삭동물, 극피동물과 함께 신구동물을 형성한다.[74][75][76][77][78] 분자 계통유전학(molecular phylogenetics)적 연구에 따르면, 척삭동물은 단계통군(monophyletic)이며, 척추동물과 가장 가까운 친척은 멍게류(tunicate)이다.[8] 사이클로필린 유사 단백질과 미토콘드리아 내막 단백질 분해효소 ATP23에서 모든 척추동물, 멍게류(tunicate), 두삭류(cephalochordate)가 독점적으로 공유하는 두 개의 보존된 시그니처 인델(conserved signature indels, CSI)(CSIs)이 확인되어 척삭동물의 단계통성을 뒷받침한다.[8]
척삭동물 중 가장 단순한 형태를 구분하기 위한 차별적(DNA 염기서열 기반) 비교 연구가 아직도 많이 진행되고 있다.[40] 초기 척삭동물의 화석 기록이 부족하기 때문에 분자 계통유전학(molecular phylogenetics)만이 그들의 출현 시기를 추정할 수 있는 합리적인 전망을 제공한다.[57] 그러나 진화적 전환의 시기를 추정하는 데 분자 계통유전학을 사용하는 것은 논란의 여지가 있다.[58][59]
해파리 및 기타 자포동물(cnidarians)보다 복잡한 대부분의 동물은 원구동물(protostome)과 후구동물(deuterostome)의 두 그룹으로 나뉘며, 후자에는 척삭동물이 포함된다.[42] 척삭동물과 가장 가까운 친척은 반삭동물과 극피동물로 여겨지며, 이들은 함께 후구동물을 형성한다.[14] 척삭동물과 후구동물은 함께 상위계통 분류군인 신구동물을 형성한다.
다음은 척삭동물의 계통 발생 나무이다.[60][61][62][63][64]
{| class="wikitable"
|-
! '''척삭동물'''
|-
|
|-
!
|-
|
{| class="wikitable"
|-
! 미삭동물
|-
|
| 탈리아류 |
| 유형동물 |
| 피낭류 |
|-
! 척추동물
|-
|
{| class="wikitable"
|-
! 원구류
|-
|
| 점액장어류 |
| 칠성장어류 |
|-
!
|-
|
{| class="wikitable"
|-
| 코노돈트†
|-
| 익갑류†
|-
| 갑피류†
|-
! 악구류
|-
|
{| class="wikitable"
|-
| 판피류†
|-
| 연골어류
|-
! 진구류
|-
|
{| class="wikitable"
|-
| 극어류†
|-
! 경골어류
|-
|
{| class="wikitable"
|-
! 조기어류
|-
|
{| class="wikitable"
|-
| 분기지느러미류
|-
!
|-
|
| 연질어류 |
| 신기어류 |
|}
|-
! 육기어류
|-
|
{| class="wikitable"
|-
!
|-
|
{| class="wikitable"
|-
! 사지동물
|-
|
{| class="wikitable"
|-
| 양서류
|-
! 양막류
|-
|
{| class="wikitable"
|-
! 단궁류
|-
|
{| class="wikitable"
|-
!
|-
|
|}
|-
! 룡궁류
|-
|
{| class="wikitable"
|-
! 이궁류
|-
|
| 린룡형류 |
| 주룡형류 |
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
|--
7. 한국의 척삭동물
참조
[1]
논문
Geochronological constraint on the Cambrian Chengjiang biota, South China
http://nora.nerc.ac.[...]
2018
[2]
논문
A new metazoan from the Vendian of the White Sea, Russia, with possible affinities to the ascidians
2012
[3]
서적
Anthropogenie oder Entwicklungsgeschichte des Menschen
Engelmann
1874
[4]
논문
Comment on "Ultrastructure reveals ancestral vertebrate pharyngeal skeleton in yunnanozoans"
2023
[5]
논문
A new interpretation of Pikaia reveals the origins of the chordate body plan
2024
[6]
논문
A long-headed Cambrian soft-bodied vertebrate from the American Great Basin region
2024
[7]
서적
The fishes of New Zealand
https://www.wikidata[...]
Te Papa Press
2015-01-01
[8]
논문
Molecular signatures that are distinctive characteristics of the vertebrates and chordates and supporting a grouping of vertebrates with the tunicates
2016-01
[9]
논문
Lack of support for Deuterostomia prompts reinterpretation of the first Bilateria
2021-03-19
[10]
웹사이트
Stratigraphic Chart 2022
https://stratigraphy[...]
International Stratigraphic Commission
2022-02
[11]
웹사이트
Chordates
https://eol.org/page[...]
[12]
논문
The authorship of higher chordate taxa
2012-07
[13]
논문
Evolution and Development of the Chordates: Collagen and Pharyngeal Cartilage
2006-03
[14]
논문
Key characters uniting hemichordates and chordates: homologies or homoplasies?
http://article.pubs.[...]
2005-01
[15]
서적
On the Origin of Phyla
University of Chicago Press
2004
[16]
서적
Invertebrates
Sinauer Associates
2003
[17]
서적
Vertebrate Palaeontology
http://palaeo.gly.br[...]
Blackwell Publishing
2004
[18]
BioRef
[19]
서적
Fishes of the World
John Wiley and Sons, Inc
2006
[20]
서적
Vertebrate Paleontology. 3rd ed.
Blackwell Science Ltd
2004
[21]
BioRef
[22]
웹사이트
Reptiles face risk of extinction
https://www.bbc.co.u[...]
BBC
2013-02-15
[23]
웹사이트
New Study Doubles the Estimate of Bird Species in the World
https://www.amnh.org[...]
Amnh.org
[24]
웹사이트
Species Statistics Aug 2019
http://www.reptile-d[...]
[25]
서적
Vertebrate Palaeontology: Biology and Evolution
https://books.google[...]
Blackwell Publishing
2000-04-14
[26]
논문
Evolutionary biology: The amphioxus unleashed
2008-06-19
[27]
웹사이트
Branchiostoma
http://lanwebs.lande[...]
Lander University
[28]
웹사이트
Appendicularia
http://www.environme[...]
Australian Government Department of the Environment, Water, Heritage and the Arts
[29]
웹사이트
Animal fact files: salp
https://www.bbc.co.u[...]
BBC
[30]
서적
Urochordata
Oxford English Dictionary
2009-01
[31]
서적
Vertebrate Palaeontology: Biology and Evolution
https://books.google[...]
Blackwell Publishing
2000-04-14
[32]
서적
Vertebrate Palaeontology: Biology and Evolution
https://books.google[...]
Blackwell Publishing
2000-04-14
[33]
웹사이트
Morphology of the Vertebrates
http://www.ucmp.berk[...]
University of California Museum of Paleontology
[34]
웹사이트
Introduction to the Myxini
http://www.ucmp.berk[...]
University of California Museum of Paleontology
[35]
서적
Biology
Benjamin Cummings
2005
[36]
논문
MicroRNAs revive old views about jawless vertebrate divergence and evolution
[37]
웹사이트
Introduction to the Petromyzontiformes
http://www.ucmp.berk[...]
University of California Museum of Paleontology
2008-10-28
[38]
논문
Monophyly of Lampreys and Hagfishes Supported by Nuclear DNA-Coded Genes
1999-12-01
[39]
논문
Complete Mitochondrial DNA of the Hagfish, Eptatretus burgeri: The Comparative Analysis of Mitochondrial DNA Sequences Strngly Supports the Cyclostome Monophyly
[40]
웹사이트
Why we have a spine when over 90% of animals don't
http://www.bbc.com/e[...]
BBC
2016-08-15
[41]
논문
Chordates
2005-11-22
[42]
논문
The last common bilaterian ancestor
http://dev.biologist[...]
2002-07-01
[43]
서적
The Rise and Fall of the Ediacaran Biota
[44]
논문
Hooking some stem-group "worms": fossil lophotrochozoans in the Burgess Shale
2006-12-01
[45]
서적
The Rise of Animals: Evolution and Diversification of the Kingdom Animalia
Johns Hopkins University Press
[46]
간행물
A Neoproterozoic chordate with possible affinity to the ascidians: New fossil evidence from the Vendian of the White Sea, Russia and its evolutionary and ecological implications
HPF-07 Rise and fall of the Ediacaran (Vendian) biota. International Geological Congress - Oslo 2008.
[47]
논문
Early skeletal fossils
http://www.nrm.se/do[...]
2008-07-18
[48]
논문
''Rhabdotubus'', a Middle Cambrian rhabdopleurid hemichordate
http://www3.intersci[...]
2007-10-01
[49]
논문
Reinterpretation of Yunnanozoon as the earliest known hemichordate
1996-04-01
[50]
논문
An early Cambrian craniate-like chordate
1999-12-01
[51]
논문
Head and backbone of the Early Cambrian vertebrate Haikouichthys
2003-01-01
[52]
논문
Lower Cambrian vertebrates from south China
http://www.bios.niu.[...]
2008-09-23
[53]
논문
A ''Pikaia''-like chordate from the Lower Cambrian of China
1996-11-01
[54]
논문
A Redescription of a Rare Chordate,'' Metaspriggina walcotti'' Simonetta and Insom, from the Burgess Shale (Middle Cambrian), British Columbia, Canada
http://jpaleontol.ge[...]
2009-04-28
[55]
논문
The first tunicate from the Early Cambrian of South China
[56]
논문
Evaluating Hypotheses of Deuterostome Phylogeny and Chordate Evolution with New LSU and SSU Ribosomal DNA Data
2002-05-01
[57]
논문
Molecular Phylogeny and Divergence Times of Deuterostome Animals
2005-11-01
[58]
논문
Molecular clock mirages
1999-01-01
[59]
논문
Do Molecular Clocks Run at All? A Critique of Molecular Systematics
2006-12-01
[60]
논문
The amphioxus genome and the evolution of the chordate karyotype
2008-06-01
[61]
논문
Cyclostome embryology and early evolutionary history of vertebrates
2007-09-01
[62]
논문
A phylogenomic framework and timescale for comparative studies of tunicates
2018-04-01
[63]
서적
ELS
John Wiley & Sons, Ltd
2015-02-16
[64]
논문
Hagfish from the Cretaceous Tethys Sea and a reconciliation of the morphological–molecular conflict in early vertebrate phylogeny
[65]
웹사이트
Introduction to the Hemichordata
http://www.ucmp.berk[...]
University of California Museum of Paleontology
2008-09-22
[66]
서적
History of Life
Blackwell Science
[67]
논문
Geochronological constraint on the Cambrian Chengjiang biota, South China
http://nora.nerc.ac.[...]
2018-01-01
[68]
논문
A new metazoan from the Vendian of the White Sea, Russia, with possible affinities to the ascidians
[69]
일반
[70]
논문
A new vetulicolian from Australia and its bearing on the chordate affinities of an enigmatic Cambrian group
https://doi.org/10.1[...]
2014-10-21
[71]
서적
Biology, Seventh Edition
Benjamin Cummings
2005
[72]
서적
On the Origin of Phyla
University Of Chicago Press
2004
[73]
저널
Evolution and Development of the Chordates: Collagen and Pharyngeal Cartilage
2006
[74]
저널
The Ediacaran emergence of bilaterians: congruence between the genetic and the geological fossil records
2008-04-27
[75]
저널
Estimating the timing of early eukaryotic diversification with multigene molecular clocks
2011-08-16
[76]
웹사이트
Raising the Standard in Fossil Calibration
http://fossilcalibra[...]
2018-03-03
[77]
저널
Support for a clade of Placozoa and Cnidaria in genes with minimal compositional bias
2018
[78]
저널
Revisions to the Classification, Nomenclature, and Diversity of Eukaryotes
2018
[79]
저널
The amphioxus genome and the evolution of the chordate karyotype
2008-06
[80]
저널
Cyclostome embryology and early evolutionary history of vertebrates
2007-09
[81]
저널
A phylogenomic framework and timescale for comparative studies of tunicates
2018-04
[82]
서적
ELS
John Wiley & Sons, Ltd
2015-02-16
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com