심해어
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
심해어는 햇빛이 도달하지 않는 깊은 바다에서 서식하는 물고기로, 독특한 신체 구조와 생태적 특징을 지니고 있다. 극한 환경에 적응하여 어둠 속에서 생존하기 위해 발광, 큰 눈, 젤리 형태의 몸 등을 진화시켰으며, 먹이를 찾고 짝을 유인하는 다양한 전략을 사용한다. 심해어는 일차성 심해어와 이차성 심해어로 분류되며, 다양한 분류군에 속한다. 일부 심해어는 식용으로 이용되기도 하지만, 남획으로 인한 자원 고갈의 위험에 직면해 있다.
더 읽어볼만한 페이지
- 심해어류 - 초롱아귀
초롱아귀는 아귀목 초롱아귀과에 속하는 물고기로, 암컷은 발달된 아래턱과 에스카를, 수컷은 암컷보다 작으며 후각으로 암컷을 찾아 부착하고, 에스카의 발광 박테리아로 먹이를 유인하며 심해에서 서식한다. - 심해어류 - 나무수염아귀과
나무수염아귀과는 심해에 사는 어류 과로, 복잡한 턱수염과 유인돌기를 가진 암컷, 암컷에 기생하는 왜소한 수컷, 몸의 왼쪽에 치우쳐진 항문 등의 특징을 가지며 5개 속, 약 27~29종이 알려져 있다. - 표시 이름과 문서 제목이 같은 위키공용분류 - 라우토카
라우토카는 피지 비치레부섬 서부에 위치한 피지에서 두 번째로 큰 도시이자 서부 지방의 행정 중심지로, 사탕수수 산업이 발달하여 "설탕 도시"로 알려져 있으며, 인도에서 온 계약 노동자들의 거주와 미 해군 기지 건설의 역사를 가지고 있고, 피지 산업 생산의 상당 부분을 담당하는 주요 기관들이 위치해 있다. - 표시 이름과 문서 제목이 같은 위키공용분류 - 코코넛
코코넛은 코코넛 야자나무의 열매로 식용 및 유지로 사용되며, 조리되지 않은 과육은 100g당 354kcal의 열량을 내는 다양한 영양 성분으로 구성되어 있고, 코코넛 파우더의 식이섬유는 대부분 불용성 식이섬유인 셀룰로오스이며, 태국 일부 지역에서는 코코넛 수확에 훈련된 원숭이를 이용하는 동물 학대 문제가 있다. - 한국어 위키백과의 링크가 위키데이터와 같은 위키공용분류 - 라우토카
라우토카는 피지 비치레부섬 서부에 위치한 피지에서 두 번째로 큰 도시이자 서부 지방의 행정 중심지로, 사탕수수 산업이 발달하여 "설탕 도시"로 알려져 있으며, 인도에서 온 계약 노동자들의 거주와 미 해군 기지 건설의 역사를 가지고 있고, 피지 산업 생산의 상당 부분을 담당하는 주요 기관들이 위치해 있다. - 한국어 위키백과의 링크가 위키데이터와 같은 위키공용분류 - 코코넛
코코넛은 코코넛 야자나무의 열매로 식용 및 유지로 사용되며, 조리되지 않은 과육은 100g당 354kcal의 열량을 내는 다양한 영양 성분으로 구성되어 있고, 코코넛 파우더의 식이섬유는 대부분 불용성 식이섬유인 셀룰로오스이며, 태국 일부 지역에서는 코코넛 수확에 훈련된 원숭이를 이용하는 동물 학대 문제가 있다.
| 심해어 | |
|---|---|
| 심해어 | |
 | |
| 서식지 | |
| 수심 범위 | '해수면에서 수심 ~' '해수면에서 수심 ~' '해수면에서 수심 ~' '해수면에서 수심 ' '해수면에서 수심 ' '해수면에서 수심 ~' |
| 환경 | |
| 수온 | |
| 특징 | |
| 특징 설명 | 심해에 사는 물고기 |
| 기타 | |
| 관련 정보 | 홋카이도 대학 종합 박물관 심포지엄 |
2. 정의
심해어는 햇빛이 비치는 해수면 아래, 즉 바다의 표해수층 또는 유광층 아래에 있는 어둠 속에서 서식하는 물고기다. 가장 흔한 심해어는 Lanternfish이며, 그 외에도 발광눈금돔, 달마 상어, 브리트슬마우스, 아귀, 바이퍼피쉬 및 등가시치의 여러 종이 심해어에 포함된다.[155]
심해는 극한 환경과 광대한 범위 때문에 천해에 비해 관찰과 연구가 어려워 생물이 존재하는지 여부는 오랫동안 불명확했다. 영국의 박물학자 에드워드 포브스는 1839년에 실시한 조사선에 의한 관측 결과를 바탕으로 "심해(300파좀(ファゾム)=수심 548미터 이하)에는 생물이 존재하지 않는다"는 "심해 무생물설"을 제창했다.[59] 그러나 그 후 저인망과 해저 케이블을 이용한 각국의 조사에 의해 심해에서 잇따라 생물이 채취되면서 이 설은 곧 부정되었다.
알려진 해양 종의 약 2%만이 원양 환경에 서식한다. 이는 해저 또는 해저에 서식하는 수저생물과는 대조적으로 물기둥에 서식하고 있음을 의미한다.[155] 심해 생물은 일반적으로 점심층(깊이 1000~4000m)과 심해원양대(깊이 4000~6000m)에 서식하지만, 생물발광과 같은 심해 생물의 특징은 중심층(깊이 200~1000m)에서도 볼 수 있다. 중심층표영대는 박광층이며 빛이 최소한이지만 여전히 측정 가능하다. 산소 최소층은 바다의 위치에 따라 다르지만 수심 700m에서 1000 사이 어딘가에 존재하며, 영양소가 가장 풍부한 곳이다. 점심층과 심해대는 무광층이며, 바다 영역에 빛이 투과하지 않는다. 심해는 거주 가능한 해양 공간의 약 75%를 구성한다.[156]
표해수대(0~200m)는 빛이 물을 투과하고 광합성이 일어나는 구역으로, 투광층으로도 알려져 있다. 일반적으로 수면 아래 수백 미터만 늘어나므로 바다 부피의 약 90%를 차지하는 심해는 어둠 속에 있다. 심해는 기온이 3°C(37.4° F)를 초과하는 경우는 거의 없으며, -1.8°C(28.76° F)까지 떨어지지 않는 매우 적대적인 환경이다. (350°를 초과할 수 있는 열수 분출 생태계 제외) C, 또는 662 °F), 저산소 수준, 20~1,000 기압(2~100 메가파스칼)의 압력[157]
3. 연구사
심해 생물의 존재를 결정적으로 증명한 것은 1872년부터 1876년에 걸쳐 실시된 영국 해군의 챌린저호에 의한 대규모 세계 일주 탐험 항해이다.[60] 이 항해가 가져온 방대한 해양학 연구 성과를 계기로 각국의 해양 조사가 본격화되고 심해어 연구의 역사도 막을 열었다.
맨몸으로 직접 심해에 잠수하는 것은 불가능하기 때문에, 심해 탐사에는 항상 어려움이 따랐다. 어망에 걸리거나 해안에 떠밀려 온 심해어도 때때로 귀중한 표본이 되었지만, 그들이 실제로 살아있는 모습(서식 환경과 생태)을 전하는 정보는 손상된 경우가 많았다. 19세기 후반 이후, 와이어 로프와 저인망의 개량으로 심해에서의 표본 채취가 가능해졌지만, 심해어를 직접 관찰하는 것은 여전히 쉽지 않았다.
1928년, 유인 잠수구(바티스피어)가 개발되어 마침내 심해어의 관찰이 가능해졌다. 바티스피어는 무동력이었지만, 수심 923미터까지의 잠수에 성공했다.[59] 그리고 1948년, 오귀스트 피카르에 의해 자체 동력을 가진 심해 탐사정 바티스카프가 건조되었다. 바티스카프는 여러 대의 후속기가 만들어져 심해어의 생태 관찰과 심해에서의 표본 채집에 강력한 수단을 제공했다. 20세기 후반부터 현대에 이르기까지, 일본의 しんかい6500, 소비에트 연방/러시아의 미르, 프랑스의 노틸 및 미국의 알빈호 등에 의한 조사를 통해 심해어의 생활 방식과 환경에 대한 적응에 관한 정보가 축적되고 있다.[62]
지금까지의 조사·연구 역사를 보면, 1960년, 바티스카프의 후속 기종 중 하나인 미국의 트리에스테호가 당시 이미 세계 최심부로 알려져 있던 마리아나 해구의 챌린저 해연을 목표로 유인 잠수를 실시했다. 탑승했던 자크 피카르(오귀스트의 아들)는 도달한 최심부(수심 10,900미터 전후)에서 "가자미와 비슷한 넙치의 일종"을 목격했다고 보고했다.[63] 반면, 일본의 무인 탐사정 "가이코"가 1998년에 실시한 조사에서는 동일 지점에서 어류를 확인할 수 없었다.[64] 21세기 초반 시점에서는 피카르의 "목격 보고"는 의문시되었고, 해삼의 일종을 잘못 본 것이 아니냐고 생각하는 연구자도 있다.[65]
확실한 과학적 근거를 가진 예로서, 지금까지 가장 깊은 곳에서 채집된 심해어는 (Cusk-eel)(아시로목)의 요미노아시로(''Abyssobrotula galatheae'')이다.[67] 덴마크의 조사대가 푸에르토리코 해구의 수심 8,372미터에서 이 종을 건져 올린 것은 1952년의 일이며, 학명에는 당시의 조사선 갈라테아(Galathea)호의 이름이 붙어 있다.[67] 그 외에, 꼼치과(망둑어목)의 심해꼼치[68] 및 아시로과의 소코보즈[69][70]가 7,000미터 이상의 깊이에서 관찰되고 있다.
해양연구개발기구(JAMSTEC)의 조사선 "카이레이"의 장비가 2017년 5월, 마리아나 해구의 수심 8,178미터에서 심해꼼치의 포식 행동 촬영에 성공했다. 2017년 8월 시점에서, 이것이 살아있는 어류의 영상 최심 기록이었다.[71]
심해어가 획득한 저수온·고압력 환경에 대한 적응 능력을 분석하기 위해서는, 가능한 한 표본을 살아있는 채로 포획하여 지상의 연구 시설에서 장기간 사육할 수 있는 것이 바람직하다. 하지만, 심해어를 생체 그대로 포획하는 데에는 실제로 많은 어려움이 있다. 가장 중요한 문제는 부상에 따른 급격한 감압과 수온 상승이며, 많은 경우 심해어에게 치명적인 손상을 준다.[72][73]
심해 탐사 기술의 발전과 함께 포획 장비의 개발·개량도 진행되어, 1970년대에는 저수온 유지 기능이 갖춰지게 되었다.[73] 1979년에 보고된 고압 유지 수조[74]는 1980년대에 걸쳐 개량이 거듭되어, 꼼치 등 부레가 발달한 저서성 심해어의 생체 포획을 가능하게 하였다.
2000년대 초 일본해양연구개발기구(JAMSTEC)에 의해 개발된 “딥 아쿠아리움”은 심해어의 포획과 고압 하에서의 사육을 단독으로 수행하기 위한 장치이다.[75] 심해 탐사정에 탑재한 딥 아쿠아리움으로 심해어를 포획한 후에는, 수조 내부의 고압 환경을 유지한 채 지상으로 운반, 사육하는 것이 가능하다. 물 교환이나 급식도 감압을 일으키지 않고 가능하며, 심해 생물 연구의 새로운 수단으로 기대되고 있다.[76]
심해의 생물학적 환경은 표층의 해류와 계절적 변화, 육지로부터의 물질 공급에 크게 의존하고 있으며, 심해어의 생태를 해양 환경과 연계하여 이해하기 위해서는 광범위하고 경시적인 조사가 필요하다. 또한, 저인망에 의한 남획이 대구류 등 대륙붕 주변에 서식하는 일부 식용종을 격감시키고 있다는 보고가 있으며, 어업상 중요한 심해어의 자원 조사의 필요성이 지적되고 있다.[77] 그러나, 특수한 탐사정과 채집 장비를 사용하는 심해어 조사는 다대한 비용을 필요로 하며, 대규모로 장기간에 걸친 생태 조사의 데이터는 매우 부족한 것이 현실이다.[78]
4. 환경

심해에서는 수심이 표영대(epipelagic zone)를 훨씬 넘어 뻗어 있으며, 이러한 심해대에 적응하여 서식하는 매우 다양한 종류의 중층성 어류를 지탱한다.[158] 심해에서는 주로 유기물질인 해양 눈이 수주 상층에서 끊임없이 떨어지는 비처럼 내린다. 그 기원은 생산적인 유광층 내 활동에 있다. 해양 눈에는 죽거나 죽어가는 플랑크톤, 원생생물(규조류 포함), 배설물, 모래, 그을음 및 기타 무기 먼지가 포함된다. 이러한 "눈송이"는 시간이 지남에 따라 성장하여 직경이 수 센티미터에 달할 수 있으며, 해저에 도달하기까지 수주 동안 이동한다. 그러나 해양 눈의 유기물 성분 대부분은 표영대 내에서 여정의 처음 1000m 이내에 미생물, 동물 플랑크톤 및 기타 여과 섭식 동물에 의해 소비된다. 이러한 방식으로 해양 눈은 심해 중층대 및 저서대 생태계의 기반으로 간주될 수 있다. 햇빛이 도달하지 않으므로 심해 유기체는 에너지원으로 해양 눈에 크게 의존한다. 심해(무광층)에는 빛이 없기 때문에 일차 생산자가 부족하다. 따라서 심해대의 대부분 유기체는 수직 기둥 상위 지역의 해양 눈에 의존한다.
랜턴피시, 릿지헤드, 해양 햇빛 물고기, 라이트피시과 같은 일부 심해 중층성 그룹은 열린 바다에서 고르게 분포하는 것이 아니라 구조적 오아시스, 특히 해산과 대륙 사면 위에서 훨씬 더 높은 풍부도로 나타나기 때문에 때때로 "pseudoceanic"(준해양성)이라고 불린다. 이 현상은 마찬가지로 구조물에 끌리는 먹이 종의 풍부함으로 설명된다.
정수압은 수심 10m당 1atm(0.1MPa)씩 증가한다.[159] 심해 유기체는 신체 내부의 압력이 외부에서 가해지는 압력과 같기 때문에 극심한 압력으로 인해 압착되지 않는다. 그러나 높은 내부 압력으로 인해 분자가 압축되어 세포막의 유동성이 감소한다. 세포막의 유동성은 생물학적 기능, 특히 단백질 생산의 효율성을 높이므로 유기체는 세포막 지질의 불포화 지방산 비율을 높임으로써 이러한 상황에 적응했다.[160] 내부 압력의 차이 외에도, 이러한 유기체는 표영대에 서식하는 유기체와 다른 대사 반응의 균형을 발달시켰다. "극한 환경의 유기체: Life at the Limits"의 저자인 David Wharton은 "생화학 반응에는 부피 변화가 수반된다. 반응의 결과로 부피가 증가하면 압력에 의해 억제되지만, 부피 감소와 관련이 있으면 향상된다"고 지적한다.[161] 즉, 그들의 대사 과정은 궁극적으로 어느 정도 유기체의 부피를 감소시켜야 한다는 것을 의미한다.
이러한 혹독한 환경에서 진화한 대부분의 물고기는 실험실 조건에서 생존할 수 없으며, 사육하려는 시도는 죽음으로 이어졌다. 심해 유기체는 기체로 채워진 공간(액포)을 포함한다. 기체는 고압 하에서 압축되고 저압 하에서 팽창한다. 이 때문에 이러한 유기체는 수면으로 올라오면 폭발하는 것으로 알려져 있다.[161]
4. 1. 수직 분포
표해수대 아래에서는 해수의 상태가 빠르게 변한다. 200미터에서 1000미터 사이에서는 빛이 거의 사라질 때까지 계속 희미해진다. 온도는 수온약층을 통과하며 7.8°C~3.9°C 사이로 떨어진다. 이 지점을 약광층[166] 또는 중층원양대라고 부른다. 이 구간에서 압력은 10m마다 1기압씩 계속 증가하며, 영양염류의 양은 용존산소량, 물의 순환 속도와 함께 감소한다.[167] 제2차 세계 대전 동안 새롭게 개발된 sonar를 사용한 이들은 낮 시간대에 보이는 300~500m 깊이의 가짜 해저[168]를 발견하고 혼란스러워했다. 이는 수중 음파 탐지기를 반사하는 부레를 가진 수백만 마리의 해양 생물들, 특히 중층원양대 어류들이 플랑크톤을 먹기 위해 해 질 녘에 얕은 물 위로 이동했기 때문에 나타난 현상이다. 이 층은 달이 떠 있을 때 더 깊어지고, 구름이 달을 가리며 지나갈 때 더 얕아질 수 있으며, 이 층을 심해 산란층이라고 부른다.[169]
대부분의 중층원양대 어류는 매일 수직 이동을 한다. 밤에는 동물성 플랑크톤을 따라 표해수층으로 이동하며, 낮에는 안전을 위해 깊은 곳으로 돌아간다.[167][170] 이러한 수직 이동은 종종 큰 수직 거리에서 발생하며, 부레의 도움을 받아 이루어진다. 물고기가 위로 올라가기를 원할 때, 부레는 부풀어 오른다. 중층원양대의 높은 압력으로 인해, 이 과정은 상당한 에너지가 필요하다. 물고기가 상승할 때, 부레가 터지는 것을 막기 위해서는 부레 내부의 압력이 조절되어야 한다. 물고기가 깊은 곳으로 돌아가고 싶을 때, 부레의 바람은 빠진다.[171] 일부 중층원양대 어류는 온도가 10°C에서 20°C 사이로 변하는 수온약층을 통해 매일 이동하므로 온도변화에 잘 견딘다.[172]
중층원양대 어류들은 근육질의 몸, 골화된 뼈, 비늘, 잘 발달된 아가미와 중추신경계, 그리고 큰 심장과 신장을 가지고 있다. 중층원양대의 플랑크톤을 먹이로 하는 생물들은 작은 입과 미세한 아가미 돌기를 가지고 있지만, 육식성 어류들은 더 큰 입과 더 거친 아가미 돌기를 가지고 있다.[167]
심해에서는 수심이 표영대(epipelagic zone)를 훨씬 넘어 뻗어 있으며, 이러한 심해대에 적응하여 서식하는 매우 다양한 종류의 중층성 어류를 지탱한다. 심해에서는 주로 유기물질인 해양 눈이 수주 상층에서 끊임없이 떨어지는 비처럼 내린다. 그 기원은 생산적인 유광층 내 활동에 있다. 해양 눈에는 죽거나 죽어가는 플랑크톤, 원생생물(규조류 포함), 배설물, 모래, 그을음 및 기타 무기 먼지가 포함된다. 이러한 "눈송이"는 시간이 지남에 따라 성장하여 직경이 수 센티미터에 달할 수 있으며, 해저에 도달하기까지 수주 동안 이동한다. 그러나 해양 눈의 유기물 성분 대부분은 표영대 내에서 여정의 처음 1000m 이내에 미생물, 동물 플랑크톤 및 기타 여과 섭식 동물에 의해 소비된다. 이러한 방식으로 해양 눈은 심해 중층대 및 저서대 생태계의 기반으로 간주될 수 있다. 햇빛이 도달하지 않으므로 심해 유기체는 에너지원으로 해양 눈에 크게 의존한다. 심해(무광층)에는 빛이 없기 때문에 일차 생산자가 부족하다. 따라서 심해대의 대부분 유기체는 수직 기둥 상위 지역의 해양 눈에 의존한다.
랜턴피시, 릿지헤드, 해양 햇빛 물고기, 라이트피시과 같은 일부 심해 중층성 그룹은 열린 바다에서 고르게 분포하는 것이 아니라 구조적 오아시스, 특히 해산과 대륙 사면 위에서 훨씬 더 높은 풍부도로 나타나기 때문에 때때로 "pseudoceanic"(준해양성)이라고 불린다. 이 현상은 마찬가지로 구조물에 끌리는 먹이 종의 풍부함으로 설명된다.
정수압은 수심 10m당 1atm(0.1MPa)씩 증가한다. 심해 유기체는 신체 내부의 압력이 외부에서 가해지는 압력과 같기 때문에 극심한 압력으로 인해 압착되지 않는다. 그러나 높은 내부 압력으로 인해 분자가 압축되어 세포막의 유동성이 감소한다. 세포막의 유동성은 생물학적 기능, 특히 단백질 생산의 효율성을 높이므로 유기체는 세포막 지질의 불포화 지방산 비율을 높임으로써 이러한 상황에 적응했다. 내부 압력의 차이 외에도, 이러한 유기체는 표영대에 서식하는 유기체와 다른 대사 반응의 균형을 발달시켰다. "극한 환경의 유기체: Life at the Limits"의 저자인 David Wharton은 "생화학 반응에는 부피 변화가 수반된다. 반응의 결과로 부피가 증가하면 압력에 의해 억제되지만, 부피 감소와 관련이 있으면 향상된다"고 지적한다. 즉, 그들의 대사 과정은 궁극적으로 어느 정도 유기체의 부피를 감소시켜야 한다는 것을 의미한다.
|thumb|right|사람들은 살아있는 주름상어를 거의 만나지 못하기 때문에 위험이 거의 없다(하지만 과학자들은 실수로 이빨을 조사하다가 자신을 다치기도 했다).
이러한 혹독한 환경에서 진화한 대부분의 물고기는 실험실 조건에서 생존할 수 없으며, 사육하려는 시도는 죽음으로 이어졌다. 심해 유기체는 기체로 채워진 공간(액포)을 포함한다. 기체는 고압 하에서 압축되고 저압 하에서 팽창한다. 이 때문에 이러한 유기체는 수면으로 올라오면 폭발하는 것으로 알려져 있다.
바다를 깊이에 따라 수직 방향으로 구분하면 표층, 중층, 심층, 심해층, 초심해층으로 나눌 수 있다.[80] 이 구분은 수층 구분대라고도 불린다. 일반적으로 중층 이하에 주된 서식 수심을 가진 어류가 심해어로 분포되어 있다.
중심층(수심 200~1,000m)에는 광합성에는 불충분하지만 미약한 햇빛이 도달한다.[81] 주요 온도약층(수온이 급격하게 변하는 수층)의 대부분이 이 영역에 존재하며, 그 아래에는 물리적으로 안정적이고 변화가 적은 심해 특유의 환경이 펼쳐져 있다. 중심층의 부유성 심해어는 현재까지 약 750종이 알려져 있으며,[53] 아귀목에 속하는 측광어과·가슴빛등어과·아귀과 어류와 멸치목의 멸치과 어류가 종류와 수량 모두에서 압도적이다. 이들 그룹은 극지방의 바다를 포함한 전 세계 해양에 널리 분포하며, 그 생물량은 막대하다. 특히 악마멸치속(측광어과)의 무리는 지구상의 척추동물 중 가장 많은 개체 수를 가진 것으로 생각된다.[82]
저서어로는 연골어류인 은상어목·각상어목의 무리 외에, 아귀과·꼼치과(대구목), 아시로과(아시로목) 및 도치과(솔치목)가 우세하다. 그 외에도 등불멸치과(청어목), 뱀장어과(뱀장어목) 및 뚝지과(농어목) 등 심해에서는 비교적 다양한 어종이 관찰되는 영역이다. 저서성 심해어의 서식 범위는 수심보다 해저 지형의 영향을 강하게 받으며, 심해층에 걸쳐 분포하는 종도 많다. 심해성 저서어는 중심층과 심해층 이하를 합쳐 1,000종 이상이 기재되어 있다.[53]
중층원(수심 1,000 - 3,000미터)은 빛이 닿지 않는 암흑의 세계이다. 수온은 2~5℃로 안정적인 반면, 생물이 이용할 수 있는 유기물의 양은 표층의 5%에도 미치지 못하며, 수심이 깊어짐에 따라 급격히 감소한다.[84] 중층원의 유영성 심해어에는 최소 200종이 포함된다.[53] 종 수로는 등불안경어류가 우세하며, 그 외에는 고래상어과(고래상어목), 등가시치과(대구목), 풍선장어목의 어류 및 악마쥐치속의 일부가 서식한다. 아귀와 도롱이치류는 이 영역에서도 많은 수를 차지하는 저서성 심해어이며, 그 외에는 뱀장어과(장어목), 홍구어과(아귀목) 등이 분포한다.
심해층(수심 3,000~6,000m)이 되면 수온은 1~2℃ 정도로 낮아지며 거의 변화하지 않는다.[85] 300기압을 넘는 수압은 생물의 세포 활동에 영향을 준다. 유영성 심해어는 거의 사라지고, 아귀과·꼼치과·솔치과의 저서어만 볼 수 있다.
초심해층(6,000미터 이하)은 해구의 심부에 한정되며, 전체 해저 면적의 2%에도 미치지 못한다. 수압이 600기압을 넘는 이 해역에 사는 심해어는 심해층과 마찬가지로 꼼치과, 히메노토로과 및 아귀과에 속하는 일부 저서어만이 알려져 있다.
4. 2. 수평 분포
심해는 수심이 표영대(epipelagic zone)를 훨씬 넘어 뻗어 있으며, 이러한 심해대에 적응하여 서식하는 매우 다양한 종류의 중층성 어류를 지탱한다. 심해에서는 주로 유기물질인 해양 눈이 수주 상층에서 끊임없이 떨어지는 비처럼 내린다. 해양 눈에는 죽거나 죽어가는 플랑크톤, 원생생물(규조류 포함), 배설물, 모래, 그을음 및 기타 무기 먼지가 포함된다. 이러한 "눈송이"는 시간이 지남에 따라 성장하여 직경이 수 센티미터에 달할 수 있으며, 해저에 도달하기까지 수주 동안 이동한다. 그러나 해양 눈의 유기물 성분 대부분은 표영대 내에서 여정의 처음 1000m 이내에 미생물, 동물 플랑크톤 및 기타 여과 섭식 동물에 의해 소비된다. 이러한 방식으로 해양 눈은 심해 중층대 및 저서대 생태계의 기반으로 간주될 수 있다. 햇빛이 도달하지 않으므로 심해 유기체는 에너지원으로 해양 눈에 크게 의존한다. 심해(무광층)에는 빛이 없기 때문에 일차 생산자가 부족하다. 따라서 심해대의 대부분 유기체는 수직 기둥 상위 지역의 해양 눈에 의존한다.
랜턴피시, 릿지헤드, 해양 햇빛 물고기, 라이트피시과 같은 일부 심해 중층성 그룹은 열린 바다에서 고르게 분포하는 것이 아니라 구조적 오아시스, 특히 해산과 대륙 사면 위에서 훨씬 더 높은 풍부도로 나타나기 때문에 때때로 "pseudoceanic"(준해양성)이라고 불린다.
정수압은 수심 10m(32.8ft)당 1atm(0.1MPa)씩 증가한다. 심해 유기체는 신체 내부의 압력이 외부에서 가해지는 압력과 같기 때문에 극심한 압력으로 인해 압착되지 않는다. 그러나 높은 내부 압력으로 인해 분자가 압축되어 세포막의 유동성이 감소한다. 세포막의 유동성은 생물학적 기능, 특히 단백질 생산의 효율성을 높이므로 유기체는 세포막 지질의 불포화 지방산 비율을 높임으로써 이러한 상황에 적응했다.
대부분의 물고기는 실험실 조건에서 생존할 수 없으며, 사육하려는 시도는 죽음으로 이어졌다. 심해 유기체는 기체로 채워진 공간(액포)을 포함한다. 기체는 고압 하에서 압축되고 저압 하에서 팽창한다. 이 때문에 이러한 유기체는 수면으로 올라오면 폭발하는 것으로 알려져 있다.
해양은 대륙붕의 가장자리를 경계로 육지에 가까운 연안역과 육지에서 멀리 떨어진 외양으로 수평적으로 구분된다. 심해에는 광합성을 하는 식물과 같은 기초생산자가 존재하지 않으며, 심해 생물의 에너지원이 되는 유기물은 주로 천해와 육지에서 공급된다. 이 때문에 일반적으로 심해어(및 다른 심해 생물)는 육지에 가까운 해역일수록 많고, 외양으로 갈수록 적어진다. 또한, 열대 해역의 외양에서는 대류가 일어나지 않기 때문에 표층의 생물이 적고, 이용 가능한 퇴적물이 부족한 황량한 해저가 펼쳐지는 경우도 있다.
해저 지형의 특징은 각 지역에 따라 다르며, 저서성 심해어의 분포에 큰 영향을 준다. 한편, 심해 중층 환경은 비교적 안정적이고 균질하기 때문에, 유영성 심해어는 광범위한 분포역을 가진 종이 많다. 태평양, 인도양, 대서양 등 세 개의 대양 모두에 분포하는 심해어도 적지 않으며, 세계종(汎世界種)이라고 불린다. 유영성 심해어의 생물군계는 주로 기후와 대륙·도서 지형의 영향을 받아 약 20개로 분류되며, 이는 다른 생물군과 비교하여 현저히 적은 구분 수이다. 햇살멸치와 긴꼬리치메소류 등 대륙붕을 따라 분포하는 유영성 심해어를 "pseudoceanic(위외양성)"이라고 특별히 구분하여 부르기도 한다.
4. 3. 고압에 대한 적응
물고기가 바다 속으로 더 깊이 들어가면, 머리 위의 물의 무게로 인해 수압이 증가한다. 이 증가된 압력은 수심 10m 마다 약 1 기압에 해당된다.[44] 점심해수층의 밑바닥에 있는 물고기의 경우 이 압력은 약 400기압에 달한다.[176]
심해 생물들은 세포와 생리적 수준에서 적응하여 압력이 큰 환경에서 생존할 수 있다. 높은 수준의 외부 압력은 대사 과정과 생화학적 반응이 어떻게 진행되는지에 영향을 미친다. 많은 화학 반응의 평형은 압력에 의해서 방해받고, 압력은 부피 증가를 일으키는 과정을 억제할 수 있다. 많은 생물학적 과정에서 중요한 요소인 물은 부피 변화에 매우 민감하게 반응하는데 이는 주로 세포 액의 성분이 물 구조에 영향을 미치기 때문이다. 그러므로 물 조직의 변화를 유도하는 효소 반응은 물의 부피를 효과적으로 변화시킨다. 반응을 촉매시키는 단백질은 일반적으로 약한 결합에 의해 함께 유지되며, 반응은 일반적으로 부피 증가를 수반한다.[177]
이러한 변화에 적응하기 위해 심해어의 단백질 구조와 반응 기준이 이러한 고압 조건에서도 반응을 수행하기 위해 압력을 견딜 수 있도록 조정되었다. 고압 환경에서 이중층 세포막은 유동성의 손실을 경험한다. 심해 세포막은 불포화 지방산의 비율이 높은 인지질 이중층을 선호하며, 이는 해수면 세포막보다 높은 유동성을 유도한다.
심해 생물종은 지표면 생물에 비해 엔트로피와 엔탈피의 변화가 낮은데, 이는 고압과 저온 환경이 엔탈피 변화를 선호하고 엔트로피가 주도하는 반응에 대한 의존도가 감소하기 때문이다. 구조적인 관점에서, G-액틴의 3차 구조로 인한 심해어의 구상 단백질은 표면에 사는 물고기의 단백질에 비해 상대적으로 단단하다. 심해어의 단백질이 표면종 물고기와 구조적으로 다르다는 사실은 심해어의 근육 섬유에서 나오는 액틴이 매우 열에 강하다는 관찰을 통해서 명백하게 밝혀졌다. 이런 단백질은 단백질의 3차 구조에서 결합 변형에 의해 구조적으로 강화되며, 이는 또한 높은 수준의 열안전성을 유도한다.[177] 단백질은 3차 구조에서 결합 변형에 의새 압력에 저항하기 위해서 구조적으로 강화된다. 그러므로 호열성 사막 파충류의 높은 체온과 유사한 높은 수준의 정수압은 단단한 단백질 구조를 선호한다.
Na+/K+-ATPase는 삼투 조절에서 중요한 역할을 하는 리포단백질 효소로, 수압의 영향을 크게 받는다. 심해어의 아가미 조직 1g당 Na+/K+ -ATPase 활동은 낮지만, 심해어의 Na+/K+ -ATPase는 얕은 물에 사는 종에 비해 수압에 대한 내성이 훨씬 높다. 약 2000m 깊이에 서식하는 ''C. acrolepis''와 약 4000m 깊이에 서식하는 심해저대 종인 ''C. armatus''를 비교하면, ''C. armatus''의 Na+/K+ -ATPase가 압력에 훨씬 덜 민감하다는 것을 알 수 있다.[46]
5. 신체 구조
심해어는 이 지역에서 살아남기 위해 다양한 적응을 진화시켰다. 이러한 물고기의 많은 수가 자연 조명이 없는 지역에 살기 때문에 먹이와 짝을 찾고 포식자를 피하는 데 시력에만 의존할 수 없다. 심해어는 자신이 사는 극한의 수광대(sub-photic region)에 적응하여 진화했다. 이러한 생물들 중 많은 수가 눈이 멀었고, 먹이를 잡고 잡히는 것을 피하기 위해 국소적인 압력과 냄새의 변화에 대한 감각과 같은 다른 감각에 의존한다. 눈이 멀지 않은 종들은 생물발광 빛을 이용할 수 있는 크고 민감한 눈을 가지고 있다. 이러한 눈은 인간의 눈보다 최대 100배나 더 빛에 민감할 수 있다. 로돕신(Rhodopsin)(Rh1)은 동물이 어두운 곳에서 볼 수 있도록 돕는 눈의 간상세포에서 발견되는 단백질이다. 대부분의 척추동물은 일반적으로 하나의 Rh1 옵신 유전자를 가지고 있는 반면, 일부 심해어는 여러 개의 Rh1 유전자를 가지고 있으며, 은빛가시돔(silver spinyfin)(''Diretmus argenteus'')라는 한 종은 38개의 유전자를 가지고 있다. 이러한 Rh1 유전자의 증가는 심해어가 바다 깊은 곳에서 볼 수 있도록 돕는다. 또한 포식을 피하기 위해 많은 종들이 주변 환경과 어울리도록 어두운 색을 띤다.
많은 심해어는 생물발광을 하며, 어둠에 적응한 매우 큰 눈을 가지고 있다. 생물발광 생물은 루시페린 분자의 교반을 통해 생물학적으로 빛을 생성할 수 있으며, 이는 빛을 생성한다. 이 과정은 산소가 존재하는 경우에만 수행되어야 한다. 이러한 생물들은 중층해수대(mesopelagic region) 이하(200m 이하)에서 흔하다. 심해어의 50% 이상과 일부 갑각류 및 오징어 종이 생물발광 능력을 가지고 있다. 이러한 생물의 약 80%는 발광 박테리아를 포함하는 발광 세포인 광포(photophores)를 가지고 있으며, 어두운 색으로 경계가 지어져 있다. 이러한 광포 중 일부는 인간의 눈과 같은 렌즈를 포함하고 있으며, 이를 통해 빛의 방출을 강화하거나 줄일 수 있다. 빛을 생성하는 능력은 생물 에너지의 1%만 필요하며, 먹이를 찾고 먹이를 유인하는 데 사용되며(예: 낚시하는 물고기), 순찰을 통해 영역을 차지하고, 의사소통하고 짝을 찾으며, 포식자를 산만하게 하거나 일시적으로 눈멀게 하여 탈출하는 등 여러 목적을 가지고 있다. 또한 일부 빛이 여전히 투과되는 중층해수대(mesopelagic)에서는 일부 생물이 위에서 오는 빛의 색깔과 강도에 맞춰 배를 밝힘으로써 그림자가 드리워지지 않도록 아래쪽 포식자로부터 위장한다. 이러한 전술을 역광조명(counter-illumination)이라고 한다.
심해어의 생활주기는 전적으로 심해에서 이루어질 수 있지만, 일부 종은 얕은 물에서 태어나 성숙하면 가라앉는다. 알과 유충이 서식하는 깊이에 관계없이, 일반적으로 원양성(pelagic)이다. 이러한 플랑크톤성 – 표류하는 – 생활 방식에는 중성 부력이 필요하다. 이를 유지하기 위해 알과 유충에는 종종 혈장에 기름 방울이 포함되어 있다. 이러한 생물이 완전히 성숙한 상태가 되면 수층에서 자신의 위치를 유지하기 위해 다른 적응이 필요하다. 일반적으로 물의 밀도는 상향력 – 생물이 뜨게 만드는 부력의 측면 – 을 유발한다. 이에 대응하기 위해 생물의 밀도는 주변 물의 밀도보다 커야 한다. 대부분의 동물 조직은 물보다 밀도가 높기 때문에 뜨도록 균형을 맞춰야 한다. 많은 생물은 부레(gas cavities)를 발달시켜 떠 있지만, 높은 환경 압력으로 인해 심해어는 일반적으로 이 기관을 가지고 있지 않다. 대신 유체역학적 양력을 제공하기 위해 하이드로포일과 유사한 구조를 보인다. 또한 물고기가 서식하는 깊이가 깊을수록 살이 더 젤리 같은 형태이고 뼈 구조가 더 최소화되는 것으로 나타났다. 높은 지방 함량, 골격 무게 감소 – 크기, 두께 및 미네랄 함량 감소를 통해 달성 – 및 물 축적을 통해 조직 밀도를 줄인다. 이로 인해 표층 어류보다 느리고 민첩성이 떨어진다. 심해어의 체형은 일반적으로 지속적인 수영보다는 주기적인 수영 돌풍에 더 적합하다.
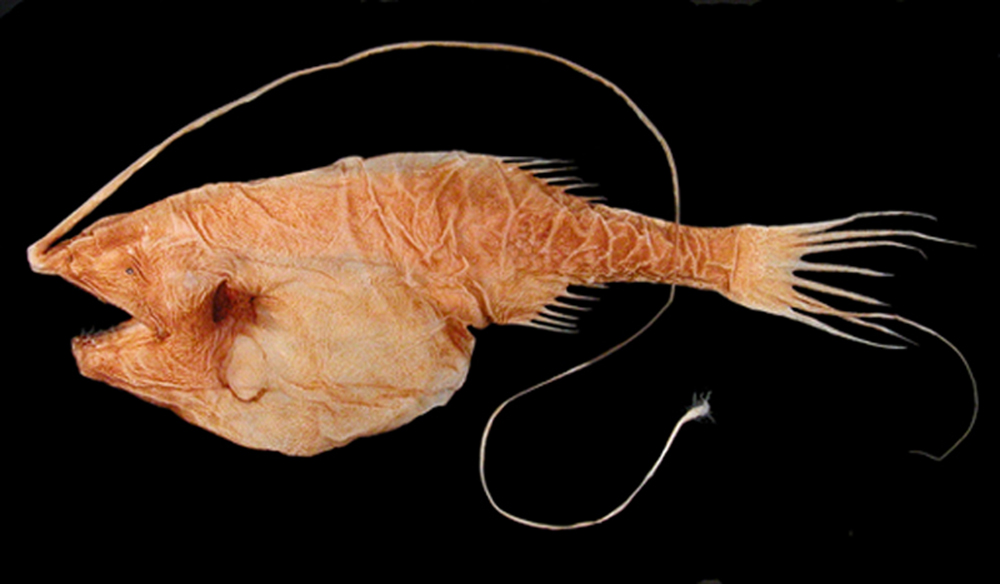
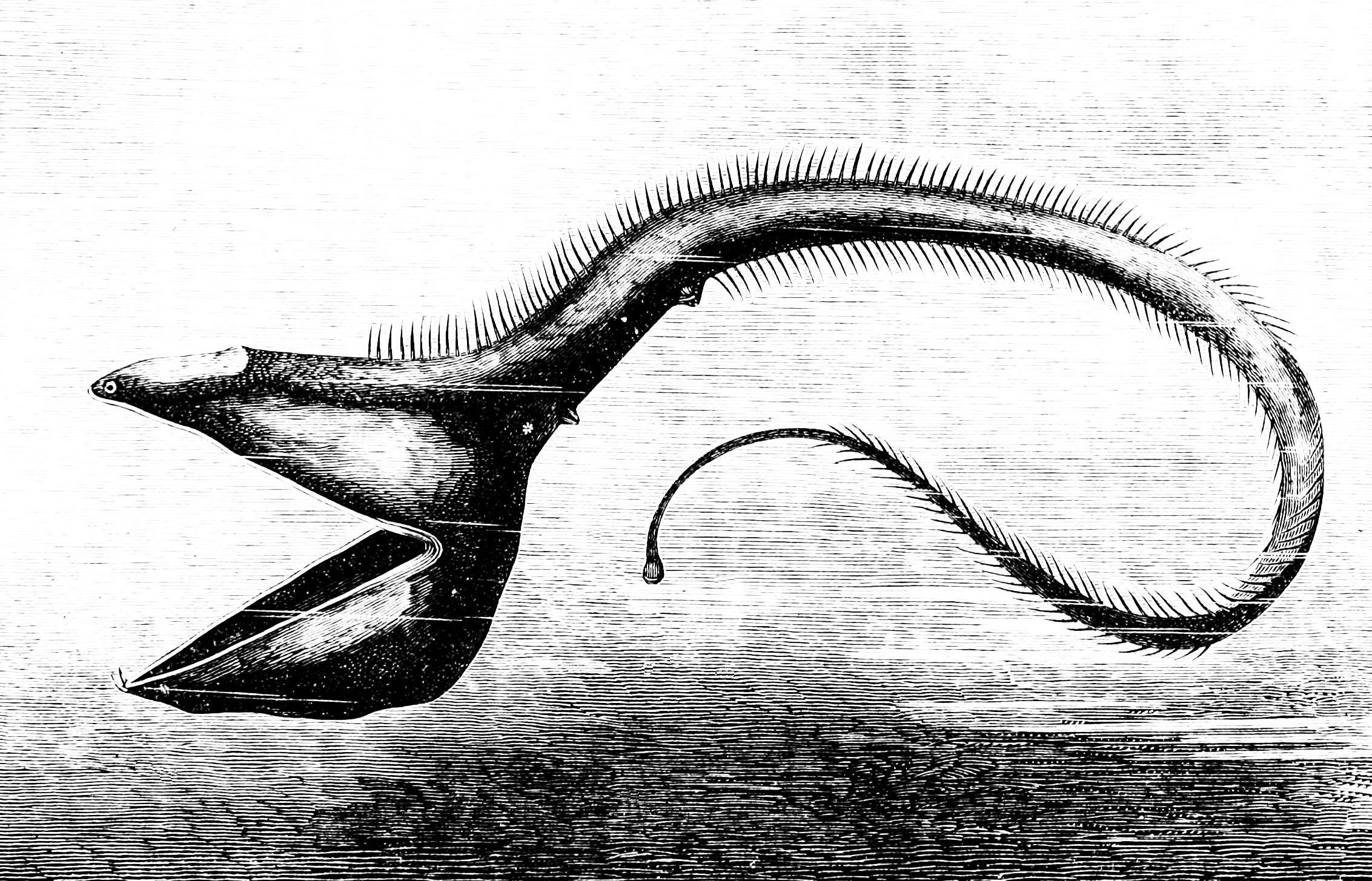
심해 환경에 도달하는 광합성 빛의 수준이 낮기 때문에 대부분의 물고기는 상층부에서 가라앉는 유기 물질이나 드문 경우 열수 분출공에 의존하여 영양분을 섭취해야 한다. 이로 인해 심해는 얕은 지역보다 생산성이 훨씬 낮다. 또한 원양 환경의 동물은 드물고 먹이가 자주 나타나지 않는다. 이 때문에 생물은 살아남을 수 있도록 하는 적응이 필요하다. 어떤 것은 깊은 바다의 칠흑 같은 어둠 속에서 먹이를 찾거나 짝을 유인하는 데 도움이 되는 긴 더듬이를 가지고 있다. 특히 심해 낚시하는 물고기는 얼굴에서 돌출된 긴 낚싯대와 같은 적응을 가지고 있으며, 그 끝에는 벌레처럼 꿈틀거리는 생물발광 피부 조각이 있어 먹이를 유인한다. 어떤 것은 자신과 크기가 같거나 더 큰 다른 물고기를 잡아먹어야 하며, 효율적으로 소화하는 데 도움이 되는 적응이 필요하다. 날카로운 이빨, 경첩이 달린 턱, 불균형적으로 큰 입, 팽창할 수 있는 몸은 심해어가 이러한 목적으로 가지고 있는 특징 중 일부이다. 큰입장어는 이러한 특징을 보이는 생물의 한 예이다.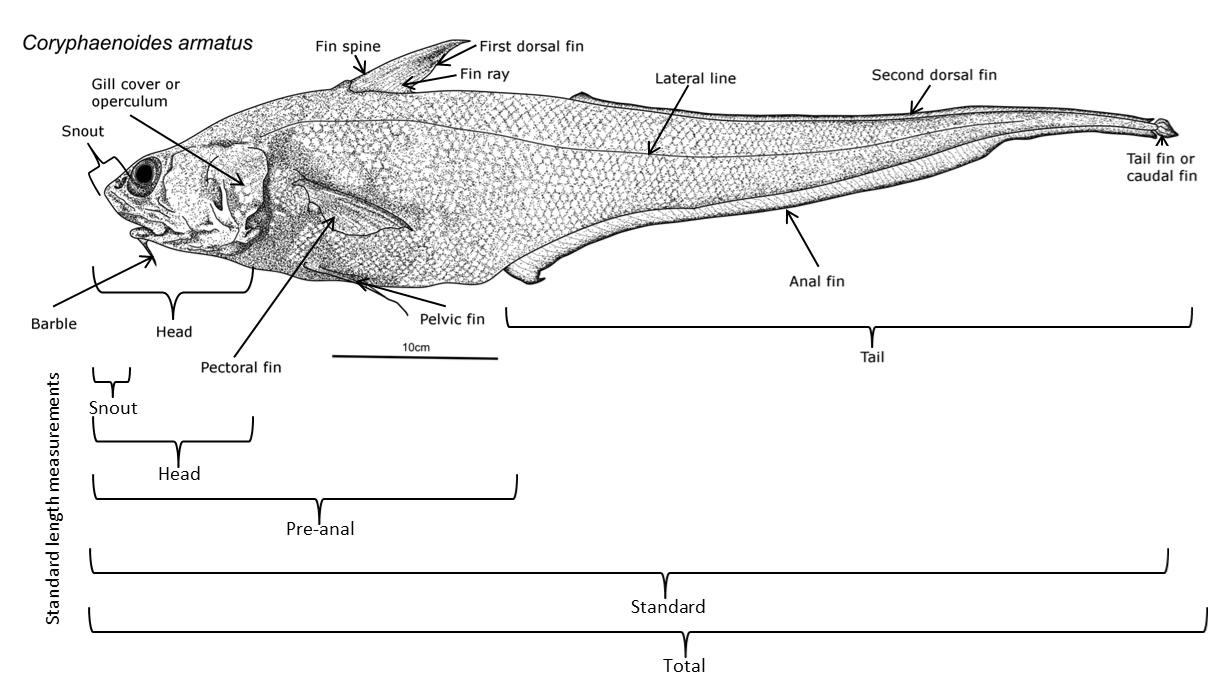

심해에는 태양빛이 거의 도달하지 않을 뿐만 아니라, 고수압, 저수온, 저산소 농도, 이용 가능한 유기물이 적다는 등 생물에게는 가혹한 조건이 갖춰져 있다. 심해 생물에 공통적으로 볼 수 있는 고수압에 대한 적응으로는 세포막의 유동성 및 압력에 대한 효소의 감수성이 저하되어 있는 것을 들 수 있다.[86]
=== 골격 및 근육 ===
어류는 몸 전체에 포함된 단백질과 골격의 비중이 일반적으로 해수보다 크다.[87] 천해어는 유영에 따른 양력이나 부레에 공기를 들이마심으로써 부력을 얻지만, 이용 가능한 에너지가 부족한 심해에서는 심해어는 가능한 한 유영하지 않고 부력을 확보해야 한다.[87] 많은 심해어는 뼈, 연골 및 근육 등 체내의 고밀도 조직이 감소하고, 대신 저비중의 수분과 지방을 다량 함유하고 있다.[87] 배지느러미와 그 지지 골격, 또는 머리의 골격을 퇴화시킨 것과 비늘, 가시를 가지지 않은 종류도 많으며, 경량화를 위한 하나의 수단으로 여겨진다.[57]
=== 부레 ===
부레(鰾)는 천해어가 부력을 얻는 일반적인 수단이지만, 심해에서는 극도의 고수압 때문에 일반적인 가스 교환에 의한 부레의 기능을 기대할 수 없다. 고수압과 급격한 압력 변화에 견디기 위해 심해어에는 부레의 벽을 견고한 구아닌결정으로 덮거나, 내용물을 기체가 아닌 지방이나 왁스로 대체하는 등의 적응이 나타난다.[95][88]
수심이 커짐에 따라 고수압에 역행하여 가스 교환(특히 분비)을 하는 데 대한 부담도 증가한다. 중심층 유영성 심해어(천해로의 이동을 하지 않는 그룹)에서는 부레는 일반적으로 퇴화되어 있으며, 더 수심이 깊은 점심층에서는 부레를 갖지 않는 종류가 많다.[95]
=== 안구 ===
심해어는 서식 환경에 따라 독특한 안구 구조를 가진다. 수심 약 1,000m까지는 미약한 태양광이 도달하므로, 이 영역에 서식하는 심해어는 몸에 비해 매우 큰 안구를 가지거나, 데메에소과, 무네에소과, 요코에소과 어류 등 최소 11개 과의 심해어는 관 모양으로 변형된 '''관상안'''을 가진다.[89] 심해에 도달하는 빛은 산란과 굴절 때문에 태양의 위치와 관계없이 항상 바로 위에서 내리쬐며, 해가 진 후에도 광량 변화가 적다.[90] 보우엔기요과 등 일부 예외를 제외하고, 대부분의 관상안은 해수면 방향의 빛에 대응하여 바로 위를 향하고 있다.
반면, 동굴 생물은 암흑 조건하에서 눈이 퇴화한 예가 많지만, 심해어는 약간의 빛이 들어오고, 종에 따라 천해로 이동하거나 발광 생물이 많다는 점이 영향을 미치는 것으로 보인다.
1,000m 이하의 심해층은 빛이 전혀 없는 암흑 세계이며, 이 영역에는 오목하게 들어간 작은 눈을 가진 심해어가 많다. 소코오쿠메우오과처럼 눈이 피부 속에 파묻히거나, 초우친하다카처럼 판상의 망막만 남은 심해어도 있지만, 빛 감지 기능은 여전히 남아 있어 퇴화가 아닌 특수화로 간주된다.[91] 심해층에서 드물게 깜빡이는 생물발광을 포착하기 위해 가늘고 긴 작은 안구가 더 적합하다는 보고도 있다.[92] 이러한 눈은 일반적인 눈보다 공간 분해능이 뛰어나 20~30m 정도 떨어진 곳의 발광을 포착하는 데 적합하며, 유영 능력이 낮은 심해어에게 시야를 좁은 범위로 제한하는 것은 에너지 효율 측면에서 합리적이다.[93]
=== 소화 기관 ===
육식성 심해어는 체형에 비해 상당히 큰 입과 이빨을 가진 경우가 많다.[95] 특히 신기한 물고기목에 속하는 심해꼼치·흡혈귀물뱀장어류는 매우 큰 입으로 잘 알려져 있는데, 이들의 두개골은 작지만 극도로 발달한 아가미뼈가 큰 입을 지탱한다.[96] 흡혈귀물뱀장어 ''Eurypharynx pelecanoides''는 전 세계 온대 해역의 심해(550~3,000미터)에서 비교적 흔하게 발견된다.[94] 심해꼼치는 날카로운 이빨로 큰 먹이를 삼키지만, 흡혈귀물뱀장어는 아가미에 이빨이 거의 없어 작은 물고기나 플랑크톤을 모아 먹는다.[96]
발달된 치열 또한 육식성 심해어의 특징인데, 귀신상어(귀신상어과)는 날카로운 송곳니 모양의 이빨을 가졌다.[95] 악어상어·등불상어류 일부는 안쪽으로 구부러진 이빨을 가져 먹이를 놓치지 않도록 한다.[95]
등불상어류와 큰입우럭과 어류 등은 식도나 위를 크게 확장시킬 수 있다.[97] 큰입우럭은 자신의 몇 배나 되는 먹이를 삼킬 수 있으며, 복부가 비정상적으로 부풀어 오른 상태로 포획되기도 한다. 멜라닌 등의 색소 침착으로 흑색화된 장관을 가진 심해어도 있는데, 이는 발광 생물을 포식했을 때 소화관을 투과한 빛이 천적을 유인하는 것을 막기 위함이다.[95]
심해 환경에서는 지속적인 포식이 어려워 에너지 축적이 중요하다. 간은 심해어에게 중요한 에너지 저장 기관이며, 긴꼬리꼼치과 등은 매우 크고 지방이 풍부한 간을 가지고 있다.[131] 요로이다라/Abyssal grenadier영어(솔치과)의 간 또한 지방·글리코겐을 풍부하게 함유하고 있으며, 약 180일 동안 먹이를 얻지 못해도 생명을 유지할 수 있다고 추정된다.[131]
지방은 바닷물보다 비중이 작아, 간에 다량의 지방분을 축적하는 것은 부력 확보에도 기여한다. 상어과·까치상어과(각상어목) 등 부레가 없는 심해성 연골어류 중에는 체중의 25%에 달하는 거대한 간을 가진 것도 있다.[88]
=== 체색 ===
심해에서는 비교적 밝은 중층에 사는 어류에서는 체표면의 '''은화'''에 의한 의태가 보인다.[98] 가시볏과 어류는 두께 수 밀리미터의 납작한 몸을 가지고 있으며, 표면은 알루미늄 호일과 같은 광택이 있는 은색을 띤다. 그들의 체표면에는 과닌에 의한 미세한 반사성 결정이 여러 층에 걸쳐 규칙적으로 나란히 배열되어 있으며,[99] 거울처럼 빛을 반사하여 포식자에게 자신의 모습을 인식시키지 않도록 한다. 가시볏과 어류의 일부는 야간에는 반사 효율을 저하시켜 생물 발광의 반사에 의한 발견의 위험성을 줄일 수 있다.[100]
수심 600m 부근부터 심해어의 체색은 은백색에서 납색으로 급격히 변화하며, 1,000m의 점심층에 이르면 거의 균일하게 암색이 된다.[101] 꼼치과의 대부분은 선명한 붉은 체색을 하고 있지만, 푸른 파장의 빛만 도달하는 심해에서는 흑색과 마찬가지로 거의 눈에 띄지 않는다고 생각된다.
=== 유인 돌기 및 미끼 모양체 ===
아귀목 어류의 등지느러미 첫 번째 가시는 낚싯대 모양으로 변형되어 있으며, '''유인 돌기'''(illicium)라고 한다. 유인 돌기 끝에 있는 술 모양으로 부풀어 오른 부분을 '''미끼 모양체'''(esca)라고 부르며, 아귀류는 미끼 모양체를 먹이처럼 움직여 먹잇감을 유인한다. 심해를 유영하는 초롱아귀상과 어류의 경우, 미끼 모양체가 공생 발광 기관으로 기능하는 경우가 많다.[102] 또한, 악마아귀과 심해어는 복잡하게 갈라진 턱수염과 같은 구조를 가지고 있으며, 역시 먹이 유인에 사용되는 것으로 생각된다.[102]

5. 1. 골격 및 근육
어류는 몸 전체에 포함된 단백질과 골격의 비중이 일반적으로 해수보다 크다.[87] 천해어는 유영에 따른 양력이나 부레에 공기를 들이마심으로써 부력을 얻지만, 이용 가능한 에너지가 부족한 심해에서는 심해어는 가능한 한 유영하지 않고 부력을 확보해야 한다.[87] 많은 심해어는 뼈, 연골 및 근육 등 체내의 고밀도 조직이 감소하고, 대신 저비중의 수분과 지방을 다량 함유하고 있다.[87] 배지느러미와 그 지지 골격, 또는 머리의 골격을 퇴화시킨 것과 비늘, 가시를 가지지 않은 종류도 많으며, 경량화를 위한 하나의 수단으로 여겨진다.[57]5. 2. 부레
부레(鰾)는 천해어가 부력을 얻는 일반적인 수단이지만, 심해에서는 극도의 고수압 때문에 일반적인 가스 교환에 의한 부레의 기능을 기대할 수 없다. 고수압과 급격한 압력 변화에 견디기 위해 심해어에는 부레의 벽을 견고한 구아닌결정으로 덮거나, 내용물을 기체가 아닌 지방이나 왁스로 대체하는 등의 적응이 나타난다.[95][88]수심이 커짐에 따라 고수압에 역행하여 가스 교환(특히 분비)을 하는 데 대한 부담도 증가한다. 중심층 유영성 심해어(천해로의 이동을 하지 않는 그룹)에서는 부레는 일반적으로 퇴화되어 있으며, 더 수심이 깊은 점심층에서는 부레를 갖지 않는 종류가 많다.[95]
5. 3. 안구
심해어는 서식 환경에 따라 독특한 안구 구조를 가진다. 수심 약 1,000m까지는 미약한 태양광이 도달하므로, 이 영역에 서식하는 심해어는 몸에 비해 매우 큰 안구를 가지거나, 데메에소과, 무네에소과, 요코에소과 어류 등 최소 11개 과의 심해어는 관 모양으로 변형된 '''관상안'''을 가진다.[89] 심해에 도달하는 빛은 산란과 굴절 때문에 태양의 위치와 관계없이 항상 바로 위에서 내리쬐며, 해가 진 후에도 광량 변화가 적다.[90] 보우엔기요과 등 일부 예외를 제외하고, 대부분의 관상안은 해수면 방향의 빛에 대응하여 바로 위를 향하고 있다.반면, 동굴 생물은 암흑 조건하에서 눈이 퇴화한 예가 많지만, 심해어는 약간의 빛이 들어오고, 종에 따라 천해로 이동하거나 발광 생물이 많다는 점이 영향을 미치는 것으로 보인다.
1,000m 이하의 심해층은 빛이 전혀 없는 암흑 세계이며, 이 영역에는 오목하게 들어간 작은 눈을 가진 심해어가 많다. 소코오쿠메우오과처럼 눈이 피부 속에 파묻히거나, 초우친하다카처럼 판상의 망막만 남은 심해어도 있지만, 빛 감지 기능은 여전히 남아 있어 퇴화가 아닌 특수화로 간주된다.[91] 심해층에서 드물게 깜빡이는 생물발광을 포착하기 위해 가늘고 긴 작은 안구가 더 적합하다는 보고도 있다.[92] 이러한 눈은 일반적인 눈보다 공간 분해능이 뛰어나 20~30m 정도 떨어진 곳의 발광을 포착하는 데 적합하며, 유영 능력이 낮은 심해어에게 시야를 좁은 범위로 제한하는 것은 에너지 효율 측면에서 합리적이다.[93]
5. 4. 소화 기관
육식성 심해어는 체형에 비해 상당히 큰 입과 이빨을 가진 경우가 많다.[95] 특히 신기한 물고기목에 속하는 심해꼼치·흡혈귀물뱀장어류는 매우 큰 입으로 잘 알려져 있는데, 이들의 두개골은 작지만 극도로 발달한 아가미뼈가 큰 입을 지탱한다.[96] 흡혈귀물뱀장어 ''Eurypharynx pelecanoides''는 전 세계 온대 해역의 심해(550~3,000미터)에서 비교적 흔하게 발견된다.[94] 심해꼼치는 날카로운 이빨로 큰 먹이를 삼키지만, 흡혈귀물뱀장어는 아가미에 이빨이 거의 없어 작은 물고기나 플랑크톤을 모아 먹는다.[96]발달된 치열 또한 육식성 심해어의 특징인데, 귀신상어(귀신상어과)는 날카로운 송곳니 모양의 이빨을 가졌다.[95] 악어상어·등불상어류 일부는 안쪽으로 구부러진 이빨을 가져 먹이를 놓치지 않도록 한다.[95]
등불상어류와 큰입우럭과 어류 등은 식도나 위를 크게 확장시킬 수 있다.[97] 큰입우럭은 자신의 몇 배나 되는 먹이를 삼킬 수 있으며, 복부가 비정상적으로 부풀어 오른 상태로 포획되기도 한다. 멜라닌 등의 색소 침착으로 흑색화된 장관을 가진 심해어도 있는데, 이는 발광 생물을 포식했을 때 소화관을 투과한 빛이 천적을 유인하는 것을 막기 위함이다.[95]
심해 환경에서는 지속적인 포식이 어려워 에너지 축적이 중요하다. 간은 심해어에게 중요한 에너지 저장 기관이며, 긴꼬리꼼치과 등은 매우 크고 지방이 풍부한 간을 가지고 있다.[131] 요로이다라/Abyssal grenadier영어(솔치과)의 간 또한 지방·글리코겐을 풍부하게 함유하고 있으며, 약 180일 동안 먹이를 얻지 못해도 생명을 유지할 수 있다고 추정된다.[131]
지방은 바닷물보다 비중이 작아, 간에 다량의 지방분을 축적하는 것은 부력 확보에도 기여한다. 상어과·까치상어과(각상어목) 등 부레가 없는 심해성 연골어류 중에는 체중의 25%에 달하는 거대한 간을 가진 것도 있다.[88]
5. 5. 체색
심해에서는 비교적 밝은 중층에 사는 어류에서는 체표면의 '''은화'''에 의한 의태가 보인다.[98] 가시볏과 어류는 두께 수 밀리미터의 납작한 몸을 가지고 있으며, 표면은 알루미늄 호일과 같은 광택이 있는 은색을 띤다. 그들의 체표면에는 과닌에 의한 미세한 반사성 결정이 여러 층에 걸쳐 규칙적으로 나란히 배열되어 있으며,[99] 거울처럼 빛을 반사하여 포식자에게 자신의 모습을 인식시키지 않도록 한다. 가시볏과 어류의 일부는 야간에는 반사 효율을 저하시켜 생물 발광의 반사에 의한 발견의 위험성을 줄일 수 있다.[100]수심 600m 부근부터 심해어의 체색은 은백색에서 납색으로 급격히 변화하며, 1,000m의 점심층에 이르면 거의 균일하게 암색이 된다.[101] 꼼치과의 대부분은 선명한 붉은 체색을 하고 있지만, 푸른 파장의 빛만 도달하는 심해에서는 흑색과 마찬가지로 거의 눈에 띄지 않는다고 생각된다.
5. 6. 유인 돌기 및 미끼 모양체
아귀목 어류의 등지느러미 첫 번째 가시는 낚싯대 모양으로 변형되어 있으며, '''유인 돌기'''(illicium)라고 한다. 유인 돌기 끝에 있는 술 모양으로 부풀어 오른 부분을 '''미끼 모양체'''(esca)라고 부르며, 아귀류는 미끼 모양체를 먹이처럼 움직여 먹잇감을 유인한다. 심해를 유영하는 초롱아귀상과 어류의 경우, 미끼 모양체가 공생 발광 기관으로 기능하는 경우가 많다.[102] 또한, 악마아귀과 심해어는 복잡하게 갈라진 턱수염과 같은 구조를 가지고 있으며, 역시 먹이 유인에 사용되는 것으로 생각된다.[102]6. 발광
많은 심해어들은 생물발광을 하며, 어둠에 적응한 매우 큰 눈을 가지고 있다. 생물발광 생물은 루시페린 분자의 교반을 통해 생물학적으로 빛을 생성할 수 있으며, 이는 빛을 생성한다. 이 과정은 산소가 존재하는 경우에만 수행되어야 한다. 이러한 생물들은 중심해 지역과 그 이하(200m 이하)에서 흔하다. 심해어의 50% 이상과 일부 종의 새우와 오징어는 생물발광을 할 수 있다. 이러한 유기체들의 80%는 발광포를 가지고 있다-어두운 색으로 둘러쌓인 발광 박테리아를 포함하는 빛을 생성하는 선 세포. 이 발광포 중 일부는 인간의 눈과 마찬가지로 렌즈를 가지고 있어 빛의 발산을 확대하거나 줄일 수 있다.[163] 빛을 생산하는 능력은 유기체 에너지의 1%만 필요하며 많은 목적을 가지고 있다. 그 능력은 먹이를 찾고 끌어들이는 데에 사용된다. 아귀처럼; 순찰을 통해 영역을 주장하고; 의사소통하고 짝을 찾고, 탈출을 위해 포식자를 정신사납게 하거나 일시적으로 눈이 멀게하는데도 사용된다. 또한, 일부의 빛이 여전히 관통하는 중심해 지역에서는 몇몇 유기체들은 그림자가 드리워지지 않도록 위레서 빛의 색과 강도에 맞게 배를 비추어 아래에 있는 포식자들로부터 위장한다. 이 전술은 카운터 일루미네이션(counter illumination)으로 알려져 있다.[163]
심해어를 포함한 많은 심해 생물은 자신이 생산한 루시페린을 이용하는 자력 발광을 한다.[106] 일반적으로 발광기의 수는 많으며, 수백에서 수천에 달하는 경우도 있다.[107] 발광기 개구부에 렌즈나 필터 모양의 구조를 가진 경우도 있으며, 광량과 조사 방향, 발광색의 조절에 도움이 된다.
심해에 도달하는 빛은 녹색과 청색의 파장으로 제한되기 때문에, 많은 심해어의 눈은 푸른빛만 감지할 수 있도록 되어 있지만, 악어도마뱀물고기과에 속하는 횃불물고기아과 및 빗자루별눈가지아과 어류에는 예외적으로 붉은빛을 인식할 수 있는 종류가 있다.[108] 이들은 발광기를 덮는 특수한 필터를 사용하여 자신이 붉은빛을 발하는 것도 가능하다.[109] 먹이 또는 다른 포식자에게 인식되지 않는 적색광을 사용하는 것은 포식과 번식을 하는 데 유리하게 작용할 것으로 생각된다.[110]
깊은 바다라고 해도 수심 1,000미터 정도까지는 아주 미약하게나마 빛이 들어온다. 따라서 낮에 바다 밑에서 해면을 올려다 볼 때, 상부에 있는 생물의 그림자가 드러난다. 복부에 발광기를 배치하여 쏟아지는 빛과 같은 수준으로 밝기를 조절하면 이 실루엣을 없앨 수 있다. 이것을 '''카운터 일루미네이션(counterillumination)'''이라고 하며, 중층성(中深層性) 심해어의 대부분이 이 방법을 이용하고 있다.[111]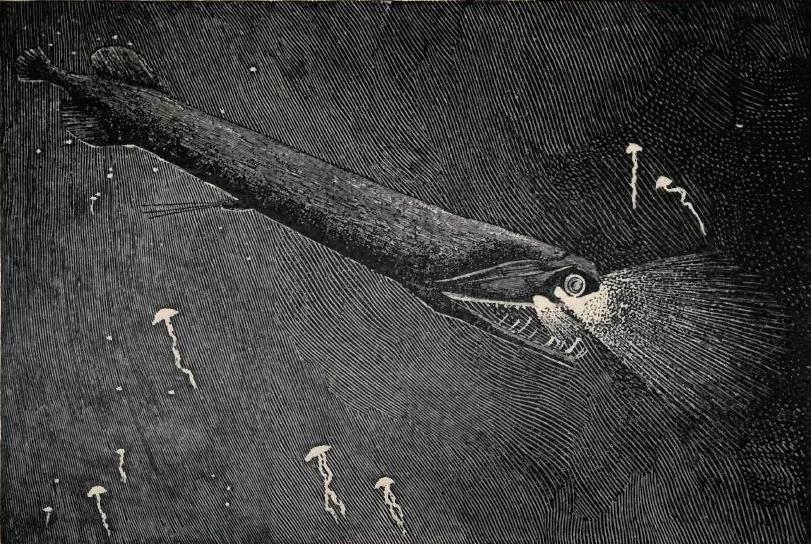
먹이가 되는 생물을 비추는 것이 생물발광의 포식에서 기본적인 용도이며, 많은 심해어가 이러한 종류의 발광을 한다.[110] 대부분의 악어상어과 어류는 눈 바로 아래 또는 뒤쪽에 대형 발광기를 가지고 있으며, 이것으로 시야를 밝게 비춘다. 등불아귀류는 미끼의 끝에서 공생 발광을 하여 먹이를 유인하며, 복잡하게 가지를 친 턱수염과 같은 자력 발광 기관을 가진 것도 있다. 또한, 가슴빛상어속이나 검은입상어속 등 몇몇 그룹은 입 안에 발광 기관을 가지고 있다.[110]
얕은 바다에 사는 물고기가 무리를 짓기 위해 모일 때는 시각 외에 반사광을 이용하고 있다.[112] 심해어도 이와 비슷하게 생물발광을 집단 유지에 사용하고 있다고 생각된다. 햇살멸치류는 복부의 발광기 외에, 머리와 꼬리에도 발광기를 가지고 있다. 발광기의 크기, 위치, 수는 암수에서 다른 성적이형을 보이며, 암수의 의사소통에 사용되고 있을 가능성이 있다.[112]
와니토카게기스과에 속하는 일부 심해어는 방어적인 강한 섬광을 발할 수 있다.[113] 매우 밝은 빛을 내뿜어 포식자를 기절시키는 경우도 있다.[114] 샛별고기과(니기스목) 어류는 아가미 아래에서 발광액을 분비하여, 포식자의 눈을 끄는 미끼 역할을 한다고 생각된다.[115]
6. 1. 발광의 기능
심해 저인망을 이용한 샘플링은 lantern fish가 모든 심해 어류 바이오매스의 65%를 차지함을 보여준다.[184] 실제로, lantern fish는 모든 척추동물 중에서 가장 널리 분포하며, 다양하며, 유기체의 먹이로 중요한 생태학적 역할을 한다. 남극해에서 Myctophids는 킹펭귄과 같은 포식자를 위한 크릴의 대체 식량 자원을 제공한다.[184]깊은 바다라고 해도 수심 1,000미터 정도까지는 아주 미약하게나마 빛이 들어온다. 따라서 낮에 바다 밑에서 해면을 올려다 볼 때, 상부에 있는 생물의 그림자가 드러난다. 복부에 발광기를 배치하여 쏟아지는 빛과 같은 수준으로 밝기를 조절하면 이 실루엣을 없앨 수 있다. 이것을 '''카운터 일루미네이션(counterillumination)'''이라고 하며, 중층성(中深層性) 심해어의 대부분이 이 방법을 이용하고 있다.[111]
먹이가 되는 생물을 비추는 것이 생물발광의 포식에서 기본적인 용도이며, 많은 심해어가 이러한 종류의 발광을 한다.[110] 대부분의 악어상어과 어류는 눈 바로 아래 또는 뒤쪽에 대형 발광기를 가지고 있으며, 이것으로 시야를 밝게 비춘다. 등불아귀류는 미끼의 끝에서 공생 발광을 하여 먹이를 유인하며, 복잡하게 가지를 친 턱수염과 같은 자력 발광 기관을 가진 것도 있다. 또한, 가슴빛상어속이나 검은입상어속 등 몇몇 그룹은 입 안에 발광 기관을 가지고 있다.[110]
얕은 바다에 사는 물고기가 무리를 짓기 위해 모일 때는 시각 외에 반사광을 이용하고 있다.[112] 심해어도 이와 비슷하게 생물발광을 집단 유지에 사용하고 있다고 생각된다. 햇살멸치류는 복부의 발광기 외에, 머리와 꼬리에도 발광기를 가지고 있다. 발광기의 크기, 위치, 수는 암수에서 다른 성적이형을 보이며, 암수의 의사소통에 사용되고 있을 가능성이 있다.[112]
ワニトカゲギス과에 속하는 일부 심해어는 방어적인 강한 섬광을 발할 수 있다.[113] 매우 밝은 빛을 내뿜어 포식자를 기절시키는 경우도 있다.[114] 샛별고기과(니기스목) 어류는 아가미 아래에서 발광액을 분비하여, 포식자의 눈을 끄는 미끼 역할을 한다고 생각된다.[115]
7. 생태
심해어의 생태에는 혹독한 심해 환경에 적응한 독특한 방식이 많이 나타난다. 심해의 생물 밀도는 천해와 비교하여 극단적으로 낮아, 개체를 유지하기 위한 포식 행동을 효율적으로 수행하는 것과 동시에 불필요한 에너지 소비를 최소한으로 억제하는 것이 요구된다. 또한, 서식 범위와 개체 수의 문제로 심해에서는 수컷과 암컷의 번식 기회가 매우 적다. 이러한 조건 하에서 확실하게 번식·성장을 위한 전략을 세우는 것은 종으로서의 생존을 도모하기 위해 필수적인 적응이다.
심해 어류는 이 지역에서 살아남기 위해서 다양한 적응법들을 진화시켜 왔다. 이런 물고기들 중 다수는 자연광이 없는 지역에 살고 있기 때문에, 먹이와 짝을 찾고 포식자를 피하기 위해 그들의 시력에만 의존할 수는 없다; 심해어는 그들이 살고 있는 극한으로 빛이 부족한 지역에 적절하게 진화해 왔다. 이 유기체의 다수는 눈이 멀었고 그들의 먹이를 잡고 잡히는 것은 피하기 위해 지역의 압력과 냄새의 변화에 대한 민감성과 같은 다른 감각에 다른 감각에 의존한다. 눈이 멀지 않은 개체들은 생물발광성 빛을 사용할 수 있는 크고 민감한 눈을 가지고 있다. 이 눈은 인간의 눈보다 빛에 100배 더 민감할 수 있다. 로돕신은 동물들이 희미한 빛으로 볼 수 있도록 도와주는 눈의 막대 세포에서 발견되는 단백질이다. 대부분의 척추동물이 보통 하나의 Rh1 옵신 유전자를 가지고 있는 반면에, 일부 심해어는 여러 Rh1 유전자를 가지고 있으며, 한 종인 은색 스피니핀(silver spinyfin)은 38개를 가지고 있다. Rh1 유전자의 증식은 심해어가 바다 깊은 곳에서 보는 데 도움이 될 수 있다. 또한, 잡아먹히는 것을 피하기 위해 많은 종들이 그들의 환경과 섞일 정도로 어둡다.[162]
많은 심해어들은 생물발광성이며, 매우 큰 눈은 어둠에 적응한다. 생발광 유기체는 루시페린 분자의 교반을 통해 생물학적으로 빛을 생산할 수 있으며 그래서 빛을 낸다. 이 과정은 산소가 있을 때 이루어져야 한다. 이런 유기체들은 중심해 지역과 그 이하(200m 이하)에서 흔하다. 심해어의 50% 이상과 일부 종의 새우와 오징어는 생물발광을 할 수 있다. 이런 유기체들의 80%는 발광포를 가지고 있다-어두운 색으로 둘러쌓인 발광 박테리아를 포함하는 빛을 생성하는 선 세포. 이 발광포 중 일부는 인간의 눈과 마찬가지로 렌즈를 가지고 있어 빛의 발산을 확대하거나 줄일 수 있다. 빛을 생산하는 능력은 유기체 에너지의 1%만 필요하며 많은 목적을 가지고 있다. 그 능력은 먹이를 찾고 끌어들이는 데에 사용된다. 아귀처럼; 순찰을 통해 영역을 주장하고; 의사소통하고 짝을 찾고, 탈출을 위해 포식자를 정신사납게 하거나 일시적으로 눈이 멀게하는데도 사용된다. 또한, 일부의 빛이 여전히 관통하는 중심해 지역에서는 몇몇 유기체들은 그림자가 드리워지지 않도록 위레서 빛의 색과 강도에 맞게 배를 비추어 아래에 있는 포식자들로부터 위장한다. 이 전술은 카운터 일루미네이션(counter illumination)으로 알려져 있다.[163]
심해어의 수명 주기는 전적으로 깊은 물이 될 수 있지만 일부 종은 얕은 물에서 태어나 성숙하면 밑으로 가라앉는다. 알과 유충이 있는 깊이와 관계 없이 그들은 일반적으로 표영성이다. 이 떠다니는 부유 생물의 생활방식은 중성 부력을 필요로 한다. 이 부력을 유지하기 위해서 알과 유충은 종종 그들의 혈장 내에 기름 방울을 함유하고 있다.[164] 이 유기체들이 완전 성숙 상태에 이를때 그들은 물기둥에서 위치를 유지하기 위해서 다른 적응방식이 필요하다. 일반적으로, 물의 밀도는 유기체를 뜨게 하는 부력의 측면인 밀어올리는 힘(upthrust)을 일으킨다. 이에 대응하기 위해 유기체의 밀도는 주변 물의 밀도보다 커야 한다. 대부분의 동물 조직들은 물보다 밀도가 크기 때문에 떠다니기 위해서는 평형 상태를 찾아야 한다. 많은 유기체는 떠있는 상태로 머물기 위해 부레를 개발하지만, 심해어들은 그들이 살고있는 환경의 고압때문에 보통 이 기관을 가지고 있지 않다. 대신에 심해어들은 유체역학적인 움직임을 주기 위해서 수중익선과 유사한 구조를 보여준다. 또한 물고기가 더 깊은 곳에 살수록 살은 더 젤리처럼, 뼈의 구조는 더 최소화된다는 것이 밝혀졌다. 그들은 높은 지방 함량, 크기, 두께 및 미네랄 함량의 감소를 통해 이루어지는 골격 중량 감소를 통해 조직 밀도를 감소시키고 해수면의 물고기보다 느리고 덜 민첩하게 만든다.
심해 환경에 도달하는 광합성 빛의 낮은 수준 때문에, 대부분의 물고기들은 더 높은 층에서 가라앉는 유기 물질에 의존해야 하며, 드물게는 영양분을 얻기 위해 열수 분출공에 의존해야 한다. 이 점은 깊은 바다가 얕은 지역보다 생산성이 훨씬 떨어지게 만든다. 또한, 표양성 환경에 있는 생물들은 드물고 먹을거리가 자주 오지 않는다. 이것 때문에, 유기체들은 그들이 생존할 수 있도록 하는 적응방식이 필요하다. 어떤 것들은 먹이를 찾거나 깊은 바다의 칠흑 같은 어둠 속에서 짝을 유혹하기 위해 긴 더듬이를 가지고 있다. 특히 심해어류는 얼굴에서 긴 낚싯대 모양의 각색이 튀어나와 있고, 그 끝에는 먹이를 유인하기 위해 지렁이처럼 꿈틀거리는 생물발광성 피부 조각이 있다. 어떤 물고기는 그들과 같은 크기나 큰 다른 물고기를 먹어야 하고, 그들은 그것들을 효율적으로 소화시키는 것을 돕기 위해 적응이 필요하다. 크고 날카로운 이빨, 경첩이 달린 턱, 불균형적으로 큰 입, 그리고 확장 가능한 몸은 심해어들이 이 목적을 위해 가지고 있는 특징들 중 몇 가지이다. 굴퍼 장어(gulper eel)는 이러한 특성을 나타내는 유기체의 한 예이다.
각기 다른 원양과 깊은 저서성 구역의 물고기는 물리적으로 구조화되어 있으며 서로 현저하게 다른 방식으로 행동한다. 각 구역 내의 공존하는 종의 집단은 수직으로 이동하는 작은 중층해양성 플랑크톤을 먹는 동물, 점심해의 아귀들, 깊은 바다의 저서성 경골어와 같이 비슷한 방식으로 작동하는 것으로 보인다.[165]
중층원양대 밑은 빛이 들지 않아 어둡다. 중층원양대 아래로 1,000m부터 심해저까지 뻗어있는 구간을 점심해수층이라고 하며, 수심이 깊은 4,000m 이하의 골짜기 지대는 심해원양대라고 부른다. 심해원양대의 기온은 1℃에서 4℃ 사이이며 점심해수층과 마찬가지로 빛이 전혀 들지 않는다. 점심해수층은 어둡고 영양염류와 용존 산소량이 적으며, 압력은 높고 온도가 낮은 극한 환경이다.[167]
점심해수층 어류는 이러한 환경을 이겨내기 위해 특별한 적응을 한다. 이들은 신진대사가 느리고 먹이로 삼을 수 있는 것이면 무엇이든 기꺼이 먹는다. 음식을 찾기 위해 돌아다니며 에너지를 낭비하기보다는 앉아서 기다리는 것을 선호하는 편이다. 점심해수층 어류의 행동은 중층원양대 어류의 행동과 대조된다. 중층원양대 어류는 이동성이 높아 활발하지만, 점심해수층 어류는 거의 모두가 제자리에 머물러있는 포식자로, 움직이는 데 에너지를 거의 소비하지 않는다.[173]
점심해수층 어류들은 주로 작은 강모 입을 가지며, 몸길이는 약 10cm 정도로 보통 25cm를 넘지 않는다. 몸은 가늘고 약하며 물기가 많은 근육과 골격 구조로 되어 있다. 물고기의 많은 부분이 물로 이루어져 있기 때문에, 점심해수층의 압력에 큰 영향을 받지 않으며, 비늘이 없고 끈적한 몸을 가진다. 이들은 먹이가 눈앞에 나타나거나 인광체에 의해 유인될 때까지 물기둥에 머무르며 기다린다. 점심해수층 어류들은 찌꺼기나 배설물, 때로는 무척추동물이나 중층원양대 어류를 먹이로 하나 이들에게 얻을 수 있는 에너지는 적다.[173] 표해수층에서 유래된 먹이의 약 20%는 중층원양대로 공급되며,[174] 5%만이 점심해수층으로 전달된다.[175]
7. 1. 식성
낮에는 심해에 사는 물고기가 밤에 먹이를 찾아 천해로 이동하는 것을 일주 수직 이동(英: diel vertical migration)이라고 하며, 중심층 유영성 심해어에 많이 나타나는 특징이다.[116] 심해어에 국한되지 않고, 요각류와 갑각류와 같은 동물 플랑크톤, 벚꽃새우 등 많은 심해 생물이 일주 수직 이동을 한다. 일주 수직 이동을 하는 심해어는 비교적 발달된 부레를 가지고 있으며, 일부 종에서는 수압 변화에 대응하기 위해 공기 대신 지방을 축적하는 등의 적응이 보인다.[95]
주로 중심층에 서식하는 햇빛멸치류는 일주 수직 이동을 하는 심해어의 대표적인 존재이며, 수심 1,000미터까지 분포하는 많은 종류가 밤에는 해면으로 이동한다. 심해에서의 서식 범위와 상승하여 먹이를 찾는 수심은 종류에 따라 다르며, 서로 경쟁하지 않도록 분포를 나누고 있다. 이러한 분포는 “수직 이동의 사다리 (ladder of migrations)”라고도 불리며, 천해의 유기물을 신속하게 심해로 운반하는 중요한 메커니즘으로 작용한다.[142]
7. 1. 1. 일주 수직 이동
표해수대 아래에서는 해수의 상태가 빠르게 변한다. 200미터에서 1000미터 사이에서는 빛이 거의 사라질 때까지 계속 희미해진다. 온도는 수온약층을 통과하며 7.8°C~3.9°C 사이로 떨어진다. 이 지점을 약광층[166] 또는 중층원양대라고 부른다. 이 구간에서 압력은 10m마다 1기압씩 계속 증가하며, 영양염류의 양은 용존산소량, 물의 순환 속도와 함께 감소한다.[167] 제2차 세계 대전 동안 새롭게 개발된 sonar를 사용한 이들은 낮 시간대에 보이는 300~500m 깊이의 가짜 해저[168]를 발견하고 혼란스러워했다. 이는 수중 음파 탐지기를 반사하는 부레를 가진 수백만 마리의 해양 생물들, 특히 중층원양대 어류들이 플랑크톤을 먹기 위해 해 질 녘에 얕은 물 위로 이동했기 때문에 나타난 현상이다. 이 층은 달이 떠 있을 때 더 깊어지고, 구름이 달을 가리며 지나갈 때 더 얕아질 수 있으며, 이 층을 심해 산란층이라고 부른다.[169]대부분의 중층원양대 어류는 매일 수직 이동을 한다. 밤에는 동물성 플랑크톤을 따라 표해수층으로 이동하며, 낮에는 안전을 위해 깊은 곳으로 돌아간다.[167][170] 이러한 수직 이동은 종종 큰 수직 거리에서 발생하며, 부레의 도움을 받아 이루어진다. 물고기가 위로 올라가기를 원할 때, 부레는 부풀어 오른다. 중층원양대의 높은 압력으로 인해, 이 과정은 상당한 에너지가 필요하다. 물고기가 상승할 때, 부레가 터지는 것을 막기 위해서는 부레 내부의 압력이 조절되어야 한다. 물고기가 깊은 곳으로 돌아가고 싶을 때, 부레의 바람은 빠진다.[171] 일부 중층원양대 어류는 온도가 10°C에서 20°C 사이로 변하는 수온약층을 통해 매일 이동하므로 온도변화에 잘 견딘다.[172]
중층원양대 어류들은 근육질의 몸, 골화된 뼈, 비늘, 잘 발달된 아가미와 중추신경계, 그리고 큰 심장과 신장을 가지고 있다. 중층원양대의 플랑크톤을 먹이로 하는 생물들은 작은 입과 미세한 아가미 돌기를 가지고 있지만, 육식성 어류들은 더 큰 입과 더 거친 아가미 돌기를 가지고 있다.[167]
낮에는 심해에 사는 물고기가 밤에 먹이를 찾아 천해로 이동하는 것을 일주 수직 이동(英: diel vertical migration)이라고 하며, 중심층 유영성 심해어에 많이 나타나는 특징이다.[116] 심해어에 국한되지 않고, 요각류와 갑각류와 같은 동물 플랑크톤, 벚꽃새우 등 많은 심해 생물이 일주 수직 이동을 한다. 일주 수직 이동을 하는 심해어는 비교적 발달된 부레를 가지고 있으며, 일부 종에서는 수압 변화에 대응하기 위해 공기 대신 지방을 축적하는 등의 적응이 보인다.[95]
주로 중심층에 서식하는 햇빛멸치류는 일주 수직 이동을 하는 심해어의 대표적인 존재이며, 수심 1,000미터까지 분포하는 많은 종류가 밤에는 해면으로 이동한다. 심해에서의 서식 범위와 상승하여 먹이를 찾는 수심은 종류에 따라 다르며, 서로 경쟁하지 않도록 분포를 나누고 있다. 이러한 분포는 “수직 이동의 사다리 (ladder of migrations)”라고도 불리며, 천해의 유기물을 신속하게 심해로 운반하는 중요한 메커니즘으로 작용한다.[142]
7. 2. 번식 행동
번식 행동의 적응은 활발하게 헤엄치는 저서성 어류인 꼼치류보다, 초우친하다카/チョウチンハダカ일본어와 같은 매복형 저서성 심해어 및 중층을 유영하는 심해어에서 현저하게 나타난다.[118] 대구목이나 솔치과 등 일부 심해어의 번식 활동에는 명확한 계절성이 있으며, 이들 종은 연안의 생물 생산이 왕성한 시기에 맞춰 산란한다.[118] 일반적으로 유영성 심해어는 작고 많은 알을 낳는 경향이 있으며, 저서성 어류의 알은 크지만 수가 적은 경향이 있다.자웅동체라면, 두 마리가 만나기만 하면 번식이 가능해진다. 두 성의 생식선을 유지해야 하므로 에너지 부담은 커지지만, 개체 밀도가 낮은 심해어에게는 큰 이점이 된다. 히메목에 속하는 솔치과·아귀과·등불해파리과·심해솔치과의 심해어는 모두 자웅동체이다.[119] 같은 히메목의 청새치과에서는, 심해성 종은 자웅동체인 반면, 천해종은 암수가 나뉜다.[119]
성전환을 하는 어류는 천해어에서도 알려져 있지만, 심해어에도 유사한 번식 방식이 있다. 천해어에서는 암컷에서 수컷으로 성전환하는 암컷선숙이 많은 반면, 심해어에서는 망둑어속이나 가시고기속 등, 수컷에서 암컷으로 성전환하는 수컷선숙이 자주 관찰된다. 주로 중심층에 서식하는 가시고기속 어류는 생후 1년차까지는 모두 수컷이지만, 대략 2년차까지는 암컷으로 성전환한다.
이러한 수컷선숙은 천해어에서는 말미잘 등에서 볼 수 있다. 수컷이 세력권이나 하렘을 형성하는 어종에서는 수컷이 대형화하는 암컷선숙이 유리하지만, 개체군 밀도가 매우 작은 심해에서는 이러한 행동 양식을 취하기 어렵다.[120] 일반적으로 정자보다 난자를 만드는 데 더 많은 에너지가 필요하므로(암컷의 성숙이 더 늦다), 심해어에게는 수컷선숙에 의한 번식이 유리하다고 생각된다.
왜웅(矮雄)이란 암컷에 비해 극도로 작은 수컷을 말하며, 특히 초롱아귀상과에서 많이 볼 수 있다.[121] 초롱아귀류의 수컷은 암컷의 1/3에서 1/13 정도 크기로밖에 자라지 않는다.[122] 미츠쿠리에나가초우친안코우과·온안코우과 등 최소 4과의 왜웅은 암컷에 기생하는 습성을 가지고 있으며, 처음에는 자유생활을 하는 수컷은 암컷을 찾으면 복부에 달라붙어 일체화한다. 복부가 가장 흔하지만, 등이나 머리, 또는 유인 돌기에 부착된 사례도 알려져 있다.[123] 수컷은 그 후 암컷의 피부에서 뻗어나온 혈관을 통해 영양을 얻게 된다. 스스로 헤엄칠 필요가 없기 때문에 수컷의 눈과 지느러미는 점차 퇴화하는 반면, 생식에 필요한 정소 기능은 유지된다. 암컷과 왜웅의 결합이 서로의 성 성숙을 달성하기 위한 필요조건이 되는 경우도 있다.
왜웅을 가진 다른 심해어로는, 미츠마타야리우오속과 오니하다카속의 일부(와니토카게기스목), 그리고 쿠지라우오과의 동료가 알려져 있으며, 어느 쪽도 암컷에 기생하지 않는다. 미츠마타야리우오( ''Idiacanthus antrostomus'')는 약 50cm까지 자라는 암컷에 비해 수컷은 약 5cm밖에 되지 않으며,[124] 이빨과 소화기관이 빈약하여 스스로 먹이를 잡는 것은 거의 불가능하다. 눈 아래 발광기와 정소는 발달되어 있으므로, 평상시에는 에너지 소비를 억제하고 부유하고 있으며, 발광으로 암컷을 불러들이는 것으로 생각된다.
이와 같이 암컷이 아닌 수컷이 소형화되는 것은, 위에서 언급한 성전환의 경우와 마찬가지로, 번식에는 암컷이 훨씬 많은 에너지를 필요로 하기 때문이다. 왜웅은 암컷을 찾아 비교적 먼 거리를 유영할 필요가 있기 때문에, 지구력이 높은 소위 적신의 근섬유가 발달되어 있다.[125] 또한, 대부분의 왜웅은 암컷보다 발달된 고정밀도의 후각과, 미량의 빛을 예민하게 감지하는 시각을 가지고 있으며, 암컷의 위치를 특정하는 데 활용하고 있다.[126][125]
7. 2. 1. 자웅동체 및 성전환
자웅동체라면, 두 마리가 만나기만 하면 번식이 가능해진다. 두 성의 생식선을 유지해야 하므로 에너지 부담은 커지지만, 개체 밀도가 낮은 심해어에게는 큰 이점이 된다. 히메목에 속하는 솔치과·아귀과·등불해파리과·심해솔치과의 심해어는 모두 자웅동체이다.[119] 같은 히메목의 청새치과에서는, 심해성 종은 자웅동체인 반면, 천해종은 암수가 나뉜다.[119]성전환을 하는 어류는 천해어에서도 알려져 있지만, 심해어에도 유사한 번식 방식이 있다. 천해어에서는 암컷에서 수컷으로 성전환하는 암컷선숙이 많은 반면, 심해어에서는 망둑어속이나 가시고기속 등, 수컷에서 암컷으로 성전환하는 수컷선숙이 자주 관찰된다. 주로 중심층에 서식하는 가시고기속 어류는 생후 1년차까지는 모두 수컷이지만, 대략 2년차까지는 암컷으로 성전환한다.
이러한 수컷선숙은 천해어에서는 말미잘 등에서 볼 수 있다. 수컷이 세력권이나 하렘을 형성하는 어종에서는 수컷이 대형화하는 암컷선숙이 유리하지만, 개체군 밀도가 매우 작은 심해에서는 이러한 행동 양식을 취하기 어렵다.[120] 일반적으로 정자보다 난자를 만드는 데 더 많은 에너지가 필요하므로(암컷의 성숙이 더 늦다), 심해어에게는 수컷선숙에 의한 번식이 유리하다고 생각된다.
7. 2. 2. 왜웅
왜웅(矮雄)이란 암컷에 비해 극도로 작은 수컷을 말하며, 특히 초롱아귀상과에서 많이 볼 수 있다.[121] 초롱아귀류의 수컷은 암컷의 1/3에서 1/13 정도 크기로밖에 자라지 않는다.[122] 미츠쿠리에나가초우친안코우과·온안코우과 등 최소 4과의 왜웅은 암컷에 기생하는 습성을 가지고 있으며, 처음에는 자유생활을 하는 수컷은 암컷을 찾으면 복부에 달라붙어 일체화한다. 복부가 가장 흔하지만, 등이나 머리, 또는 유인 돌기에 부착된 사례도 알려져 있다.[123] 수컷은 그 후 암컷의 피부에서 뻗어나온 혈관을 통해 영양을 얻게 된다. 스스로 헤엄칠 필요가 없기 때문에 수컷의 눈과 지느러미는 점차 퇴화하는 반면, 생식에 필요한 정소 기능은 유지된다. 암컷과 왜웅의 결합이 서로의 성 성숙을 달성하기 위한 필요조건이 되는 경우도 있다.
왜웅을 가진 다른 심해어로는, 미츠마타야리우오속과 오니하다카속의 일부(와니토카게기스목), 그리고 쿠지라우오과의 동료가 알려져 있으며, 어느 쪽도 암컷에 기생하지 않는다. 미츠마타야리우오( ''Idiacanthus antrostomus'')는 약 50cm까지 자라는 암컷에 비해 수컷은 약 5cm밖에 되지 않으며,[124] 이빨과 소화기관이 빈약하여 스스로 먹이를 잡는 것은 거의 불가능하다. 눈 아래 발광기와 정소는 발달되어 있으므로, 평상시에는 에너지 소비를 억제하고 부유하고 있으며, 발광으로 암컷을 불러들이는 것으로 생각된다.
이와 같이 암컷이 아닌 수컷이 소형화되는 것은, 위에서 언급한 성전환의 경우와 마찬가지로, 번식에는 암컷이 훨씬 많은 에너지를 필요로 하기 때문이다. 왜웅은 암컷을 찾아 비교적 먼 거리를 유영할 필요가 있기 때문에, 지구력이 높은 소위 적신의 근섬유가 발달되어 있다.[125] 또한, 대부분의 왜웅은 암컷보다 발달된 고정밀도의 후각과, 미량의 빛을 예민하게 감지하는 시각을 가지고 있으며, 암컷의 위치를 특정하는 데 활용하고 있다.[126][125]
7. 3. 성장
생물은 성숙하는 데 많은 에너지를 필요로 한다. 심해에서는 충분한 먹이를 얻기 어렵기 때문에, 심해어는 얕은 바다에서 유생 시기를 보내는 경우가 많다.[127] 명태처럼 얕은 바다에서 산란하는 것과, 긴꼬리치어처럼 깊은 바다에서 산란하고 자연스럽게 부상하도록 하는 것이 있다.표층에서 성장하는 심해어의 자치어는, 포식자에게 발견되기 어려운 투명한 몸을 가지는 등, 성체와는 전혀 다른 특이한 형태를 취하는 경우가 많다.[127] 긴꼬리치어나 세줄아귀과의 자치어는, 지느러미나 눈 등 신체 일부를 가늘고 긴 돌기처럼 늘이고 있다. 또, 톱상어과・멸치과의 일부 자치어는 장관을 체외로 노출시켜, 체장의 수 배에 달하는 긴 장을 매달고 유영한다.[128] 이와 같이 신체 일부를 늘인 형태는 부유 생활에 대한 적응으로 보이며, 체표면적을 확대하여 부력을 높이는 효과를 가진다. 또, 노출된 소화관은 장의 표면적을 넓혀 큰 먹이를 소화 흡수할 수 있도록 하는 등의 의미가 있다고 여겨진다.
얕은 바다에서 성장한 심해어는 변태를 거쳐 성어와 거의 같은 모습의 치어가 되어, 원래의 서식지인 심해로 이동한다. 한랭하고 먹이가 적은 환경에서 생활하는 심해어의 성장 속도는 느리고, 특히 저서어는 수명도 긴 것으로 생각된다.[129] 심해어의 나이는 다른 어류와 마찬가지로, 이석이나 비늘에 새겨진 동심원상의 무늬로 추정할 수 있다. 그러나, 성장 주기의 계절적 변화가 적은 심해어의 이석에 명확한 나이테가 형성되는 것은 드물고, 나이 추정은 매우 미세한 일주륜으로 이루어진다.[129]
남방심해꼼치(심해꼼치과)나 세키토리이와시과의 일종 (''Conocara macropterum'')은 소형의 치어와 대형의 성어만이 현저하게 많고, 양자의 중간에 해당하는 크기가 매우 적다는 이봉성의 체장 분포를 보인다.[118] 이들 어류는 치어기에 어떤 원인에 의한 선택적 포식을 받고, 이 시기를 극복한 것만이 급속한 성장을 이루는 것으로 보인다.
7. 4. 유영 행동
심해어는 에너지 소비를 억제하기 위해 활발하게 움직이지 않는 종류가 많다. 중심층에 분포하는 햇살치나 쥐치류는 평상시 떠 있는 듯한 자세로 가만히 있는데,[130] 이는 자신의 그림자를 최대한 작게 하여 포식자에게 발견되기 어렵게 하는 효과가 있다.중심층에 많은 악어장어류, 점심층에 널리 분포하는 낚시벌레류는 유영성 매복형(float-and-wait) 포식어의 대표이다. 낚시벌레과 등을 제외하고 둥글둥글한 구형의 체형을 하고 있으며, 부력 유지에는 적합하지만 빠른 유영에는 적합하지 않다. 이들의 근육은 흰살이며, 순발력에는 뛰어나지만 지구력은 거의 없다. 은어과·검은입술장어과 등은 수렵 채집형(active foraging)의 유영성 심해어이다.
저서성 심해어에는 긴꼬리에소(낚시벌레과), 빨강아귀(빨강아귀과), 노르웨이가자미(가자미과)와 같이 해저에 정지하여 먹이를 기다리는 것(benthic fish)과, 속갈치·꼼치·장어·뱀장어·상어류 등 활발하게 헤엄치며 먹이를 찾는 것(benthopelagic fish)이 있다.[84]
매복형(sit-and-wait)의 저서어는 근육 조직이 발달한 체격을 가지며, 부레가 없는 경우가 많다.[131] 육식성 종류는 긴 이빨이 늘어선 큰 입과 눈을 갖추는 한편, 플랑크톤 식성 어류의 눈은 퇴화하는 경우가 많다. 모래 해저에서 배지느러미와 가슴지느러미를 사용하여 몸을 지탱하고, 가까이 온 먹이를 잡아 삼킨다. 심해에소 등 체비중이 큰 저서성 심해어는 해저에서 50cm 이상 떨어지는 것은 드물다.[131] 대형 먹이를 포식하는 매복형 심해어는 큰 눈을 효율적으로 이용할 수 있는 대륙 사면 상부에서 중부에 걸쳐 분포한다.
수렵 채집형은 저서성 심해어로서 일반적인 행동 양식이며, 속갈치과·꼼치과 등은 모든 수심에서 관찰된다.[131] 이들은 잘 발달된 부레와 기저가 긴 등지느러미·뒷지느러미를 가지고 있으며, 해저 바로 위를 호버링하듯이 헤엄칠 수 있다. 시각에 대한 의존도는 낮고, 먹이 탐색은 후각과 측선이 주로 이용된다.[131]
왕돌묵과 같은 심해어가 해안에 떠밀려 오거나 얕은 해역에서 어획·목격되면 지진의 전조로 여겨지기도 한다. 동해대학교 오리가라 요시아키 특임준교수 연구팀은 1992년부터 2011년 3월 11일까지의 심해어 목격 정보 101건과 규모 6 이상 지진 161건을 대조한 결과, 시기·장소의 일치는 적고 지진 예지에 의한 방재·감재에는 도움이 되지 않는다는 견해를 2017년에 발표했다.[132]
8. 심해어의 이용
심해어는 강과 같은 민물에서 살아가는 물고기, 생물, 육지에서 살아가는 생물, 얕은 표해수층에서 살아가는 물고기나 생물 등에 비해 대체적으로 사람이 식용으로 이용하기엔 부적합하지만 일부의 심해어는 사람의 식용으로 이용이 된다. 식용으로 이용되는 대표적인 심해어론 메로가 있으며 메로는 맛이 좋은 최고급 생선의 종류이다. 그외에 은대구나 돗돔도 심해어이지만 사람의 식용으로 이용이 된다. 또한 초롱아귀는 대체적으로 사람의 식용에 부적합하다는 편견도 있지만 일본에서는 심해어 초밥집에서 식용으로 사용이 된다.[47]
일상생활과는 거리가 먼 인상을 주는 심해어이지만, 식용으로 이용되는 종류는 많다.[133] 일본에서 식용으로 이용되는 앙코우(アンコウ), 타치우오(タチウオ), 무츠(ムツ), 긴메다이(キンメダイ), 스케토다라(スケトウダラ), 오히요(オヒョウ), 하타하타(ハタハタ) 등은 모두 수심 수백 미터의 심해에 서식한다. 어획 대상이 되는 유용 어종은 흉상어목(ツノザメ目), 가오리목(エイ目), 대구목(タラ目), 긴메다이목(キンメダイ目), 가자미목(カレイ目), 농어목(スズキ目) 등 대부분 저서성 심해어이다.[78] 해저 부근을 활발하게 유영하는 포식성 어류는 그 운동량을 뒷받침하기 위한 근육을 발달시키는 반면, 중층을 부유하는 심해어는 고압 하에서 부력을 확보하기 위해 몸 전체를 물처럼 만들거나 과도한 지방을 축적하는 경우가 많다. 미즈우오(ミズウオ)는 1미터를 넘는 중심층 유영성의 대형 심해어로, 삶으면 살이 녹아 없어진다는 데서 이름이 유래되었다.[134]
앞서 설명한 바와 같이, 유영성 심해어는 체내에 지질을 축적하는 경우가 많은데, 그 중에는 유지 성분으로 왁스를 포함하는 종류도 있다. 인체는 왁스를 소화할 수 없기 때문에, 이러한 어류의 고기를 다량 섭취하면 설사와 복통의 원인이 된다.[135] 왁스 성분이 특히 많은 クロタチカマス科의 바람치(バラムツ)와 아브라소코무츠(アブラソコムツ)는 일본에서는 식품위생법(食品衛生法)에 의해 판매가 금지되어 있다.[136] 햇살치류(ハダカイワシ類) 중에서도, 일주 수직 이동을 하지 않는 일부 종류는 체내에 왁스를 축적하고 있다.[140]
8. 1. 새로운 수산 자원
심해어는 강과 같은 민물에서 살아가는 물고기나 얕은 표해수층에 서식하는 생물에 비해 식용으로 부적합하지만, 메로, 은대구, 돗돔 등 일부는 식용으로 이용된다. 일본에서는 초밥 재료로 초롱아귀를 사용하기도 한다. 스페인 시장에서는 헤이크(hake)로 총칭되는 메르루사과의 심해어가 식용으로 많이 판매된다.
트롤어망을 이용한 저층 트롤 어업에서는 많은 양의 심해어가 함께 어획된다. 종래에는 시장 가치가 없어 폐기되던 심해어도 지역소비를 통해 식용화가 진행되고 있다. 스루가만 동북부 시즈오카현 누마즈시의 누마즈항이나 토다(헤다) 어항 등에서는 다양한 심해어가 어획되어, 누마즈항 심해수족관이 개설되거나 이즈시 등 서이즈 지역에서 심해어 요리를 홍보하는 등[137] 관광 자원으로 활용되기도 한다. 토다항의 음식점 주인에 따르면, 심해어는 기름기가 많고 폭신폭신한 식감을 가진 경우가 많다.[138]
1970년대 이후, 일본에서는 새로운 어업 자원으로 이용 가능한 심해어 탐색이 활발하게 진행되었다.[133] 수산청과 해양수산자원개발센터(현・수산종합연구센터)는 심해 어장 개발을 실시했고, 그 결과 메르루사(Merluza)나 마젤란아이나메 등 많은 유용 어종이 수입 식용어로 이용되게 되었다.[133] しんかい2000 등의 잠수 조사선은 키치지(キチジ)나 하타하타(ハタハタ) 등 심해성 수산 어종의 자원량 조사와 생태 해명에 기여하고 있다.[139]
저서성 심해어 외에도, 중층에 막대한 자원량을 가진 멸치과 어류는 세계적인 식량 수요를 충당할 에너지원으로 주목받고 있다.[140] 멸치과 어류는 지방 때문에 식용에 부적합하다는 인식이 있었지만, 일반적인 식용 어종과 유사한 지방질 조성을 가진다는 보고도 있다.[141] 중심층 유영성 심해어의 총 생물량은 적어도 9억 5천만 톤으로 추정되며, 이러한 미이용 자원을 활용하는 방법이 모색되고 있다.[142]
8. 2. 남획 문제
일부 심해어는 사람의 식용으로 이용되는데, 대표적으로 메로, 은대구, 돗돔 등이 있다. 일본에서는 초롱아귀를 심해어 초밥집에서 사용하기도 한다.[47]캐나다 과학자들의 2006년 연구에 따르면, 상업 어업이 대륙붕에서 대륙붕 경사면(수심 약 1600m)으로 이동함에 따라 청대구와 가시장어 등 심해어 5종이 멸종 위기에 처해 있다고 한다. 심해어는 사람과 비슷한 나이에 성적 성숙에 이르며 번식 속도가 느려 과도한 어획으로부터 회복하기 어렵다.[47]
심해어 자원 개발과 이용이 진행되는 한편, 심해 어업의 대규모화에 따른 남획은 큰 문제이다. 저수온에 노출되는 심해어는 성장이 느리고, 특히 저서성의 중·대형 종에서 그 경향이 현저하다.[129] 암수의 접촉이 적고 번식 기회가 제한되어 자원 회복량을 웃도는 규모의 남획은 멸종 위기를 초래할 수 있다. 2006년에는 대서양 북서부에서 대구류의 급속한 감소가 보고되었고,[77] 국제연합식량농업기구(FAO)는 어업 국가에 자원 보호를 위한 적절한 관리를 요구하고 있다.[143]
9. 심해어의 진화와 계통
wikitext
심해 생태계는 육지에서 속씨식물의 출현에 따른 후기 쥐라기와 초기 백악기의 해양으로의 영양분 유입 증가 이전에는 척추동물의 서식에 적합하지 않았을 것이라는 추측이 제기되어 왔다.[1] 이는 심해 무척추동물의 증가를 야기하여, 결과적으로 어류가 이러한 생태계를 식민지화할 수 있게 했다. 그러나 홀로세팔리류와 같은 일부 현생 심해어류는 훨씬 오래된 계통의 후손이라는 점을 고려하면, 화석 증거는 없지만 훨씬 이전에 척추동물이 심해에 진출했을 가능성도 있다.[1]
심해어류에 대한 가장 오래된 기록은 알려지지 않은 신텔레오스테이류의 먹이 활동 및 유영 행동의 생흔 화석으로, 초기 백악기(1억 3천만 년 전)의 이탈리아 팔롬비니 셰일에서 발견되었다. 이 셰일은 과거 피에몬테-리구리아 해의 심해저 평원에 퇴적된 것으로 생각된다.[1] 이 화석이 발견되기 전까지는 고제3기에서 5천만 년보다 오래된 심해 경골어류의 증거는 없었다.[1] 대부분의 현생 심해어류의 백악기 기원설은 오룰포르미스류 어류에 대한 계통 발생 연구와 같이, 이들 그룹의 많은 심해 계통이 이 시기에 기원했음을 보여주는 연구를 통해 더욱 확증되었다.
팔롬비니 셰일의 기록이 가장 오래된 심해 경골어류 기록을 나타내지만, 백악기 후기에 심해 상어 화석을 보존하는 지층도 알려져 있다. 여기에는 캐나다의 노섬벌랜드 지층과 비슷한 시대의 앙골라 퇴적층이 포함되는데, 모두 오늘날 심해 서식지에서 알려져 있지만 당시 다른 지층에서는 드문 분류군인 곱상어류, 클라미도셀라키드류, 그리고 고양이상어류의 화석을 보존하고 있다. 고제3기 지층에서는 뉴질랜드의 중기 팔레오세에서 심해 상어 이빨 화석이 알려져 있으며, 덴마크, 프랑스, 오스트리아, 모로코의 에오세 지층에서도 발견된다. 파라테티스해는 마이오세까지 헝가리 지층에 보존된 심해 상어와 가오리를 지지했다.
고제3기에는 잘 관절된 심해 경골어류 표본을 보존하는 주요 지층들이 알려져 있다. 여기에는 초기 에오세 이탈리아의 몬테 솔라네 라거슈타테가 포함되는데, 이곳은 바다 아래 300~600미터에 퇴적된 것으로 여겨지는 심해대 서식지를 보존하고 있으며, 후기 에오세 파브데 지층(이란)도 포함된다. 두 지층 모두에서 보존된 심해 환경은 풍부한 스토미이포르미스류 어류 화석으로 명확하게 드러난다. 심해 경골어류 화석을 보존하는 주목할 만한 신생대 지층은 이탈리아, 일본, 캘리포니아의 마이오세 지층에서 알려져 있다.
어류가 심해로 진출하기 시작한 시기는 정확히 알 수 없다. 심해는 매우 안정적인 환경이며, 적어도 경골어류의 현저한 다양화가 일어난 중생대 백악기 이전부터 이미 어류가 심해에 서식하고 있었던 것으로 여겨진다.[144] 고생대 석탄기 후기(약 3억 년 전) 지층에서 발견된 꼼치과의 유일한 화석 종은 많은 점에서 현생종과 다르지 않은 형태를 가지고 있었지만,[145] 그 안구는 현존하는 꼼치류와는 다른 기능적인 것이었다.[146] 심해 적응이 어떻게 진행되었는지 알기 위해서는 화석 기록에 기반한 경시적 분석이 필요하지만, 지금까지 알려진 심해어 화석은 매우 부족하다.
현생 심해어의 대부분은 조기강에 속하는 어류, 특히 경골어류의 무리이다. 경골어류 중에서도 원시적인 그룹이 많은 것이 특징이며, 특히 중층 유영성 심해어에서 그 경향이 뚜렷하게 나타난다.[148] 더 진화가 진행된 고위 그룹인 농어목은 현대의 천해에서 가장 번성하는 그룹이지만, 포함된 심해어의 비율은 현저히 적다.
초기 출현 그룹에 심해어가 많고, 비교적 새로운 그룹에는 적은 이유로, 천해에서의 생존 경쟁에서 뒤처진 오래된 어류가 피난처로 삼은 곳이 심해였다고 여겨져 왔다.[56] 그러나 이 설은 1950년대에 부정되었고, 이후 심해어는 진화 계통적으로 크게 두 세대(일차성 ''ancient'' 및 이차성 심해어 ''secondary'')로 나누어 생각하게 되었다.[48][56][58]
일차성 심해어는 외양성 심해어라고도 불리며, 톱상어목이나 앨퉁이목 등 유영성 심해어가 주로 포함된다. 이들은 출현 초기부터 심해로 진출하여, 관상안·발광기 등 천해어와는 거리가 먼 특이한 형태 및 일주 수직 이동 등 독자적인 생태를 매우 긴 시간에 걸쳐 특화시켰다고 여겨진다.[56]
이차성 심해어는 대륙붕성 심해어라고도 불리며, 대구목이나 쥐치목 등 저서성 어류가 속한다. 이들은 초기 진화를 천해의 해저에서 경험한 후, 일차성 심해어보다 늦게 심해저로 진출하게 된 것으로 생각된다. 이 때문에 이차성 심해어가 속하는 분류군에는 얕은 바다에 사는 어류도 많이 포함되며, 형태적으로도 천해어와 극단적인 변화가 없는 경우가 흔하다.[56]
9. 1. 심해어의 진출 시기
심해 생태계는 육지에서 속씨식물의 출현에 따른 후기 쥐라기와 초기 백악기의 해양으로의 영양분 유입 증가 이전에는 척추동물의 서식에 적합하지 않았을 것이라는 추측이 제기되어 왔다.[1] 이는 심해 무척추동물의 증가를 야기하여, 결과적으로 어류가 이러한 생태계를 식민지화할 수 있게 했다. 그러나 홀로세팔리류와 같은 일부 현생 심해어류는 훨씬 오래된 계통의 후손이라는 점을 고려하면, 화석 증거는 없지만 훨씬 이전에 척추동물이 심해에 진출했을 가능성도 있다.[1]심해어류에 대한 가장 오래된 기록은 알려지지 않은 신텔레오스테이류의 먹이 활동 및 유영 행동의 생흔 화석으로, 초기 백악기(1억 3천만 년 전)의 이탈리아 팔롬비니 셰일에서 발견되었다. 이 셰일은 과거 피에몬테-리구리아 해의 심해저 평원에 퇴적된 것으로 생각된다.[1] 이 화석이 발견되기 전까지는 고제3기에서 5천만 년보다 오래된 심해 경골어류의 증거는 없었다.[1] 대부분의 현생 심해어류의 백악기 기원설은 오룰포르미스류 어류에 대한 계통 발생 연구와 같이, 이들 그룹의 많은 심해 계통이 이 시기에 기원했음을 보여주는 연구를 통해 더욱 확증되었다.
팔롬비니 셰일의 기록이 가장 오래된 심해 경골어류 기록을 나타내지만, 백악기 후기에 심해 상어 화석을 보존하는 지층도 알려져 있다. 여기에는 캐나다의 노섬벌랜드 지층과 비슷한 시대의 앙골라 퇴적층이 포함되는데, 모두 오늘날 심해 서식지에서 알려져 있지만 당시 다른 지층에서는 드문 분류군인 곱상어류, 클라미도셀라키드류, 그리고 고양이상어류의 화석을 보존하고 있다. 고제3기 지층에서는 뉴질랜드의 중기 팔레오세에서 심해 상어 이빨 화석이 알려져 있으며, 덴마크, 프랑스, 오스트리아, 모로코의 에오세 지층에서도 발견된다. 파라테티스해는 마이오세까지 헝가리 지층에 보존된 심해 상어와 가오리를 지지했다.
고제3기에는 잘 관절된 심해 경골어류 표본을 보존하는 주요 지층들이 알려져 있다. 여기에는 초기 에오세 이탈리아의 몬테 솔라네 라거슈타테가 포함되는데, 이곳은 바다 아래 300~600미터에 퇴적된 것으로 여겨지는 심해대 서식지를 보존하고 있으며, 후기 에오세 파브데 지층(이란)도 포함된다. 두 지층 모두에서 보존된 심해 환경은 풍부한 스토미이포르미스류 어류 화석으로 명확하게 드러난다. 심해 경골어류 화석을 보존하는 주목할 만한 신생대 지층은 이탈리아, 일본, 캘리포니아의 마이오세 지층에서 알려져 있다.
어류가 심해로 진출하기 시작한 시기는 정확히 알 수 없다. 심해는 매우 안정적인 환경이며, 적어도 경골어류의 현저한 다양화가 일어난 중생대 백악기 이전부터 이미 어류가 심해에 서식하고 있었던 것으로 여겨진다.[144] 고생대 석탄기 후기(약 3억 년 전) 지층에서 발견된 꼼치과의 유일한 화석 종은 많은 점에서 현생종과 다르지 않은 형태를 가지고 있었지만,[145] 그 안구는 현존하는 꼼치류와는 다른 기능적인 것이었다.[146] 심해 적응이 어떻게 진행되었는지 알기 위해서는 화석 기록에 기반한 경시적 분석이 필요하지만, 지금까지 알려진 심해어 화석은 매우 부족하다.
현생 심해어의 대부분은 조기강에 속하는 어류, 특히 경골어류의 무리이다. 경골어류 중에서도 원시적인 그룹이 많은 것이 특징이며, 특히 중층 유영성 심해어에서 그 경향이 뚜렷하게 나타난다.[148] 더 진화가 진행된 고위 그룹인 농어목은 현대의 천해에서 가장 번성하는 그룹이지만, 포함된 심해어의 비율은 현저히 적다.
초기 출현 그룹에 심해어가 많고, 비교적 새로운 그룹에는 적은 이유로, 천해에서의 생존 경쟁에서 뒤처진 오래된 어류가 피난처로 삼은 곳이 심해였다고 여겨져 왔다.[56] 그러나 이 설은 1950년대에 부정되었고, 이후 심해어는 진화 계통적으로 크게 두 세대(일차성 ''ancient'' 및 이차성 심해어 ''secondary'')로 나누어 생각하게 되었다.[48][56][58]
일차성 심해어는 외양성 심해어라고도 불리며, 톱상어목이나 앨퉁이목 등 유영성 심해어가 주로 포함된다. 이들은 출현 초기부터 심해로 진출하여, 관상안·발광기 등 천해어와는 거리가 먼 특이한 형태 및 일주 수직 이동 등 독자적인 생태를 매우 긴 시간에 걸쳐 특화시켰다고 여겨진다.[56]
이차성 심해어는 대륙붕성 심해어라고도 불리며, 대구목이나 쥐치목 등 저서성 어류가 속한다. 이들은 초기 진화를 천해의 해저에서 경험한 후, 일차성 심해어보다 늦게 심해저로 진출하게 된 것으로 생각된다. 이 때문에 이차성 심해어가 속하는 분류군에는 얕은 바다에 사는 어류도 많이 포함되며, 형태적으로도 천해어와 극단적인 변화가 없는 경우가 흔하다.[56]
9. 2. 심해어의 분류
현생종은 갯지렁이목과 칠성장어목 두 종류뿐이며, 칠성장어목은 주로 담수에 서식한다. 무악류에 속하는 갯지렁이목은 약 70여 종으로, 모래 진흙 바닥에 서식하며 심해에서 썩은 고기를 먹는 동물로서 중요한 위치를 차지한다. 갯지렁이목에는 갯지렁이와 가는갯지렁이가 있다.연골어류에는 상어, 가오리, 은상어류가 속하며, 저서성 심해어가 많이 포함된다.
- '''은상어목''' 은 약 30종 모두 심해 저층에 서식한다. 조개류, 갑각류 등 단단한 껍질을 가진 매몰성 동물을 강력한 턱으로 포식한다. 은상어목에는 코끼리상어과, 긴코상어과, 은상어(은상어과)가 있다.
- '''악상어목''' 은 표층에서 심해까지 넓은 서식 범위를 가지는 종이 많고, 순수한 심해성 어류는 적다. 고블린상어(고블린상어과)와 메가마우스(메가마우스과)가 악상어목에 속한다.
- '''흉상어목''' 은 상어류 중 가장 큰 그룹이며, 대부분은 천해에 서식하지만 까치상어과의 일부 속에는 심해어가 포함된다. 이들은 대부분 대륙붕에 서식하는 저서어류이며, 거의 헤엄치지 않는다. 흉상어목에는 까치상어과가 있다.
- '''갈고리상어목''' 에 포함된 5종은 모두 중심층 해저 근처에 사는 심해어이다. 낙지상어(낙지상어과)와 갈고리상어(갈고리상어과)가 갈고리상어목에 속한다.
- '''가시상어목''' 에 속한 2종은 모두 저서성 심해어이다. 가시상어과가 이 목에 속한다.
- '''뿔상어목''' 은 약 100종의 대부분이 심해 저서성이다. 기름뿔상어(뿔상어과), 쥐상어과, 푸른쥐상어(까마귀상어과), 톱상어(톱상어과), 박쥐상어(요새상어과), 온딘상어(온딘상어과)가 뿔상어목에 속한다. 쥐상어과에는 6,000미터 이상의 심해에서 채집된 기록이 있지만, 그 신뢰성은 의문시되고 있다.
- '''홍어목''' 은 대부분 대륙붕에서 대륙사면에 걸쳐 서식하는 저서어류이며, 분포 범위는 천해에서 심해까지 다양하다. 긴가오리(긴가오리과)와 가시가오리(가시가오리과)가 홍어목에 속한다.
조기어강에는 현생 경골어류 대부분이 포함되며, 속하는 약 40개의 목 중 절반은 심해에 적응한 형태를 보인다.
- '''소토이와시목''' 소코기스아목의 대부분이 저서성 심해어이다. 소코기스과의 무리는 심해성 표재동물(말미잘, 거미불가사리, 바다나리, 성게 등)을 선택적으로 포식한다. 도카게기스(도카게기스과)와 소코기스(소코기스과)가 소토이와시목에 속한다.
- '''뱀장어목''' 에서는 주로 붕장어아목에 심해어가 많이 포함된다. 시기우나기과와 노코바우나기과는 중층 유영성이며, 그 외 그룹은 저서성이다. 이라코아나고와 콩고우아나고(호라아나고과), 바케후사아나고(후사아나고과), 헤라아나고과, 시기우나기(시기우나기과), 쿠즈아나고과, 노코바우나기과가 뱀장어목에 속한다.
- '''풍선장어목''' 에 속하는 28종은 모두 심해어이며, 중심층에서 점심층을 표영한다. 야바네우나기과, 풍선장어(풍선장어과), 주머니장어(주머니장어과), 탕가쿠우나기과가 풍선장어목에 속한다.
- '''니기스목''' 은 과거 오이우오목에 속했던 무리이다. 약 200종의 대부분이 중심층 표영성 심해어이며, 일부 저서어도 포함된다. 니기스(니기스과), 데메니기스(데메니기스과), 소코이와시과, 하나메이와시(하나메이와시과), Bathylaconidae 과, 세키토리이와시(세키토리이와시과)가 니기스목에 속한다.
- '''악어고기목''' 에 속하는 약 400종은 거의 모든 종이 중심층 유영성이다. 개체 수가 매우 많으며, 발광기관을 가진 종류도 많다. 유메하다카과, 오니하다카(요코에소과), 호우넨에소(무네에소과), 긴하다카과, 호우라이에소와 오오쿠치호시에소, 미츠마타야리우오(악어고기과)가 악어고기목에 속한다.
- '''샤치브리목''' 은 12종이 포함되며, 모두 저서성이다. 샤치브리(샤치브리과)가 이 목에 속한다.
- '''히메목''' 은 초우친하다카 등의 저서성 어류와 미즈우오 등 유영성 심해어를 함께 포함한다. 자웅동체인 어류가 많이 포함되는 것이 특징이다. 나가즈에에소와 이토히키이와시(초우친하다카과), 후데에소과, Bathysauropsidae 과, 데메에소과, 야리에소과, 미즈우오(미즈우오과), 하다카에소과, 신카이에소(신카이에소과), 보우엔굥(보우엔굥과)이 히메목에 속한다.
- '''벌레고기목''' 은 약 250종이 속하며, 악어고기목 어류와 함께 중심층 유영성 심해어의 대표적인 존재이다. 거의 모든 무리가 발광기를 가지며, 극해를 포함한 전 세계 해양에 분포하여 총 생물량이 막대하다. 소토오리이와시과와 벌레고기(벌레고기과)가 벌레고기목에 속한다.
- '''붉은돔목''' 은 약 20종의 대부분이 심해성이며, 드문 종류가 많다. クサアジ(クサアジ과), 붉은돔(붉은돔과), 스튜레포르스(스튜레포르스과), 붉은날개(붉은날개과), 라디이케팔루스과, 긴날개물고기(긴날개물고기과), 류우구우노츠카이(류우구우노츠카이과)가 붉은돔목에 속한다.
- '''은색돔목''' 은 10종만을 포함하는 작은 그룹으로, 모든 종이 중심층 저부에 서식한다. 은색돔(은색돔과)이 은색돔목에 속한다.
- '''대구목''' 에 속하는 500종 이상의 대부분이 심해어이다. 소코다라, 치고다라 무리는 중심층에서 심해층에 걸쳐 넓게 분포하는 저서성 어류이며, 종류와 개체 수가 모두 많다. 붕장어대구과, 긴다리대구과, 요로이다라와 신카이요로이다라(소코다라과), 치고다라(치고다라과), 대구와 실버헤이크(대구과), 명태(대구과)가 대구목에 속한다.
- '''아시로목''' 에서 아시로과의 어류는 소코다라류와 마찬가지로 수가 많은 중요한 저서성 심해어이다. 소코보우즈와 요미노아시로(아시로과), 소코오쿠메우오과가 아시로목에 속한다.
- '''아귀목''' 에 기재된 300종 가까이 되는 대부분이 심해에서 생활한다. 초우친앙코우상과의 무리는 점심층 유영성 심해어로서 대표적인 존재이다. 다른 그룹은 저서성이다. 아귀와 키앙코우(아귀과), 후사앙코우과, 붉은갯가지(붉은갯가지과), 긴지느러미초롱아귀과, 송곳니아귀과, 검은아귀과, 초롱아귀(초롱아귀과), 두가지아귀과, 낙타아귀과, 타우마티크티우스과, 거친아귀과, 긴꼬리초롱아귀(긴꼬리초롱아귀과), 고사리아귀과, 악마아귀(악마아귀과)가 아귀목에 속한다.
- '''고래물고기목'''(관무리킨메다이목)은 체형이나 체색에 특색이 많고, 다른 목과 구별이 용이한 무리이다. 중심층에서 점심층에서 유영 생활을 하며, 특히 고래물고기과 어류는 1,800미터 이하에서 지배적인 존재이다. 투구물고기(투구물고기과), 관무리킨메다이과, 히스피도베리쿠스과, 기이한물고기(기이한물고기과), 아귀물고기과, 붉은고래물고기과, 고래물고기(고래물고기과), 톡비레이와시과, 소코쿠지라우오과가 고래물고기목에 속한다.
- '''금눈돔목''' 은 산호초 지역에서 심해까지 넓은 분포역을 가진다. 오니킨메류는 점심층 유영성이고, 히우치다이, 킨메다이 무리는 저서성이다. 오니킨메(오니킨메과), 나카무라긴메과, 히우치다이과, 금눈돔(금눈돔과)이 금눈돔목에 속한다.
- '''쥐돔목''' 에 속하는 30종 가까이 대부분이 대륙붕에서 대륙사면에 걸쳐 서식하는 저서성 심해어이며, 분포 범위가 넓은 종류가 많다. 큰눈쥐돔과, 붉은쥐돔과, 바닥쥐돔과, 마름모쥐돔과가 쥐돔목에 속한다.
- '''꼼치목''' 은 대부분 넓은 분포역을 가지며, 심해에 특화된 과는 적다. 쿠사우오과에는 해구 심부에 서식하는 초심해종이 포함된다. 키호우보우과, 붉은꼼치과, 대구와 아브라보우즈(대구과), 쥐치과, 심해쿠사우오(쿠사우오과)가 꼼치목에 속한다.
- '''농어목''' 은 현대 천해에서 가장 번성하며, 최대 어종(약 1만 종)을 자랑하는 그룹이다. 심해어도 다수 포함되지만, 꼼치목과 마찬가지로 과 수준에서 심해에 적응한 예는 적다. 북대서양에서의 추산에 따르면, 농어목 어류는 유영성 심해어의 6%, 저서성 심해어의 9%를 차지하는 데 불과하다. 200종 이상을 포함하는 겐게과는 북반구의 심해저에 넓게 분포하는 중요한 저서어이며, 짙은 무리를 형성하기도 한다. 이시나기과, 야세무츠과, 야에기스과, 소코니신과, 겐게과, 아르테디드라콘과, 악마농어(검은입큰농어과), 이레즈미곤냐쿠아지(이레즈미곤냐쿠아지과), 무카시쿠로타치과, 바람치와 아브라소콤츠(갈치과), 큰눈돔과, 쥐돔(쥐돔과)이 농어목에 속한다.
- '''넙치목''' 은 많은 과가 천해종과 심해종을 고루 포함한다. 우시노시타과에 속하는 2아과 중, 아즈마가레이아과는 심해성이다. 가와라가레이과와 벨로가레이과가 넙치목에 속한다.
- '''복어목''' 은 천해어가 많지만, 베니카와무키과의 약 20종은 저서성 심해어이다. 베니카와무키과와 만보우과(Molidae)가 복어목에 속한다.
육기류에 속하며 사지동물의 조상으로 여겨지는 무리이다. 현생종은 폐어류와 실러캔스류뿐이다.
- '''실러캔스목''' 은 살아있는 화석이라고도 불리며, 현생종은 2종이다. 라티메리아(라티메리아과)가 실러캔스목에 속한다.
10. 한국의 심해어 연구
참조
[1]
논문
[36]
서적
Progenetic tendencies in deep-sea fishes
Fisheries Society of the British Isles
[37]
논문
Detection of the Largest Deep-Sea-Endemic Teleost Fish at Depths of Over 2,000 m Through a Combination of eDNA Metabarcoding and Baited Camera Observations
2022-07-01
[38]
논문
The swim bladder as a juvenile organ in stromateoid fishes
https://archive.org/[...]
[39]
웹사이트
In situ observation of a macrourid fish at 7259 m in the Japan Trench: swimbladder buoyancy at extreme depth
https://journals.bio[...]
[40]
논문
Location by olfaction: a model and application to the mating problem in the deep-sea Hatchetfish Argyropelecus hemigymnus
[41]
논문
Precocious sexual parasitism in the deep sea ceratioid anglerfish, Cryptopsaras couesi Gill
http://www.nature.co[...]
[42]
서적
A Guide to the Study of Fishes
https://archive.org/[...]
H. Holt and Company
[43]
웹사이트
[44]
서적
The Universe as It Really Is: Earth, Space, Matter, and Time
https://columbia.deg[...]
Columbia University Press
[45]
논문
Distribution, composition and functions of gelatinous tissues in deep-sea fishes
2017
[46]
논문
Adaptations to High Hydrostatic Pressure
http://www.annualrev[...]
[47]
논문
Fisheries: Deep-sea fishes qualify as endangered
[48]
서적
深海魚 暗黒街のモンスターたち
[49]
뉴스
世界一深い場所で泳ぐ魚 東京海洋大など:水深8336メートルで撮影 ギネス記録
https://www.yomiuri.[...]
読売新聞
2023-04-05
[50]
웹사이트
北海道大学総合博物館第8回公開シンポジウム・深海魚の多様性
http://museum-sv.mus[...]
北海道大学総合博物館、北海道大学 水産学部/大学院水産科学院/大学院水産科学研究院|大学院水産科学研究科
2011-05-21
[51]
서적
Fishes of the World Third Edition
[52]
서적
Fishes of the World Fourth Edition
[53]
서적
The Diversity of Fishes Second Edition
[54]
논문
How many recent fishes are there?
[55]
서적
Fishes of the World Third Edition
[56]
서적
Deep-Sea Fishes
[57]
서적
The Diversity of Fishes Second Edition
[58]
서적
魚学入門
[59]
서적
深海調査船が観た深海生物
[60]
논문
Search for life in deep biospheres
[61]
서적
Deep-Sea Fishes
[62]
서적
深海と深海生物 美しき神秘の世界
[63]
서적
深海の生物学
[64]
웹사이트
マリアナ海溝チャレンジャー海淵において世界で初めて底生生物の採集に成功
https://www.jamstec.[...]
海洋科学技術センター
2014-04-19
[65]
서적
深海の生物学
[66]
웹사이트
DEEP OCEAN 超深海/地球最深(フルデプス)への挑戦
https://web.archive.[...]
NHKホームページ
[67]
논문
The deepest living fish Abyssobrotula galatheae. A new genus and species of oviparous ophidioids (Pisces, Brotulidae)
[68]
논문
A new fish of the snailfish family (Pisces, Liparidae) found at a depth of more than 7 kilometers
[69]
논문
A hadal fish (Bassogigas profundissimus) with a functional swimbladder
[70]
서적
深海生物図鑑
[71]
웹사이트
マリアナ海溝の水深8178mにおいて魚類の撮影に成功~魚類の世界最深映像記録を更新~
https://www.jamstec.[...]
JAMSTECプレスリリース
2017-08-25
[72]
서적
海洋生物の機能
[73]
서적
Deep-Sea Fishes
[74]
논문
Hyperbaric fish trap operation and deployment in the deep sea
[75]
서적
潜水調査船が観た深海生物
[76]
서적
深海生物ファイル
[77]
논문
Deep-sea fishes qualify as endangered
[78]
서적
Deep-Sea Fishes
[79]
서적
深海生物図鑑
[80]
서적
潜水調査船が観た深海生物
[81]
서적
Deep-Sea Fishes
[82]
서적
Fishes of the World Fourth Edition
[83]
서적
The Diversity of Fishes Second Edition
[84]
서적
Deep-Sea Fishes
[85]
서적
深海生物図鑑
[86]
서적
深海の生物学
[87]
논문
The buoyancy of bathypelagic fishes without a gas-filled swimbladder
[88]
서적
The Diversity of Fishes Second Edition
[89]
서적
Adaptations to the deep-sea environment. In Handbook of sensory physiology. Vol. VII/5. The visual system in vertebrates
Springer-Verlag
[90]
서적
深海生物図鑑
[91]
서적
深海の生物学
[92]
논문
Vision in the deep sea.
[93]
논문
The eyes of deep-sea fishes and the changing nature of visual scenes with depth.
[94]
서적
日本の海水魚
[95]
서적
Deep-Sea Fishes
[96]
서적
深海生物図鑑
[97]
서적
深海の生物学
[98]
서적
深海の生物学
[99]
논문
Mechanism of reflexion in silvery layers of fish and cephalopods
[100]
논문
Modulation of light localization in the iridophores of the deep-sea highlight hatchetfish Sternoptyx pseudobscura under magnetic field
https://aip.scitatio[...]
2021-09-14
[101]
서적
深海の生物学
[102]
서적
Oceanic Anglerfishes
[103]
서적
深海の生物学
[104]
논문
Light organ symbioses in fishes
[105]
서적
深海の生物学
[106]
서적
深海の生物学
[107]
서적
Luminescent organs. In The Mollusca. Vol. 11. Form and function
Academic Press
[108]
논문
The eyes of deep-sea fish. I: Lens pigmentation, tapeta and visual pigments
[109]
논문
On the 'filters' in the photophores of mesopelagic fish and on a fish emitting red light and especially sensitive to red light
[110]
서적
深海の生物学
[111]
서적
深海の生物学
[112]
서적
深海の生物学
[113]
서적
深海の生物学
[114]
논문
Review of oceanic luminescence
[115]
서적
深海生物図鑑
[116]
서적
深海の生物学
[117]
서적
Deep-Sea Fishes
[118]
서적
Deep-Sea Fishes
[119]
서적
深海の生物学
[120]
서적
The economy of nature and the evolution of sex
University of California Press
[121]
논문
Dimorphism, parasitism and sex: reproductive strategies among deep-sea ceratioid anglerfishes
[122]
서적
Fishes of the World Fourth Edition
[123]
서적
The Diversity of Fishes Second Edition
[124]
서적
日本の海水魚
[125]
서적
The Diversity of Fishes Second Edition
[126]
논문
The olfactory organs of bathypelagic fishes
[127]
서적
深海生物図鑑
[128]
서적
深海魚 暗黒街のモンスターたち
[129]
서적
Deep-Sea Fishes
[130]
서적
深海生物図鑑
[131]
서적
Deep-Sea Fishes
[132]
뉴스
深海魚と地震は無関係 場所一致わずかと東海大
https://www.sankei.c[...]
산케이 신문
2017-07-13
[133]
서적
深海魚 暗黒街のモンスターたち
[134]
서적
日本の海水魚
[135]
서적
深海生物ファイル
[136]
간행물
厚生省通達
1970-09-04, 1981-01-10
[137]
웹사이트
ほかでは味わえない「深海魚」
http://www.city.numa[...]
沼津市観光WEB
2017-08-02
[138]
뉴스
食感の「旬」到来 ふっくら脂乗る
일본경제신문
2017-09-30
[139]
서적
潜水調査船が観た深海生物
[140]
서적
魚類環境生態学入門
[141]
저널
Characterization of lipids in myctophid fish in the subarctic and tropical Pacific
[142]
서적
Deep-Sea Fishes
[143]
웹사이트
Better management for fishing's 'last frontier'
http://www.fao.org/n[...]
FAO
2010-05-02
[144]
서적
The Diversity of Fishes Second Edition
[145]
서적
海の動物百科2 魚類I
[146]
서적
The Diversity of Fishes Second Edition
[147]
서적
The Diversity of Fishes Second Edition
[148]
서적
Deep-Sea Fishes
[149]
서적
Deep-Sea Fishes
[150]
웹사이트
NZ IPY-CAML Voyage 2008
http://www.fish.govt[...]
2008
[151]
서적
日本産魚類検索 全種の同定 第三版
[152]
서적
日本産魚類検索 全種の同定 第三版
[153]
서적
深海魚 暗黒街のモンスターたち
[154]
웹사이트
北海道大学総合博物館第8回公開シンポジウム・深海魚の多様性
http://museum-sv.mus[...]
北海道大学総合博物館・大学院水産科学研究科
2011-05-21
[155]
서적
Hazardous Constructions
http://dx.doi.org/10[...]
University of Minnesota Press
2011-09-27
[156]
서적
Bamboo material characterisation
http://dx.doi.org/10[...]
Elsevier
2016
[157]
서적
Physical Constants and Formulas
http://dx.doi.org/10[...]
Elsevier
2011
[158]
저널
Poly-L-Lactic Acid
http://dx.doi.org/10[...]
2004
[159]
저널
Discussion
http://dx.doi.org/10[...]
2002
[160]
서적
Geschlechterforschung und Organisationssoziologie
http://dx.doi.org/10[...]
VS Verlag für Sozialwissenschaften
2002
[161]
저널
Discussion
http://dx.doi.org/10[...]
2002
[162]
저널
Trujillo, Guillermo
http://dx.doi.org/10[...]
Oxford University Press
2011-10-31
[163]
저널
Trujillo, Guillermo
http://dx.doi.org/10[...]
Oxford University Press
2011-10-31
[164]
저널
Deep-sea fish
https://en.wikipedia[...]
2022-05-15
[165]
저널
Deep-sea fish
https://en.wikipedia[...]
2022-05-15
[166]
기타
[167]
저널
Moyle 2004
[168]
기타
[169]
웹사이트
Deep-sea creatures: The mesopelagic zone
https://teara.govt.n[...]
2007-09-21
[170]
서적
Bone & Moore 2008
[171]
논문
Fishes in oxygen-minimum zones: blood oxygenation characteristics
http://www.sciencema[...]
[172]
저널
Moyle 2004
[173]
저널
Moyle 2004
[174]
웹사이트
Deep-sea creatures: The mesopelagic zone
http://www.teara.gov[...]
2007-09-21
[175]
웹사이트
Deep-sea creatures: The bathypelagic zone
http://www.teara.gov[...]
2007-09-21
[176]
저널
The Universe as It Really Is
http://dx.doi.org/10[...]
2018-08-27
[177]
서적
Deep-Sea Fishes
http://dx.doi.org/10[...]
Cambridge University Press
2017
[178]
서적
Fishes of the World Fourth Edition
[179]
서적
The Diversity of Fishes Second Edition
[180]
서적
Fishes of the World Third Edition; The Diversity of Fishes Second Edition
1970
[181]
일반
[182]
저널
How many recent fishes are there?
[183]
서적
Fishes of the World Third Edition
[184]
서적
GLEANINGS AND ORIGINAL MEMORANDA
http://dx.doi.org/10[...]
Cambridge University Press
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com