스쿠토사우루스
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
스쿠토사우루스는 후기 페름기에 살았던 멸종된 파레이아사우루스과의 파충류이다. 러시아에서 화석이 처음 발견되었으며, 단단한 골판으로 덮인 몸과 뾰족한 뿔이 특징이다. 스쿠토사우루스는 초식성이었으며, 2.5~3m의 크기로 자랐다. 연구를 통해, 스쿠토사우루스는 육상 생활을 했을 가능성이 높으며, 다른 페름기 동물들과 함께 살았다.

더 읽어볼만한 페이지
- 페름기의 동물 - 아르트로플레우라
아르트로플레우라는 석탄기와 페름기에 번성했던 멸종된 다지류로, 넓은 등판과 3개의 엽을 가진 외형을 특징으로 하며, 육상에서 생활하며 초식 또는 부식성 식성을 가졌을 것으로 추정된다. - 페름기의 동물 - 바나롭스
바라놉스는 1911년 텍사스에서 발견된 멸종된 대형 양막류 속으로, 날카로운 이빨과 큰 사지를 가진 약 1.2미터 길이의 페름기 육상 포식자이며, 바라노돈 및 와통기아와 근연 관계이고 바라노돈티나이의 기원이다. - 측파충류 - 메소사우루스
페름기에 서식한 메소사우루스는 유선형 몸체, 긴 꼬리, 물갈퀴 모양 발을 가진 수생 파충류로, 남아프리카와 남아메리카에서 발견된 화석은 대륙 이동설의 증거로 제시되며, 가늘고 긴 턱의 바늘 모양 이빨로 작은 갑각류나 플랑크톤을 섭취했을 것으로 추정된다. - 측파충류 - 중룡목
중룡목은 멸종된 해양 파충류 무리로, 파충류 계통 분류에서 논쟁의 대상이 되었으며, 얕은 물에서 서식하며 갑각류를 섭취하고 태생을 했을 가능성이 있다. - 1930년 기재된 화석 분류군 - 마크로플라타
마크로플라타는 쥐라기 초기에 서식한 로말레오사우루스과의 플레시오사우루스로, 독특한 형태의 두개골 뼈들과 26개의 경추를 가진 것이 특징이다. - 1930년 기재된 화석 분류군 - 만추로사우루스
만추로사우루스는 백악기 후기에 살았던 하드로사우루스과 공룡으로, 1914년 아무르강 유역에서 발견된 화석을 바탕으로 1953년 명명되었으며, 몸길이 약 8m, 키 약 4.5m로 추정되는 만추로사우루스 아무렌시스 한 종이 인정된다.

2. 연구사
''스쿠토사우루스''는 북드비나 유적지에서 흔한 화석이며, 최소 6개의 비교적 완전한 골격과 수많은 다양한 고립된 신체 및 두개골 유해, 그리고 피골로 알려져 있어 가장 완전하게 알려진 파레이아사우루스이다. 모든 ''스쿠토사우루스'' 표본은 후기 타타르 시대(비아트키안) 러시아 동물상 단계로 거슬러 올라가며,[7] 이는 후기 페름기의 로핑기안 시대(2억 5900만 년~2억 5200만 년 전)와 대략 일치한다.[9][10] 1996년, 러시아 고생물학자 발레리 K. 골루베프는 해당 유적지의 동물상 구역을 설명하며, ''스쿠토사우루스'' 구역이 초기 우차피안에서 시작된 "''Proelginia''" 단계를 따라 대략 중간 우차피안에서 중간 창싱기안까지 확장된다고 언급했다.[11][12]
2. 1. 초기 연구 및 명명
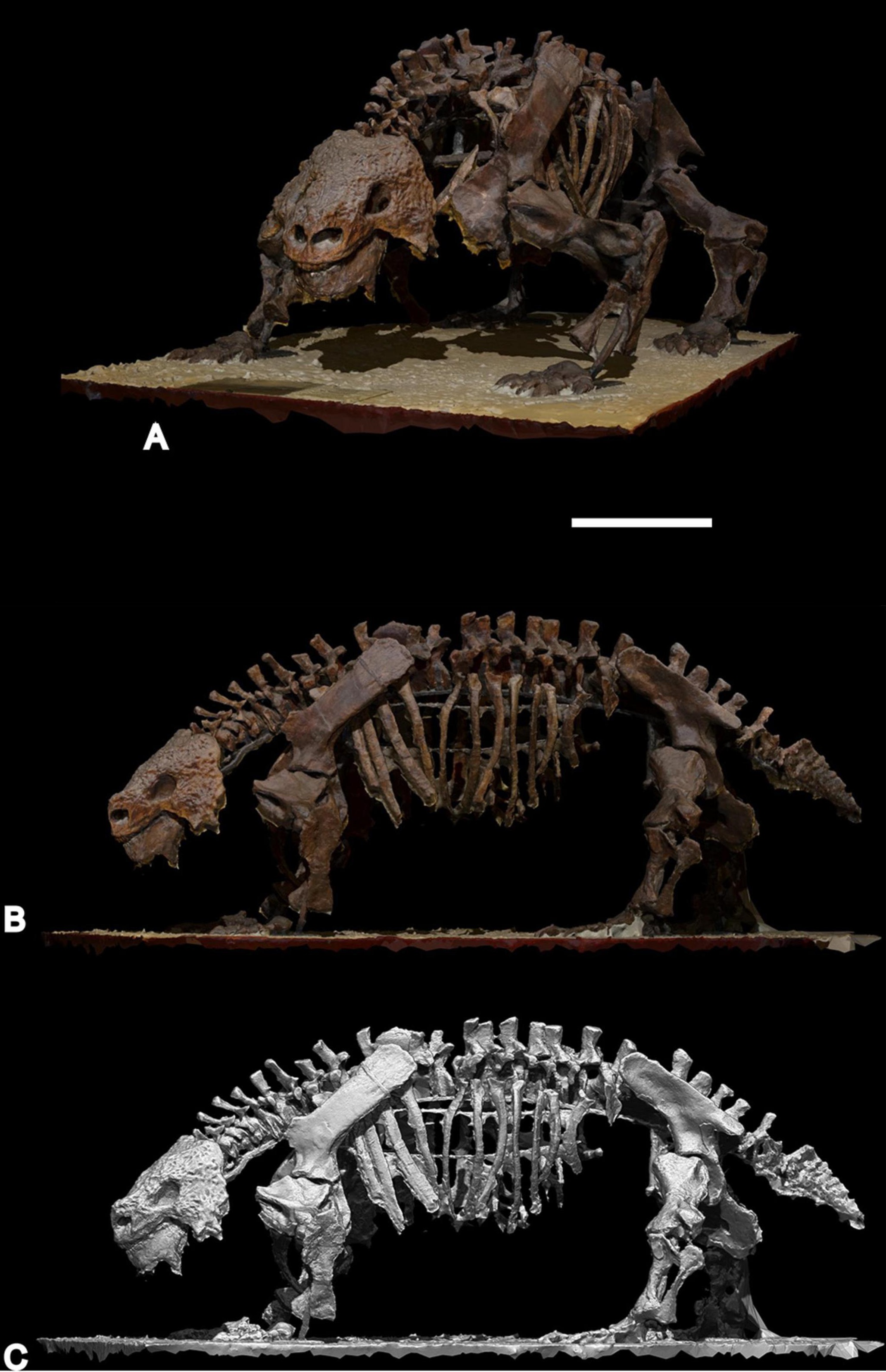
최초의 화석은 러시아 고생물학자 블라디미르 프로코로비치 아말리츠키가 북드비나강, 아르한겔스크주 북부 유럽 러시아의 후기 페름기 퇴적층에서 동식물 종을 기록하면서 발견했다. 아말리츠키는 1899년에 해당 유적지를 발견했으며, 그와 그의 아내 안네 아말리츠키는 1914년까지 발굴 작업을 감독하며 다양한 동물들의 거의 완전하고 관절이 연결된(자연적인 위치에 있는) 수많은 골격을 발굴했다.[2] 이 표본들에 대한 공식적인 진단은 제1차 세계 대전으로 인해 지연되었다.[8] 현재 ''스쿠토사우루스 카르핀스키''라고 불리는 종의 최초 이름은 1917년에 영국 동물학자 데이비드 메러디스 시어스 왓슨에 의해 발표되었으며, 그는 보존 상태가 좋지 않은 표본 PIN 2005/1535를 기반으로 한 견갑골상완골 재구성에 "''Pariasaurus Karpinskyi'', Amalitz"라는 설명을 붙였다(이 이름에 대한 공로는 아말리츠키에게 돌렸다).[3] 아말리츠키는 그 해 말에 사망했고, 동물의 실제 진단은 사후에 1922년에 발표되었으며, 이름은 "''Pareiosaurus''" ''karpinskii''로,[2] 모식 표본은 거의 완전한 골격 PIN 2005/1532로 지정되었다.[7] 세 개의 부분적인 두개골도 발견되었지만, 아말리츠키는 이들을 "''P. elegans''", "''P. tuberculatus''", "''P. horridus''"라는 새로운 종으로 분류하기로 결정했다.[2]
"''Pariasaurus''"와 "''Pareiosaurus''"는 모두 남아프리카의 ''파레이아사우루스''의 오기였다.[8] 1930년, 소비에트 척추 고생물학자 알렉산드라 파울리노브나 안나 하트만-바인베르크는 북드비나에서 발견된 파레이아사우루스 자료는 단 1종만을 나타내며, 이 종은 다른 ''파레이아사우루스''와 구별되어 새로운 속으로 분류될 만하다고 말했다. 아말리츠키는 독특한 속명 "''Pareiosaurus''"를 사용했지만, 이는 실수였으며, 그녀는 "''Pareiosaurus''"가 ''파레이아사우루스''의 주니어 동의어라고 선언하고 ''스쿠토사우루스''라는 속을 만들었다. 그녀는 종명으로 "''karpinskyi''"의 철자를 사용했지만,[4] 1937년에 ''karpinskii''로 변경했다. 동시에 그녀는 부분적인 두개골 PIN 156/2를 기반으로 또 다른 독특한 속 "''Proelginia permiana''"을 분리했다.[5] 1968년, 러시아 고생물학자 N. N. 칼란다제와 그의 동료들은 "''Proelginia''"을 ''스쿠토사우루스''의 분류학적 동의어로 간주했다.[6] 유해의 보존 상태가 좋지 않기 때문에, "''Proelginia''"의 유효성은 불분명하다. 1987년, 러시아 고생물학자 미하일 페오도시예비치 이바흐넨코는 두개골 조각 PIN 3919를 기반으로 새로운 종 "''S. itilensis''"를 만들고 "''S. tuberculatus''"를 부활시켰지만, 호주 생물학자 마이클 S. Y. 리는 2000년에 이 두 가지 조치가 정당하지 않다고 여겼다.[7] 2001년, 리는 국제동물명명규약위원회에 ''karpinskyi''의 철자를 공식적으로 무효화하고(왓슨은 자신의 연구가 이 종의 공식적인 설명이 될 의도가 없었고, ''karpinskii''가 훨씬 더 널리 사용되었기 때문에) 저자 인용을 Amalitskii, 1922로 기록해 달라고 청원했다.[8]
2. 2. 스쿠토사우루스속의 확립
최초의 화석은 러시아 고생물학자 블라디미르 프로코로비치 아말리츠키가 북드비나강, 아르한겔스크주 북부 유럽 러시아의 후기 페름기 퇴적층에서 동식물 종을 기록하면서 발견했다. 아말리츠키는 1899년에 해당 유적지를 발견했으며, 그와 그의 아내 안네 아말리츠키는 1914년까지 발굴 작업을 감독하며 다양한 동물들의 거의 완전하고 관절이 연결된(자연적인 위치에 있는) 수많은 골격을 발굴했다.[2] 이 표본들에 대한 공식적인 진단은 제1차 세계 대전으로 인해 지연되었다.[8]
현재 ''스쿠토사우루스 카르핀스키''라고 불리는 종의 최초 이름은 1917년에 영국 동물학자 데이비드 메러디스 시어스 왓슨에 의해 발표되었으며, 그는 보존 상태가 좋지 않은 표본 PIN 2005/1535를 기반으로 한 견갑골상완골 재구성에 "''Pariasaurus Karpinskyi'', Amalitz"라는 설명을 붙였다(이 이름에 대한 공로는 아말리츠키에게 돌렸다).[3] 아말리츠키는 그 해 말에 사망했고, 동물의 실제 진단은 사후에 1922년에 발표되었으며, 이름은 "''Pareiosaurus''" ''karpinskii''로,[2] 모식 표본은 거의 완전한 골격 PIN 2005/1532로 지정되었다.[7] 세 개의 부분적인 두개골도 발견되었지만, 아말리츠키는 이들을 "''P. elegans''", "''P. tuberculatus''", "''P. horridus''"라는 새로운 종으로 분류하기로 결정했다.[2]
"''Pariasaurus''"와 "''Pareiosaurus''"는 모두 남아프리카 공화국의 ''파레이아사우루스''의 오기였다.[8] 1930년, 소비에트 척추 고생물학자 알렉산드라 파울리노브나 안나 하트만-바인베르크는 북드비나에서 발견된 파레이아사우루스 자료는 단 1종만을 나타내며, 이 종은 다른 ''파레이아사우루스''와 구별되어 새로운 속으로 분류될 만하다고 말했다. 아말리츠키는 독특한 속명 "''Pareiosaurus''"를 사용했지만, 이는 실수였으며, 그녀는 "''Pareiosaurus''"가 ''파레이아사우루스''의 주니어 동의어라고 선언하고 ''스쿠토사우루스''라는 속을 만들었다. 그녀는 종명으로 "''karpinskyi''"의 철자를 사용했지만,[4] 1937년에 ''karpinskii''로 변경했다.
동시에 그녀는 부분적인 두개골 PIN 156/2를 기반으로 또 다른 독특한 속 "''Proelginia permiana''"을 분리했다.[5] 1968년, 러시아 고생물학자 N. N. 칼란다제와 그의 동료들은 "''Proelginia''"을 ''스쿠토사우루스''의 분류학적 동의어로 간주했다.[6] 유해의 보존 상태가 좋지 않기 때문에, "''Proelginia''"의 유효성은 불분명하다.
1987년, 러시아 고생물학자 미하일 페오도시예비치 이바흐넨코는 두개골 조각 PIN 3919를 기반으로 새로운 종 "''S. itilensis''"를 만들고 "''S. tuberculatus''"를 부활시켰지만, 호주 생물학자 마이클 S. Y. 리는 2000년에 이 두 가지 조치가 정당하지 않다고 여겼다.[7] 2001년, 리는 국제동물명명규약위원회에 ''karpinskyi''의 철자를 공식적으로 무효화하고(왓슨은 자신의 연구가 이 종의 공식적인 설명이 될 의도가 없었고, ''karpinskii''가 훨씬 더 널리 사용되었기 때문에) 저자 인용을 Amalitskii, 1922로 기록해 달라고 청원했다.[8]
''스쿠토사우루스''는 북드비나 유적지에서 흔한 화석이며, 최소 6개의 비교적 완전한 골격과 수많은 다양한 고립된 신체 및 두개골 유해, 그리고 골판(피골)으로 알려져 있다. 그것은 가장 완전하게 알려진 파레이아사우르이다.
3. 해부학적 특징
파레이아사우루스과는 페름기에 살았던 가장 큰 파충류 중 하나였다. 스쿠토사우루스는 몸길이가 약 2.5m, 무게가 최대 1160kg에 달하는 비교적 큰 파레이아사우루스과 동물이다.[46] 몸 전체는 척추뼈 가운데에 돌출부가 있는 거친 골배엽(피부 골판)으로 덮여 있었을 것으로 추정된다. 이 골배엽들은 서로 분리되어 있었지만, 엘기니아처럼 어깨와 골반 부위에서는 서로 밀착되어 봉합되었을 가능성도 있다. 팔다리에는 작은 원뿔 모양의 돌기가 있었다.
파레이아사우루스과는 짧고 튼튼한 몸통과 짧은 꼬리를 가지고 있으며, 스쿠토사우루스는 19개의 천추 앞 척추뼈를 가지고 있다.[47] 등뼈는 짧고 높으며 튼튼하고, 크고 강하게 굽은 늑골을 지탱한다. 넓은 몸통은 큰 소화 기관을 수용했을 것으로 추정된다.[51]
상악골에는 콧구멍 바로 뒤에 뿔이 있다. 구개 뒤쪽의 두 구멍(interpterygoid vacuity)은 크다.[7] 스쿠토사우루스는 위턱에 18개(9~11개의 치관을 가짐), 아래턱에 16개(13~17개의 치관)의 이빨을 가지고 있다. 위턱 이빨 끝은 약간 바깥쪽으로 향하고, 아래턱 이빨의 혀 쪽에는 삼각형 능선이 있으며, 양쪽 턱의 일부 이빨에는 치관이 있는 치경이 있을 수 있다. 다른 파레이아사우루스류와 달리 스쿠토사우루스는 두개골 기저의 결절 사이에 작은 결절을 가지고 있다.[7]
3. 1. 골격 및 외피

파레이아사우루스과는 페름기에 살았던 가장 큰 파충류 중 하나였다. 스쿠토사우루스는 몸길이가 약 2.5m, 무게가 최대 1160kg에 달하는 비교적 큰 파레이아사우루스과 동물이다.[46] 몸 전체는 척추뼈 가운데에 돌출부가 있는 거친 골배엽(피부 골판)으로 덮여 있었을 것으로 추정된다. 이 골배엽들은 서로 분리되어 있었지만, 엘기니아처럼 어깨와 골반 부위에서는 서로 밀착되어 봉합되었을 가능성도 있다. 팔다리에는 작은 원뿔 모양의 돌기가 있었다.
파레이아사우루스과는 짧고 튼튼한 몸통과 짧은 꼬리를 가지고 있으며, 스쿠토사우루스는 19개의 천추 앞 척추뼈를 가지고 있다.[47] 파레이아사우루스를 포함한 페름기 초식성 사지동물들은 공통적으로 큰 몸집, 통 모양의 늑골, 튼튼한 팔다리, 그리고 가슴과 골반이 결합된 특징을 보인다.[48]
파레이아사우루스과의 견갑골은 크고 접시 모양이며, 팔 쪽으로 약간 확장되어 있고 수직 방향으로 향한다. 큰 쇄골과 연결되는 견봉은 초기 거북과 마찬가지로 짧고 뭉툭하며 견갑골 아래쪽에 위치한다. 관절 표본(관절된 뼈의 위치가 보존된 경우)에서는 쇄골과 견갑골 사이에 작은 틈이 있다. 초기 파레이아사우루스과는 견갑골을 따라 뻗어 있는 쇄골을 가졌지만, 스쿠토사우루스를 포함한 후기 표본에서는 이것이 사라졌다.[49] 손과 발의 뼈는 짧다.[50] 등뼈는 짧고 높으며 튼튼하고, 크고 강하게 굽은 갈비뼈를 지탱한다. 넓은 몸통은 큰 소화 기관을 수용했을 것으로 추정된다.[51]
뺨은 바깥쪽으로 강하게 돌출되어 길고 뾰족한 돌기로 끝난다. 두개골의 돌기는 다른 파레이아사우루스류보다 훨씬 두드러진다. 상악골에는 콧구멍 바로 뒤에 뿔이 있다. 구개 뒤쪽의 두 구멍(interpterygoid vacuity)은 크다.[7] 모든 파레이아사우루스류는 빽빽하게 배열된 높고 칼날 모양의 이형치를 가진 넓은 주둥이를 가지고 있으며, 치아와 종에 따라 다양한 수의 치관을 갖는다.[17] 스쿠토사우루스는 위턱에 18개(9~11개의 치관), 아래턱에 16개(13~17개의 치관)의 이빨을 가지고 있다. 위턱 이빨 끝은 약간 바깥쪽으로 향하고, 아래턱 이빨의 혀 쪽에는 삼각형 능선이 있으며, 양쪽 턱의 일부 이빨에는 치관이 있는 치경이 있을 수 있다. 다른 파레이아사우루스류와 달리 스쿠토사우루스는 두개골 기저의 결절 사이에 작은 결절을 가지고 있다.[7]
3. 2. 치아 및 소화 기관
스쿠토사우루스는 넓은 주둥이에 빽빽하게 박힌, 칼날 모양의 이형치성 이빨을 가지고 있었다. 위턱에는 18개(9~11개의 치관을 가짐), 아래턱에는 16개(13~17개의 치관)의 이빨이 나 있었다. 윗니 끝은 약간 바깥쪽으로 튀어나와 있었고, 아랫니의 혀 쪽에는 삼각형 능선이 있었다. 양쪽 턱의 일부 이빨은 치관이 있는 치경을 가질 수 있었다.[7] 넓은 몸통은 넓은 소화 기관을 가졌을 것으로 추정된다.[17]4. 고생물학
스쿠토사우루스는 튼튼한 체격을 가진 파충류로, 몸은 뼈로 된 갑옷으로 덮여 있었고, 두개골에는 여러 개의 뾰족한 뿔이 나 있었다.[22] 무거운 몸과 짧은 다리 때문에 빠르게 움직이기 어려워 포식자에게 취약했을 것으로 보인다. 팀 헤인즈와 폴 챔버스는 스쿠토사우루스의 두꺼운 골격이 강한 근육으로 덮여 있었고, 특히 목 부분이 발달했다고 설명한다. 피부 아래의 단단한 골판은 사슬 갑옷과 같은 역할을 했다.[40] 또한, 이들은 광대뼈가 커서 울음소리가 컸을 것이며, 발달된 청각으로 소리를 통해 의사소통을 했을 가능성이 있다고 추정한다.[40] 먹이는 부드러운 식물이었을 것으로 추정된다.[23][35]
2021년 연구에 따르면, 스쿠토사우루스의 체중은 약 1160kg으로, 현생 검은코뿔소나 소와 비슷한 수준으로 추정되었다.[42]
4. 1. 이동 방식 및 생활사
스쿠토사우루스는 뼈로 된 갑옷과 두개골을 장식하는 여러 개의 뾰족한 뿔을 가진 거대한 체구의 파충류였다.[45] 비교적 작은 몸집에도 불구하고 무겁고 짧은 다리로 인해 오랜 시간 동안 빠른 속도로 움직일 수 없어 큰 포식자의 공격에 취약했다. 스스로를 방어하기 위해 스쿠토사우루스는 특히 목 부위에 강력한 근육으로 덮인 두꺼운 골격을 가지고 있었다. 피부 아래에는 골판이 줄지어 있었고, 이 판은 마치 두정갑 갑옷의 한 형태였다.[52]
사막을 포함한 반건조 기후에 사는 초식동물인 스쿠토사우루스는 먹을 수 있는 신선한 잎을 찾기 위해 오랫동안 돌아다녔을 것이다. 강둑과 범람원에 가까이 붙어있어서 식물 생물이 더 풍부했을 것이고 가뭄이 들 때에만 더 멀리 떠돌았을 것이다. 이빨은 납작했고 잎과 어린 가지를 갈아서 큰 내장에서 길게 소화시킬 수 있었다. 스쿠토사우루스는 식물을 소화시키기 위해 위석을 삼켰다. 스쿠토사우루스는 먹이를 계속 먹어야 한다는 점을 고려하면, 아마도 먹을 수 있는 식물의 많은 부분을 배출하지 않기 위해 혼자 살거나 아주 작은 무리를 지어 살았을 것이다.
파레이아사우루스는 오랫동안 육상 동물로 여겨져 왔지만, 현대 해부학적 유사성이 부족하여 그들의 이동 범위를 평가하기 어렵다. 1987년, 이바흐넨코는 깊고 낮은 흉부 거들, 짧지만 통통한 사지, 사지 관절의 두꺼운 연골 때문에 수생 또는 양서류일 것이라고 가설을 세웠는데, 이는 수생 듀공을 연상시킨다. 이후 아프리카와 유라시아 유해에 대한 연구(안정 동위원소 분석 및 발자국 분석 포함)는 모두 육상 행동과 일치하는 결과를 보고했다. 카세이드는 파레이아사우루스와 전반적으로 유사한 체형을 가지고 있으며, 아마도 동일한 이동 습성을 보였을 것이다. 두 종 모두 얇고 다공성인 긴 뼈를 가지고 있는데, 이는 현대 잠수 생물과 일치하지만, 전반적으로 무거운 몸통은 그러한 행동을 방해했을 것이다. 그럼에도 불구하고, 이와 유사한 중력 보행 생물은 훨씬 더 두꺼운 긴 뼈를 가지고 있다. 2016년, 동물학자 마르쿠스 람베르츠와 동료들은 얇은 뼈와 낮은 식물에 도달하기에 적합하지 않은 짧은 목을 근거로, 카세이드는 주로 수생 동물이며 짧은 간격 동안만 육지로 나왔다고 제안했다. 전반적으로 해부학적 증거는 동위원소 증거와 직접적으로 상반되는 것으로 보인다. 뼈 해부학은 동물의 생활 방식보다 체중에 더 관련이 있었을 가능성이 있다.[19]
다른 파레이아사우루스와 마찬가지로, 스쿠토사우루스는 빠른 초기 성장률과 주기적인 성장 간격을 보인 것으로 나타났다. 이러한 비교적 짧은 어린 시절 이후, 개체는 전체 크기의 75%에 도달하고 몇 년 동안 더 느린 속도로 계속 성장했을 것이다. 빠른 성장에서 느린 성장으로의 이러한 전환은 잠재적으로 성적 성숙의 시작을 알리는 신호였다.[19]

5. 고생태학

스쿠토사우루스는 고토양층으로 구성된 살라레프스카야 지층에서 발견된다. 이 지층은 우기 동안 건조하거나 반 건조한 평원을 덮친 여러 차례의 대규모 홍수로 설명되며, 여러 개의 임시로 채워진 수로와 영구적인 마른 호수가 특징이다.[20]
스쿠토사우루스는 상부 타타리아 시대의 파레이아사우루스-고르곤옵스 동물군에 속했으며, 당시 생태계는 파레이아사우루스, 이형치류, 고르고놉스, 테로케팔리아류, 키노돈트류가 지배적이었다. 이전 지층과는 달리 디노케팔리아류는 완전히 사라졌다.[21] 이 지역에서는 다양한 이끼류, 레피도파이트류, 고사리류, 펠타스페르마세아류가 발견되었다.[20]
5. 1. 서식 환경
스쿠토사우루스는 얕은 수역과 건조 지대를 주기적으로 반복하는 퇴적층인, 균일하게 붉은 살라레프스카야 지층에서 산출되었다. 층준의 형태나 두께는 지층 전체에서 크게 변화하며, 각 층이 서로 다른 공급원에서 유래했을 가능성이 있다. 지층 상부에서 고토양은 점차 소실되고, 층 두께는 수 밀리미터~수 미터로 불규칙하고 불연속적이 되며, 산화철이 환원되어 생긴 것으로 보이는 청색 반점이 나타나게 된다. 이 층은 탄산염암 층으로 덮여 있는데, 그 두께는 작은 매듭 정도에서 최대 1미터까지 다양하다. 고토양이나 탄산염암에는 식물의 뿌리로 인한 구멍이 뚫려 있지만, 점토암-실트암의 각력암이나 렌즈 모양의 얇은 사층에서는 보존되지 않았다. 이 지층은 건조한 평원이 우기 동안 여러 차례의 대규모 홍수로 씻겨 내려가 형성되었다고 설명되는 것이 일반적이며, 영구적으로 건조한 마른 호수 등의 흔적도 보인다[43]。스쿠토사우루스가 서식했던 동물상에는, 파레이아사우루스과, 이치아목, 고르곤놉스아목, 테로케팔루스류, 키노돈류가 서식하고 있었다. 더 오래된 시대의 층과는 달리, 디노케팔루스류는 완전히 사라졌다. 스쿠토사우루스는 Sokolki 동물상에서 확인되었으며, 해당 동물상에서는 앞선 세 분류군이 주요 종이다. 스쿠토사우루스를 제외하면, 비박사우루스가 유일하게 확인된 초식 동물이다. 그 대신 육식성 동물은 많았으며, 그 중에서도 가장 큰 속이 이노스트란체비아( ''I. latifrons'' 와 ''I. alexandri'')이다. 다른 고르곤놉스아목 동물에는 프라보슬라블레비아와 Sauroctonus progressus가 있었다. 다른 육식성 동물에는 테로케팔루스류의 ''Annatherapsidus petri'', 키노돈류의 도비니아, 크로니오수키드와 세이모리아모르파의 양서류 (카르핀스키오사우루스, 코틀라시아, 드비노사우루스)가 확인되었다[44]。 식물에서는, 다양한 선류와 대엽시드식물, 씨앗고사리류 등이 퇴적되어 있다[43]。
5. 2. 공존 동물
''스쿠토사우루스''는 상부 타타리아 시대의 파레이아사우루스-고르고놉스 동물군에 속했으며, 파레이아사우루스, 이형치류, 고르고놉스, 테로케팔리아류, 키노돈트류가 지배적이었다. 이전 지층과는 달리 디노케팔리아류는 완전히 사라졌다. ''스쿠토사우루스''는 소콜키 동물군에서 확인되었으며, 주로 앞서 언급한 3개의 그룹이 특징이다. ''스쿠토사우루스'' 외에 유일한 초식동물은 ''비박사우루스''이다. 육식동물이 훨씬 더 흔했으며, 가장 큰 육식동물은 ''이노스트란체비아''(''I. latifrons'' 와 ''I. alexandri'')로 확인되었고, 다른 고르고놉스는 ''프라보슬라블레비아'' 와 ''사우로크토누스 프로그레수스''이다. 다른 육식동물로는 테로케팔리아 ''안나테라프시두스 페트리'' 와 키노돈트 ''드비니아''가 있으며, ''카르핀스키오사우루스'', ''코틀라시아'', ''드비노사우루스''를 포함한 크로니오수키드와 세이모리아모르파 양서류도 확인되었다.[21] 식물의 경우, 이 지역에서는 다양한 이끼류, 레피도파이트류, 고사리류, 펠타스페르마세아류가 발견되었다.[20]참조
[1]
서적
The Marshall Illustrated Encyclopedia of Dinosaurs and Prehistoric Animals
Marshall Editions
[2]
논문
Diagnoses of the new forms of vertebrates and plants from the Upper Permian on North Dvina
http://www.mathnet.r[...]
[3]
논문
The evolution of the tetrapod shoulder girdle and fore-limb
[4]
논문
Zur Systematik der Nord-Düna-Pareiasauridae
[5]
논문
Pareiasauriden als Leitfossilien
http://publikationen[...]
[6]
서적
Upper Paleozoic and Mesozoic Amphibians and Reptiles in the USSR
Nauka
[7]
서적
The Age of Dinosaurs in Russia and Mongolia
Cambridge University Press
2003-12-04
[8]
논문
"''Pareiasaurus karpinskii'' Amalitzky, 1922 (currently ''Scutosaurus karpinskii'', Reptilia, Pareiasauria): proposed conservation of the specific name"
https://archive.org/[...]
[9]
논문
Non-marine ostracods of the Permian-Triassic transition from sections of the East European platform
https://www.research[...]
[10]
웹사이트
International Chronostratigraphic Chart
https://stratigraphy[...]
International Commission on Stratigraphy
[11]
서적
Stratotipy i opornye razrezyverkhnei permi Povolzh'ya i Prikam'ya
Ekotsentr
[12]
논문
Sequence of Permian tetrapod faunas of Eastern Europe and the Permian–Triassic ecological crisis
[13]
논문
Volumetric Body Mass Estimate and in vivo Reconstruction of the Russian Pareiasaur ''Scutosaurus karpinskii''
2021
[14]
서적
Evolution of Herbivory in Terrestrial Vertebrates: Perspectives from the Fossil Record
Cambridge University Press
[15]
논문
The homologies and early evolution of the shoulder girdle in turtles
[16]
논문
Pareiasaurian Tracks from the Upper Permian of Eastern Europe
http://golubeff.naro[...]
[17]
서적
Evolution of Herbivory in Terrestrial Vertebrates
[18]
서적
よみがえる恐竜・古生物
SB Creative
2006-07-15
[19]
논문
Bone histology of two pareiasaurs from Russia (''Deltavjatia rossica'' and ''Scutosaurus karpinskii'') with implications for pareiasaurian palaeobiology
[20]
논문
Fossil Roots from the Upper Tatarian Deposits in the basin of the Sukhona and Malaya Severnaya Dvina Rivers: Stratigraphy, Taxonomy, and Paleoecology
https://www.research[...]
[21]
서적
The Age of Dinosaurs in Russia and Mongolia
Cambridge University Press
2003-12-04
[22]
서적
The Marshall Illustrated Encyclopedia of Dinosaurs and Prehistoric Animals
Marshall Editions
[23]
서적
リアルサイズ古生物図鑑 古生代編
2018-08-04
[24]
논문
Diagnoses of the new forms of vertebrates and plants from the Upper Permian on North Dvina
http://www.mathnet.r[...]
[25]
논문
The evolution of the tetrapod shoulder girdle and fore-limb
[26]
웹사이트
NHK海外ドラマ 恐竜SFドラマ プライミーバル 古生物ファイル
http://www9.nhk.or.j[...]
NHK
2020-07-18
[27]
논문
Zur Systematik der Nord-Düna-Pareiasauridae
[28]
논문
"''Pareiasaurus karpinskii'' Amalitzky, 1922 (currently ''Scutosaurus karpinskii'', Reptilia, Pareiasauria): proposed conservation of the specific name"
https://archive.org/[...]
[29]
논문
Pareiasauriden als Leitfossilien
http://publikationen[...]
[30]
서적
Upper Paleozoic and Mesozoic Amphibians and Reptiles in the USSR
Nauka
[31]
서적
The Age of Dinosaurs in Russia and Mongolia
Cambridge University Press
2003-12-04
[32]
논문
Non-marine ostracods of the Permian-Triassic transition from sections of the East European platform
https://www.research[...]
[33]
서적
Stratotipy i opornye razrezyverkhnei permi Povolzh'ya i Prikam'ya
Ekotsentr
[34]
논문
Sequence of Permian tetrapod faunas of Eastern Europe and the Permian–Triassic ecological crisis
[35]
웹사이트
東海大自然史:展示:3階
http://www.scc.u-tok[...]
東海大学
2021-09-24
[36]
서적
Evolution of Herbivory in Terrestrial Vertebrates: Perspectives from the Fossil Record
Cambridge University Press
[37]
저널
The homologies and early evolution of the shoulder girdle in turtles
[38]
저널
Pareiasaurian Tracks from the Upper Permian of Eastern Europe
http://golubeff.naro[...]
[39]
서적
Evolution of Herbivory in Terrestrial Vertebrates
[40]
서적
よみがえる恐竜・古生物
SBクリエイティブ
2006-07-15
[41]
저널
Bone histology of two pareiasaurs from Russia (''Deltavjatia rossica'' and ''Scutosaurus karpinskii'') with implications for pareiasaurian palaeobiology
[42]
저널
Volumetric Body Mass Estimate and in vivo Reconstruction of the Russian Pareiasaur Scutosaurus karpinskii
2021
[43]
저널
Fossil Roots from the Upper Tatarian Deposits in the basin of the Sukhona and Malaya Severnaya Dvina Rivers: Stratigraphy, Taxonomy, and Paleoecology
https://www.research[...]
[44]
서적
The Age of Dinosaurs in Russia and Mongolia
Cambridge University Press
2003-12-04
[45]
서적
The Marshall Illustrated Encyclopedia of Dinosaurs and Prehistoric Animals
https://archive.org/[...]
Marshall Editions
[46]
저널
Volumetric Body Mass Estimate and in vivo Reconstruction of the Russian Pareiasaur ''Scutosaurus karpinskii''
2021
[47]
서적
The Age of Dinosaurs in Russia and Mongolia
Cambridge University Press
2003-12-04
[48]
서적
Evolution of Herbivory in Terrestrial Vertebrates: Perspectives from the Fossil Record
Cambridge University Press
[49]
저널
The homologies and early evolution of the shoulder girdle in turtles
[50]
저널
Pareiasaurian Tracks from the Upper Permian of Eastern Europe
http://golubeff.naro[...]
[51]
서적
Evolution of Herbivory in Terrestrial Vertebrates
[52]
서적
よみがえる恐竜・古生物
SB Creative
2006-07-15
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com