2025. 8. 5. 오전 9:02:44
움츠릴 때 움츠릴 줄 아는 ‘무적의 극한 생물’
출처: 한겨레 ( 한국 / 한국어 )
절지동물은 관절 다리를 가진 무척추동물로, 곤충, 거미, 게 등을 포함하는 매우 다양한 동물 문이다. 외골격, 체절 구조, 부속지, 개방혈관계, 그리고 탈피를 통해 성장하는 특징을 갖는다. 절지동물은 협각류, 다지류, 갑각류, 육각류의 4개 아문으로 분류되며, 해양, 담수, 육상 등 다양한 환경에서 서식한다. 절지동물은 식량, 수분, 의약품, 생체 모방 로봇 등 인간에게 다양한 긍정적 측면을 제공하지만, 질병 매개, 농작물 피해 등 부정적인 영향도 미친다.
| 절지동물 - [생물]에 관한 문서 | |
|---|---|
| 기본 정보 | |
 | |
| 학명 | Arthropoda |
| 명명자 | 그라벤호르스트, 1843 |
| 이명 | Condylipoda Latreille, 1802 |
| 화석 기록 범위 | 최초 캄브리아기 (포르투나절)–현세 |
| 다양성 | 약 1,170,000종 |
| 어원 | 그리스어 ἄρθρον(arthron, '관절') + 그리스어 πούς(pous, '발') |
| 동물계 | 동물계 |
| 계통군 | 좌우대칭동물 전구동물 탈피동물상문 범절지동물 |
| 분류 | |
| 아문, 미지정 속, 강 | "디노카리디다" † (측계통군, 때로는 엽족동물로 취급) 케리그마켈라과 † 팜델루리온 † (가능한 엽족동물) 미에리두린 † (가능한 오파비니아류) 파르비벨루스 † (가능한 시베리드 엽족동물) 오파비니아과 † 라디오돈타 † 쿠쿠메리크루스 † (가능한 라디오돈트) 카리오신트립스 † (가능한 라디오돈트) 브라도리다 † 듀테로포다 아르티오포다 † 삼엽충류 † 아그노스티다 (삼엽충류일 가능성) † 네크타스피다 † 아글라스피디다 † 켈로니엘리다 † 부시지헤이아 † 에라투스 † 펑정기아 † 푹시안후이이다 † 이소키시다 † 키이소르토키아 † 킬링크시아 † 메가케이라 † (가능한 측계통군) 협각류 하벨리다 † 거미불가사리류 메가케이라? † 디바스테리움 † 오파콜리다 † 프로소마포다 "신지포수리나" (측계통군) 검미류 데카트리아타 카스마테스피다 † 호우이아 † 위네시에키아 † 광익류 † 거미강 포스파토코피나 (가능한 줄기턱동물) † 대악류 히메노카리나 † 유티카르시노이데아 † 틸라코케팔라? † 다지류 지네류 톡토기류 야스류 노래기류 범갑각류 올리고스트라카 패충류 미스타코카리다류 익티오스트라카 멀티크루스타케아 사이클리다 † 테코스트라카 탄툴로카리다류 요각류 연갑강 알로트리오카리다 세팔로카리다류 새각강 레미피드류 육각류 톡토기목 낫발이목 별벌레목 곤충강 불확실한 위치 아베크아스피스 † 아르트로기리누스 † 베네타르트라 † 캄브로파키코페 † 캄브로포두스 † 캄토필리아 † 추안디아넬라 † 고티카리스 † 마렐로모르파 † 파리오스코르피오 † 리니몬스트룸 † 사로트로세르쿠스 † 스트라보피다류 † 윙게르트셸리쿠스 † 정헤카리스 † |
| 한국어 | |
| 주요 분류군 | |
| 아문 | 협각아문 다지아문 갑각아문 육각아문 |
절지동물은 무척추동물이며, 분절된 몸과 마디가 있는 다리를 가지고 있다.[29] 체제는 환형동물에서 볼 수 있는 체절 구조가 기본을 이루나, 체표에 키틴질의 외골격이 있다는 점이 다르다. 또 좌우 대칭으로 대부분 머리·가슴·배의 세 부분, 또는 머리가슴과 배의 두 부분으로 나뉘어 있다. 부속지는 각 체절마다 한 쌍씩 존재하나 몸의 부분에 따라 모양과 기능이 다르다. 모든 종에서 공통적으로 나타나는 체제는 몸 앞쪽 끝에 입, 뒤쪽 끝에 항문, 배쪽에 부속지, 내부에서는 등쪽에 심장, 중축에 소화관 등이 위치한다는 점이다. 부속지의 대부분은 2교차형이 기본이다.
절지동물은 호흡기, 운동용 부속지, 입틀, 감각기의 발달 정도에 따라 생활 방식이 매우 다양하다. 바다에서는 갑각류(새우, 게 등)가 주류를 이루며, 육지에서는 거미류나 곤충류가, 하늘에서는 곤충류만이 살고 있다. 생식 기간 외에는 단독 생활을 하는 종류가 많지만, 먹이를 찾기 위해 일시적 또는 영속적으로 무리를 짓기도 한다. 특히 개미나 벌은 완벽한 사회생활을 하며, 먹이를 찾아 먼 곳까지 이동하거나, 일정한 지역에서 먹이를 구하고 세력권을 형성하기도 한다.
외골격 또는 큐티클은 키틴으로 구성되는데, 이는 N-아세틸글루코사민의 중합체이다.[30] 많은 갑각류, 응애류, 풍뎅이 아과 프레나파티나에 내의 Penetini 및 Archaeoglenini 분지군,[31] 그리고 (털많은 쥐며느리를 제외한) 지네의 큐티클은 생체 광물화되어 탄산칼슘을 포함하고 있다. 근육 부착에 사용되는 내부 구조인 내골격의 석회화는 일부 거미류에서도 발생하며,[32] 초파리 ''동남아시아과실파리''의 번데기 큐티클에는 인산칼슘이 포함되어 있다.[33]
모든 절지동물의 배아는 체절로 구성되며, 일련의 반복되는 모듈로 이루어져 있다. 현존하는 절지동물의 최종 공통 조상은 아마도 각각 다리 역할을 하는 한 쌍의 부속지를 가진 미분화된 체절들의 연속으로 구성되었을 것이다. 그러나 알려진 모든 현존 및 화석 절지동물은 체절과 그들의 부속지가 다양한 방식으로 특수화된 체절군으로 체절들을 그룹화하고 있다.[37]
많은 곤충의 몸이 3부분으로, 거미의 몸이 2부분으로 보이는 것은 이러한 그룹화의 결과이다. 진드기에는 체절의 외부 징후가 없다.[37] 절지동물은 또한 이러한 일련의 반복되는 체절 패턴의 일부가 아닌 두 가지 신체 요소를 가지고 있는데, 입과 눈이 기원한 앞쪽의 눈 체절[37][41]과 항문 뒤쪽의 미절이 그것이다.
원래 각 부속지가 있는 체절은 두 쌍의 별개 부속지를 가진 것으로 보인다. 하나는 위쪽에 있는 분절되지 않은 외지이고, 다른 하나는 아래쪽에 있는 분절된 내지이다. 이들은 나중에 기저부분(원족 또는 기저절)에 의해 결합된 한 쌍의 이지형 부속지로 융합되었는데, 위쪽 가지는 아가미 역할을 하고 아래쪽 가지는 이동에 사용되었다.[42][86][40] 대부분의 갑각류와 삼엽충과 같은 일부 멸종 분류군의 부속지는 외지로 알려진 또 다른 분절된 가지를 가지고 있지만, 이러한 구조가 단일 기원을 가지고 있는지 여부는 여전히 논란의 여지가 있다.[43][44][40] 알려진 모든 절지동물의 일부 체절에서 부속지는 변형되었는데, 예를 들어 아가미, 입 부분, 정보 수집을 위한 더듬이, 또는 잡는 데 사용하는 발톱을 형성하기 위해 변형되었다.[45] 절지동물은 "스위스 아미 나이프와 같아서 각각 고유한 특수 도구 세트를 갖추고 있다."[37] 많은 절지동물에서 부속지는 신체의 일부 영역에서 사라졌는데, 특히 배 부속지가 사라지거나 크게 변형되는 것이 일반적이다.[37]
체절의 가장 두드러진 특수화는 머리에서 볼 수 있다. 절지동물의 4개 주요 그룹 – 협각류(바다거미, 투구게, 거미류), 다지류(좀벌레, 무각류, 지네, 노래기), 판갑류(빈갑류, 요각류, 말라코스트라카류, 새각류, 육각류 등) 및 멸종된 삼엽충 – 은 다양한 체절 조합으로 형성된 머리를 가지고 있으며, 부속지는 누락되거나 서로 다른 방식으로 특수화되어 있다.[37][46] 다지류와 육각류 모두 유사한 머리 조합을 가지고 있음에도 불구하고, 육각류는 갑각류 내에 깊이 중첩되어 있지만 다지류는 그렇지 않기 때문에 이러한 특징은 별도로 진화한 것으로 여겨진다. 또한, ''마렐라''와 같은 일부 멸종된 절지동물은 고유한 체절과 특수화된 부속지 조합으로 머리가 형성되었기 때문에 이러한 그룹 중 어느 그룹에도 속하지 않는다.[47]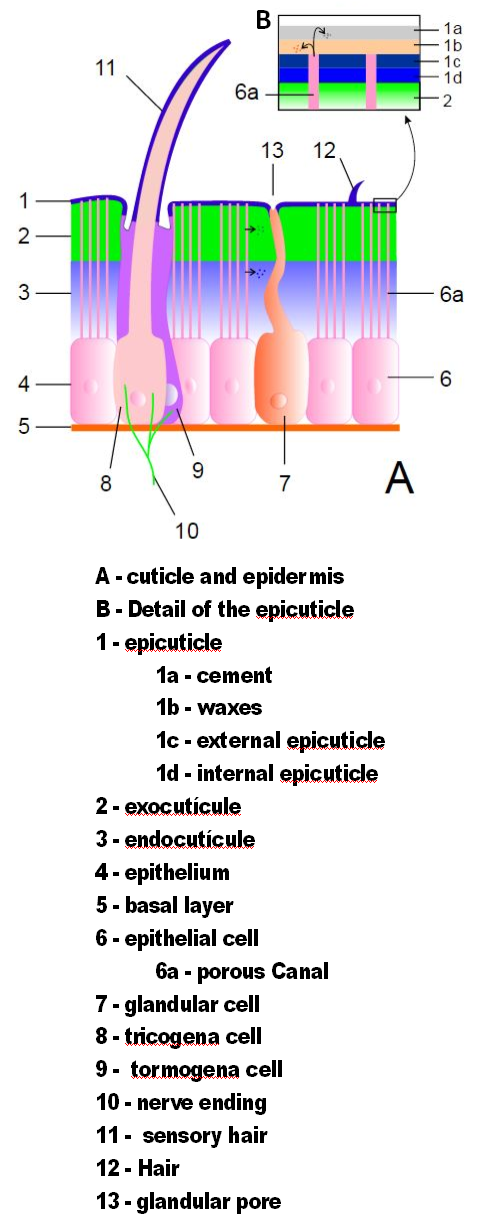
절지동물의 외골격은 큐티클로 이루어져 있으며, 이는 표피에서 분비되는 비세포성 물질이다.[37] 큐티클의 구조는 세부적으로 다양하지만, 일반적으로 세 가지 주요 층으로 구성된다. 표피층은 얇은 외층의 왁스 코팅으로 다른 층의 방수 기능을 제공하고 어느 정도의 보호 기능을 제공한다. 외피층은 키틴과 화학적으로 경화된 단백질로 구성된다. 내피층은 키틴과 경화되지 않은 단백질로 구성된다. 외피층과 내피층을 합쳐 전피층이라고 한다.[50] 각 체절과 다리 부분은 경화된 큐티클로 둘러싸여 있다. 체절과 다리 부분 사이의 관절은 유연한 큐티클로 덮여 있다.[37]
대부분의 수생 갑각류의 외골격은 물에서 추출한 탄산칼슘으로 생체 광물화된다. 일부 육상 갑각류는 광물을 저장하는 수단을 개발했는데, 육지에서는 용해된 탄산칼슘의 안정적인 공급에 의존할 수 없기 때문이다.[51] 생체 광물화는 일반적으로 외피층과 내피층의 바깥 부분에 영향을 미친다.[50] 절지동물과 다른 동물 그룹에서 생체 광물화의 진화에 대한 두 가지 최근 가설은 더 강력한 방어 갑옷을 제공한다는 것[52]과 더욱 단단한 골격을 제공함으로써 동물이 더 크고 강하게 성장할 수 있게 한다는 것이다.[53] 어느 경우든, 비슷한 강도의 완전 유기성 외골격보다 광물-유기 복합 외골격을 만드는 것이 더 저렴하다.[53][54]
큐티클에는 표피의 특수 세포에서 자라는 강모가 있을 수 있다. 강모는 부속지처럼 형태와 기능이 다양하다. 예를 들어, 공기 또는 물의 흐름이나 물체와의 접촉을 감지하는 센서로 자주 사용된다. 수생 절지동물은 수영 부속지의 표면적을 늘리고 물에서 먹이 입자를 여과하는 데 깃털 모양의 강모를 사용한다. 공기 호흡을 하는 수생 곤충은 두꺼운 펠트 모양의 강모 코트를 사용하여 공기를 가두어 물속에서 보낼 수 있는 시간을 연장한다. 무겁고 단단한 강모는 방어용 가시 역할을 한다.[37]
모든 절지동물은 외골격 안쪽에 부착된 근육을 사용하여 다리를 구부리지만, 일부는 여전히 유압 압력을 사용하여 다리를 뻗는다. 이 시스템은 절지동물 이전 조상으로부터 물려받은 것이다.[55] 예를 들어, 모든 거미는 다리를 유압으로 뻗으며 정지 수준의 최대 8배에 달하는 압력을 생성할 수 있다.[56]
절지동물의 몸은 내부적으로도 분절되어 있으며, 신경계, 근육계, 순환계 및 배설계는 반복되는 구성 요소를 가지고 있다.[37] 절지동물은 체강을 가진 동물 계통에서 유래하였는데, 체강은 내장과 체벽 사이에 막으로 둘러싸인 공간으로 내부 기관을 수용한다. 절지동물의 강하고 분절된 다리는 체강의 주요 조상 기능 중 하나인 수정체 골격의 필요성을 없애는데, 이는 근육이 압축되어 동물의 형태를 바꾸고 움직임을 가능하게 한다. 따라서 절지동물의 체강은 생식기관과 배설기관 주변의 작은 영역으로 축소된다. 그 자리는 대부분 혈강에 의해 대체되는데, 혈강은 몸의 대부분 길이에 걸쳐 뻗어 있으며 혈액이 흐르는 공간이다.[58]
절지동물의 형태는 매우 다양하여 종류에 따라 다양한 외형을 가진다. 현생 종의 크기는 1mm 미만의 진드기부터 수십 cm의 바닷가재까지 다양하며, 고생물까지 범위를 넓히면 2m 이상으로 추정되는 거대한 종류도 포함된다.[178][239][179]
절지동물은 '''체절'''(segmentation)을 가진다. 즉, 몸은 체절(somite)이라는 단위의 반복 구조로 이루어져 있으며, 각 체절은 원칙적으로 한 쌍의 부속지를 가진다.[181]
단, 절지동물은 '''이규체절'''(heteronomous metamerism)[182]이 어느 정도 발달하여, 여러 체절이 결합하여 '''합체절'''(tagma)이라는 외관상 또는 기능상의 단위를 구성하는(체절융합, tagmosis, tagmatization) 예가 많이 볼 수 있다.[183][181]
절지동물의 각 체절에는 원칙적으로 1쌍의 '''관절지'''(arthropodized appendage)라는 이 분류군 특유의 부속지가 나와 있다. 이것이 "절지동물"이라는 이름과 그 학명의 유래가 되고 있다.[176] 관절지는 체절과 마찬가지로 외골격으로 덮여 있으며, 관절에 의해 나뉜 '''지절'''(podomere)로 구성된다. 이것은 분류군이나 위치에 따라 다리, 유영각, 집게, 낫, 턱, 촉각, 아가미, 생식지 등 다양한 기능에 따라 다양한 형태로 특화되어 있다.[181]
절지동물의 관절지는, 전체 길이가 가지치지 않은 '''단지형 부속지'''(단지형 부속지, uniramous appendage), 또는 안쪽의 내지(endopod)와 바깥쪽의 외지(exopod)로 가지치기한 '''이지형 부속지'''(이지형 부속지, biramous appendage)로 나타난다. 많은 현생 절지동물(육각류, 다지류, 대부분의 협각류)는 단지형 부속지만 가지고 있지만, 갑각류[185]나 삼엽충, 메가케이라류 등의 고생대의 멸종군에서는 이지형 부속지가 더 일반적이다.[343][186]
절지동물의 외골격은 분절된 관절과 그 사이의 부드러운 '''절간막'''(arthrodial membrane)으로 인해 가동 범위를 확보한다. 분절된 체절은 관절이 신축에서 굴곡까지 다양한 방향으로 움직일 수 있는 경우가 많지만, 절지동물의 부속지의 관절은 대개 하나 또는 한 쌍의 '''관절융기'''(condyle)에 의해 외골격의 받침점이 고정되고, 특히 한 쌍의 경우 축이나 경첩처럼 하나의 평면에서 안정적으로 구부러진다.[187][189][188]
또한, 절지동물의 운동 기구는 상술한 외골격뿐만 아니라 체내 근육에 부착면을 제공하는 '''내골격'''(endoskeleton)도 갖추고 있다. 이것은 주로 외골격 내벽에서 돌출된 것 또는 근육의 뿌리에서 경화된 '''건'''(내돌기, internal tendon, apodeme)이다.[193]
또한, 근육 이외의 기구로 관절을 움직이는 예도 많이 볼 수 있다. 예를 들어 많은 거미류는 다리 중간 관절에 신근을 가지지 않고, 체내 혈림프의 수압(거미 등) 또는 탄력 있는 특수한 외골격(전갈 등)으로 관절을 펼쳐 다리를 뻗는다.[189][200][201][202]
3. 생태
거미류는 그물 같은 포획 장치를 이용하거나 돌아다니며 먹이를 얻는다. 개미와 같은 사회생활을 하는 곤충들은 버섯류를 재배하거나 다른 동물을 기르는 흥미로운 행동을 보인다. 천적이 많은 만큼 방어 수단도 발달했는데, 분비물로 관을 만들어 숨거나, 튼튼한 집이나 고치를 만들기도 한다. 또한 독샘, 독침, 집게발, 발톱 등의 무기를 사용해 적극적으로 공격하며, 의태나 보호색을 통해 몸을 보호하기도 한다.
절지동물문은 가장 큰 동물 문으로, 절지동물 종의 수는 1,170,000종에서 500만~1000만 종으로 추산되며, 알려진 모든 생물 종의 80% 이상을 차지한다.[34][35] 절지동물의 한 아문인 곤충은 다른 어떤 분류학적 강보다 더 많은 종이 기재되어 있다.[36] 종의 총 수를 결정하는 것은 여전히 어렵다. 이는 특정 지역의 개체 수를 전 세계로 확대하기 위해 다른 지역에 투영되는 조사 모델링 가정 때문이다. 1992년 연구에 따르면 코스타리카에만 50만 종의 동물과 식물이 있는데, 그중 36만 5천 종이 절지동물이었다고 한다.[36]
절지동물은 해양, 담수, 육지 및 공기 생태계의 중요한 구성원이며, 건조한 환경에서의 삶에 적응한 두 가지 주요 동물 그룹 중 하나이다. 다른 하나는 파충류, 조류, 포유류를 포함하는 양막류이다.[37] 가장 작은 절지동물과 가장 큰 절지동물 모두 갑각류이다. 가장 작은 절지동물은 탄툴로카리다강에 속하며, 그중 일부는 길이가 100µm 미만이다.[38] 가장 큰 절지동물은 갑각류에 속하며, 일본 대게의 다리 길이는 최대 4m에 달하고,[39] 미국 바닷가재는 20kg이 넘는 무게에 달한다.
3. 1. 생활
절지동물은 호흡기, 운동용 부속지, 입틀, 감각기의 발달 정도에 따라 생활 방식이 매우 다양하다. 바다에서는 갑각류(새우, 게 등)가 주류를 이루며, 육지에서는 거미류와 곤충류가, 하늘에서는 곤충류만이 서식한다. 대부분 생식 기간 외에는 단독 생활을 하지만, 먹이를 찾기 위해 일시적 또는 영속적으로 무리를 짓기도 한다. 특히 개미나 벌은 완벽한 사회생활을 하며, 먹이를 찾아 먼 곳까지 이동하거나, 일정한 지역에서 먹이를 구하고 세력권을 형성하기도 한다.
거미류는 그물 같은 포획 장치를 이용하거나 돌아다니며 먹이를 얻는다. 개미와 같은 사회생활을 하는 곤충들은 버섯류를 재배하거나 다른 동물을 기르는 흥미로운 행동을 보인다. 천적이 많은 만큼 방어 수단도 발달했는데, 분비물로 관을 만들어 숨거나, 튼튼한 집이나 고치를 만들기도 한다. 또한 독샘, 독침, 집게발, 발톱 등의 무기를 사용해 적극적으로 공격하며, 의태나 보호색을 통해 몸을 보호하기도 한다.
절지동물의 외골격은 분절된 관절과 부드러운 '''절간막'''으로 연결되어 넓은 가동 범위를 가진다. 절지동물의 부속지 관절은 보통 하나 또는 한 쌍의 '''관절융기'''에 의해 받침점이 고정되어, 축이나 경첩처럼 한 평면에서 안정적으로 구부러진다.[187][189][188] 부속지 기부는 여러 관절로 나뉘어 다양한 움직임에 대응하며,[187][189][190][191] 외골격 관절의 마찰을 줄이기 위해 윤활 물질을 분비한다.[192]
절지동물은 외골격뿐 아니라 '''내골격'''도 갖추고 있는데, 이는 체내 근육에 부착면을 제공한다. 내골격은 외골격 내벽에서 돌출된 것, 또는 근육 뿌리에서 경화된 '''건'''(내돌기)이다.[193] 예를 들어, 대부분의 절지동물은 부속지 기부의 외재근에 연결되는 '''내복판'''을 체절 내에 가진다.[193][195][196][202] 게처럼 강력한 집게발에는 가동지 내측 관절에 연결된 굴근과 판상의 건이 있다.[197][198][199]
많은 거미류는 다리 중간 관절에 신근이 없어, 체내 혈림프의 수압(거미 등)이나 탄력 있는 외골격(전갈 등)으로 관절을 펴 다리를 뻗는다.[189][200][201][202]
3. 2. 생식과 보육
절지동물은 대부분 암수딴몸이며 체내 수정을 한다. 곤충류나 갑각류 중에는 양성 생식과 단위 생식을 번갈아 하는 종류가 있고, 수컷이 드물거나 없어 단위 생식을 하는 종류도 있다.[69] 개미나 벌처럼 사회 생활을 하는 곤충은 번식 능력을 갖는 암수컷 외에 생식 능력 없이 특정 일만 하는 개체도 있다. 절지동물은 생식 습성이 매우 다양하며, 특히 독립 생활을 하는 종은 배우자를 얻기 위해 경쟁, 유인, 잠복 등 성 본능 행동이 발달되어 있다. 대부분 알을 낳지만(난생), 일부는 알이 어미 뱃속에서 부화하는 난태생이다.[69] 특히 기생성 무리에서는 유생 생식, 유체 성숙, 다배 현상 같은 특수한 생식 방법도 알려져 있다. 절지동물의 알은 대부분 중황란으로, 표면의 세포질만 난할이 진행되는 표할을 한다.
몇몇 절지동물(예: 따개비)은 자웅동체로, 한 개체가 암수 기관을 모두 가지고 있다. 그러나 대부분은 일생 동안 한 성별을 유지한다.[69] 일부 곤충과 갑각류는 특정 조건에서 단위생식으로 번식하기도 하지만, 대부분은 유성생식을 하며, 단위생식 종도 조건이 나빠지면 유성생식으로 돌아간다.[65] 감수분열은 유성생식 및 단위생식 절지동물 모두에게서 나타나는 주요 특징이지만, 그 적응적 이점에 대한 이해는 오랫동안 미해결 문제로 남아있다.[66][67]
수생 절지동물은 투구게처럼 체외 수정을 하거나,[68] 체내 수정을 할 수 있다.[69] 알려진 모든 육상 절지동물은 체내 수정을 한다. 거미류(수확기), 지네, 일부 갑각류는 변형된 부속지(교미각, 음경)로 정자를 암컷에게 전달한다. 그러나 대부분의 수컷 육상 절지동물은 정포(방수 처리된 정자 꾸러미)를 생성하여 암컷이 몸 안으로 가져가게 한다. 일부 종은 암컷이 땅에 놓인 정포를 찾지만, 대부분은 수컷이 복잡한 구애 행동을 성공시킨 후에만 정포를 놓는다.[69]
대부분의 절지동물은 알을 낳지만,[69] 전갈은 난태생으로, 어미 뱃속에서 알이 부화한 후 새끼를 낳고, 어미가 오랫동안 돌본다.[70] 새로 태어난 절지동물은 다양한 형태를 가지며, 곤충만 해도 그 범위가 넓다. 어떤 것은 작은 성체로 부화하고(직접 발생), 좀처럼 부화 후 첫 탈피까지 먹이를 먹지 않고 무력한 경우도 있다. 많은 곤충은 분절된 다리나 단단한 큐티클이 없는 애벌레나 유충으로 부화하여, 유충 조직이 분해되고 성체 몸을 만드는 데 재사용되는 비활동 단계를 거쳐 성체로 변태한다.[71] 잠자리 유충은 절지동물 특유의 큐티클과 관절 다리를 가졌지만, 날개가 없는 수생 동물이며 늘어나는 턱을 가지고 있다.[72] 갑각류는 보통 세 개의 체절과 한 쌍의 부속지만 있는 작은 노플리우스 유충으로 부화한다.[69]
절지동물은 다양한 보육 형태를 보인다. 알을 낳은 뒤 방치하는 원시형, 어미가 알을 품는 경우, 유충이 부화 후 바로 먹이를 얻도록 적당한 장소에 알을 낳는 경우, 유충을 위해 정교한 집을 짓고 먹이를 저장하는 경우, 유충에게 먹이를 공급하는 경우 등이 있다. 유충의 발달 형식은 무변태, 불완전 변태, 완전 변태, 개형 변태, 정형 변태 등으로 나뉜다.
절지동물은 크게 협각류(Chelicerata), 다지류(Myriapoda), 갑각류(Crustacea), 육각류(Hexapoda)의 네 아문으로 분류된다. 화석 기록까지 포함하면, 삼엽충 등을 포함하는 아문과, 이들 아문에 명확히 포함되지 않는 다양한 화석 분류군도 알려져 있다.[171][278]
각 아문의 주요 특징은 앞쪽 체절들이 융합하여 만들어진 합체절(두부 및 전체)에 있는 부속지의 형태이다. 각 아문별 부속지 형태는 다음과 같다.[173][285][219]
 |  |
 | |
 |  |
다음은 기존 분류군(†협각류, †다지류, †갑각류, †육각류)의 크라운 그룹에 포함되지 않는(예: 해당 기존 분류군에 대해 기저적인 화석 분류군, 스템 그룹), 또는 속한 기존 분류군이 불확실한 화석 분류군을 나열한다.
; †공룡갑강 (Dinocaridida)
: 판브델리온, 케리그마켈라, 오파비니아류, 라디오돈타류 등. 절지동물의 스템 그룹, 측계통군[284]. 캄브리아기 - 데본기에 서식.
; †이소키스목 (Isoxyda)
: 이소키스류. 이소키스, 술시카리스 등. 진절지동물[343][286]/대악류[287][336] 중 하나의 스템 그룹. 캄브리아기에 서식.
; (목) (Hymenocarina)[309]
: 카나다스피스, 브랑키오카리스, 오다라이아, 와프티아 등. 진절지동물[337][345][343]/대악류[309][341][338][287][339][340][336]/범갑각류[341][354][342] 중 하나의 스템 그룹. 캄브리아기에 서식.
 |
 |
; †후시엔후이아목 (Fuxianhuiida)[343]
: 후시엔후이아류. 후시엔후이아, 청장고카리스 등. 진절지동물[337][345][284][343]/대악류[309][341][287] 중 하나의 스템 그룹. 캄브리아기에 서식.
; †거대협강 (Megacheira)[313]
: 거대협류[330](대부속지절지동물). 요호이아, 하이쿠카리스, 레안코일리아, 포르티포르셉스 등. 진절지동물[284][343][344][342][338]/협각류[310][287][336][286] 중 하나의 스템 그룹. 캄브리아기(-데본기?)에 서식.
; †브라도리아목 (Bradoriida)
: 브라도리아류. 브라도리아, 쿤밍겔라 등. 진절지동물[344]/대악류[345]/범갑각류[346] 중 하나의 스템 그룹. 캄브리아기에 서식.
; (목) (Phosphatocopina)
: (Hesslandona), (Vestrogothia) 등. 대악류의 스템 그룹[345] 또는 갑각류[347]. 캄브리아기에 서식.
;†마렐로모르프강 (Marrellomorpha)
: 마렐로모르프류[330][332](마렐로모르프류[333]). 마렐라, 바코니시아, 풀카, 미메타스터 등. 진절지동물[309]/대악류[345][344] 중 하나의 스템 그룹. 캄브리아기 - 데본기에 서식.
; †유시칼시노이드강 (Euthycarcinoidea)
: 유시칼시노이드류[330]. 유시칼시누스, 헤테로크라니아, 아판쿠라 등. 대악류[309]/다지류[348] 중 하나의 스템 그룹. 캄브리아기 - 트라이아스기에 서식.
 |
; †낭두강 (Thylacocephala)
: 낭두류(티라코세팔루스류[330]). (Thylacares), 돌로카리스, 앙키토카조카리스[349] 등. 갑각류로서의 위치는 불확실[350]. 오르도비스기[351](캄브리아기?) - 백악기에 서식.
; (목) (Cyclida)
: (Cyclus), (Schramine), (Americlus) 등. 갑각류로서의 위치는 불확실[352]. 석탄기 - 백악기에 서식.
; †하벨리아목 (Habeliida)[353]
: 하벨리아류. 하벨리아, 상크타카리스 등. 협각류/진협각류 중 하나의 스템 그룹[353][354]. 캄브리아기에 서식.
; †몰리소니아목 (Mollisoniida)[355]
: 몰리소니아류. 몰리소니아[356], 텔크시오페 등. 협각류/진협각류 중 하나의 스템 그룹[354][355]. 캄브리아기 - 오르도비스기에 서식.
; (아문[357]) '''(Artiopoda)'''[358][313]
: 삼엽충, 광순류 등. 진절지동물[309]/협각류[345][286]/대악류[308][287][336] 중 하나의 스템 그룹. 캄브리아기 - 페름기에 서식.



| 아문 | 강 | 구성원 | 예시 종 |
|---|---|---|---|
| 협각류 | 요각류 검투게류 거미류 | 바다거미 투구게 위족류, 솔풍뎅이류, 진드기, 전갈, 거미, 진드기 등 |  |
| 다지류 | 좀다지류 소형다지류 노래기류 지네류 | 가짜지네 육각류, 사각류 털노래기류, 알노래기류, 편구노래기류 등 지네류, 석지네류, 지네류 등 |  (노래기류, Spirostreptida) |
| 갑각류 | 개형류 수염새우류 구두충류 흡구류 완족류 요각류 갑각류 두흉류 새각류 레미페디아류 | 씨드 새우 수염새우류 혀벌레 물고기 이 완족류 등 칼라노이드, 시클로포이데아, 미소프리오이데아, 흡구류 등 갯가재, 유령새우, 쥐며느리, 새우, 게, 가재, 크릴 등 말굽새우 요정새우, 올챙이새우, 물벼룩, 조개새우 레미페디아류 |  |
| 육각류 | 곤충 내구강류 | 곤충 좀붙이 등 |  |
절지동물은 '''체절'''(segmentation)을 가진다. 즉, 몸은 체절(somite)이라는 단위의 반복 구조로 이루어져 있으며, 각 체절은 원칙적으로 한 쌍의 부속지를 가진다.[181] 몸은 원칙적으로 위아래가 등판(tergite, tergum)과 배판(sternite, sternum)으로 덮여 있으며, 더욱이 좌우에 측판(pleuron)을 가진 것도 있다. 이러한 외골격도 체절 단위로 되어 있으며, 체절 사이는 관절로 나뉘어 가동성이 있는 경우가 많다. 몸의 선두 체절은 두부체절(ocular somite)이라고 하며, 절지동물의 눈과 입은 여기에 유래한다.[285] 몸의 끝에 미절(telson)이라는 비체절성 꼬리에 해당하는 구조를 갖는 경우도 있다.[181]
단, 절지동물은 '''이규체절'''(heteronomous metamerism)[182]이 어느 정도 발달하여, 여러 체절이 결합하여 '''합체절'''(tagma)이라는 외관상 또는 기능상의 단위를 구성하는(체절융합, tagmosis, tagmatization) 예가 많이 볼 수 있다.[183][181] 예를 들어, 몸을 "머리(head, cephalon)·몸통(trunk)", "두흉부(cephalothorax)·배(abdomen)", "전체(prosoma)·후체(opisthosoma)" 등 2부 또는 "머리·가슴(thorax)·배", "머리·가슴·미부(pygidium)", "전체·중체(mesosoma)·종체(metasoma)" 등 3부로 나누어 부르는 경우가 있으며, 이것은 절지동물의 각 분류군별 특징으로 사용된다.[181] 특히 앞쪽의 합체절(두부융합절 head tagma, 머리·전체 등)은 종종 체절의 융합이 진행되어 외관상으로는 원래의 체절 구조가 보이지 않고 모두 단일 외골격으로 덮여 있다.[181]. 한편, 체절의 융합이나 퇴화가 극단적으로 진행되어 외관상의 체절 구조가 전혀 보이지 않는 예도 있다.[181]
소화관은 대개 순서대로 인두(pharynx)·식도(esophagus)·전장(foregut, 전위 proventriculus)·중장(midgut)·후장(hindgut) 등으로 분화가 진행되고, 발달된 분지(diverticula, 소화선 digestive gland, 중장선 midgut gland)를 중장에 가지는 경우도 있다.[282] 소화관의 전후, 즉 입과 항문 바로 안쪽의 내벽은 큐티클성이며, 성장 시에는 외부의 표피와 마찬가지로 탈피하여 갱신된다.[203]
기본적인 배설기로는 쌍을 이루는 신관(nephridium)이 있다. 이것은 분류군에 따라 특정 부속지 기부에서 개구하여 촉각선(antennal gland, 갑각류의 제2촉각)[204]·하악선(maxillary gland, 갑각류의 하악)[205]·기절선(coxal gland, 협각류의 다리)[206] 등으로 불린다. 그 외의 배설기로는 소화관에서 가지를 친 말피기관(malpighian tubule)이 있으며, 곤충류, 다지류와 거미류에서 볼 수 있다.[207][208][209]
절지동물이라는 단어는 그리스어 ἄρθρον(árthron, 관절)과 πούς(pous, 발/다리)(속격: ποδός, podos)에서 유래하였으며, 이 두 단어를 합쳐 "마디가 있는 다리"를 의미한다.[24] "arthropodes"라는 용어는 1832년 바르텔레미 샤를 조제프 뒤무르티에가 해부학적 설명에서 처음 사용하였다.[1] "Arthropoda"라는 명칭은 독일 동물학자 요한 루트비히 크리스티안 그라벤호르스트(1777~1857)가 1843년에 처음 사용한 것으로 보인다.[25][1] 이 명칭의 기원은 상당한 혼란을 야기했으며, 피에르 앙드레 라트레유나 카를 테오도어 에른스트 폰 지볼트 등 여러 사람에게 잘못 돌려지기도 했다.[1]
절지동물은 인간과 다양한 관계를 맺고 있다. 식량 생산과 소비, 농업, 의학 등 여러 분야에서 긍정적, 부정적 영향을 모두 미친다.
식량 자원: 게, 새우 등 갑각류는 전 세계적으로 인기 있는 수산물이다. 곤충 또한 곤충 식용을 통해 일부 지역에서 중요한 식량 자원으로 활용된다. 꿀벌의 꿀이나 치즈 진드기를 이용한 밀벤케제[359]처럼 절지동물의 생태적 특성을 이용한 식품도 존재한다. 깍지벌레에서 추출한 셸락이나 코치닐 색소는 식품첨가물로 사용된다.[360][361][362]
농업: 절지동물은 농작물의 수분 매개자로서 중요한 역할을 한다. 2005년 기준 절지동물 수분의 경제적 가치는 1530억유로로 추정되며, 이는 당시 식량 생산의 9.5%에 해당한다.[363] 반면, 농업해충으로 분류되는 절지동물은 농작물에 큰 피해를 입힌다. 메뚜기, 노린재, 진딧물, 딱정벌레, 나비목 곤충, 진드기, 노래기 등이 대표적이다.[364]
애완동물: 장수풍뎅이, 사슴벌레, 소라게, 가재, 타란툴라 등 다양한 절지동물이 애완동물로 사육된다.[365][366][367] 곤충젤리와 같은 사육 용품이나 사육 키트도 판매된다.[368]
의학: 말굽게의 혈림프는 세균 내독소 검출 시약 제조에 사용된다.[369][370] 일부 곤충은 민간요법에 사용되거나, 새로운 의약품 개발의 잠재적 자원으로 연구되기도 한다.[371]

| 질병 | 곤충 | 연간 발생 건수 | 연간 사망자 수 |
|---|---|---|---|
| 말라리아 | 말라리아모기 | 2억 6700만 건 | 100만~200만 명 |
| 뎅기열 | 이집트숲모기 | 500만 건 | 5,000명 |
| 황열 | 이집트숲모기 | 4,432건 | 1,177명 |
| 사상충증 | 깔따구 모기 | 2억 5000만 건 | 알 수 없음 |
한편, 인간에게 물거나, 흡혈하거나, 접촉하거나, 기생하거나, 매개 등을 통해 질병을 일으키는 절지동물도 있으며, 이는 위생 해충에 포함된다. 이는 흡혈성으로 감염병을 매개하는 모기나 진드기가 대표적이며, 예를 들어 말라리아모기가 매개하는 말라리아는 2000년부터 2020년 사이에 전 세계적으로 약 1억 7000만 명이 감염되었고, 그중 약 1060만 명이 사망한 것으로 추정된다.[372]
[1]
논문
The correct authorship of Arthropoda—A reappraisal
2023-05-04
[2]
서적
Vergleichende Zoologie
https://www.biodiver[...]
Druck und Verlag von Graß, Barth und Comp
[3]
논문
Burgess Shale fossils shed light on the agnostid problem
2019-01-01
[4]
논문
The evolution of biramous appendages revealed by a carapace-bearing Cambrian arthropod
[5]
논문
A new euarthropod with large frontal appendages from the early Cambrian Chengjiang biota
https://palaeo-elect[...]
2022-01-01
[6]
논문
Arthropod fossil data increase congruence of morphological and molecular phylogenies
2013-09-30
[7]
논문
Description of ''Acheronauta'' gen. nov., a possible mandibulate from the Silurian Waukesha Lagerstätte, Wisconsin, USA
[8]
논문
Redescription of ''Americlus rankini'' (Woodward, 1868) (Pancrustacea: Cyclida: Americlidae) and interpretation of its systematic placement, morphology, and paleoecology
https://eprints.gla.[...]
2020-01-01
[9]
논문
A new Arthropod from the Lower Cambrian Sirius Passet Fossil-Lagerstätten of North Greenland
http://www.geology.c[...]
[10]
논문
New Euthycarcinoids and an Enigmatic Arthropod from the British Coal Measures
2001-02-01
[11]
논문
A large arthropod from the Lower Old Red Sandstone (Early Devonian) of Tredomen Quarry, south Wales: ARTHROPOD FROM THE LOWER ORS
2010-05-01
[12]
논문
Inferring Arthropod Phylogeny: Fossils and their Interaction with Other Data Sources
2017-09-01
[13]
논문
Earliest-known uniramous arthropod
1990-01-01
[14]
논문
The enigmatic arthropod Camptophyllia
http://palaeo-electr[...]
2012-06-11
[15]
논문
Chuandianella ovata: An early Cambrian stem euarthropod with feather-like appendages
https://palaeo-elect[...]
2022-02-22
[16]
논문
Upper Cambrian stem-lineage crustaceans and their bearing upon the monophyly of Crustacea and the position of Agnostus
https://www.research[...]
1990-10-01
[17]
논문
Redescription of the cheloniellid euarthropod ''Triopus draboviensis'' from the Upper Ordovician of Bohemia, with comments on the affinities of ''Parioscorpio venator''
2022-06-13
[18]
논문
An Early Devonian arthropod fauna from the Windyfield cherts, Aberdeenshire, Scotland
2003-05-01
[19]
논문
Sarotrocercus oblitus - Small arthropod with great impact on the understanding of arthropod evolution?
2011-11-01
[20]
논문
The phylogeny of aglaspidid arthropods and the internal relationships within Artiopoda
2013-01-01
[21]
논문
''Devonohexapodus bocksbergensis'' is a synonym of ''Wingertshellicus backesi'' (Euarthropoda) – no evidence for marine hexapods living in the Devonian Hunsrück Sea
[22]
논문
The diverse radiodont fauna from the Marjum Formation of Utah, USA (Cambrian: Drumian)
2021-01-19
[23]
웹사이트
arthropod
2024-05-12
[24]
웹사이트
Arthropoda
http://www.etymonlin[...]
Online Etymology Dictionary
2013-05-23
[25]
서적
Vergleichende Zoologie
https://www.biodiver[...]
Graß, Barth & Comp.
1843
[26]
웹사이트
What is a bug? Insects, arachnids, and myriapods
https://www.tepapa.g[...]
2022-03-10
[27]
서적
The Handy Bug Answer Book
https://archive.org/[...]
Visible Ink
[28]
서적
The Handy Bug Answer Book
https://archive.org/[...]
Visible Ink
[29]
서적
On the Origin of Phyla
https://books.google[...]
University of Chicago Press
[30]
논문
Arthropod cuticle features and arthropod monophyly
1980-08-01
[31]
서적
Australian Beetles Volume 2: Archostemata, Myxophaga, Adephaga, Polyphaga
https://books.google[...]
[32]
논문
Natural calcification of the prosomatic endosternite in the Phalangiidae (Arachnida: Opiliones).
1978-01-01
[33]
논문
Amorphous calcium phosphate in the pupal cuticle of ''Bactrocera dorsalis'' Hendel (Diptera: Tephritidae): A new discovery for reconsidering the mineralization of the insect cuticle
https://linkinghub.e[...]
2019-11-01
[34]
웹사이트
The Arthropod Story
http://evolution.ber[...]
University of California, Berkeley
2008-09-29
[35]
논문
How many species of arthropods? Erwin's estimate revised
http://si-pddr.si.ed[...]
2010-05-06
[36]
서적
The Coevolutionary Process
https://books.google[...]
University of Chicago Press
[37]
문서
Ruppert, Fox & Barnes (2004)
[38]
논문
Tantulocarida (Crustacea) of the Southern Ocean deep sea, and the description of three new species of ''Tantulacus''
2010-10-01
[39]
서적
Scaling: Why is animal size so important?
https://archive.org/[...]
Cambridge University Press
[40]
논문
Exites in Cambrian arthropods and homology of arthropod limb branches
2021-07-30
[41]
논문
Origin and evolution of the panarthropod head – A palaeobiological and developmental perspective
2017-01-01
[42]
웹사이트
Giant sea creature hints at early arthropod evolution
http://news.yale.edu[...]
2015-03-11
[43]
논문
Clonal analysis of Distal-less and engrailed expression patterns during early morphogenesis of uniramous and biramous crustacean limbs
2004-10-01
[44]
논문
The clonal composition of biramous and uniramous arthropod limbs
2008-05-07
[45]
서적
Shaking the Tree: Readings from Nature in the History of Life
University of Chicago Press
[46]
논문
Segmentation and tagmosis in Chelicerata
https://linkinghub.e[...]
[47]
논문
Redescription of ''Marrella splendens'' (Trilobitoidea) from the Burgess Shale, Middle Cambrian, British Columbia
[48]
논문
A palaeontological solution to the arthropod head problem
2002-05-16
[49]
논문
Facts and theories concerning the insect head
[50]
서적
Mechanical Design in Organisms
https://archive.org/[...]
Princeton University Press
[51]
서적
On biomineralization
https://books.google[...]
Oxford University Press
[52]
서적
The Rise and Fall of the Ediacaran Biota
[53]
논문
Not armour, but biomechanics, ecological opportunity and increased fecundity as keys to the origin and expansion of the mineralized benthic metazoan fauna
http://eprints.gla.a[...]
[54]
서적
Neoproterozoic-Cambrian Biological Revolutions
http://www.starregis[...]
[55]
서적
The Invertebrates: A Synthesis
Blackwell Publishing
[56]
논문
The hydraulic mechanism of the spider leg
http://jeb.biologist[...]
[57]
문서
Ruppert, Fox & Barnes (2004)
[58]
문서
Ruppert, Fox & Barnes (2004)
[59]
문서
Ruppert, Fox & Barnes (2004)
[60]
문서
Ruppert, Fox & Barnes (2004)
[61]
문서
Ruppert, Fox & Barnes (2004)
[62]
문서
Ruppert, Fox & Barnes (2004)
[63]
문서
Ruppert, Fox & Barnes (2004)
[64]
논문
Miniaturized imaging systems
http://www.suss-micr[...]
2003-06-01
[65]
백과사전
Encyclopedia of Life Sciences
John Wiley & Sons
[66]
논문
Inventory and phylogenomic distribution of meiotic genes in Nasonia vitripennis and among diverse arthropods
2010-02-01
[67]
서적
Molecular Genetics of Development
1987-01-01
[68]
웹사이트
Facts About Horseshoe Crabs and FAQ
https://myfwc.com/re[...]
[69]
문서
Ruppert, Fox & Barnes (2004)
[70]
서적
European Arachnology 2000
http://www.european-[...]
Aarhus University Press
[71]
논문
The origins of insect metamorphosis
http://www.insecta.u[...]
1999-09-01
[72]
간행물
Diversity and Adaptations of the Aquatic Insects
http://faculty.ncf.e[...]
New College of Florida
[73]
서적
Crustacea and Arthropod Relationships
Taylor & Francis
[74]
웹사이트
Arthropod ancestor had the mouth of a penis worm
http://www.nhm.ac.uk[...]
2016-09-30
[75]
논문
New fossils from the base of the Cambrian in South Australia
http://www.samuseum.[...]
[76]
논문
A ''Parvancorina''-like arthropod from the Cambrian of South China
[77]
논문
''Spriggina'' is a trilobitoid ecdysozoan
http://gsa.confex.co[...]
[78]
논문
Early fossil record of Euarthropoda and the Cambrian Explosion
2018-05-22
[79]
논문
First Early Cambrian Radiolaria
http://biosys-serv.b[...]
[80]
논문
Towering sponges in an Early Cambrian Lagerstätte: Disparity between nonbilaterian and bilaterian epifaunal tierers at the Neoproterozoic-Cambrian transition
https://www.research[...]
2002-04-01
[81]
논문
Bivalved arthropods from the Lower Cambrian Mernmerna Formation of South Australia and their implications for the identification of Cambrian 'small shelly fossils'
https://www.research[...]
[82]
논문
A new early Cambrian bradoriid (Arthropoda) assemblage from the northern Flinders Ranges, South Australia
https://www.research[...]
2014-01-01
[83]
논문
Testing the Darwinian legacy of the Cambrian radiation using trilobite phylogeny and biogeography
http://jpaleontol.ge[...]
1999-03-01
[84]
뉴스
A 520-million-year-old, five-eyed fossil reveals arthropod origin
https://phys.org/new[...]
[85]
논문
An early Cambrian euarthropod with radiodont-like raptorial appendages
https://www.nature.c[...]
2020-12-01
[86]
논문
The evolution of biramous appendages revealed by a carapace-bearing Cambrian arthropod
[87]
서적
Early arthropods, their appendages and relationships
Academic Press
[88]
서적
The Burgess Shale
Yale University Press
[89]
논문
Moulting arthropod caught in the act
2004-05-01
[90]
서적
The Fossil Record of the Pancrustacea
http://dx.doi.org/10[...]
Oxford University Press
2020-09-10
[91]
논문
Collective Behavior in an Early Cambrian Arthropod
https://www.science.[...]
2008-10-10
[92]
논문
Crustaceans and the "Cambrian Explosion"
2001-12-01
[93]
논문
A New Arthropod in Chainlike Associations from the Cnengjiang Lagerstätte (Lower Cambrian), Yunnan, China
2009-01-01
[94]
논문
An epipodite-bearing crown-group crustacean from the Lower Cambrian
2007-10-01
[95]
논문
Early Terrestrial Animals, Evolution, and Uncertainty
[96]
논문
The colonization of land by animals: molecular phylogeny and divergence times among arthropods
[97]
서적
History of Life
Blackwell Science
[98]
논문
Giant claw reveals the largest ever arthropod
[99]
논문
A trigonotarbid arachnid from the Upper Silurian of Shropshire
http://palaeontology[...]
1996-09-01
[100]
논문
A replacement name for the trigonotarbid arachnid ''Eotarbus'' Dunlop
1999-01-01
[101]
논문
Fossil evidence for the origin of spider spinnerets
2008-12-01
[102]
논문
Fossil mesothele spiders
1996-02-01
[103]
논문
The Role of Behavior in the Evolution of Spiders, Silks, and Webs
http://homepage.mac.[...]
2007-12-01
[104]
논문
Stranger than a scorpion: a reassessment of Parioscorpio venator, a problematic arthropod from the Llandoverian Waukesha Lagerstätte
2021-01-01
[105]
논문
Book-lungs in a Lower Carboniferous scorpion
1990-01-01
[106]
논문
Exploring the evolution and terrestrialization of scorpions (Arachnida: Scorpiones) with rocks and clocks
2019-03-01
[107]
논문
The Lower Devonian scorpion ''Waeringoscorpio'' and the respiratory nature of its filamentous structures, with the description of a new species from the Westerwald area, Germany
2008-12-01
[108]
논문
New light shed on the oldest insect
2004-02-01
[109]
논문
The presumed oldest flying insect: more likely a myriapod?
2017-05-30
[110]
서적
The Fossil Record of Insect Diversity and Disparity
http://www.santafe.e[...]
Witwatersrand University Press
2008-10-21
[111]
서적
Entomology
Springer
[112]
웹사이트
Arthropod Fossils and Phylogeny, edited by Gregory D. Edgecomb
http://palaeo-electr[...]
Palaeontologia Electronica
1999-03-15
[null]
논문
Arthropod Fossils and Phylogeny
Columbia University Press
[113]
논문
Head segmentation in Early Cambrian ''Fuxianhuia'': implications for arthropod evolution
1995-06-02
[114]
논문
The morphology of ''Opabinia regalis'' and the reconstruction of the arthropod stem-group
[115]
논문
A Cambrian gilled lobopod from Greenland
[116]
서적
Animal Evolution: Interrelationships of the living phyla
https://books.google[...]
Oxford University Press
[117]
논문
Distinguishing anomalocaridids from arthropods and priapulids
[118]
보도자료
Misunderstood worm-like fossil finds its place in the Tree of Life
http://www.cam.ac.uk[...]
Cambridge University
2017-01-24
[119]
논문
Cambrian origins and affinities of an enigmatic fossil group of arthropods
2004-07-29
[120]
논문
The position of the Arthropoda in the phylogenetic system
1999-01-01
[121]
논문
The evolution of the Ecdysozoa
2008-01-01
[122]
논문
Arthropod Origins: Integrating Paleontological and Molecular Evidence
[123]
논문
Making sense of 'lower' and 'upper' stem-group Euarthropoda, with comments on the strict use of the name Arthropoda von Siebold, 1848
[124]
논문
The origin and early evolution of arthropods
http://paleorxiv.org[...]
2022-04-26
[125]
논문
Challenging received wisdoms: Some contributions of the new microscopy to the new animal phylogeny
2006-04-01
[126]
웹사이트
Fossil Focus: Chelicerata
http://www.palaeonto[...]
2011-01-31
[127]
ITIS
2006-08-15
[128]
논문
Phylogenetic analysis of mitochondrial protein coding genes confirms the reciprocal paraphyly of Hexapoda and Crustacea
2007-08-16
[129]
논문
Pancrustacean phylogeny in the light of new phylogenomic data: support for Remipedia as the possible sister group of Hexapoda
[130]
논문
Phylogeny of Arthropoda inferred from mitochondrial sequences: Strategies for limiting the misleading effects of multiple changes in pattern and rates of substitution
http://www.csulb.edu[...]
2010-04-16
[131]
서적
The position of crustaceans within Arthropoda – Evidence from nine molecular loci and morphology
http://www.amonline.[...]
2006-08-23
[132]
논문
Arthropod relationships revealed by phylogenomic analysis of nuclear protein-coding sequences
[133]
논문
A Phylogenomic Solution to the Origin of Insects by Resolving Crustacean-Hexapod Relationships
http://dx.doi.org/10[...]
2017-06-19
[134]
논문
Pancrustacean Evolution Illuminated by Taxon-Rich GenomicScale Data Sets with an Expanded Remipede Sampling
[135]
논문
The Phylogeny and Evolutionary History of Arthropods
2019-06-01
[136]
논문
Phylogenomics resolves the timing and pattern of insect evolution
[137]
서적
Crustacean Farming: Ranching and Culture
http://www.blackwell[...]
Blackwell
2008-10-03
[138]
웹사이트
Bugfood II: Insects as Food!?!
http://www.uky.edu/A[...]
University of Kentucky Department of Entomology
2008-10-03
[139]
웹사이트
Bugfood III: Insect Snacks from Around the World
http://www.uky.edu/A[...]
University of Kentucky Department of Entomology
2008-10-03
[140]
뉴스
Tuck into a Tarantula
http://www.rhymer.ne[...]
2009-08-24
[141]
웹사이트
Spiderwomen serve up Cambodia's creepy caviar
http://abc.net.au/ne[...]
2009-08-24
[142]
서적
Lonely Planet Cambodia
Lonely Planet Publications
[143]
서적
Fierce Food
https://archive.org/[...]
Plume
2008-10-03
[144]
서적
Butterflies in My Stomach (or: Insects in Human Nutrition)
Woodbridge Press Publishing Company
[145]
웹사이트
Codex Standard 152 of 1985 (on "Wheat Flour")
http://www.codexalim[...]
Food and Agriculture Organization
2010-05-08
[146]
웹사이트
Complete list of Official Standards
http://www.codexalim[...]
Food and Agriculture Organization
2010-05-08
[147]
웹사이트
The Food Defect Action Levels
http://www.cfsan.fda[...]
U. S. Food and Drug Administration
2006-12-16
[148]
서적
Ecological implications of minilivestock: potential of insects, rodents, frogs, and snails
https://books.google[...]
Science Publishers
[149]
논문
Economic valuation of the vulnerability of world agriculture confronted with pollinator decline
https://halshs.archi[...]
2008-08
[149]
뉴스
Economic value of insect pollination worldwide estimated at 153 billion euros
http://www.eurekaler[...]
[150]
웹사이트
Apiservices — International honey market — World honey production, imports & exports
http://www.beekeepin[...]
[151]
웹사이트
Time line of fabrics
http://threadsintyme[...]
Threads In Tyme, LTD
[152]
웹사이트
The bug that changed history
http://www.gcrg.org/[...]
[153]
웹사이트
Canary Islands cochineal producers homepage
http://www.arrakis.e[...]
[154]
서적
Forensic Entomology: the Utility of Arthropods in Legal Investigations
https://archive.org/[...]
CRC Press
[155]
논문
Insects and their chemical weaponry: New potential for drug discovery
2010-12
[156]
논문
Distributed mechanical feedback in arthropods and robots simplifies control of rapid running on challenging terrain
http://polypedal.ber[...]
2007-03
[157]
서적
Adaptive Motion of Animals and Machines
[158]
서적
The Economic Importance of Insects
Springer
[159]
서적
Tick-borne diseases of humans
https://books.google[...]
ASM Press
[160]
웹사이트
Parasitic Mites of Humans
http://www.ca.uky.ed[...]
University of Kentucky College of Agriculture
[161]
웹사이트
House dust mite allergy
http://www.netdoctor[...]
NetDoctor
[162]
서적
Ecological Entomology
John Wiley & Sons
[163]
서적
Insect and Mite Pests in Food: An Illustrated Key
http://www.afpmb.org[...]
United States Department of Agriculture
[164]
논문
Mite Pests of Honey Bees
1982-01
[165]
서적
Introduction to insect pest management
https://books.google[...]
Wiley-IEEE
[166]
서적
Encyclopedia of Life Sciences
John Wiley & Sons, Ltd.
[167]
논문
Which predatory mite can control both a dominant mite pest, ''Tetranychus urticae'', and a latent mite pest, ''Eotetranychus asiaticus'', on strawberry?
2002-03
[168]
문서
절지동물 계통수 그림
[169]
논문
The correct authorship of Arthropoda—A reappraisal
[170]
서적
Lehrbuch der vergleichenden Anatomie der Wirbellosen Thiere. Erster Theil. In Lehrbuch der vergleichenden Anatomie (eds C. T. von Siebold and H. Stannius)
https://www.biodiver[...]
Verlag von Veit & Comp.
1848
[171]
논문
The Phylogeny and Evolutionary History of Arthropods
http://www.sciencedi[...]
2019-06-17
[172]
웹사이트
Arthropoda (arthropods)
[173]
서적
The Arthropoda: A Phylogenetic Framework. In book: Arthropod Biology and Evolution – Molecules, Development, Morphology (pp.17-40)Chapter: The Arthropoda: a phylogenetic framework
https://www.research[...]
Springer Berlin Heidelberg
2013
[174]
서적
절지동물의 다양성과 계통
裳華房
2008-04-11
[175]
문서
육생을 주체로 하는 분류군에 대한 설명
[176]
서적
A Dictionary of Entomology
https://books.google[...]
CABI
2011
[177]
문서
クチクラ, セメント, 蝋, 外角皮, 上皮組織, 基底層, 上皮細胞, 腺細胞, 神経, 剛毛
[178]
논문
Giant claw reveals the largest ever arthropod
https://royalsociety[...]
2008-02-23
[179]
논문
The largest arthropod in Earth history: insights from newly discovered ''Arthropleura'' remains (Serpukhovian Stainmore Formation, Northumberland, England)
https://doi.org/10.1[...]
2021-12-20
[180]
논문
How the Ecdysozoan Changed Its Coat
https://journals.plo[...]
2005-10-11
[181]
서적
Arthropod Biology and Evolution: Molecules, Development, Morphology
https://doi.org/10.1[...]
Springer
2013
[182]
서적
Biology of Arthropoda
https://books.google[...]
Discovery Publishing House
2004
[183]
논문
甲殻類系統発生雑記(動物分類学会シンポジウム講演要旨)
https://doi.org/10.1[...]
1963
[184]
서적
Invertebrate Zoology
Brooks/Cole
2004
[185]
웹사이트
Exopodites , Epipodites and Gills in Crustaceans
https://www.semantic[...]
2009
[186]
논문
Exites in Cambrian arthropods and homology of arthropod limb branches
https://www.nature.c[...]
2021-07-30
[187]
논문
Invertebrate paraxial locomotory appendages: design, deformation and control.
https://www.semantic[...]
1999
[188]
서적
Chapter 5 - Systems and Scaling
https://www.scienced[...]
Academic Press
2017-01-01
[189]
논문
Morphology of locomotor appendages in Arachnida: evolutionary trends and phylogenetic implications
https://doi.org/10.1[...]
1989-09-01
[190]
논문
Gimbals in the insect leg
https://www.scienced[...]
2009-01-01
[191]
논문
Neofunctionalization of a Duplicate dachshund Gene Underlies the Evolution of a Novel Leg Segment in Arachnids
https://doi.org/10.1[...]
2016-01-01
[192]
논문
Lubrication in the joints of insects (Arthropoda: Insecta)
https://onlinelibrar[...]
2022-01
[193]
논문
The endoskeletal structures in arthropods: cytology, morphology and evolution
https://linkinghub.e[...]
2002-02
[194]
웹사이트
endosterniteの意味・使い方
https://eow.alc.co.j[...]
[195]
논문
Morphology of the prosomal endoskeleton of Scorpiones (Arachnida) and a new hypothesis for the evolution of cuticular cephalic endoskeletons in arthropods
https://www.scienced[...]
2007-03-01
[196]
논문
Animal Evolution: The Hard Problem of Cartilage Origins
https://www.scienced[...]
2016-07-25
[197]
논문
Dimorphism and the functional basis of claw strength in six brachyuran crabs
https://onlinelibrar[...]
2001-09
[198]
논문
THE EVOLUTION OF ARMAMENT STRENGTH: EVIDENCE FOR A CONSTRAINT ON THE BITING PERFORMANCE OF CLAWS OF DUROPHAGOUS DECAPODS
http://www.bioone.or[...]
2001
[199]
논문
Crabs grab strongly depending on mechanical advantages of pinching and disarticulation of chela: Functional Morphology Of Decapod Chela
https://onlinelibrar[...]
2016-10
[200]
논문
Mechanics of cuticular elastic energy storage in leg joints lacking extensor muscles in arachnids
https://doi.org/10.1[...]
2003-02-15
[201]
서적
Biology of spiders
https://www.academia[...]
Oxford University Press
2011
[202]
논문
Evolutionary and functional substitution of extrinsic musculature in Solifugae (Arachnida)
https://onlinelibrar[...]
2020-12
[203]
웹사이트
Digestive System
https://genent.cals.[...]
[204]
웹사이트
Crustacea Glossary::Definitions (Antennal gland)
https://research.nhm[...]
[205]
웹사이트
Crustacea Glossary::Definitions (Maxillary gland)
https://research.nhm[...]
[206]
서적
The Colonisation of Land: Origins and Adaptations of Terrestrial Animals
https://books.google[...]
Cambridge University Press
1983-12-15
[207]
논문
Insect-Like Characteristics of the Malpighian Tubules of a Non-Insect: Fluid Secretion in the Centipede ''Uthobius Forficatus'' (Myriapoda: Chilopoda)
https://doi.org/10.1[...]
1991-07-01
[208]
논문
Morphology and ultrastructure of the malpighian tubules of the Chilean common tarantula (Araneae: Theraphosidae)
https://onlinelibrar[...]
2002-01
[209]
논문
Inorganic and organic anion transport by insect renal epithelia
https://www.scienced[...]
2003-12-30
[210]
논문
The Biochemistry of Insect Hemolymph
https://www.annualre[...]
1961-01
[211]
논문
Hematopoiesis and Hematopoietic Organs in Arthropods
https://www.ncbi.nlm[...]
2013-03
[212]
논문
Phylogenetic analysis of the Malacostraca (Crustacea)
http://doi.wiley.com[...]
2001-09
[213]
논문
Arthropod neurons and nervous system
https://www.ncbi.nlm[...]
2016-10-24
[214]
서적
動物学の百科事典
丸善出版
2018-09-28
[215]
논문
The evolution of arthropod heads: reconciling morphological, developmental and palaeontological evidence
https://doi.org/10.1[...]
2006-07-01
[216]
논문
Serial Homology and Segment Identity in the Arthropod Head
https://doi.org/10.1[...]
2022-01-01
[217]
서적
動物学の百科事典
丸善出版
2018-09-28
[218]
논문
Trichobothria
https://doi.org/10.1[...]
Springer
2022-07-29
[219]
논문
Segmentation and tagmosis in Chelicerata
https://www.academia[...]
2017-05
[220]
논문
The Cerci and Abdominal Giant Fibres of the House Cricket, ''Acheta domesticus''. I. Anatomy and Physiology of Normal Adults
https://www.jstor.or[...]
1974
[221]
논문
Primary processing neuropils associated with the malleoli of camel spiders (Arachnida, Solifugae): a re-evaluation of axonal pathways
https://doi.org/10.1[...]
2019-08-02
[222]
논문
The tettigoniid (Orthoptera : Tettigoniidae) ear: Multiple functions and structural diversity
https://www.scienced[...]
1993-04-01
[223]
웹사이트
Acoustic Receivers: From Insect Ear to Next-Generation Sensor {{!}} ARCHIE-WeSt
https://www.archie-w[...]
2018-12-20
[224]
논문
The midline metathoracic ear of the praying mantis, ''Mantis religiosa''
https://www.research[...]
1988-01-01
[225]
논문
The structure and function of auditory chordotonal organs in insects
https://onlinelibrar[...]
2004-04-15
[226]
서적
Biology of spiders
https://www.academia[...]
New York: Oxford University Press
2011
[227]
논문
Homology of Head Sclerites in Burgess Shale Euarthropods
http://www.sciencedi[...]
2015-06-15
[228]
논문
Lateral eye evolution in the arachnids
https://bioone.org/j[...]
2016-07
[229]
논문
A Great-Appendage Arthropod with a Radial Mouth from the Lower Devonian Hunsrück Slate, Germany
http://science.scien[...]
2009-02-06
[230]
논문
Acute vision in the giant Cambrian predator Anomalocaris and the origin of compound eyes
https://www.nature.c[...]
2011-12
[231]
논문
Arthropod eyes: The early Cambrian fossil record and divergent evolution of visual systems
https://www.research[...]
2016-03
[232]
서적
XI. The Hexapoda
https://www.degruyte[...]
Cornell University Press
2019-05-20
[233]
논문
The Function of the Tracheal Gills in Larvae of the Caddis Fly, Macronema zebratum Hagen
https://www.jstor.or[...]
1931
[234]
논문
The bimodal gas exchange strategies of dragonfly nymphs across development
https://www.scienced[...]
2020-01-01
[235]
서적
VIII. The Diplopoda
https://www.degruyte[...]
Cornell University Press
2019-05-20
[236]
논문
The tracheal system of scutigeromorph centipedes and the evolution of respiratory systems of myriapods
https://www.scienced[...]
2021-01-01
[237]
서적
X. The Symphyla
https://www.degruyte[...]
Cornell University Press
2019-05-20
[238]
논문
Respiratory gut peristalsis by sea spiders
https://www.cell.com[...]
2017-07-10
[239]
논문
Miniaturisation in Chelicerata
https://www.scienced[...]
2019-01-01
[240]
논문
On the structure of the respiratory organs of the terrestrial Isopoda
https://eprints.utas[...]
1931
[241]
논문
Evolutionary adaptation of oniscidean isopods to terrestrial life: Structure, physiology and behavior
https://www.research[...]
2011
[242]
논문
The morphology and vasculature of the respiratory organs of terrestrial hermit crabs (''Coenobita'' and ''Birgus''): gills, branchiostegal lungs and abdominal lungs
https://linkinghub.e[...]
2005-01
[243]
서적
Chapter 20 - Cladocera and Other Branchiopoda
https://www.scienced[...]
Academic Press
2022-08-01
[244]
논문
Evolutionary Biology of Centipedes (Myriapoda: Chilopoda)
https://www.annualre[...]
2007-01-01
[245]
논문
The Development of Arthropod Segmentation Across the Embryonic/Post-embryonic Divide – An Evolutionary Perspective
https://www.frontier[...]
2021
[246]
웹사이트
An Animated Guide to This Year's Massive Brood X Cicada Emergence
https://time.com/604[...]
2022-08-05
[247]
논문
Mud and silk in the dark: A new type of millipede moulting chamber and first observations on the maturation moult in the order Callipodida
https://www.scienced[...]
2016-05-01
[248]
서적
Biology of spiders
https://www.academia[...]
Oxford University Press
2011
[249]
논문
The evolution of the Ecdysozoa
https://www.ncbi.nlm[...]
2008-04-27
[250]
논문
Palaeontological and Molecular Evidence Linking Arthropods, Onychophorans, and other Ecdysozoa
https://evolution-ou[...]
2009-06
[251]
논문
Current Understanding of Ecdysozoa and its Internal Phylogenetic Relationships
https://doi.org/10.1[...]
2017-09-01
[252]
서적
Generelle Morphologie der Organismen. Allgemeine Grundzüge der Organischen formen-wissenschaft, mechanisch begründet durch die von Charles Darwin reformirte descendenztheorie
Georg Reimer
1866
[253]
간행물
Evolution of the annelida onychophora and arthropoda
https://repository.s[...]
1938
[254]
서적
Arthropod phylogeny with special reference to insects
https://archive.org/[...]
New York : Wiley
1979
[255]
논문
The oldest ‘onychophoran’Xenusion: a link connecting phyla?
https://onlinelibrar[...]
1989
[256]
논문
Phylogeny, biogeography and reproductive trends in the Onychophora
https://www.research[...]
1995-05-01
[257]
논문
Memoirs: The Structure and Classification of the Arthropoda
https://doi.org/10.1[...]
1904-03-01
[258]
서적
Arthropodes: Généralités, composition de l’embranchement
Masson
1949
[259]
간행물
Los órdenes de los artrópodos
http://bba.bioucm.es[...]
1971
[260]
논문
Evidence for a clade of nematodes, arthropods and other moulting animals
https://doi.org/10.1[...]
1997-05
[261]
논문
The new animal phylogeny: Reliability and implications
https://www.research[...]
2000-05-01
[262]
논문
Expression of ‘segmentation’ genes during larval and juvenile development in the polychaetes Capitella sp. I and H. elegans
https://www.scienced[...]
2006-01-01
[263]
논문
Origin and differentiation of nephridia in the Onychophora provide no support for the Articulata
https://doi.org/10.1[...]
2006-03-01
[264]
논문
Parallel evolution of segmentation by co-option of ancestral gene regulatory networks
https://onlinelibrar[...]
2010
[265]
논문
Further spermatological evidence for including the pentastomida (tongue worms) in the Crustacea
https://www.scienced[...]
1992-02-01
[266]
논문
Phylogenetic position of the Pentastomida and (pan)crustacean relationships
https://royalsociety[...]
2004-03-07
[267]
서적
Animal Evolution, Interrelationships of the Living Phyla
Oxford University Press
1995
[268]
논문
Neuroanatomy of ''Halobiotus crispae'' (Eutardigrada: Hypsibiidae): Tardigrade brain structure supports the clade panarthropoda
http://onlinelibrary[...]
2012-11-01
[269]
논문
Partial mitochondrial gene arrangements support a close relationship between Tardigrada and Arthropoda
https://www.research[...]
2008-01-01
[270]
논문
Ecdysozoan mitogenomics: evidence for a common origin of the legged invertebrates, the Panarthropoda
http://gbe.oxfordjou[...]
2010
[271]
서적
Ultrastruktur und Phylogenie von Seison nebaliae (Grube 1859) und Seison annulatus (Claus 1876) : Hypothesen zu phylogenetischen Verwandtschaftsverhältnissen innerhalb der Bilateria
https://www.worldcat[...]
Cuvillier
1995
[272]
서적
Cycloneuralia
https://www.research[...]
Springer
2015
[273]
논문
Segmentation in Tardigrada and diversification of segmental patterns in Panarthropoda
https://www.scienced[...]
2017-05-01
[274]
논문
A Cambrian gilled lobopod from Greenland
https://doi.org/10.1[...]
1993-08
[275]
논문
The morphology of Opabinia regalis and the reconstruction of the arthropod stem-group
https://onlinelibrar[...]
1996-03
[276]
논문
Brain structure resolves the segmental affinity of anomalocaridid appendages
https://doi.org/10.1[...]
2014-07-16
[277]
논문
Anomalocaridid trunk limb homology revealed by a giant filter-feeder with paired flaps
https://www.nature.c[...]
2015-03-11
[278]
논문
Arthropod Origins: Integrating Paleontological and Molecular Evidence
https://www.annualre[...]
2020-11-02
[279]
논문
Early fossil record of Euarthropoda and the Cambrian Explosion
https://www.research[...]
2018-05-21
[280]
논문
The “evolution” of Anomalocaris and its classification in the arthropod class Dinocarida (nov.) and order Radiodonta (nov.)
https://www.cambridg[...]
1996-03
[281]
논문
Cephalic and limb anatomy of a new Isoxyid from the Burgess Shale and the role of "stem bivalved arthropods" in the disparity of the frontalmost appendage
https://pubmed.ncbi.[...]
2015
[282]
논문
Sophisticated digestive systems in early arthropods
https://www.nature.c[...]
2014-05-02
[283]
논문
New opabiniid diversifies the weirdest wonders of the euarthropod stem group
https://www.research[...]
2022-02-09
[284]
논문
Making sense of ‘lower’ and ‘upper’ stem-group Euarthropoda, with comments on the strict use of the name Arthropoda von Siebold, 1848
http://eprints.esc.c[...]
2016
[285]
논문
Origin and evolution of the panarthropod head – A palaeobiological and developmental perspective
https://www.scienced[...]
2017-05-01
[286]
논문
A three-eyed radiodont with fossilized neuroanatomy informs the origin of the arthropod head and segmentation
https://www.cell.com[...]
2022-07-08
[287]
논문
An early Cambrian euarthropod with radiodont-like raptorial appendages
https://www.nature.c[...]
2020-12
[288]
논문
The Evolution of the Insect Head: the Endless Dispute
https://www.biodiver[...]
1975
[289]
서적
On the relationships and phylogeny of fossil and recent Arachnomorpha: a comparative study on Arachnida, Xiphosura, Eurypterida, Trilobita, and other fossil Arthropoda
https://paleoarchive[...]
Jacob Dybwad
1944
[290]
서적
Arthropod Relationships
https://doi.org/10.1[...]
Springer Netherlands
1998
[291]
논문
Expression of homeobox genes shows chelicerate arthropods retain their deutocerebral segment
http://www.pnas.org/[...]
1998-09-01
[292]
논문
Development of the nervous system in the "head" of Limulus polyphemus (Chelicerata: Xiphosura): morphological evidence for a correspondence between the segments of the chelicerae and of the (first) antennae of Mandibulata
https://doi.org/10.1[...]
2003-02-01
[293]
논문
Immunohistochemical localization of neurotransmitters in the nervous system of larval Limulus polyphemus (Chelicerata, Xiphosura): evidence for a conserved protocerebral architecture in Euarthropoda
https://www.scienced[...]
2005-07-01
[294]
논문
A conserved genetic mechanism specifies deutocerebral appendage identity in insects and arachnids
https://royalsociety[...]
2015-06-07
[295]
논문
A Review of Arthropod Phylogeny: New Data Based on Ribosomal DNA Sequences and Direct Character Optimization
https://doi.org/10.1[...]
2000-06
[296]
논문
Hexapod Origins: Monophyletic or Paraphyletic?
http://science.scien[...]
2003-03-21
[297]
논문
Pancrustacean phylogeny: hexapods are terrestrial crustaceans and maxillopods are not monophyletic
https://www.ncbi.nlm[...]
2005-02-22
[298]
논문
Arthropod relationships revealed by phylogenomic analysis of nuclear protein-coding sequences
https://doi.org/10.1[...]
2010-02
[299]
논문
Phylotranscriptomics to Bring the Understudied into the Fold: Monophyletic Ostracoda, Fossil Placement, and Pancrustacean Phylogeny
https://doi.org/10.1[...]
2012-09-12
[300]
웹사이트
Linking Insects with Crustacea : Comparative Physiology of the Pancrustacea Organized by
https://www.semantic[...]
2015
[301]
논문
Pancrustacean Evolution Illuminated by Taxon-Rich Genomic-Scale Data Sets with an Expanded Remipede Sampling
https://doi.org/10.1[...]
2019-08-01
[302]
서적
16 Phylogeny of the Chelicerates: Morphological and molecular evidence
https://www.degruyte[...]
De Gruyter
2014-02-27
[303]
논문
Biodiversity and phylogeny of Ammotheidae (Arthropoda: Pycnogonida)
https://europeanjour[...]
2017-02-24
[304]
논문
Arthropod phylogeny based on eight molecular loci and morphology
https://www.nature.c[...]
2001-09
[305]
논문
Pycnogonid affinities: A review
2005
[306]
논문
Revised systematics of Palaeozoic ‘horseshoe crabs’ and the myth of monophyletic Xiphosura
https://www.research[...]
2013-01-01
[307]
논문
Heads, Hox and the phylogenetic position of trilobites
https://www.research[...]
CRC Press
2005-04-27
[308]
논문
Trilobite compound eyes with crystalline cones and rhabdoms show mandibulate affinities
https://www.nature.c[...]
2019-06-07
[309]
논문
Burgess Shale fossils illustrate the origin of the mandibulate body plan
https://www.nature.c[...]
2017-05
[310]
논문
Chelicerate neural ground pattern in a Cambrian great appendage arthropod
https://www.nature.c[...]
2013-10
[311]
논문
The impact of fossils on arthropod phylogeny
http://spiral.imperi[...]
2013-06
[312]
간행물
節足動物全体の分類体系・系統の現状
裳華房
2008
[313]
서적
Arthropods of the Lower Cambrian Chengjiang fauna, southwest China
https://foreninger.u[...]
Scandinavian University Press
1997
[314]
간행물
三葉虫綱
裳華房
2008
[315]
논문
Homeotic evidence for the appendicular origin of the labrum in Tribolium castaneum
https://doi.org/10.1[...]
2001-03-08
[316]
논문
Embryonic development of the scorpionfly ''Panorpa emarginata'' Cheng with special reference to external morphology (Mecoptera: Panorpidae)
https://onlinelibrar[...]
2009-08
[317]
논문
Arthropod Diversity in a Tropical Forest
https://www.research[...]
2012-12-14
[318]
논문
Phylum Arthropoda von Siebold, 1848
https://doi.org/10.1[...]
Magnolia Press
2011
[319]
웹사이트
What are trilobites? - Australian Museum
https://australianmu[...]
2018-10-25
[320]
문서
2010년대 이후의 많은 계통 분석에 있어서 거미류에 대해 측계통군으로 여겨지지만, 단계통군으로 여겨지는 분석 결과도 있다.
[321]
논문
What Is an “Arachnid”? Consensus, Consilience, and Confirmation Bias in the Phylogenetics of Chelicerata
https://www.mdpi.com[...]
2021-11
[322]
서적
ぞわぞわした生きものたち: 古生代の巨大節足動物
https://www.worldcat[...]
ソフトバンククリエイティブ
2012
[323]
문서
2010년대 이후의 계통 분석에서 단계통성이 찬반 양론이 있다.
[324]
논문
Increasing species sampling in chelicerate genomic-scale datasets provides support for monophyly of Acari and Arachnida
https://www.nature.c[...]
2019-05-24
[325]
논문
顎脚類(甲殻類)の分類と系統に関する研究の最近の動向
https://doi.org/10.1[...]
日本動物分類学会
2020
[326]
웹사이트
岡山県野生生物目録2019 - 岡山県ホームページ(自然環境課)
https://www.pref.oka[...]
2019-08-16
[327]
문서
갯지렁이류는 딱정벌레류에서 분기했을 가능성이 있다.
[328]
문서
강 계급이라고도 한다.
[329]
논문
The evolutionary diversity of barnacles, with an updated classification of fossil and living forms
https://doi.org/10.1[...]
2021-02-25
[330]
서적
古生物
https://www.worldcat[...]
学研プラス
2017
[331]
서적
アノマロカリス解体新書
https://www.worldcat[...]
2020
[332]
서적
大むかしの生きもの
https://www.worldcat[...]
講談社
2020
[333]
서적
ゼロから楽しむ古生物: 姿かたちの移り変わり
https://www.worldcat[...]
2021
[334]
서적
大昔の生きもの
https://www.worldcat[...]
ポプラ社
2014
[335]
서적
生命史図譜 = History of life
https://www.worldcat[...]
2017
[336]
논문
A new euarthropod with large frontal appendages from the early Cambrian Chengjiang biota
https://palaeo-elect[...]
[337]
논문
Cambrian bivalved arthropod reveals origin of arthrodization
https://www.research[...]
2012-12-07
[338]
논문
Evolutionary trade-off in reproduction of Cambrian arthropods
https://www.science.[...]
2020-05
[339]
논문
Fuxianhuiids are mandibulates and share affinities with total-group Myriapoda
https://doi.org/10.1[...]
2021-03-22
[340]
논문
A Burgess Shale mandibulate arthropod with a pygidium: a case of convergent evolution
https://onlinelibrar[...]
2021-11
[341]
논문
''Waptia fieldensis'' Walcott, a mandibulate arthropod from the middle Cambrian Burgess Shale
https://royalsociety[...]
[342]
논문
Fossils from South China redefine the ancestral euarthropod body plan
https://doi.org/10.1[...]
2020-01-08
[343]
논문
Early Cambrian fuxianhuiids from China reveal origin of the gnathobasic protopodite in euarthropods
https://www.nature.c[...]
2018-02-01
[344]
논문
Variation in appendages in early Cambrian bradoriids reveals a wide range of body plans in stem-euarthropods
https://www.nature.c[...]
2019-09-03
[345]
논문
Arthropod fossil data increase congruence of morphological and molecular phylogenies
https://www.research[...]
[346]
논문
Soft-part anatomy of the Early Cambrian bivalved arthropods Kunyangella and Kunmingella: significance for the phylogenetic relationships of Bradoriida
https://royalsociety[...]
2010-06-22
[347]
서적
Fossils and Strata, Morphology, Ontogeny and Phylogeny of the Phosphatocopina (Crustacea) from the Upper Cambrian Orsten of Sweden.
https://www.worldcat[...]
Wiley
2009
[348]
논문
Aquatic stem group myriapods close a gap between molecular divergence dates and the terrestrial fossil record
https://www.pnas.org[...]
2020-04-01
[349]
논문
REVIEW OF EARLY TRIASSIC THYLACOCEPHALA
https://riviste.unim[...]
2021-02-25
[350]
논문
New Genus and Species from the Cretaceous of Lebanon Links the Thylacocephala To the Crustacea
https://onlinelibrar[...]
2001
[351]
저널
Upper Ordovician Thylacocephala (Euarthropoda, Eucrustacea) from Bohemia indicate early ecological differentiation
https://onlinelibrar[...]
2021
[352]
저널
A new interpretation of the enigmatic fossil arthropod Anhelkocephalon handlirschi Bill, 1914 – important insights in the morphology of Cyclida
https://bioone.org/j[...]
2020-05
[353]
저널
Mandibulate convergence in an armoured Cambrian stem chelicerate
https://bmcevolbiol.[...]
2017-12
[354]
저널
A middle Cambrian arthropod with chelicerae and proto-book gills
https://www.nature.c[...]
2019-09
[355]
저널
Revision of the mollisoniid chelicerate(?) Thelxiope , with a new species from the middle Cambrian Wheeler Formation of Utah
https://peerj.com/ar[...]
2020-04-09
[356]
서적
ワンダフル・ライフ: バージェス頁岩と生物進化の物語
https://www.worldcat[...]
2000
[357]
저널
The Vicissicaudata revisited – insights from a new aglaspidid arthropod with caudal appendages from the Furongian of China
https://www.research[...]
2017-12
[358]
저널
A xandarellid artiopodan from Morocco – a middle Cambrian link between soft-bodied euarthropod communities in North Africa and South China
https://www.nature.c[...]
2017-02-17
[359]
웹사이트
Manufaktur
https://www.milbenka[...]
2019-08-17
[360]
웹사이트
Scientific Opinion on the re-evaluation of cochineal, carminic acid, carmines (E 120) as a food additive {{!}} EFSA
https://www.efsa.eur[...]
2022-08-12
[361]
웹사이트
GSFA Online Food Additive Details for Shellac
https://www.fao.org/[...]
2022-08-12
[362]
저널
Shellac as a multifunctional biopolymer: A review on properties, applications and future potential
https://www.research[...]
2022-08-31
[363]
저널
Economic valuation of the vulnerability of world agriculture confronted with pollinator decline
https://www.scienced[...]
2009-01-15
[364]
저널
Pest damage in sweet potato, groundnut and maize in north-eastern Uganda with special reference to damage by millipedes (Diplopoda)
http://www.sciencedi[...]
2005-01-01
[365]
저널
観賞用として扱われている甲殻類の現状: 企画趣旨とシンポジウムの内容(シンポジウム報告 観賞用として扱われている甲殻類の現状)
https://doi.org/10.1[...]
2014
[366]
웹사이트
Query on global tarantula trade
http://biodiversityr[...]
2022-08-12
[367]
저널
Searching the web builds fuller picture of arachnid trade
https://www.nature.c[...]
2022-05-19
[368]
웹사이트
History of Sea-Monkeys – The Original Sea-Monkeys
https://www.sea-monk[...]
2022-08-08
[369]
저널
Biochemical principle of ''Limulus'' test for detecting bacterial endotoxins
https://doi.org/10.2[...]
2007
[370]
저널
''Tachypleus'' Amebocyte Lysate Test Using in Transfusion Reaction
http://wprim.whocc.o[...]
2006-01-01
[371]
저널
Insects and their chemical weaponry: New potential for drug discovery
https://pubs.rsc.org[...]
2010-11-17
[372]
웹사이트
World malaria report 2021
https://www.who.int/[...]
2022-08-12
[373]
서적
Dr.夏秋の臨床図鑑 虫と皮膚炎
学研プラス
[374]
서적
Dr.夏秋の臨床図鑑 虫と皮膚炎
学研プラス
[375]
서적
Dr.夏秋の臨床図鑑 虫と皮膚炎
学研プラス
[376]
웹사이트
CDC - DPDx - American Trypanosomiasis
https://www.cdc.gov/[...]
2019-01-26
[377]
서적
Dr.夏秋の臨床図鑑 虫と皮膚炎
学研プラス
[378]
저널
Evolution of the bilaterian mouth and anus
2018-08-22
[379]
저널
The Ediacaran origin of Ecdysozoa: Integrating fossil and phylogenomic data
2022-03-10
[380]
저널
Arthropod Origins: Integrating Paleontological and Molecular Evidence
2020
[381]
저널
Pancrustacean Evolution Illuminated by Taxon-Rich Genomic-Scale Data Sets with an Expanded Remipede Sampling
2019
( 최근 20개의 뉴스만 표기 됩니다. )
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com