곡비원류
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
곡비원류(Strepsirrhini)는 젖은 코와 콤마 모양 콧구멍을 특징으로 하는 분류학적 명칭으로, 여우원숭이, 로리스, 갈라고 등을 포함하는 영장류의 한 무리이다. 이들은 5500만 년에서 9000만 년 전에 활비원류와 분화되었으며, 아다피형류에서 진화했다. 곡비원류는 빗 모양의 치아, 그루밍 발톱, 휘판을 통한 야간 시력 향상 등의 해부학적 특징을 가지고 있다. 현재 여우원숭이형 하목과 로리스형 하목으로 나뉘며, 멸종된 아다피스류도 포함된다. 대부분 야행성이며, 냄새 표시를 통해 소통한다. 곡비원류는 열대 지역에 분포하며, 산림 벌채와 같은 인간 활동으로 인해 멸종 위기에 처해 있다.

더 읽어볼만한 페이지
- 1812년 기재된 분류군 - 협비원류
협비원류는 콧구멍이 아래쪽을 향하고 잡는 꼬리가 없으며 납작한 손톱과 발톱, 관 모양 외고막을 가진 구세계원숭이와 유인원을 포함하는 영장류 분류군으로, 약 3,000만~4,000만 년 전 신세계원숭이와 분기되었고 일부 종은 3색형 색각을 갖는다. - 1812년 기재된 분류군 - 늘보로리스
늘보로리스는 남아시아와 동남아시아에 사는 로리스과의 야행성 영장류로, 둥근 머리와 뭉툭한 꼬리를 가지며 몸에 독성이 있고, 삼림 벌채와 야생동물 거래로 인해 멸종 위기에 처해 있으며 7종이 알려져 있다. - 영장류 - 유인원
유인원은 꼬리 없는 영장류로, 인간을 포함하여 긴팔원숭이과와 사람과로 나뉘며, 높은 지능과 복잡한 사회 구조를 공유하지만 서식지 파괴와 사냥으로 멸종 위기에 처해 있다. - 영장류 - 원숭이
원숭이는 신세계원숭이와 구세계원숭이를 포함하는 영장류의 한 종류로, 크기와 몸무게, 서식지가 다양하며 인간과 유사한 특징을 지니고 있어 다양한 문화권에서 상징적인 의미를 지니고 있고 여러 목적으로 활용되지만 윤리적 논쟁 또한 존재한다. - 에티엔 조프루아 생틸레르가 명명한 분류군 - 무덤박쥐속
무덤박쥐속은 대꼬리박쥐과에 속하는 박쥐 속으로, 해안대꼬리박쥐, 대꼬리박쥐, 힐데가르트무덤박쥐, 모리셔스무덤박쥐, 이집트무덤박쥐 등 다양한 종을 포함하며, 이들은 주로 어두운 곳에 서식하며 밤에 곤충을 잡아먹는다. - 에티엔 조프루아 생틸레르가 명명한 분류군 - 이집트과일박쥐
이집트과일박쥐는 아프리카, 중동, 인도 등지에 분포하며 과일을 주식으로 하고 동굴에 군집생활을 하며 반향정위를 사용하는 큰박쥐류의 일종으로, 인간과 질병 매개, 농작물 피해, 전시 등의 관계를 맺고 있다.

2. 어원
분류학적 명칭인 곡비원류(Strepsirrhini)는 고대 그리스어의 στρέψις|스트렙시스grc(strepsis, "돌아감")와 ῥίς|리스grc(rhis, "코, 주둥이, (복수형) 콧구멍")에서 유래되었으며,[2] 이는 젖은 코인 비갑개의 구불구불한(콤마 모양의) 콧구멍 모양을 가리킨다. 이 명칭은 프랑스 박물학자 에티엔 조프루아 생틸레르가 1812년에 아목으로 처음 사용했다.
곡비원류는 약 5500만 년에서 9000만 년 전에 활비원류(원숭이, 유인원, 안경원숭이)와 분화되었다. 이 분화 시기는 유전자 분석과 화석 기록에 따라 차이가 있는데, 유전자 분석은 더 오래된 시기를, 화석 기록은 더 최근 시기를 제시한다.
3. 진화 역사
원숭이류는 아시아에서 기원했을 가능성이 있는 케르카모니나이 또는 시바라다피다이와 같은 아다피형류에서 진화했을 수 있다. 과거에는 유럽의 아다피다이에서 진화했다고 생각되기도 했다.
여우원숭이는 4700만 년에서 5400만 년 사이에 아프리카에서 마다가스카르로 이동했으며,[4] 로리스는 약 4000만 년 전에 아프리카 갈라고에서 분리되어 아시아로 이동했다.[4] 여우원숭이는 '살아있는 화석' 등으로 불리기도 하지만, 이는 냄새 의존성, 골격 특징, 작은 뇌 크기 등 곡비원류의 진화를 이해하는 데 방해가 되는 견해이다. 여우원숭이는 마다가스카르에서 다양한 생태적 지위를 채우도록 진화했다.
초기 영장류의 기원은 불분명하며, 화석 기록과 유전자 분석 결과 간의 차이로 인해 논쟁이 계속되고 있다. 초기 영장류는 야행성 및 주행성 소형 종을 모두 포함하며, 나무 위 생활에 적합한 손과 발을 가지고 있었다.
최초의 진정한 영장류(유영장류)는 초기 에오세 (~5500만 년 전)에 화석 기록에 나타나며, 여기에는 칸티우스, 돈루셀리아, 알타니우스, 테일하르디나 등이 포함된다.
3. 1. 아다피스형류의 진화
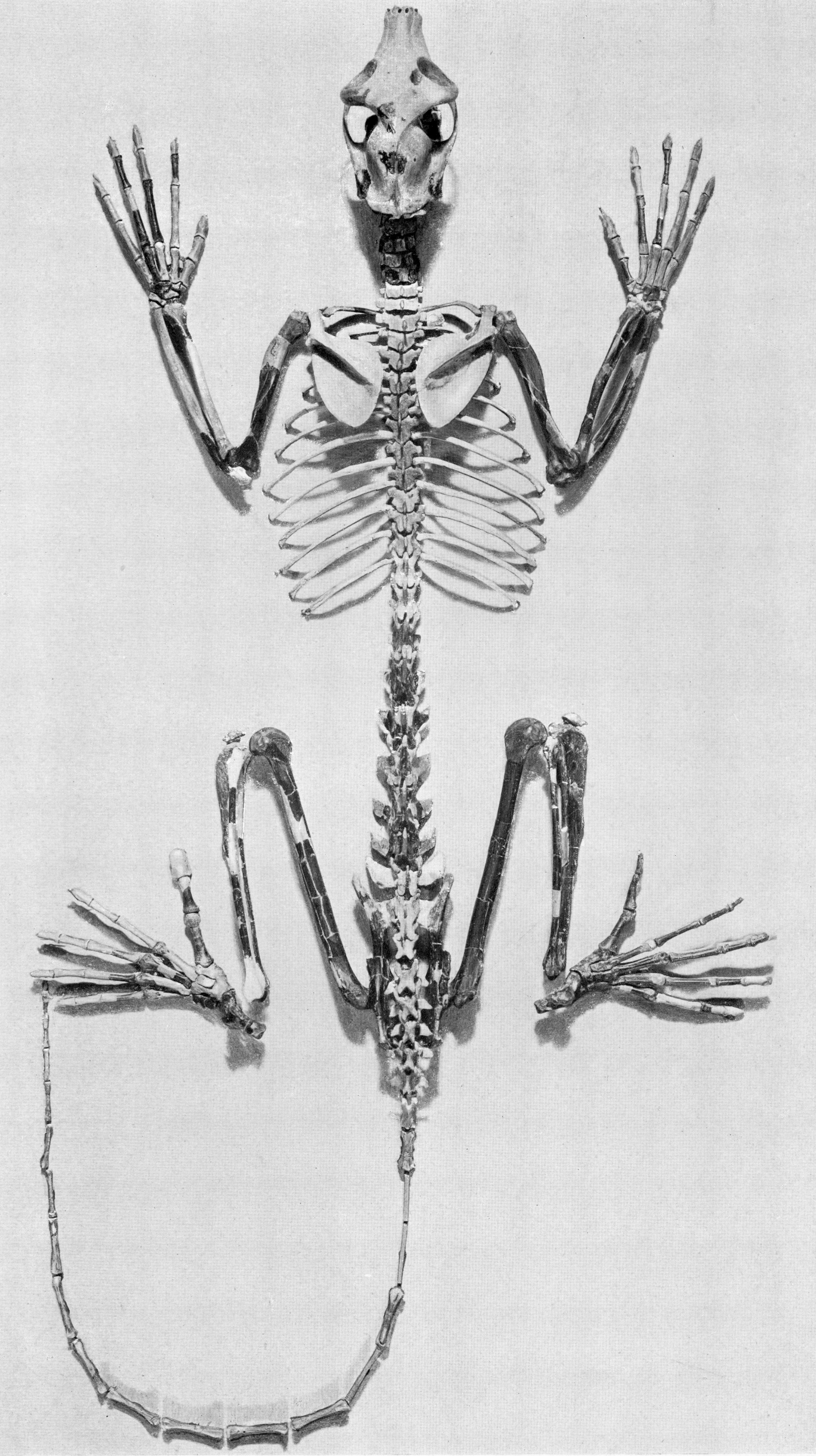
아다피형 영장류는 여우원숭이와 많은 해부학적 유사점을 공유했던 멸종된 곡비원류이다. 여우원숭이와 같은 영장류로 언급되기도 하지만, 여우원숭이와 아다피형류의 다양성은 이러한 유추를 뒷받침하지 않는다.
현존하는 곡비원류와 마찬가지로 아다피형류는 매우 다양했으며, 2000년대 초반 기준으로 화석 기록에서 최소 30개 속과 80개 종이 알려져 있다. 이들은 에오세 동안 라시아 전역에서 다양화되었으며, 일부는 육교를 통해 북아메리카에 도달했다. 당시 화석층에서 발견되는 가장 흔한 포유류 중 하나였으며, 몇몇 희귀종은 북아프리카에서도 발견되었다. 아다피형류 중 가장 기저에 있는 종에는 북아메리카와 유럽의 ''칸티우스''(Cantius) 속과 유럽의 ''돈루셀리아''(Donrussellia) 속이 있다. ''돈루셀리아''는 가장 많은 원시 형질을 지니고 있어 다른 아다피형류의 자매군 또는 줄기군으로 여겨진다.
아다피형류는 종종 세 개의 주요 그룹으로 나뉜다.3. 2. 여우원숭이형류의 진화
여우원숭이류의 기원은 불분명하며 논쟁의 대상이다. 미국의 고생물학자 필립 진저리치는 여우원숭이류 영장류가 유럽의 몇몇 아다피스과 속에서 진화했다는 가설을 제시했는데, 이는 아다피스과의 앞쪽 아래쪽 치아와 현존하는 여우원숭이류의 치아빗 사이의 유사성에 근거한 것이었다. 그러나 명확한 과도기 화석의 부족으로 인해 이 견해는 강력한 지지를 받지 못하고 있다. 대신, 여우원숭이류는 아시아의 아주 초기의 세르카모니과 또는 시발라다피스과 분기에서 유래하여 북아프리카로 이주했을 수 있다.[1][2]
1997년과 2005년 사이에 이집트의 엘 파이움 퇴적물에서 4천만 년 된 3개의 화석 로리스류(''카라니시아'', ''사하라갈라고'', 그리고 ''와딜레무르'')가 발견되기 전까지, 가장 오래된 여우원숭이류는 케냐와 우간다의 초기 미오세(~2천만 년 전)에서 발견되었다. 이 새로운 발견은 여우원숭이류 영장류가 아프로-아라비아에서 에오세 중기에 존재했으며, 여우원숭이류 계통과 다른 모든 협비원류 분류군이 그 이전에 분화되었음을 보여준다.[3][4] 튀니지의 ''제벨레무르''는 후기 초기 또는 초기 중기 에오세(5천 2백만 년에서 4천 6백만 년 전)로 거슬러 올라가며, 세르카모니과로 간주되어 왔지만, 줄기 여우원숭이류일 수도 있다.[1] 아지비드는 알제리에서 발견되었으며, 대략 같은 시기에 살았고, 제벨레무르류의 자매 그룹일 수 있다. 후기 에오세 이집트에서 발견된 ''플레시오피테쿠스''와 함께, 이 셋은 아프리카의 줄기 여우원숭이류로 분류될 수 있다.[2]
분자 시계 추정에 따르면, 여우원숭이와 로리스류는 고신세 동안 아프리카에서 약 6천 2백만 년 전에 분화되었다. 4천 7백만 년에서 5천 4백만 년 전 사이에, 여우원숭이들은 뗏목을 이용하여 마다가스카르로 이동했다.[4] 고립된 상태에서 여우원숭이들은 다양화되었고, 오늘날 원숭이와 유인원이 채우는 생태적 지위를 채웠다.[5] 아프리카에서 로리스와 갈라고는 에오세 동안 약 4천만 년 전에 분화되었다.[4] 마다가스카르의 여우원숭이들과 달리, 그들은 원숭이와 유인원뿐만 아니라 다른 포유류들과도 경쟁해야 했다.[6]
4. 분류
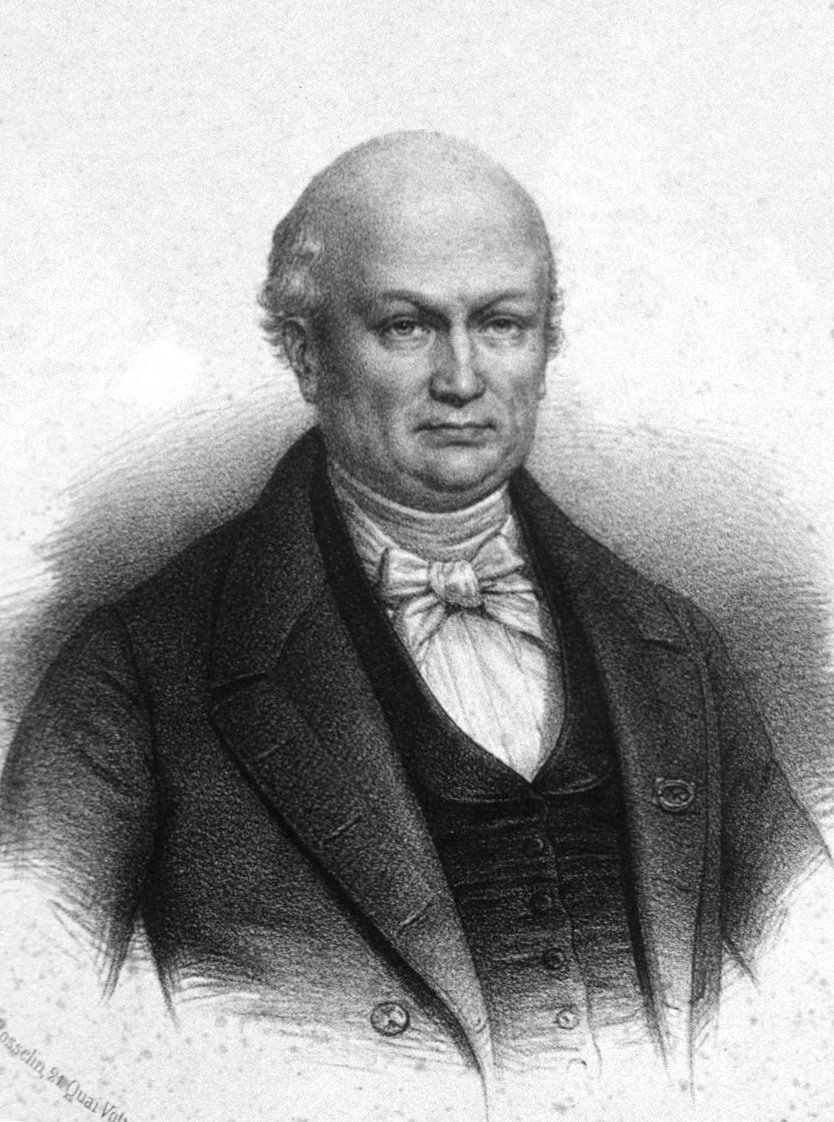
굽은코원류의 분류는 복잡하며 논란의 여지가 있다. 1758년 칼 린네는 ''Lemur'' 속에 단 세 종만을 분류했으나, 이후 여러 학자들이 다양한 분류 체계를 제시하면서 혼란이 발생했다. 특히 1812년 E. Geoffroy가 굽은코원류(Strepsirrhini)라는 아목을 제안했지만, 1918년까지 주목받지 못했다. 1953년 이후 굽은코원류와 직비원류(Haplorhini)로 나누는 분류가 널리 사용되기 시작했지만, 여전히 여우원숭이형 하목과 로리스형 하목을 포함하는 분류 체계와 원원류 아목(Prosimii)과 진원류 아목(Anthropoidea)으로 나누는 분류 체계가 혼용되고 있다.[5]
학명 Strepsirrhini는 고대 그리스어에서 "꼬인·구부러진"을 의미하는 streptos와 "코"를 의미하는 rhis에서 유래되었다.[5] 일본어 명칭은 1989년에 곡비원류로 번역되었다.[5]
굽은코원류는 크게 여우원숭이형 하목과 로리스형 하목으로 나뉜다. 일부 학자들은 로리스형 하목을 여우원숭이형 하목에 포함시키기도 한다.[9]
4. 1. 하위 분류
여우원숭이형 하목(Lemuriformes)과 로리스형 하목(Lorisiformes)으로 나뉜다.[6][7]- 여우원숭이형 하목(Lemuriformes)
- * 아이아이상과(Daubentonioidea)
- ** 아이아이과(Daubentoniidae)
- * 여우원숭이상과(Lemuroidea)
- ** 난쟁이여우원숭이과(Cheirogaleidae)
- ** 여우원숭이과(Lemuridae)
- ** 소형여우원숭이과(Lepilemuridae)
- ** 인드리과(Indriidae)
- 로리스형 하목(Lorisiformes)
- * 갈라고과(Galagonidae)
- * 로리스과(Lorisidae)
로리스형 하목에 분류되는 과를, 여우원숭이형 하목에 포함시키는 설도 있다.[9]
5. 해부학적 특징
곡비원류는 젖은 코(비강)와 빗 모양의 치아(치아빗) 등 원숭이류와 공유되지 않는 특징을 가지고 있다.[5] 콧구멍은 굽어진 틈새 모양이며, 코끝은 축축하고 코 중앙에서 윗입술까지 홈(인중)이 나 있다.[5] 아래턱의 앞니와 송곳니는 빗 모양(치아빗)을 이룬다.[9]
대부분 손가락과 발가락에 납작한 발톱(평발톱)이 있지만, 뒷다리 두 번째 발가락에는 털 손질에 사용되는 갈고리 모양의 발톱(그루밍 발톱)이 있다.[9] 다만, 아이아이는 예외적으로 치아빗이 없고, 손가락과 발가락에 갈고리 발톱이 있으며 뒷다리 첫 번째 발가락에만 평발톱이 있다.[9]
망막 뒤쪽에 빛을 반사하는 막(타페텀)이 있어 야간 시력에 유리하다.[8] 또한, 대부분의 곡비원류는 비타민 C를 자체적으로 생합성할 수 있어 식단에서 비타민 C를 섭취할 필요가 없다.[10]
5. 1. 생식 생리
곡비원류는 상피융모막 태반을 가지고 있으며, 이는 혈액융모막 태반을 가진 단비원류와 달리, 모체의 혈액이 태아의 융모막과 직접 접촉하지 않는 특징이 있다.[1] 곡비원류의 자궁은 두 개의 뚜렷한 방(쌍각 자궁)을 가지고 있다.[2] 크기가 비슷한 단비원류와 임신 기간은 유사하지만, 곡비원류의 태아 성장률은 일반적으로 더 느린 편이다. 이로 인해 단비원류 신생아 크기의 3분의 1 정도 되는 신생아를 출산한다.[2][3] 존속하는 곡비원류는 낮은 기초 대사율을 가지는데, 이는 임신 기간 동안 암컷에게서 상승하여 어미에게 더 큰 부담을 준다.[4]6. 행동
대부분의 곡비원류는 야행성이지만, 마다가스카르에는 아이아이, 생쥐여우원숭이, 솜털여우원숭이, 스포츠여우원숭이처럼 야행성인 종과 고리꼬리여우원숭이와 그 친척, 시파카, 인드리처럼 주행성인 종이 있다. 일부 갈색 여우원숭이(''Eulemur'')는 주야 활동성을 보여, 온도 및 포식 등의 요인에 따라 낮이나 밤에 활동한다.[8] 사회적 행동은 종에 따라 다양하여, 고독하게 먹이를 찾는 종부터 복잡한 사회적 조직을 보이는 종까지 있다. 털두루원숭이처럼 일부일처 번식 쌍으로 생활하기도 한다.[8] 주행성과 연일활동성 여우원숭이는 원숭이와 유사하게 다수 수컷/다수 암컷 집단으로 진화했으며, 인드리과와 여우원숭이과에서 이러한 사회적 특성이 독립적으로 나타났다.[8] 암컷 우위는 여우원숭이에게 꽤 흔하다.[8]
의사소통에는 냄새 표시가 많은 부분을 차지하며, 표피 냄새샘 분비물을 나무 가지에 바르거나 소변, 대변을 이용한다. 소변 세척을 하는 경우도 있다.[8] 몸 자세와 몸짓도 사용되지만, 긴 주둥이, 움직이지 않는 입술, 감소된 안면 신경으로 인해 얼굴 표정 사용은 제한적이다. 단거리 및 장거리 호출, 경보 신호도 사용되며,[8] 야행성 종은 빛 부족으로 인해 영역 주장을 위한 장거리 호출을 사용하기도 한다.[8] 곡비원류는 상호 그루밍을 통해 서로를 돌보며, 여우원숭이형 영장류는 털을 핥고 이빨 빗으로 빗고, 그루밍 발톱으로 입이 닿지 않는 곳을 긁는다.[8]
6. 1. 이동 방식
대부분의 곡비원류는 나무 위에서 생활하며, 네 다리로 걷거나 뛰는 방식으로 이동한다.[8] 난쟁이여우원숭이는 나무 위에서 네 다리로 걷는 수목성 사지보행을 한다. 갈라고와 인드리과는 수직으로 매달리거나 뛰어오르는 데 특화되어 있다.[8] 특히 인드리과는 수직 매달리기와 도약에 매우 특화되어 있다.[8] 반면, 로리스는 느리고 신중하게 기어오른다.[8] 고리꼬리여우원숭이는 예외적으로 땅에서 많은 시간을 보낸다.[8]멸종된 아다피형류의 경우, 유럽의 ''고원숭이''(Adapis)와 ''Leptadapis''는 로리스처럼 느리게 기어올랐을 것으로 보이며, 작은 신세계원숭이처럼 네 다리로 달렸을 수도 있다. 북미의 ''Notharctus''와 ''Smilodectes'', 유럽의 ''Europolemur''는 수직 매달리기와 도약을 하는 여우원숭이와 유사했지만, 인드리과만큼 특화되지는 않아 나뭇가지를 따라 달리면서 도약을 덜 했을 것으로 보인다.[8] 노타르키드 ''Cantius''와 ''Pronycticebus''는 갈색 여우원숭이와 유사한 민첩한 수목성 사지보행을 보였을 것으로 추정된다.[8]
6. 2. 식성
곡비원류는 과일(씨앗 포함), 잎(꽃 포함), 동물(절지동물, 작은 척추동물, 알) 등 다양한 먹이를 먹는다.[8] 일부 종은 특정 먹이에 특화되어 있는데, 예를 들어 포크마크 여우원숭이와 바늘발톱갈라고는 나무 수액을, 인드리과, 스포츠여우원숭이, 대나무여우원숭이는 잎을 주로 먹는다.[8] 갈라고, 늘씬로리스, 앙와티보는 주로 곤충을 먹으며,[8] 꼬리여우원숭이와 쥐여우원숭이는 과일, 잎, 동물을 섞어 먹는 잡식성이다.[8]아다피스류 중에서는 ''Cantius'', ''Pelycodus'', ''Cercamonius''와 같이 중간 크기에서 큰 종들은 주로 과일을 먹었으며,[8] ''Smilodectes'', ''Notharctus'', ''Adapis'', ''Leptadapis'' 등은 잎을 주로 먹었다.[8] ''Anchomomys'' 및 ''Donrussellia''와 같이 작은 아다피스류는 곤충을 먹었을 것으로 추정된다.[8]
7. 분포 및 서식지
곡비원류는 적도 아프리카와 동남아시아에서 발견되는 로리스와 사하라 이남 아프리카의 숲과 숲에 제한되어 있는 갈라고, 그리고 마다가스카르에 고유종으로 서식하는 여우원숭이로 나뉜다. 이들은 위도 28° S에서 26° N 사이의 열대 지역에 분포한다.[8]
거의 모든 영장류와 마찬가지로 곡비원류는 일반적으로 열대 우림에 서식한다. 마다가스카르 동부 우림에서는 11~12종이 같은 숲을 공유하며, 인간이 도착하기 전에는 일부 숲에서 그 두 배에 가까운 다양성을 보였다. 몇몇 종의 여우원숭이는 섬 남단의 가시 숲을 포함하여 더 건조하고 계절적인 숲에서 발견되지만, 이러한 지역의 여우원숭이 군집은 그만큼 풍부하지 않다.
8. 보존
원숭이하목류는 인간의 활동, 특히 열대 지역의 산림 벌채로 인해 멸종 위험이 높다. 서식지 대부분이 농업 및 목초지 등으로 전환되었다.[1] 원숭이하목류 영장류가 직면한 위협은 서식지 파괴, 사냥(부시미트 또는 전통 의학을 위해), 수출 또는 지역 애완동물 거래를 위한 생포 등으로 나뉜다. 사냥은 종종 금지되어 있지만, 법률은 거의 시행되지 않는다. 마다가스카르에서는 ''파디''(fady)라고 알려진 지역 금기가 때때로 여우원숭이 종을 보호하는 데 도움이 되지만, 일부는 여전히 전통 의학을 위해 사냥된다.[2]
2012년, 국제 자연 보전 연맹(IUCN)은 여우원숭이가 "가장 멸종 위기에 처한 포유류"라고 발표했는데, 이는 주로 2009년 정치 위기 이후 불법 벌목과 사냥이 증가했기 때문이다.[3] 동남아시아에서 천천히 로리스는 서식지 파괴 외에도 애완동물 거래와 전통 의학에 의해 위협받고 있다.[4][5] 여우원숭이와 천천히 로리스는 모두 CITES 부록 I에 따라 상업적 국제 거래로부터 보호받고 있다.[6]
참조
[1]
웹사이트
Strepsirrhini
https://paleobiodb.o[...]
2021-07-22
[2]
LSJ
στρέψις
null
[3]
문서
Greek [[rho]] ⟨ρ⟩ ''r'' is written with a [[spiritus asper]] when word-initial, i.e. ⟨ῥ⟩ ''rh'', and often gets [[gemination|doubled]] to ⟨ῤῥ⟩ ''rrh'' (also spelled ⟨ρρ⟩) when such a word constitutes the second part of a [[compound word]]. See [[Ancient Greek phonology#Liquids]], [[Ancient Greek phonology#Doubled consonants|#Doubled consonants]] and [[Ancient Greek phonology#Consonant spelling|#Consonant spelling]].
null
[4]
논문
Comparative morphology of the eye in primates
2004
[5]
간행물
サルの分類名(その7:総説とメガネザル)
https://doi.org/10.2[...]
日本霊長類学会
1989
[6]
웹사이트
日本モンキーセンター 霊長類和名リスト 2018年11月版
http://www.j-monkey.[...]
日本モンキーセンター霊長類和名編纂ワーキンググループ
2018-12-16
[7]
간행물
世界哺乳類標準和名目録
https://doi.org/10.1[...]
2018
[8]
서적
原始的なサルである「原猿類」は、その名のとおり、古いタイプの哺乳類の特徴をもっている。
朝日新聞社
1992
[9]
간행물
サルの分類名(その8:原猿)
https://doi.org/10.2[...]
日本霊長類学会
1989
[10]
논문
Vitamin C biosynthesis in prosimians: evidence for the anthropoid affinity of Tarsius
1987-05
[11]
간행물
霊長類の手の構造: 樹上生活における把握能力の意義
https://doi.org/10.2[...]
日本霊長類学会
2004
[12]
서적
Mammal Species of the World: A Taxonomic and Geographic Reference
http://www.departmen[...]
존스 홉킨스 대학교 출판사
2005
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com