세포질 유동
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
세포질 유동은 세포 내에서 세포질이 움직이는 현상으로, 식물 세포에서 다양한 형태로 나타난다. 회전형, 순환형, 왕복형 등 여러 유형이 있으며, 세포 종류에 따라 다른 양상을 보인다. 세포질 유동은 주로 엽록체의 운동으로 관찰되며, 미오신과 액틴 필라멘트의 상호 작용을 통해 발생한다. 이러한 유동은 영양분 수송, 광합성 효율 증가, 중력 감지 등 다양한 이점을 제공한다. 18세기부터 관찰되었으며, 20세기 중반부터 메커니즘 연구가 활발히 진행되었다.
식물 세포나 점균류의 변형체, 또는 원생 동물인 아메바 등에서 세포질 내부의 여러 가지 운동을 볼 수 있다. 이러한 운동은 원형질 유동이라 불리며, 세포의 생활 활동을 보여주는 지표이다. 이는 ATP 등의 에너지를 사용하여 일어나며, 물 분자 운동에 기반한 브라운 운동과는 다르다.
식물 세포에서 세포질 유동은 주로 엽록체의 운동으로 관찰된다. 미오신 필라멘트는 세포 소기관을 액틴 필라멘트에 연결하며, 미오신 분자가 액틴 필라멘트를 따라 이동하면서 세포질 유체를 밀거나 당겨 세포질 흐름을 유발한다.[8] 세포질 흐름 속도는 초당 1~100 마이크로미터 범위일 수 있다.[8][9]
2. 원형질 유동의 양상
원형질 유동은 방향성을 가지는데, 세포막을 따라 흐르거나 부분적으로 순환하기도 하고, 일단 정지했다가 반대 방향으로 흐르는 듯한 운동(변형체의 왕복 운동)도 있다. 유동하는 것은 졸 부분이며, 방향 결정이나 운동을 일으키는 구조에는 세포막, 핵, 엽록체 등의 겔 부분이 관여한다. 아메바 운동에서는 바깥쪽(주변부)의 겔과 안쪽의 졸이 상호 변화하며, 원형질 유동은 원형질 내 물질 수송뿐만 아니라 세포 자체의 이동에도 기여한다.
2. 1. 회전형
액포가 발달한 식물 세포에서 전형적으로 나타나는 유동으로, 세포질은 세포막과 액포막 사이의 영역을 한 방향으로 회전한다. 차축조의 절간 세포에서는 유속이 초당 80um 정도이다. 검정말 등에서 흔히 관찰된다.
2. 2. 순환형
액포 내에서 원형질이 가느다란 실처럼 관통하며 순환한다. 자주달개비, 양파 등에서 관찰된다.[1]
2. 3. 왕복형
점균 변형체 등에서 관찰되는, 주기적으로 방향이 바뀌는 유동이다. 2~3분 주기로 왕복하며, 한 방향으로의 최고 유속은 초당 약 1밀리미터에 달한다.
3. 원형질 유동의 메커니즘
원형질 유동에는 방향성이 있어 세포막을 따라 흐르거나 부분적으로 순환하는 운동(예: 자주달개비 수꽃술의 털세포) 외에, 일시 정지 후 반대 방향으로 흐르는 듯한 운동(예: 변형체의 왕복 운동)도 있다. 유동하는 것은 졸 부분이며, 방향 결정이나 운동 유발에는 세포막, 핵, 엽록체 등의 겔 부분이 관여한다. 아메바 운동에서는 바깥쪽(주변부) 겔과 안쪽 졸이 상호 변화하며, 원형질 유동은 물질 수송뿐 아니라 세포 이동에도 기여한다.
3. 1. 차축조 (''Chara corallina'')
말 코랄리나(''Chara corallina'')는 큰 중심 액포 주위로 순환하는 세포질 유동을 보인다.[8] 이 큰 중심 액포는 식물 세포에서 가장 큰 세포 소기관 중 하나이며 주로 저장에 사용된다.[10] ''말 코랄리나'' 세포는 최대 10cm 길이, 1mm 직경까지 자랄 수 있다.[8] 액포의 직경은 세포 직경의 약 80%를 차지할 수 있다.[11] 따라서 직경 1mm인 세포의 경우 액포 직경은 0.8mm가 될 수 있으며, 액포 주변 세포질이 흐를 수 있는 경로는 약 0.1mm에 불과하다. 세포질은 초당 100 마이크로미터의 속도로 흐르는데, 이는 알려진 모든 세포질 흐름 현상 중 가장 빠르다.[8]
''Chara corallina'' 세포 내 세포질 흐름은 엽록체의 "이발사 기둥" 운동으로 관찰된다.[8] 현미경으로 관찰하면 엽록체 흐름에는 두 구역이 있다. 이 구역들은 세포의 종축을 따라 나선형으로 배열된다.[8] 한 구역에서는 엽록체가 나선의 한 띠를 따라 위쪽으로 움직이고, 다른 구역에서는 아래쪽으로 움직인다.[8] 이 구역들 사이 영역을 무관심 구역이라고 한다. 엽록체는 이 구역을 절대 넘지 않는 것으로 관찰되었으며,[8] 그 결과 세포질과 액포 내 유체 흐름도 이와 유사하게 제한될 것이라고 생각했지만, 이는 사실이 아니다.
먼저, Kamiya와 Kuroda는 세포질 유속이 세포 내에서 반경 방향으로 변화한다는 것을 실험적으로 밝혀냈는데, 이는 엽록체의 움직임으로는 명확하게 나타나지 않는다.[12] 둘째, 레이몬드 골드스타인(Raymond Goldstein) 등은 세포질 흐름에 대한 수학적 유체 모델을 개발하여, Kamiya와 Kuroda가 관찰한 현상을 예측할 뿐만 아니라, 무관심 구역을 통과하는 세포질 흐름 궤적까지 예측했다.[8] 골드스타인 모델은 액포막을 무시하고, 전단력이 세포질로부터 액포 내 유체로 직접 전달된다고 가정한다.
골드스타인 모델은 순 흐름이 한 무관심 구역에서 다른 구역으로 향한다고 예측한다.[8] 이는 실제로 엽록체의 흐름에서 관찰된다. 한 무관심 구역에서는 아래쪽으로 기울어진 엽록체가 위쪽으로 기울어진 엽록체 위에 위치한다. 이 구역을 마이너스 무관심 구역(IZ-)이라고 한다. 여기서 각 방향을 세타(수평) 및 z(수직) 방향 성분으로 분해하면, 이 성분들의 합이 z 방향에서는 서로 반대 방향으로 작용하고, 세타 방향에서는 발산한다.[8] 다른 무관심 구역은 위쪽으로 기울어진 엽록체 운동이 위에 있으며, 이를 플러스 무관심 구역(IZ+)이라고 한다. 따라서 z 방향 성분은 다시 서로 반대 방향으로 작용하지만, 세타 성분은 수렴한다.[8] 이 힘들의 순 효과는 세포질/액포 유동이 마이너스 무관심 구역에서 플러스 무관심 구역으로 이동하는 것이다.[8]
이러한 방향 성분은 엽록체 운동으로 관찰되지만 명확하지는 않다. 세포질/액포 유동이 한 무관심 구역에서 다른 구역으로 이동하면, 표면의 엽록체는 그렇지 않더라도 세포질 입자가 무관심 구역을 가로지른다. 입자는 세포 내에서 상승하면서 마이너스 무관심 구역 근처에서 반원형으로 나선형을 그리며, 한 무관심 구역을 지나 플러스 무관심 구역 근처에서 끝난다.[8]
Characean 세포에 대한 추가 실험은 액포 유체 흐름에 대한 골드스타인 모델을 뒷받침한다.[11] 그러나 (골드스타인 모델에서 무시된) 액포막 때문에, 세포질 흐름은 다른 흐름 패턴을 따른다. 또한, 최근 실험에서는 세포질 내 속도가 균일하다는 것을 시사했던 Kamiya와 Kuroda가 수집한 데이터가 완전히 정확하지 않다는 것을 보여주었다.[11] Kikuchi는 ''Nitella flexillis'' 세포를 가지고 연구하여 유체 흐름 속도와 세포막으로부터의 거리 사이에 지수 관계가 있음을 발견했다.[11] 이 연구는 ''Chara'' 세포에 대한 것이 아니지만, ''Nitella flexillis''와 ''Chara coralina'' 사이의 흐름은 시각적으로나 구조적으로 유사하다.[11]
3. 2. 점균 (''Physarum polycephalum'')
점균으로 비공식적으로 불리는 단세포 원생생물 ''Physarum polycephalum'' 내 미오신과 액틴 분자에 대한 생물학적 연구는 인간 근육의 미오신 및 액틴 분자와 놀라운 물리적, 기계적 유사성을 보여주었다. 이 분자들의 수축과 이완은 세포 길이에 따라 압력 기울기를 유발하며, 세포질 유동을 한 방향으로 밀어내어 성장에 기여한다.[16] 인간의 분자와 유사하지만, 미오신이 액틴에 결합하는 부위를 막는 분자는 다르다는 것이 입증되었다. 인간의 경우 트로포미오신이 해당 부위를 덮어 칼슘 이온이 존재할 때만 수축이 가능하게 하지만, 이 아메바에서는 칼모듈린이라는 다른 분자가 해당 부위를 막아 높은 칼슘 이온 농도에서 이완을 가능하게 한다.[16]
3. 3. 누룩곰팡이 (''Neurospora crassa'')
누스포라 크라싸는 여러 개의 뻗어 나가는 균사를 가진 다세포 곰팡이이다. 세포는 최대 10cm까지 길 수 있으며, 작은 격막으로 구분된다.[17] 격막의 작은 구멍을 통해 세포질과 세포질 내용물이 세포에서 세포로 흐를 수 있다. 삼투압 구배는 세포의 길이를 따라 발생하여 이러한 세포질 흐름을 유도한다. 흐름은 성장과 세포 하위 구획의 형성에 기여한다.[17][18]
세포질 유동은 삼투압 구배를 통해 생성되어 균사체를 따라 종방향으로 흐르다가 끝에 부딪혀 성장을 유발한다. 균사체 끝 부분의 압력이 클수록 성장 속도가 빠르다는 것이 입증되었다. 균사체가 길수록 길이 방향의 압력 차이가 커서 세포질 유동 속도가 빨라지고 균사체 끝 부분의 압력이 커진다.[17] 이것이 더 긴 균사체가 더 짧은 균사체보다 빨리 자라는 이유이다. 끝 부분의 성장은 세포질 유동 속도가 증가함에 따라 24시간 동안 증가하며, 최대 1마이크론/초의 성장 속도가 관찰된다.[17] 주 균사체에서 갈라져 나온 부분은 더 짧고 세포질 유동 속도가 느리며, 이에 따라 성장 속도도 느리다.[17]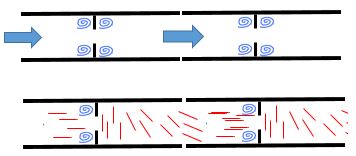
뉴로스포라 크라사(Neurospora crassa)의 세포질 유동은 미세소관을 운반한다. 미세소관의 존재는 유동에 흥미로운 측면을 만들어낸다. 곰팡이 세포를 중앙에 구멍이 있는 격막으로 정기적으로 분리된 파이프로 모델링하면 매우 대칭적인 유동이 생성되어야 한다. 기본적인 유체 역학은 각 격막의 전후에 소용돌이가 형성되어야 한다고 제안한다.[19] 그러나 뉴로스포라 크라사(Neurospora crassa)에서는 소용돌이가 격막 전에만 형성된다. 이는 미세소관이 격막 구멍으로 들어가면 유동에 평행하게 정렬되어 유동 특성에 거의 기여하지 않지만, 격막 구멍을 나가면 유동에 수직으로 정렬되어 가속을 늦추고 소용돌이 형성을 방지하기 때문이다.[17] 격막 바로 전에 형성된 소용돌이는 특수 단백질로 점철된 핵이 모이는 하위 구획의 형성을 가능하게 한다.[17] 이 단백질 중 하나인 SPA-19는 격막 유지에 기여한다. 그것이 없으면 격막이 분해되어 세포가 이웃 세포로 많은 양의 세포질을 누출하여 세포 사멸을 초래할 것이다.[17]
3. 4. 생쥐 난모세포
중심립과 방추사는 세포 분열, 감수 분열 등 여러 과정에서 세포 내 핵을 중앙에 위치시키는 역할을 한다. 하지만 생쥐 난모세포는 중심립이 없음에도 세포질 유동을 통해 핵을 중앙에 위치시킨다.[20]
미세 섬유는 미세 소관, 미오신 2와는 독립적으로 세포 전체에 망상 네트워크를 형성한다. 중심에서 벗어난 핵은 25마이크론 이상 떨어진 거리에서 세포 중심으로 이동하지만, 이 네트워크가 존재하면 6마이크론 이상 벗어나지 않는다.[20] 미세 섬유 네트워크는 미오신 Vb 분자에 의해 세포 소기관과 결합되어 있다.[20] 세포질 유체는 세포 소기관의 움직임에 따라 함께 이동하지만, 방향성은 없다. 이러한 움직임은 브라운 운동의 특징을 보인다.
세포 소기관의 방향성 있는 움직임은 세포질이 세포를 채우는 구형 구조 때문에 나타난다. 구의 반지름이 커질수록 표면적도 증가하므로, 중심을 향한 움직임이 중심에서 멀어지는 움직임보다 커서 핵을 세포 중심으로 미는 순수한 힘이 발생한다. 즉, 세포질 입자의 무작위적인 움직임이 세포 중심을 향하는 순 힘을 생성한다.[20]
세포질의 증가된 움직임은 세포질 점성을 낮춰 핵이 세포 내에서 더 쉽게 움직이게 한다. 이러한 세포질 유동의 두 가지 요인이 난모세포에서 핵을 중앙에 위치시키는 역할을 한다.[20]
4. 원형질 유동의 이점
원형질 유동은 식물 세포, 점균류의 변형체, 아메바 등에서 관찰되는 세포질 내부의 운동이다.[1] 이는 세포의 생활 활동을 보여주는 현상으로, ATP 등의 에너지를 사용하여 일어난다. 물 분자 운동에 의한 브라운 운동과는 다르다.
원형질 유동은 특정한 방향성을 가진다. 세포막을 따라 흐르거나, 부분적으로 순환하는 운동(자주달개비 수꽃술의 털세포) 외에도, 일시 정지 후 반대 방향으로 흐르는 듯한 운동(변형체의 왕복 운동)도 있다. 유동하는 것은 졸(sol) 상태의 세포질이며, 세포막, 핵, 엽록체 등의 겔(gel) 상태 구조물이 운동 방향과 발생에 관여한다. 아메바 운동에서는 바깥쪽(주변부)의 겔과 안쪽의 졸이 서로 변환되기도 한다.
원형질 유동은 세포 내 물질 수송뿐만 아니라 세포 자체의 이동에도 기여한다.
4. 1. 영양분 수송 증진
세포질 유동으로 인해 발생하는 복잡한 유동 궤적은 액포 막을 가로지르는 농도 기울기를 크게 만들어 확산을 통한 영양분 수송을 증진시킨다.[8] 피크의 확산 법칙에 따르면, 더 큰 농도 기울기는 더 큰 확산 흐름을 유발한다.[13] 따라서, ''Chara coralina''의 세포질 흐름의 독특한 유동 궤적은 확산을 통해 저장 액포로의 영양분 수송을 증진시킨다.골드스타인은 세포질 흐름이 빠를수록 더 큰 농도 기울기가 발생하고, 저장 액포로의 더 큰 확산 영양분 수송이 발생한다는 것을 증명했다. 액포로의 향상된 영양분 수송은 성장률과 전반적인 성장 크기에 현저한 차이를 가져온다.[9]
애기장대(Arabidopsis thaliana)를 이용한 실험에서, 야생형 식물은 ''Chara coralina''와 유사하게 유체를 포획하여 세포질 유동을 나타내지만, 유속은 더 느리다.[9] 야생형 미오신 운동 분자를 더 빠른 미오신 분자로 대체한 결과 세포질 흐름 속도가 증가했다. 반면, 느린 인간 미오신 Vb로 대체한 식물은 세포질 유동이 거의 나타나지 않았다.
이후 유사한 환경에서 식물을 키운 결과, 더 빠른 세포질 유동 속도를 보인 식물이 더 크고 풍성한 잎을 가진 더 큰 식물로 성장했다.[9] 이는 골드스타인 모델에서 증명된 바와 같이, 향상된 영양분 저장이 식물이 더 크고 빠르게 성장하도록 한다는 것을 보여준다.[8][9]
4. 2. 광합성 효율 증가
광합성은 빛 에너지를 아데노신 삼인산(ATP) 형태로 바꾸는 화학 에너지이다.[14] 광합성은 식물 세포의 엽록체에서 발생한다. 빛 광자는 엽록체의 다양한 막간 단백질과 상호 작용하여 빛 에너지를 화학 에너지로 바꾼다. 그러나 이러한 단백질은 광자에 포화되어 포화가 풀릴 때까지 제 기능을 수행할 수 없게 된다. 이것을 카우스키 효과라고 부르며, ATP 생산 메커니즘이 비효율적인 원인이 된다. 그러나 ''말 코랄리나''(Chara corallina)의 세포질 유동은 엽록체가 식물의 줄기 주위로 이동할 수 있게 한다. 따라서 엽록체는 밝은 영역과 그늘진 영역으로 이동한다.[14] 이러한 세포질 유동으로 광자에 간헐적으로 노출되면 실제로 엽록체의 광합성 효율이 증가한다.[14] 광합성 활동은 보통 클로로필 형광 분석을 사용하여 평가한다.4. 3. 중력 감지
차축조 식물은 세포질 유동 속도의 차이를 이용하여 중력을 감지한다.[15] 세포질 유동으로 인해 엽록체는 한쪽 방향으로는 위로, 다른 한쪽 방향으로는 아래로 흐르는 운동을 보인다.[8] 이때 아래쪽으로 움직이는 엽록체의 운동이 위쪽으로 흐르는 것보다 약간 더 빠르며, 이 속도의 비율(극성 비율)은 1.1 정도이다.[8][15] 이 극성 비율은 중력의 힘에 따라 달라진다.[15]이러한 속도 증가는 중력의 힘이 직접적으로 작용한 결과가 아니라 간접적인 결과이다. 중력은 식물 원형질이 세포벽 내에 침전되도록 한다. 이에 따라 세포막은 위쪽에서는 인장력을, 아래쪽에서는 압축력을 받는다. 세포막에 가해지는 이러한 압력의 차이가 ''말 코랄리나''(Chara corallina)에서 관찰되는 세포질 흐름 속도의 차이를 유발하며, 이를 통해 중력을 감지하게 된다.[15] 이러한 중력 감지 이론은 아밀로플라스트 침전에 의해 나타나는 정지립 이론과 정반대된다.[15]
5. 연구사
1772년, 이탈리아의 보나벤투라 코르티는 현미경을 이용해 샤지쿠모의 세포 내용물이 순환 운동을 한다는 논문을 발표했지만, 큰 주목을 받지 못했다.[1]
1807년, 독일의 식물학자 루돌프 트레비라누스는 이 현상을 재발견했지만, 세포 내 열의 불균등 분포에 따른 대류와 같은 현상이라고 생각했다.[1]
20세기 중반부터 가미야 노리오 등의 샤지쿠모와 점균을 이용한 연구를 통해 이 현상의 발생 기작이 밝혀지기 시작했다.[1]
1956년, 가미야 등은 원형질 유동은 원형질의 졸=겔 계면에서의 능동적인 "미끄럼"에 의해 발생한다고 하는 '''미끄럼설'''을 제창했다.[1]
1974년 샤지쿠모류에서 액틴 필라멘트가, 1994년에는 차축조에서 미오신이 동정되어 유동력이 액틴과 미오신의 상호 작용에 의한 것이라는 기작이 입증되었다.[1]
참조
[1]
논문
A physical perspective on cytoplasmic streaming
2015-08
[2]
서적
The Physiology of Characean Cells
Springer Science & Business Media
2013-11-19
[3]
논문
Cytoplasmic streaming in plant cells emerges naturally by microfilament self-organization
2013-08
[4]
논문
Cytoplasmic streaming in plants
2004-02
[5]
논문
Effects of cyclosis on chloroplast-cytoplasm interactions revealed with localized lighting in Characean cells at rest and after electrical excitation
2011-09
[6]
논문
A physical perspective on cytoplasmic streaming
2015-08
[7]
논문
A cytochalasin-sensitive actin filament meshwork is a prerequisite for local wound wall deposition in Nitella internodal cells
1997-03
[8]
논문
Microfluidics of cytoplasmic streaming and its implications for intracellular transport
2008-03
[9]
논문
Cytoplasmic streaming velocity as a plant size determinant
2013-11
[10]
논문
Structure, function, and motility of vacuoles in filamentous fungi
1998-06
[11]
논문
Diffusive Promotion by Velocity Gradient of Cytoplasmic Streaming (CPS) in Nitella Internodal Cells
2015
[12]
논문
Velocity Distribution of the Protoplasmic Streaming in ''Nitella'' Cells
1956
[13]
서적
The Principles of Engineering Materials
Prentice-Hall
1973
[14]
논문
Effect of Cytoplasmic Streaming on Photosynthetic Activity of Chloroplasts in Internodes of ''Chara Corallina''
[15]
논문
The effect of the external medium on the gravity-induced polarity of cytoplasmic streaming in Chara corallina (Characeae)
1997-11
[16]
논문
Calcium wave for cytoplasmic streaming of Physarum polycephalum
2009-12
[17]
논문
Cellular Subcompartments through Cytoplasmic Streaming
2015-08
[18]
논문
How does a hypha grow? The biophysics of pressurized growth in fungi
https://zenodo.org/r[...]
2011-06
[19]
서적
Fluid Mechanics
McGraw Hill
[20]
논문
Active diffusion positions the nucleus in mouse oocytes
https://resolver.sub[...]
2015-04
[21]
논문
A physical perspective on cytoplasmic streaming
2015-08
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com