식물의 진화
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
식물의 진화는 지구 생명체의 역사에서 중요한 사건으로, 초기 종속 영양 생물에서 시작하여 엽록소를 가진 독립 영양 생물로의 전환을 거쳐 다양한 조류와 육상 식물로 이어졌다. 식물의 계통은 남조식물, 홍조식물, 녹조식물 등 조류를 포함하며, 선태식물과 관속식물로 분화되었다. 식물의 출현은 대기 중 산소 농도를 증가시키고 오존층 형성에 기여하여 생물의 육상 진출을 가능하게 했다. 육상 식물은 녹조류에서 진화했으며, 차축조류가 가장 가까운 친척이다. 초기에는 단세포에서 다세포로 진화하며, 특히 크리오기 시대의 빙하기가 스트렙토식물의 분화에 영향을 미쳤다. 조류는 동화 색소와 동화 산물의 차이에 따라 다양한 계통으로 진화했으며, 균류는 조류와는 다른 기원을 가진 제3의 생물군으로 여겨진다. 식물의 육상 진출은 큐티클층 발달을 통해 가능했으며, 관다발 식물은 물 수송 시스템을 발달시켜 건조에 적응했다. 선태식물과 양치식물의 생활사에서 우세한 세대가 다르며, 종자 식물은 포자에서 씨앗으로 번식 방식을 전환하여 건조 환경에 더욱 적응했다.
더 읽어볼만한 페이지
- 분류군별 진화 - 진핵생물 발생
진핵생물 발생은 원핵생물에서 진핵생물로의 진화 과정을 연구하는 분야로, 세포 내 공생 가설을 통해 미토콘드리아와 엽록체의 형성을 설명하며 LECA의 특징과 크라운 그룹의 진화를 포함하고, 공생 영양 모델과 포식 영양 모델을 주요 모델로 제시한다. - 고식물학 - 규화목
나무가 땅에 묻혀 광물질에 의해 화석화된 규화목은 일본과 미국 등지에서 발견되며, 특히 일본에서는 다양한 종류가 천연기념물로 지정되어 보호받고 있다. - 고식물학 - 석화목
석화목은 매몰된 나무가 지하수 속 광물질, 주로 실리카에 의해 치환되어 형성된 화석으로, 세포 구조와 형태가 보존되며 고생물학 연구 및 다양한 분야에 응용되는 '모의 석화목' 연구에도 활용된다. - 식물 - 종자식물
종자식물은 씨앗으로 번식하는 관다발식물로서 겉씨식물과 속씨식물로 나뉘며 약 3억 1900만 년 전에 출현하여 육상 생태계에서 번성했고 동물의 먹이가 되면서 공진화 관계를 맺어왔다. - 식물 - 상록식물
상록식물은 1년 내내 잎이 푸르게 유지되는 식물로, 침엽수와 일부 활엽수, 겉씨식물이 속하며, 조경, 방풍, 건축 자재 등 다양한 용도로 활용되고 척박한 환경에서도 잘 자라는 특징을 가진다.
| 식물의 진화 | |
|---|---|
| 개요 | |
 | |
| 시기 | 오르도비스기 ~ 현재 |
| 주요 사건 | 최초의 육상 식물 출현 관다발 식물 진화 종자 식물 진화 속씨 식물 진화 |
| 시대별 주요 식물 | |
| 고생대 | 선캄브리아 시대: 조류 오르도비스기: 최초의 육상 식물 추정 실루리아기: 쿡소니아 데본기: 원시 종자 식물 석탄기: 양치식물 우세 |
| 중생대 | 트라이아스기: 겉씨식물 출현 쥐라기: 겉씨식물 번성 백악기: 속씨식물 출현 및 번성 |
| 신생대 | 속씨식물의 번성 및 다양화 |
| 환경 변화 | |
| 초기 육상 환경 | 척박한 환경, 자외선 노출, 물 부족 |
| 식물의 적응 | 큐티클층 형성 (수분 손실 방지) 배우자낭 및 포자낭 발달 (생식 세포 보호) 관다발 발달 (물과 영양분 운반) 종자 발달 (건조한 환경에서 생존) |
| 기후 변화와 식생 변화 | |
| 석탄기 | 온난 습윤한 기후, 거대한 양치식물 숲 형성 |
| 페름기 | 건조한 기후, 겉씨식물 증가 |
| 백악기 | 온난한 기후, 속씨식물 번성 |
| 추가 정보 | |
| Strother et al., 2011 | 지구상에서 가장 오래된 비해양 진핵생물 |
| Knauth & Kennedy, 2009 | 늦은 선캄브리아 시대 지구 녹화 |
| Stewart & Rothwell, 1993 | 고식물학 및 식물 진화 |
| Rothwell, G. W., Scheckler, S. E. & Gillespie, W. H., 1989 | 늦은 데본기의 구과식물 엘킨시아 속 |
2. 식물의 계통
화석 기록을 통해 생물의 변천 과정을 추측할 수 있는 지질시대 이후, 동물은 식물이 만든 유기물을 이용해왔다. 이 때문에 지구 최초의 생물은 독립 영양 생물이었을 것이라는 추측도 있었으나, 현재는 알렉산드르 오파린이 제창한 코아세르베이트설이 유력하다. 오파린에 따르면, 원시 바다의 단백질, 핵산, 당류 등 유기물 분자들이 전하를 띠고 물 분자를 끌어당겨 콜로이드 입자 상태로 존재했다. 이 입자들이 모여 막으로 둘러싸인 액체 방울, 즉 코아세르베이트를 형성했으며, 이것이 원시 생명체의 출발점이라는 것이다. 코아세르베이트는 주변의 무기물과 유기물을 흡수하며 원시세포 형태로 발전했고, 효소나 핵산이 존재할 경우 물질대사, 분열, 증식 등 생명체와 유사한 활동을 보였다. 이러한 과정을 거쳐 자기 복제 능력과 물질대사 기능을 갖춘 원시 생명체가 탄생했을 것으로 추정된다. 이 가설에 따르면, 원시 지구에 최초로 출현한 생물은 주변 유기물을 흡수하여 생활하는 종속 영양 생물이었을 것이다.
엽록소와 같은 색소를 가진 생물이 출현하여 태양 광선을 이용해 이산화탄소와 물로부터 당류를 합성하고 산소 (O2)를 방출하기 시작했다. 이로 인해 대기 중 산소 농도가 점차 증가하면서 유기 호흡을 하는 독립 영양 생물이 나타났다. 초기 독립 영양 생물은 홍색황세균이나 녹색황세균 등의 광합성 세균이었으나, 이후 물을 분해하고 태양 에너지를 더 효율적으로 사용하는 조류 등으로 진화한 것으로 보인다. 녹색 식물의 광합성으로 원시 해양 속 유기물이 다시 공급되고, 대기 중 산소 농도가 높아지면서 대기 조성이 변하기 시작했다. 증가된 산소는 태양의 자외선을 흡수하여 일부가 오존 (O3)으로 변했고, 이 오존이 대기 상층에 오존층을 형성하여 생물에게 해로운 강한 자외선을 차단하게 되었다. 이로써 물속에서만 살던 생물들이 육상으로 진출할 수 있는 환경이 마련되었다. 또한, 대기 중의 암모니아와 메탄은 산소와 반응하여 각각 기체 상태의 질소와 이산화탄소로 바뀌어 오늘날과 같은 대기가 조성되었다.
오파린이 제창한 코아세르베이트설에 따르면, 원시 지구에 최초로 출현한 생물은 원시 해양의 유기물을 이용하는 종속 영양 생물이었을 것으로 추정된다. 이들 초기 종속 영양 생물이 증식하면서 해양 속 유기물은 점차 감소하고, 무기 호흡의 결과로 이산화탄소 농도는 증가했을 것이다. 이러한 환경 변화 속에서 새로운 생존 방식이 필요하게 되었다.
초기 종속 영양 생물이 증식하면서 해양 속 유기물은 급격히 감소하고, 지구 냉각으로 유기물 자연 합성도 줄어들었다. 당시 산소가 없던 환경에서 종속 영양 생물의 무기 호흡 결과로 이산화탄소 농도는 점차 증가했다. 이러한 환경 변화에 적응하기 위해 새로운 생존 방식이 필요했고, 엽록소와 같은 색소를 이용하여 태양 빛 에너지로 이산화탄소와 물로부터 유기물을 합성하고 산소를 방출하는 독립 영양 생물이 출현하게 되었다. 초기 독립 영양 생물은 홍색황세균이나 녹색황세균 같은 광합성 세균이었으며, 이후 물을 분해하고 태양 에너지를 더 효율적으로 이용하는 조류 등으로 진화한 것으로 보인다. 광합성 세균에서 남조식물이 진화했고, 이들의 광합성 활동으로 대기 중 산소 농도가 증가하면서 녹조식물이 출현했을 것으로 추정된다.
가장 오래된 생물 화석은 식물이며, 남아프리카 짐바브웨의 32억~35억 년 전 암석에서 발견된 지름 10~100μ의 둥근 물체는 남조식물 화석으로 추정된다. 약 25억 년 전 석회암에서는 길이 2~3μ의 세포 집합체가 발견되어, 이 시기 석회질 조류나 세균의 대량 생존을 시사한다. 북아메리카 슈피리어호 부근의 약 20억 년 전 암석에서는 철세균, 남조식물, 망간 및 철산화균, 사상균 등이 발견되었다. 전 세계 철광상의 대부분이 약 20억 년 전에 형성되었는데, 이는 물속 철세균에 의해 산화철 형태로 침전된 것으로 보인다. 이는 당시 대기 중에 상당량의 산소가 존재했음을 의미하며, 녹조식물의 출현 시기와 일치할 가능성이 있다. 캐나다 온타리오주 건플린트 철광상에서는 19억 년 전의 다양한 형태(균사 모양, 별 모양, 둥근 모양 등)의 조류 화석이 발견되었고, 탄소 동위원소 분석 결과 광합성으로 만들어진 유기물임이 확인되었다. 이 시기 생물은 단세포에서 사상체, 다핵체 등으로 점차 복잡해지는 경향을 보였다.
조류의 각 문(門)은 동화 색소와 동화 산물이 다른데, 이는 각기 다른 경로를 통해 별개의 계통으로 진화했음을 시사한다. 자외선이 오존층에 흡수되지 않고 지표에 도달하던 초기, 조류는 깊은 물속에서 생활했다. 이곳에는 파장이 짧은 청색광만 도달하므로, 청색광을 흡수하는 홍조소(피코에리드린)를 가진 홍조식물과 남조식물이 번성했으며, 이 둘은 유연 관계가 깊고 세포 구조도 유사하다. 이후 대기 중 산소와 오존층이 증가하면서 자외선이 차단되자, 조류는 점차 얕은 물로 이동하며 갈조식물, 녹조식물 등 다양한 종류로 분화하며 생활권을 넓혀갔다. 이처럼 환경 변화는 조류의 동화 색소 다양화에 영향을 미쳤다. 고생대 캄브리아기 이후 오르도비스기, 실루리아기에는 현생 조류와 유사한 형태의 화석(디모르포시폰, 카에토크라주스, 베르니포레라 등)이 발견된다.
육상 식물은 약 5억 1천만 년 전쯤[158] 또는 그 이전(기원전 8억 5천만 년경[3])에 녹조류의 한 부류에서 진화한 것으로 보인다. 현생 육상 식물과 가장 가까운 녹조류는 차축조 식물문, 특히 차축조류이다.[5][159] 이들의 조상은 얕은 민물, 아마도 계절적으로 마르는 작은 웅덩이 가장자리에 서식하던[85][158] 가지를 치는 실 모양의 조류였을 것으로 추정된다. 이 조류는 반수체였을 것이다. 최근에는 현존하는 클렙소르미디움강과 유사한 단세포 육상 차축조류에서 유래했을 가능성도 제기된다.[6] 균류와의 공생 관계가 초기 식물의 육상 환경 적응에 도움을 주었을 수 있다.[7][158][160]
육상 식물이 최초의 육상 광합성 생물은 아니었다. 암석 풍화 연구에 따르면 약 12억 년 전[158], 담수호 퇴적층 화석 증거에 따르면 약 10억 년 전[161]부터 생물이 육상에 존재했을 가능성이 있다. 그러나 대기 구성에 큰 영향을 미칠 정도의 번성은 8억 5천만 년 전 이후에 시작된 것으로 보인다.[162] 이들 초기 육상 생물은 작고 단순한 형태였을 것이다.[158]
육상 식물의 가장 오래된 직접적인 증거는 약 4억 7천만 년 전 중기 오르도비스기의 4분자 포자[163] 화석이다.[164][165] 이는 감수 분열 결과 생성된 4개의 포자가 연결된 형태로, 모든 육상 식물과 일부 조류에서 나타난다.[163] 초기 포자의 미세 구조는 현생 선태식물의 것과 매우 유사하여 같은 단계의 생물이었음을 시사한다.[166] 이후 오르도비스기 후기에는 4분자 포자가 분리된 삼조형 포자가 나타난다.[169] 포자 표면의 Y자형 삼조 흔적[163]은 포자벽이 견고했음을 의미하며, 이는 건조 환경에 대한 저항성을 시사한다. 건조 저항성은 수중 생활을 벗어난 포자의 중요한 특성이다.[163] 최초의 육상 식물 대형 화석은 실루리아기의 엽상체 형태로 발견되며, 물에 잠겼을 때만 생존 가능한 습지 식물이었음을 보여준다.[170]
육상으로 진출한 식물은 건조 환경에 적응해야 했다. 선태식물은 습한 환경을 선호하거나 건조 시 휴면 상태에 들어가는 방식으로 적응했다. 반면 관다발식물은 방수성 외피(큐티클)를 발달시켜 수분 손실을 막고, 기공을 통해 가스 교환을 하며, 관다발 조직으로 물 수송 효율을 높이는 방식으로 건조에 저항했다. 또한, 세대 교번에서 포자체 세대가 우세해지는 방향으로 진화했다.
육상 식물의 확산은 광합성을 통해 대기 중 산소 농도를 크게 높였다. 산소 농도가 13%를 넘어서면서 산불 발생이 가능해졌고, 이는 실루리아기 전기 이후 발견되는 탄화된 식물 화석으로 확인된다.[171] 탄화 작용은 식물의 단단한 탄소 껍질만 남겨 보존성을 높이며, 세포 수준의 미세 구조 관찰을 가능하게 하는 중요한 화석화 과정이다.
3. 식물의 출현
선캄브리아기의 생물은 모두 물속에서 살았지만, 오존층 형성으로 자외선이 차단되자 식물계에서는 녹조식물과 같은 녹색 식물들이 육상으로 진출하기 시작했다. 갈조식물은 미역, 감태, 모자반류처럼 몸 구조가 더 복잡하고 조직 분화가 나타나는 경우도 있었지만, 체표에 큐티클층이 없어 건조에 약해 육상에 정착하지 못했다. 반면, 상대적으로 구조가 단순했던 녹조식물은 큐티클층이 발달하여 체내 수분 증발을 막고 건조에 견딜 수 있었기 때문에 육상 진출에 성공할 수 있었다.
육상 식물은 약 8억 5천만 년 전에 녹조류의 한 부류에서 진화했을 것으로 추정되지만,[3] 조류와 같은 식물은 10억 년 전부터 진화했을 수 있다.[2] 육상 식물의 가장 가까운 현존 친척은 차축조류이며, 특히 차라류이다. 현대 차라류의 조상이 육상 식물의 조상과 유사하다면, 이는 육상 식물이 얕은 민물에 서식하는 분지형 실 모양의 반수체 조류에서 진화했음을 의미하며,[5] 아마도 계절적으로 건조되는 웅덩이 가장자리에서 진화했을 것이다.[85] 그러나 일부 최근 증거에 따르면 육상 식물은 현존하는 클렙소르미디움강과 유사한 단세포 육상 차축조류에서 유래되었을 수 있다.[6] 초기 식물은 균류와의 공생 관계를 통해 육상 환경의 스트레스에 적응하는 데 도움을 받았을 수 있다.[7]
육상 식물이 최초의 육상 광합성 생물은 아니었다. 풍화 속도 연구는 광합성 능력이 있는 생물이 이미 12억 년 전에 육지에 살았음을 시사하며,[85] 10억 년 전 담수호 퇴적물에서 미생물 화석이 발견되기도 했다.[8] 그러나 탄소 동위원소 기록에 따르면, 이들 생물은 8억 5천만 년 전까지 대기 조성에 큰 영향을 미치지는 못했던 것으로 보인다.[3] 이들은 계통 발생학적으로 다양했지만,[9] 아마도 작고 단순하여 조류 찌꺼기 이상을 형성하지 못했을 것이다.[85] 육상 식물이 출현하는 데 시간이 걸린 이유로는 대기의 '독성' 문제,[11] 필요한 복잡성 진화에 걸린 시간,[12] 또는 적절한 토양의 부재[13] 등이 거론된다. 초기 육상 토양은 질소나 인과 같은 필수 자원이 부족하고 물 보유 능력도 낮았다.
대기 중 오존량이 현재와 비슷해져 식물이 육상에 정착할 수 있게 된 시기는 화석 증거로 볼 때 고생대 오르도비스기에서 실루리아기에 이르는 시기이다. 초기 육상 식물의 증거는 약 4억 7천만 년 전 사우디 아라비아[14]와 곤드와나[15]의 하부 중기 오르도비스기 암석에서 발견된 포자 화석이다. 이 포자들은 스포로폴레닌이라는 매우 단단한 물질로 된 벽을 가지고 있어 화석으로 잘 보존되었으며, 단독(단립), 쌍(이립), 또는 네 개가 뭉친 사립체 형태로 발견된다. 그 미세 구조는 현대 우산이끼류 포자와 유사하여 비슷한 수준의 조직을 가졌음을 시사한다.[16] 포자 벽에 스포로폴레닌이 포함된 것은 이들이 배아 식물(육상 식물)과 관련 있음을 더욱 입증한다.[17] 약 4억 5천 5백만 년 전 상부 오르도비스기 암석에서는 관다발 식물과 유사한 삼렬 포자(사립체가 분리되면서 Y자형 흔적이 남은 포자)가 나타난다.[18][19] 이러한 삼렬 흔적은 포자벽이 초기부터 튼튼하고 건조에 강했음을 의미하며, 이는 육상 환경 적응의 중요한 특징이다. 물 밖에서 생존해야 할 때 유용한 특성이기 때문이다. 실제로 물로 돌아간 배아식물은 저항성 벽이 없어 삼렬 흔적을 가지지 않는다.[20]
가장 오래된 육상 식물의 거대 화석은 엽상체 형태의 유기체로, 실루리아기 초기(란도베리절) 하천 습지 환경의 범람원에서 발견되었다. 이들은 땅이 물에 잠겼을 때만 생존할 수 있었던 것으로 보인다.[21] 미생물 매트도 존재했다.[22] 실루리아기 말에는 최초의 관다발식물이자 육상에 처음 적응한 양치식물 중 하나인 쿠크소니아가 출현했다. 그러나 포자 증거 등을 고려할 때, 선태식물은 이보다 다소 이른 시기에 육상에 진출했을 것으로 추정된다.
식물은 육상 환경의 건조에 대처하기 위해 두 가지 주요 전략을 발전시켰다. 현대 선태식물처럼 습한 환경으로 서식지를 제한하거나, 건조 시 신진대사를 멈추고 수분이 공급될 때까지 기다리는 방식(Targionia 속 우산이끼 등)이다. 반면, 관다발 식물은 몸 표면에 방수성 외부 큐티클 층을 발달시켜 수분 손실을 줄이고(일부 선태식물도 마찬가지), 가변 개구부인 기공을 통해 대기 중의 CO2 흡수와 가스 교환 속도를 조절하며 건조에 저항했다. 또한 물과 양분을 운반하는 관다발 조직을 발달시켜 몸을 지지하고 수직 성장을 가능하게 하여 더 큰 식물의 진화를 위한 길을 열었다.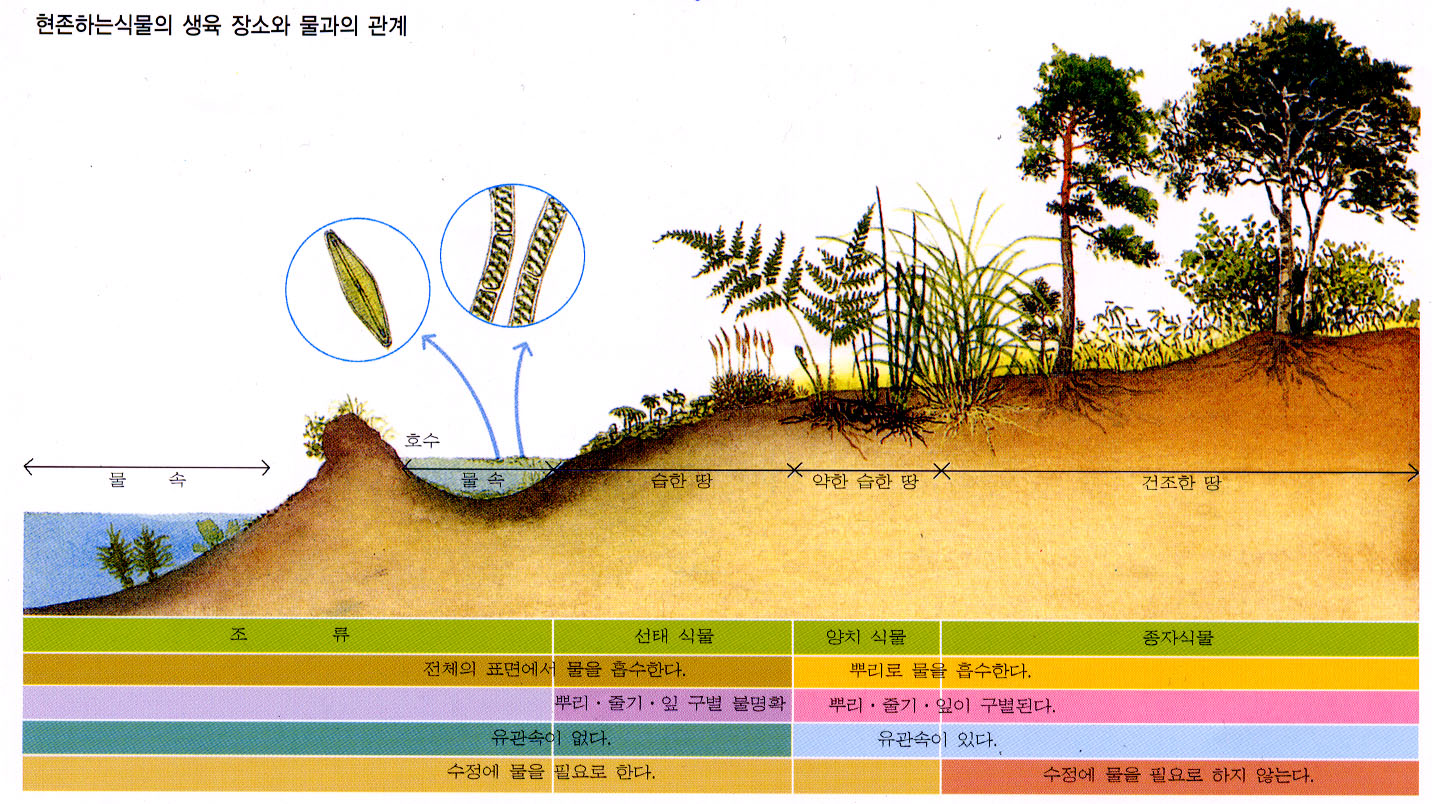
4. 단세포에서 다세포로
이러한 상황에서 엽록소와 같은 색소를 가지고 태양 광선을 이용하여 이산화탄소와 물로부터 당류를 합성하고 산소를 방출하는 독립 영양 생물이 출현했다. 최초의 광합성 생물은 혐기성인 홍색황세균, 녹색황세균, 홍색세균 등으로, 이들은 물 대신 황화수소나 수소 가스를 이용해 광합성을 했다. 이후 물을 분해하여 수소를 이용하고 산소를 방출하는 광합성 방식이 나타났으며, 이는 남조식물(시아노박테리아)로 이어지는 진화 과정에서 더욱 발전했다. 남조식물의 광합성 활동은 대기 중 산소 농도를 점차 증가시키는 중요한 계기가 되었다.
화석 기록을 통해 초기 생물의 흔적을 찾아볼 수 있다.
이러한 증거들을 종합하면, 지구상 생명체는 약 35억 년 전 단세포 형태로 출현하여 점차 사상체, 다핵체 등으로 체제가 복잡화되는 경향을 보였다. 특히 약 20억 년 전후로는 다양한 종류의 세균과 조류가 번성하며 다세포 생물로 진화하는 기반을 마련했다.
한편, 약 7억 2천만 년 전에서 6억 3천 5백만 년 전 사이의 크리오기 시대에 발생한 전 지구적 빙하기(눈덩이 지구)는 초기 광합성 생물이 이산화탄소 농도를 감소시켜 온실 효과를 약화시킨 결과일 수 있다.[23] 2022년 연구에 따르면, 다세포 스트렙토식물의 기원이 이 추운 크리오기 시대와 일치하며, 이후 따뜻한 에디아카라기에 이들의 분화가 이루어졌다는 점이 관찰되었다. 이는 빙하기가 광합성 생물에게 가한 선택압의 결과로 해석되며, 일부 생물은 상대적으로 따뜻한 환경에서 살아남아 이후 육상 환경으로 진출하는 기반을 마련했을 수 있다.[24] 윤조강(Zygnematophyceae)의 단세포 형태 등은 저온 환경에 대한 추가적인 적응을 반영할 수 있다.[24]
5. 조류의 다원성
조류의 각 문(門) 사이에 동화 색소와 동화 산물이 다른 점은 이들이 각기 다른 과정을 통해 서로 다른 계통으로 진화했음을 보여준다. 자외선이 대기 중의 오존층에 흡수되지 않고 직접 지표면에 도달하던 시기에는 조류가 주로 깊은 물 속에서 생활했다. 깊은 물 속에는 대부분의 빛이 물을 통과하며 흡수되고 파장이 짧은 청색광만이 도달할 수 있는데, 이러한 환경에서는 청색광을 흡수하는 동화 색소인 홍조소(피코에리드린)를 가진 생물이 유리했다. 홍조소를 가진 홍조식물과 남조식물은 이러한 환경에 적응했으며, 이 둘은 생식 세포나 세포 구조 면에서도 매우 유사하여 유연 관계가 깊은 것으로 여겨진다.
이후 광합성 생물의 활동으로 대기 중에 산소가 축적되고 오존층이 형성되면서 자외선이 효과적으로 차단되기 시작했다. 이에 따라 조류는 점차 얕은 물에서도 생활할 수 있게 되었다. 이러한 환경 변화 속에서 갈조식물, 녹조식물 등 다양한 종류의 조류가 출현했으며, 이들은 수면을 넘어 지상으로까지 생활권을 넓혀나갔다. 이처럼 다양한 조류들이 서로 다른 동화 색소를 갖게 된 것은 지구 환경 변화에 적응한 결과이다.
일반적으로 조류의 세포벽은 단단하지 않아 화석으로 남기 어렵다. 하지만 일부 조류는 세포벽에 석회질이 침적되는 특징을 가져 화석화되기도 한다. 이러한 조류 화석은 고생대 캄브리아기 이후, 특히 오르도비스기와 실루리아기 지층에서 발견되며, 현생 조류와 유사한 형태를 보여준다.
| 분류 | 속명 | 발견 시기 |
|---|---|---|
| 녹조식물 (청각과) | Dymorphosiphon (디모르포시폰) | 오르도비스기 |
| 녹조식물 (청각과) | Chaetocladus (카에토크라주스) | 실루리아기 |
| 녹조식물 (삿갓말과) | Vermiporella (베르니포레라) | 오르도비스기 |
| 녹조식물 (삿갓말과) | Rhabdoporella (라프도포레라) | 실루리아기 |
| 홍조식물 (석회조) | Delesserites (데레세리데스) | 실루리아기 |
| 홍조식물 (석회조) | Solenopora (솔레노포라) | 캄브리아기 |
6. 균류의 출현
균류는 조류와 마찬가지로 화석으로 남기 어려운 생물군이다. 균사가 단독으로 화석이 되는 경우는 매우 드물고, 대부분 식물 조직 속에서 기생하거나 공생하는 형태로 발견된다. 예를 들어, 데본기의 양치식물 뿌리나 뿌리줄기 화석 속에서 공생하던 균사를 관찰할 수 있다.
과거에는 균류가 광합성 능력을 상실한 조류로부터 유래했을 것이라는 견해가 있었다. 균류의 기원에 대해서는 여러 학설이 제기되어 왔다:
- 홍조식물 기원설: 자낭을 가진 자낭균류가 낭과를 가진 홍조식물에서 유래했다는 주장이다. 두 생물군은 생식 세포에 편모가 없다는 공통점을 가진다. 또한, 담자균류에 속하는 녹균 역시 홍조식물에서 분화되었다고 보는 시각도 있다.
- 원생동물 기원설: 동물과 식물의 구분이 모호한 원생동물로부터 균류가 유래했다는 설이다. 이 관점에서는 아메바와 유사한 원생동물에서 이른바 고생균류(하등한 균류의 모임)와 점균이 발생하였고, 다시 고생균류로부터 난균류나 통곰팡이류 등이 갈라져 나왔다고 설명한다. 다른 한편으로는, 원생동물에서 생식 세포의 편모 유무와 개수에 따라 ▲편모가 1개인 무리(일모균류) ▲편모가 2개인 무리(이모균류) ▲편모가 없는 무리(접합균류, 자낭균류, 담자균류)로 각각 진화했다고 보기도 한다.
최근에는 균류가 동물도 식물도 아닌, 독자적인 기원을 가진 제3의 생물군이라는 견해가 설득력을 얻고 있다. 생명의 기원이 종속 영양 생물에서 시작되었다는 학설이 유력해짐에 따라, 스스로 양분을 만들지 못하고 외부 유기물에 의존하는 종속 영양 생물인 균류 역시 식물과는 구별되는 독자적인 생물군으로 인정하는 추세이다.
7. 식물의 상륙
선캄브리아기의 생물은 모두 물 속에서 살았다. 당시 원시 지구의 대기에는 산소가 거의 없었고, 따라서 오존층도 존재하지 않았다. 이 때문에 생물에게 해로운 강한 자외선이 지표면에 직접 도달하여, 생물은 자외선을 막아주는 깊은 물 속에서만 살 수 있었다.[85]
그러나 녹색 식물의 광합성 활동으로 대기 중에 산소(O2)가 점차 증가하게 되었다. 이 산소의 일부가 태양의 자외선이나 번개 방전 등에 의해 오존(O3)으로 변하면서 대기 상층에 오존층을 형성했다. 오존층은 생물에게 유해한 강한 자외선을 막아주는 역할을 하여, 물 속에서만 살던 생물들이 육지로 올라와 생활할 수 있는 환경을 만들었다. 또한, 대기 중의 암모니아와 메탄은 산소와 반응하여 각각 기체 상태의 질소와 이산화탄소(CO2)로 바뀌면서 오늘날과 유사한 대기 조성이 만들어졌다.
육상 식물은 약 8억 5천만 년 전[3] 또는 5억 1000만 년 전쯤[158]에 녹조류의 한 부류에서 진화한 것으로 추정된다. 조류와 같은 식물은 이미 10억 년 전부터 진화했을 수 있다.[2] 현존하는 육상 식물과 가장 가까운 친척은 차축조류이며, 특히 차라류이다. 만약 현대 차라류가 육상 식물의 조상과 유사하다면, 육상 식물은 얕은 민물에 서식하는, 가지를 치는 실 모양의 반수체(haplontic) 조류에서 진화했을 가능성이 있다.[5][159] 이들은 아마도 계절적으로 건조해지는 웅덩이 가장자리에서 살았을 것이다.[85][158] 일부 최근 증거는 육상 식물이 현존하는 클렙소르미디움강과 유사한 단세포 육상 차축조류에서 유래했을 수도 있음을 시사한다.[6] 초기 식물이 육상 환경의 스트레스에 적응하는 데에는 균류와의 공생 및 협력적 상호 작용이 도움이 되었을 수 있다.[7][158][160]
육상 식물이 최초의 육상 광합성 생물은 아니었다. 암석 풍화 속도 연구에 따르면 광합성 생물은 이미 12억 년 전에 육지에 존재했을 수 있으며,[85][158] 약 10억 년 전 담수호 퇴적물에서 미생물 화석이 발견되기도 했다.[8][161] 그러나 탄소 동위원소 기록은 이들 생물이 8억 5천만 년 전까지는 대기 조성에 큰 영향을 미칠 만큼 충분하지 않았음을 시사한다.[3][162] 이 초기 육상 생물들은 계통 발생학적으로 다양했지만,[9] 아마도 작고 단순하여 조류 피막(algal scum) 이상을 형성하지는 못했을 것이다.[85][158] 지의류가 먼저 육상에 정착하여 식물의 진출을 도왔다는 가설도 있지만, 분자 계통 발생 및 화석 기록은 이를 지지하지 않는다.[10]
육상 식물이 출현하는 데 오랜 시간이 걸린 이유로는 대기의 '독성' 문제,[11] 필요한 복잡성을 진화시키는 데 걸린 시간,[12] 또는 적절한 토양의 부재[13] 등이 제기된다. 초기 육상 토양은 질소, 인과 같은 필수 영양소가 부족했고 물을 보유하는 능력도 거의 없었다.
화석 기록에 따르면, 식물이 육상으로 진출할 수 있게 된 것은 고생대 오르도비스기에서 실루리아기에 이르는 시기이다. 이 시기에 대기 중 오존량이 현재와 비슷해진 것으로 추정된다. 초기 육상 식물의 증거는 약 4억 7천만 년 전 중기 오르도비스기 암석에서 발견된 포자 화석이다.[14][15][164][165] 이 포자들은 단독, 쌍, 또는 네 개가 뭉친 사분체(tetrad) 형태로 발견되며, 스포로폴레닌이라는 매우 단단한 물질로 된 벽을 가지고 있어 화석으로 잘 보존된다. 이 포자들의 미세 구조는 현대 우산이끼류의 포자와 유사하여 비슷한 수준의 조직을 가졌음을 시사하며,[16][166] 스포로폴레닌 벽은 이들이 배아식물임을 뒷받침한다.[17]
사분체 포자가 분리되면서 각 포자에 Y자 모양의 삼렬 흔적(trilete mark)이 생기는데, 이는 포자들이 서로 눌렸던 자국이다.[20][163] 이러한 삼렬 포자는 약 4억 5500만 년 전 후기 오르도비스기 암석에서 나타나기 시작한다.[18][19][169] 삼렬 흔적이 생기려면 포자벽이 초기부터 튼튼해야 하며, 이는 건조에 강한 특성과 관련이 있다. 건조 저항성은 물 밖에서 생존해야 하는 포자에게 필수적인 특징이다. 실제로 물 속 생활로 돌아간 배아식물들은 튼튼한 포자벽과 삼렬 흔적이 없다.[20][163] 조류 포자 역시 삼렬 흔적을 가지지 않는데, 이는 포자벽이 충분히 튼튼하지 않거나, 튼튼하더라도 흔적이 생기기 전에 분리되거나, 정사면체 형태의 사분체를 이루지 않기 때문이다.[20][163]
최초의 육상 식물 대형 화석은 실루리아기 초기(란도베리절)의 하천 습지에서 발견된 엽상체 유기체이다. 이들은 땅이 물에 잠겼을 때만 생존할 수 있었으며, 당시 범람원의 상당 부분을 덮었던 것으로 보인다.[21][170] 미생물 매트도 함께 발견되었다.[22] 실루리아기 말기에는 최초의 관다발식물이자 가장 오래된 육상 양치식물 중 하나인 쿠크소니아가 나타났다. 하지만 실제로는 선태식물이 이보다 조금 더 일찍 상륙했을 것으로 추정된다.
조류 중에서는 녹조식물보다 갈조식물이 체제나 내부 구조가 더 복잡하다. 갈조식물 중에는 미역, 감태, 모자반류처럼 뿌리, 줄기, 잎과 유사한 기관이 분화된 경우도 있고, 모자반류처럼 줄기 내부에 통도 조직과 유사한 조직 분화를 보이는 경우도 있다. 그럼에도 불구하고 체제가 비교적 단순한 녹조식물이 육상에 진출하고 복잡한 갈조식물은 그러지 못한 주된 이유는, 녹조식물은 몸 표면에 큐티클층이 발달하여 체내 수분 증발을 막고 건조에 견딜 수 있었던 반면, 갈조식물은 큐티클층이 없어 건조에 매우 약했기 때문이다.
식물이 육지에 정착한 후 건조에 대처하는 방식은 크게 두 가지로 나뉘었다. 현대 선태식물과 같은 그룹은 건조를 피하거나(습한 환경에 서식) 건조에 굴복하여(물이 다시 공급될 때까지 신진대사를 멈춤) 생존한다. 반면, 관다발 식물은 물 손실 속도를 조절하여 건조에 저항한다. 이들은 공기에 노출된 모든 표면에 방수성 큐티클 층을 가지고 있어 수분 손실을 줄인다(일부 선태식물도 마찬가지). 그러나 큐티클이 몸 전체를 덮으면 대기 중의 CO2 흡수가 어려워지므로, 관다발 식물은 가스 교환을 조절하기 위해 여닫을 수 있는 구멍인 기공을 발달시켰다. 또한, 몸 전체로 물을 효율적으로 운반하기 위해 관다발(물관부 등) 조직을 발달시켰고, 세대 교번에서 포자체가 우세한 생활 주기로 전환했다. 이러한 적응, 특히 관다발 조직의 발달은 식물이 물의 지지 없이도 수직으로 성장할 수 있게 하여 더 큰 육상 식물의 진화를 가능하게 했다.
육상 식물 군락이 확산되면서 대기 중 산소 농도는 이전보다 훨씬 높아졌다. 산소 농도가 약 13%를 넘어서면서부터 산불이 발생할 가능성이 생겼으며, 이는 실루리아기 전기의 탄화된 식물 화석을 통해 확인된다.[171] 데본기 말기의 공백기를 제외하면, 탄화된 식물 화석은 이후 지질 시대에서도 계속 발견된다. 탄화는 중요한 화석화 과정 중 하나로, 산불로 인해 식물의 연한 부분이 제거되고 단단한 탄소 껍질만 남아 보존되는 방식이다. 이 탄화된 화석은 세포 수준의 미세 구조까지 잘 보존하는 경우가 많다.
8. 선태식물과 양치식물
현재 알려진 초기 육상 식물로는 선태식물과 양치식물이 있는데, 이 둘 중 어느 쪽이 먼저 육지에 정착했는지, 즉 녹조식물에서 어느 쪽이 먼저 분화되었는지에 대해서는 두 가지 학설이 존재한다.
하나는 녹조식물에서 선태식물을 거쳐 양치식물로 진화했다는 학설이고, 다른 하나는 녹조식물에서 먼저 양치식물이 분화된 후, 이들이 축소·퇴화하여 선태식물이 되었다는 학설이다. 과거에는 녹조식물, 선태식물, 양치식물의 체제와 내부 구조를 비교했을 때 전자의 학설이 더 타당하다고 여겨져 많은 학자들의 지지를 받았다.
그러나 다음과 같은 이유로 후자의 학설(양치식물 → 선태식물)을 지지하는 견해도 있다.
- 화석 기록: 양치식물 화석(쿠크소니아 등)은 실루리아기 말에 나타나 데본기에 다양하게 분화된 반면, 선태식물 화석은 데본기 기록이 드물고 석탄기 이후에 주로 발견된다. 이는 화석상으로는 양치식물이 더 오래되었음을 시사한다.
- 현존 선태식물의 축소 경향: 현재 살아있는 선태식물들을 비교하면, 몸체가 점차 작아지는 방향으로 진화했을 가능성이 보인다. 예를 들어, 데본기 중기의 양치식물인 타에니오크라다는 특정 우산이끼류와 구조적으로 유사하며, 역시 데본기 중기 양치식물인 호르네오피톤의 포자낭 내 축기둥 구조는 뿔이끼류와의 연관성을 시사한다. 이는 양치식물에서 선태식물로의 축소 진화 가능성을 보여주는 해석이다.
하지만 화석 기록만으로는 진화 방향을 단정하기 어렵다. 타에니오크라다는 포자체(포자낭을 가진 개체) 화석만 알려져 있고 배우체(장란기, 장정기를 가진 개체)는 발견되지 않았는데, 우산이끼의 몸체는 배우체이므로 단순히 생김새만으로 직접 비교하기는 어렵다.
오히려 식물계 전체의 진화 흐름과 여러 생물학적 특징을 고려할 때, 녹조식물에서 선태식물을 거쳐 양치식물로 진화했다는 첫 번째 학설이 더 설득력이 있다.
- 체제 및 구조: 조류(녹조식물 포함)에서 갑자기 복잡한 관다발 조직과 분화된 기관을 가진 양치식물이 직접 진화했다고 보기에는 무리가 있다. 두 그룹 사이의 체제 차이가 크고, 이를 연결할 중간 단계의 화석도 발견되지 않았다.
- 육상 적응과 배우체 축소: 육상 환경, 특히 건조한 환경에 적응하기 위해 식물은 배우체를 축소시켜 적은 물로도 수정이 가능하도록 진화해왔다. 양치식물의 작은 전엽체나 종자식물의 꽃가루관, 배낭 등이 그 예이다. 선태식물에서 관찰되는 몸체 축소화 경향은 이러한 육상 적응의 결과이지, 양치식물에서 퇴화했다는 의미와는 다르다. 배우체가 커지는 방향의 진화는 건조 환경에서 불리하며, 수정 기관인 장란기가 더 크고 복잡해지는 방향으로 진화했다는 이론도 이해하기 어렵다.
- 장란기 구조: 선태식물의 장란기는 배우체 표면에 완전히 노출되어 있고 목 부분이 길며 여러 개의 경구 세포를 가진다. 반면, 양치식물의 장란기는 알을 가진 배 부분이 배우체 조직 속에 묻혀 보호받고, 짧은 목 부분만 밖으로 나와 있으며 경구 세포도 1개(2핵성)인 경우가 많다. 보호받는 구조에서 노출된 구조로 진화했다고 보기는 어려우므로, 장란기 형태는 선태식물에서 양치식물로의 진화 방향을 지지한다.
- 차축조식물과의 관계: 녹조식물의 일종 또는 독립된 문으로 분류되는 차축조식물은 여러 면에서 선태식물과 유사성을 보인다. 장란기와 장정기가 다세포성 영양 세포로 덮여 있고, 생란기의 형태가 선태식물 장란기의 원시 형태로 보이며, 접합 포자 발아 시 태류의 원사체와 비슷한 발아관을 만들고, S자 모양의 정자를 만드는 점 등이 그렇다. 또한, 조류에는 없는 플라보노이드 화합물, 특히 선태식물에 흔한 글루코플라본을 함유하고 있어 녹조식물보다는 선태식물에 더 가깝다고 볼 수 있다. 이는 녹조식물과 선태식물의 연결성을 강화한다.
- 생활사 비교: 선태식물은 배우체(유성 세대)가 우세하며 포자체(무성 세대)는 배우체에 기생한다. 반면 양치식물은 포자체가 우세하고 배우체는 작지만 독립 생활을 한다. 녹조식물과 차축조식물은 대부분 배우체가 발달해 있으므로, 생활사 측면에서도 녹조식물은 양치식물보다 선태식물과 더 가깝다.
이러한 증거들을 종합해 볼 때, 배우체가 우세한 녹조식물에서 유래한 선태식물의 일부가 스포로고니테스, 호르네오피톤, 리니아와 같은 원시적인 양치식물을 거쳐 포자체가 우세한 양치식물로 세대 교번 형태를 전환하며 진화했다고 보는 것이 타당하다.
한편, 초기 육상 식물의 육상 정착 과정에서 균류와의 공생이 중요한 역할을 했다. 실루리아기 및 데본기 초기의 식물 화석에서는 뿌리가 발견되지 않지만, ''호르네오피톤'' 등 일부 종에서 뿌리털과 유사한 구조가 관찰된다. 이 시기 식물들은 물과 영양분을 흡수하는 관다발 조직도 없었다. 라이니 셰일에서 발견된 데본기 식물 ''아글라오피톤''[35]은 뿌리가 없었지만, 줄기 세포 내에 수지상 균근(Arbuscular mycorrhiza)을 형성하는 곰팡이와 공생 관계를 맺은 최초의 육상 식물 중 하나로 밝혀졌다.[36] 곰팡이는 토양에서 인산염과 같은 영양분을 흡수하여 식물에게 제공하고, 식물로부터 당분을 얻었다. ''아글라오피톤''을 포함한 초기 육상 식물들은 물과 영양분 흡수를 위해 이러한 균근 곰팡이에 크게 의존했을 것으로 추정된다.[37]
이 공생 곰팡이는 글로메로미코타문(Glomeromycota)에 속하며,[38] 약 10억 년 전에 출현하여 현재까지도 선태식물, 양치식물, 겉씨식물, 속씨식물 등 대부분의 육상 식물(관다발 식물의 80% 이상)과 공생 관계를 맺고 있다.[41] DNA 염기 서열 분석 결과, 수지상 균근 공생은 육상 식물의 공통 조상이 육지로 진출할 때 시작되었으며,[39] 어쩌면 육상화를 가능하게 한 결정적인 요인이었을 수도 있다.[40] 식물이 뿌리를 진화시키기 전, 균근 곰팡이는 식물이 스스로 얻기 힘든 인과 같은 무기 영양분과 물을 공급해주고, 그 대가로 유기 화합물을 얻었다.[41] 이러한 공생은 우산이끼와 같은 단순한 식물의 생존과 번성에도 기여한다.[42][43]
9. 차축조식물의 특징
차축조식물은 녹조식물의 일부로 다루어지기도 하고, 독립된 문으로 취급되기도 한다. 녹조식물과 동화 색소와 동화 산물이 같다는 점에서는 녹조식물의 계통으로 볼 수 있다.
그러나 여러 특징에서 선태식물과 유사성을 보인다. 차축조식물의 장란기와 장정기는 다른 조류처럼 단세포가 아니고 영양체 세포로 덮여 있다. 특히 생란기는 겉보기에 선태식물 장란기의 조상형을 닮았으며, 접합 포자가 감수 분열하여 싹 틀 때 태류의 짧은 원사체와 비슷한 발아관을 만든다. 또한 2개의 편모를 가진 S자 모양의 정자가 생기는 점도 태류와 비슷하다.
차축조식물의 몸은 마디 부분과 마디사이 부분이 되풀이되어 이루어져 있다. 자성 생식 기관인 장란기는 마디 사이 세포(절간 세포)에서 유래하며, 이를 둘러싸는 5개의 관세포는 마디 부분 세포(절부 세포)에서 나뉘어 나온 것이다. 반면, 선태식물이나 양치식물의 장란기는 배우체 표면에 있는 1개의 세포로부터 난자와 그것을 둘러싸는 벽세포가 만들어진다. 이 때문에 차축조식물의 장란기와 선태·양치식물의 장란기를 겉모양만으로 비교하기는 어려울 수 있다. 그럼에도 불구하고, 노출된 단세포성 생란기가 영양 세포에 덮이게 된 것은 육상 식물로 진화하기 위한 준비 단계로 해석될 수 있다.
식물체 성분에서도 차이가 나타난다. 차축조식물에는 플라보노이드 화합물이 함유되어 있는데, 이는 다른 조류에서는 볼 수 없는 물질이다. 특히 글루코플라본은 대부분의 선태식물에 흔히 들어 있는 물질이다.
이러한 점들을 종합해 볼 때, 차축조식물은 녹조식물보다 선태식물에 더 가깝다고 할 수 있다. 또한, 양치식물보다는 선태식물이 차축조식물을 통해 녹조식물과 더 밀접하게 관련되어 있다고 볼 수 있다.
10. 화석이 되기 어려운 선태식물
식물의 육상 진출 순서를 고려하면 선태식물이 양치식물보다 먼저 녹조식물에서 분화되어 상륙했을 것으로 추정된다. 그러나 현재까지 발견된 화석 기록상으로는 양치식물(실루리아기의 쿠크소니아) 화석이 선태식물(데본기 후기의 헤파티키테스) 화석보다 더 오래된 것으로 나타난다.
이러한 시간적 불일치는 선태식물의 몸 구조와 생태가 화석이 되기 어려웠기 때문이다. 데본기 중기부터 화석 기록에 등장하기 시작한 양치식물은 그 몸 구조로 미루어 건조한 환경에 적응한 초본이었을 가능성이 높다. 이는 데본기에 퇴적된 적색 사암의 존재로 뒷받침되는데, 적색 사암은 고온 건조한 조건, 즉 당시 환경이 열대 사막과 유사했음을 시사한다. 이러한 환경에서는 큰 나무가 자라기 힘들었을 뿐만 아니라, 습한 환경을 선호하는 선태식물 역시 거의 생육하지 못했을 것이다. 데본기의 육상 식물 화석 대부분이 초본성 양치식물인 이유가 여기에 있다.
하지만 데본기에도 사막 속의 오아시스나 지형적 영향으로 늘 구름과 안개가 끼는 습한 언덕 같은 미소 서식지가 존재했을 수 있다. 이러한 곳에서는 아르카에오프테리스와 같은 비교적 큰 나무들이 작은 규모의 삼림을 형성하고, 그 아래에는 다양한 선태식물이 번성했을 것으로 추정된다. (이는 오늘날 칠레 아타카마 사막 남쪽 반사막 지대에 존재하는 '프라이 호르헤의 삼림'과 유사한 생태 환경을 상상해볼 수 있다.)
문제는 이렇게 고온 다습한 환경에서는 선태식물의 유체가 쉽게 부패하여 화석으로 남기 어려웠다는 점이다. 반면, 세포벽이 단단하게 목화(木化)된 관다발식물, 특히 그 목질부는 상대적으로 화석화되기에 유리했다. 아르카에오프테리스는 이러한 데본기 습윤 지역 삼림을 구성했던 주요 수종 중 하나로 여겨진다. 결국 선태식물은 화석 기록상으로는 양치식물보다 늦게 나타나지만, 이는 실제 출현 순서라기보다는 화석화 조건의 차이 때문일 가능성이 높다.
11. 세대 교번의 역전
선태식물과 양치식물의 주요 차이점 중 하나는 생활사에서 우세한 세대가 다르다는 점이다. 선태식물은 유성 세대인 배우체가 발달하고, 무성 세대인 포자체는 배우체에 의존한다. 반면 양치식물은 포자체가 발달하고, 배우체는 독립 생활을 하지만 크기가 작다. 녹조식물은 대부분 배우체가 발달하며, 차축조식물은 배우체만 존재한다. 따라서 생활사 측면에서 녹조식물은 양치식물보다 선태식물에 더 가깝다고 볼 수 있다.
원시 육상 식물인 선태식물과 양치식물 중 어느 쪽이 녹조식물에서 먼저 분화했는지에 대해서는 두 가지 학설이 있다.
1. 선태식물에서 양치식물로 진화: 녹조식물에서 선태식물을 거쳐 양치식물로 진화했다는 학설이다. 식물체의 구조나 내부 조직 비교 시 타당성이 높아 많은 지지를 받는다.
2. 양치식물에서 선태식물로 퇴화: 녹조식물에서 먼저 양치식물이 분화된 후 선태식물로 축소·퇴화되었다는 학설이다. 근거는 다음과 같다.
- 화석 기록상 선태식물이 양치식물보다 더 새로운 시대에서 발견된다. 양치식물(쿡소니아)은 실루리아기 말에 나타나 데본기에 다양하게 분화했지만, 선태식물은 데본기 기록이 적고 석탄기 이후에 많이 발견된다.
- 현존 선태식물을 비교하면 몸체가 축소되는 방향으로 진화했다고 볼 수 있다. 데본기 중기 양치식물(타에니오크라다, 호르네오피톤)과 현생 선태식물(우산이끼류, 뿔이끼류)의 구조적 유사성을 근거로 든다.
그러나 화석 기록만으로 진화 방향을 단정하기는 어렵다. 식물계 전체의 진화 흐름을 볼 때, 조류에서 복잡한 관다발과 분화된 조직을 가진 양치식물이 직접 진화했다고 보기는 어렵다. 조류와 양치식물을 직접 연결하는 화석 증거는 없으며 체제 차이도 크다. 또한 타에니오크라다는 포자체만 알려져 있어 배우체가 주인 선태식물과 직접 비교하기 어렵다.
육상 식물 전반에서 몸이 작아지는 경향은 건조한 환경에 적응하는 과정으로 해석될 수 있다. 즉, 식물은 배우체를 축소시켜 적은 물로도 수정이 가능하도록 진화했다. 양치식물의 전엽체, 종자식물의 꽃가루관이나 배낭이 그 예이다. 선태식물의 몸 축소화는 이러한 적응의 일부이며, 양치식물에서 선태식물로의 퇴화와는 다른 의미를 갖는다. 건조 환경에서 배우체가 커지거나, 수정 기관인 장란기가 더 크고 복잡해지는 방향으로 진화하기는 어렵다. 오히려 물이 부족하면 장란기는 단순해지고 보호받는 구조로 진화하는 것이 자연스럽다. 양치식물의 장란기는 배우체 조직 속에 묻혀 보호받지만, 선태식물의 장란기는 노출되어 있고 구조적으로 더 원시적이다. 따라서 장란기 형태로 보아도 양치식물에서 선태식물로 진화했다고 보기는 어렵다.
결론적으로, 배우체가 우세한 선태식물의 일부가 스포로고니테스, 호르네오피톤, 리니아와 같은 원시양치강 식물을 거쳐 포자체가 우세한 양치식물로 세대 교번형을 전환했을 가능성이 높다고 여겨진다.
모든 다세포 육상 식물은 배우체 단계와 포자체 단계를 포함하는 세대 교번 생활 주기를 가진다. 배우체는 단상(n) 상태로 배우자(정자와 난자)를 만들고, 포자체는 복상(2n) 상태로 포자를 만든다. 모든 현생 육상 식물(배아식물)은 두 세대의 형태가 매우 다른 이형 세대 교번을 보인다.[172]
식물 진화의 주요 경향은 배우체 세대의 축소와 포자체 세대의 증대이다. 육상 식물의 조상인 조류는 거의 확실히 단상 생활사를 가졌으나, 모든 육상 식물은 단상과 복상 단계 모두 다세포인 단복상 생활사를 영위한다.[1] 선태식물은 배우체를 생활 주기의 주요 단계로 발달시켰고 포자체는 배우체에 의존하게 되었다. 반대로 관다발 식물은 포자체를 주요 단계로 발달시켰으며, 특히 종자식물에서는 배우체가 매우 축소되었다.
복상(2n) 단계가 우세해진 이유 중 하나로, 복상 상태가 유전자 보완을 통해 해로운 돌연변이의 영향을 줄일 수 있다는 점이 제안되었다.[28][29] 이는 복제 정확도를 크게 높이지 않고도 게놈 크기와 정보량을 늘려 새로운 환경 적응에 유리했을 수 있다. 그러나 이 견해는 이끼와 속씨식물에서 단상체보다 복상체 단계에서 선택이 더 효과적이지 않다는 증거로 반박되기도 했다.[30]
단복상 생활 주기의 출현을 설명하는 두 가지 주요 이론이 있다.
- 보간 이론 (Interpolation theory 또는 Antithetic/Insertion theory):[31][173] 두 배우체 세대 사이에 다세포 포자체 단계가 새롭게 삽입되었다고 본다. 접합자가 감수분열 전 유사분열을 통해 복상(2n) 포자체를 형성하고, 이 포자체가 나중에 감수분열하여 포자를 생성한다는 것이다. 초기 포자체는 배우체와 형태가 다르고 구조가 단순했을 것으로 보며,[31] 이는 배우체에 의존하는 단순한 포자체를 가진 선태식물의 특징과 잘 맞는다. 이후 포자체가 복잡해지면서 광합성 능력을 갖추고(일부 뿔이끼류처럼) 독립하며, 결국 기관과 관다발 조직을 발달시켜 관다발 식물처럼 포자체가 우세한 단계가 되었다는 설명이다.[1][172] 작은 쿡소니아 화석이 배우체에 의해 지지되었을 것이라는 관찰도 이 이론을 뒷받침한다.[31][173]
- 전환 이론 (Transformation theory 또는 Homologous theory): 포자체가 감수분열 시기를 늦춤으로써 갑자기 출현했다고 가정한다. 동일 유전 정보가 사용되므로 초기에는 배우체와 포자체의 형태가 비슷했을 것으로 본다. 이는 배우체와 포자체 형태가 같은 일부 조류(Ulva lactuca)의 생활사를 설명한다. 이후 건조한 육상 환경에 적응하면서 생식에 불리한 배우체는 단순화되고, 포자를 효과적으로 퍼뜨리기 위한 포자체는 복잡하게 발달했다는 것이다.[1][172] 라이니 처트(Rhynie chert)에서 발견된 초기 관다발 식물(Rhynia)의 포자체와 배우체가 비슷한 복잡성을 보인다는 점이 이 가설의 근거로 제시된다.[31][32][33][173] 그러나 솔잎란(Psilotum)을 제외한 대부분의 현생 관다발 식물은 포자체와 배우체의 형태가 매우 다르며, 배우체에는 관다발 조직이 거의 없다.[34]
12. 포자에서 씨로
식물이 육상으로 진출한 후 가장 큰 어려움은 수정을 위해 물을 얻는 것이었다. 선태식물과 양치식물은 배우체를 습한 곳에서 자라게 해야 했으므로, 더 건조한 곳으로 분포를 넓히기 위해서는 배우체를 건조로부터 보호할 필요가 있었다. 선태식물은 배우체가 포자체보다 크고 포자체가 배우체에 영양적으로 의존하기 때문에 배우체 보호에 한계가 있었다.
반면, 양치식물은 포자체가 훨씬 크며, 일부는 배우체를 매우 작게 만들어 포자의 두꺼운 벽 안에서 성숙시키는 방식으로 진화했다. 석송류(소엽류)인 부처손속·물부추속과 고사리류(대엽류)인 수생고사리류가 이러한 이형 포자성을 보인다. 이들은 크고 작은 두 종류의 포자를 만드는데, 소포자는 수배우체를, 대포자는 암배우체를 형성한다. 특히 석탄기의 인목(레피도덴드론)과 근연 관계인 일부 화석 식물(레피도카르폰, 미아데스미아)에서는 대포자낭이 포자엽이나 돌기(주피)로 싸여 더욱 보호되었고, 대포자낭 속에서 단 하나의 대포자만 성숙하는 특징을 보여 씨의 조상 형태로 여겨진다. 이러한 구조는 대포자낭(주심) 안에 하나의 대포자가 생기고, 주피를 가지며, 대포자가 만들어진 곳에서 배우자가 생겨 수정 후 배를 만드는 씨의 기본 요건을 갖추기 시작했음을 보여준다.
양치식물의 수정에는 정자가 헤엄쳐 이동할 물이 필요하지만, 부처손속 등 일부 이형 포자성 양치류는 비교적 적은 양의 물만 필요로 하며, 전엽체가 완전히 노출되지 않아 건조에 더 잘 견딜 수 있었다.
종자식물은 여기서 더 나아가, 밑씨(대포자낭과 주피)가 어미 식물에 붙어 영양을 공급받고, 꽃가루가 꽃가루관을 통해 정자를 직접 운반함으로써 수정 과정에서 물에 대한 의존성을 거의 또는 완전히 극복했다. 이는 식물이 건조한 환경으로 확산하는 데 결정적인 역할을 했다. 씨는 배를 보호하고 휴면을 가능하게 하여 생존율을 높였다.
12. 1. 양치식물과 겉씨식물
선태형의 세대 교번을 거듭하던 원시양치류에서 솔잎란류(나경강)로 진화하여 양치형의 세대 교번이 확립되었고, 이로부터 석송류(소엽강), 속새류(설엽강), 그리고 고사리류(대엽강)가 분화된 것은 데본기 중엽이다. 거의 동시에 원시겉씨강 역시 솔잎란류로부터 진화한 것으로 여겨진다. 이 원시겉씨강에서 소철류와 침엽수류라는 겉씨식물의 두 주요 계열이 갈라져 나왔다.[178]뿌리의 발달초기 육상 식물들은 뿌리가 없었으나[78], 땅속으로 뻗어 영양분을 흡수하는 구조는 점차 발달했다. 실루리아기-데본기의 ''Rhynia''나 ''Horneophyton'' 같은 식물들은 뿌리와 생리적으로 동일한 기능을 하는 구조를 가졌지만[79][80], 근관을 가진 진정한 뿌리는 아니었다.[85] 뿌리와 유사한 작은 구조인 근모는 매우 일찍 진화했을 가능성이 있으며[85], Rhynie chert 화석에서도 명확히 관찰된다.[81] 데본기 동안 뿌리 구조는 점점 더 흔해지고 깊이 침투했다. 석송류 나무는 중기 데본기에 약 20cm 길이의 뿌리를 형성했고, 후기 데본기에는 전나자식물(progymnosperm)이 약 1m 깊이까지 뿌리를 내렸다.[100] 진정한 겉씨식물과 자이곱테리스 양치류도 후기 데본기에 얕은 뿌리 시스템을 형성했다.[100]
특히 석송류의 뿌리내림 방식은 독특했는데, 이들의 근경(''Stigmaria'')은 잎과 같은 기관이 뿌리 역할을 하는 줄기와 동일한 구조였다.[85] 이는 현존하는 석송류 ''Isoetes''에서도 유사한 구조가 관찰되어, 뿌리가 석송류와 다른 식물(유엽식물)에서 적어도 두 번 독립적으로 진화했음을 시사한다.[85][83] 뿌리의 발달은 토양을 변화시키고 CO2 농도에 영향을 미쳐 지구 환경에 큰 영향을 주었으며[75][76], 후기 데본기 대멸종의 원인 중 하나로 지목되기도 한다.[77]


나무 형태와 목재의 진화초기 데본기의 육상 식물은 키가 작았으나, 더 높은 키는 햇빛 경쟁과 포자(나중에는 씨앗) 분산에 유리했다. 이를 위해 효과적인 관다발 시스템과 지지 구조가 필요했고, 목재 조직과 2차 생장 능력이 진화했다. 2차 생장은 관다발 형성층이라는 분열 조직 고리가 안쪽으로 물관부(목재)를, 바깥쪽으로 체관부를 만들어내며 이루어진다.[86] 목재는 원래 물 수송을 위해 진화했으며, 이후 기계적 지지 기능이 추가된 것으로 보인다.[87]
최초로 2차 생장과 목질 습성을 개발한 것은 고사리류로, 중기 데본기의 ''Wattieza''는 이미 8m 높이에 달하는 나무 형태를 갖추었다.[88] 후기 데본기에는 트리메로파이트에서 진화한 겉씨식물의 전구체인 ''Archaeopteris''가 나타나 높이 30m에 이르렀다.[89] 진정한 목재는 양면 형성층에서 자라며, 이는 lignophyte라는 분기군에서 단 한 번 진화한 것으로 생각된다. 그중 하나인 ''Rellimia''는 중기 데본기에 처음 나타났다.[90]

후기 데본기와 석탄기에는 Lepidodendrales 형태의 나무 같은 석송류가 숲을 지배하며 석탄 매장층을 형성했다.[91] 이들은 높이 50m 이상, 밑동 지름 2m를 초과하기도 했다. Lepidodendrales는 현대 나무와 달리 단일 면 형성층을 가져 줄기가 계속 굵어지지 못했고, 일정 높이까지 자란 뒤 가지를 치고 포자를 퍼뜨린 후 죽는 제한적인 성장 방식을 보였다.[92][1] 석탄기에는 속새류인 ''Calamites''도 등장했는데, 이들은 단일 면 형성층을 가졌지만 목재를 발달시켜 10m가 넘는 높이로 자랄 수 있었다.

종자의 출현초기 육상 식물은 포자를 통해 번식했으며, 수정 과정에 물이 필요하여 습한 환경에 제한되었다.[213] 이러한 한계를 극복하는 과정에서 이형 포자성(크기가 다른 두 종류의 포자, 즉 소포자와 대포자를 만드는 것)이 발달했고, 이는 종자의 출현으로 이어졌다. 대포자낭 안에 하나의 대포자만 남게 되고, 이 대포자가 포자낭 안에서 발아하여 주피(integument)라는 보호층에 싸인 배주가 형성되었다. 소포자에서 발아한 수배우체(웅성배우체)는 화분 형태로 퍼져나가 배주에 도달하여 수정한다.[178]
소엽 식물과 스페노필룸류는 거의 종자에 가까운 구조를 만들었다. 예를 들어 ''Lepidocarpon'' 및 ''Achlamydocarpon''과 같은 화석 소엽 식물의 대포자는 지름이 1cm에 달했으며, 영양 조직에 둘러싸여 포자체 내에서 대배우체로 발아했다. 그러나 매우 작은 틈새이지만 대포자가 대기에 접촉하고, 주심(내부의 포자를 덮는 층에 싸여 있는 부분)이 포자를 덮지 않기 때문에 진정한 종자라고 할 수 없다. 그 결과로 건조에 대한 내성이 불충분하며, 정자가 대포자의 조란기에 도달하기 위해 파고 들어갈 필요가 없었다.[178]
진정한 종자를 가진 최초의 "종자 식물"은 종자고사리류(Pteridospermatophyta)이다. 이 명칭은 잎이 고사리 잎과 유사했기 때문이지만, 반드시 대엽 고사리 식물과 가까운 계통 관계에 있는 것은 아니다. 종자 식물의 가장 오래된 화석 증거는 후기 데본기에서 유래되었으며, 이는 전나자식물(progymnosperm)로 알려진 그룹에서 진화했다. 이 초기 종자 식물은 나무에서 덩굴성 관목 등으로, 한편 초기 전나자식물도 목본성으로 고사리 모양의 잎을 가지고 있어서 매우 유사했다. 이들은 배주는 가지고 있었지만, 구과(솔방울 등)나 과실과 같은 것은 가지고 있지 않았다. 초기 종자의 진화를 추적하는 것은 어렵지만, 단순한 트리멜로피톤류에서 대칭 포자의 전나자식물 ''Aneurophytales'' 등으로의 계통을 추적할 수 있다.[178]
이 종자의 형식은 기본적으로 나자식물에 공유되어 있다. 종자는 대부분 목질 또는 육질(주목류 등의 경우)의 구과에 싸여 있지만, 완전히 싸여 있지는 않다. 피자식물은 심피에 완전히 싸여 있는 종자를 가진 유일한 분류군이다.
완전히 싸인 종자에 의해 휴면이라는 새로운 능력이 발견되었다. 배는 완전히 외부 공기로부터 차단되어 건조로부터 보호되기 때문에 발아 전 건조를 수년 동안 견딜 수 있다. 후기 석탄기의 나자식물 종자에서 배를 포함하는 것이 발견되었다. 이는 수정과 발아 사이에 긴 기간이 있음을 보여준다.[214] 이 시기는 지구가 온난기가 되었을 때와 겹치며, 건조도가 상승하고 있다. 이것은 휴면이 건조 기후에 대한 대응으로 발생했음을 시사한다. 습윤기까지 발아를 기다리는 것이 이점이 된다.[214] 이러한 진화적 혁신은 큰 가능성을 열었다. 건조 산지 등 이전에는 생육에 부적합했던 지역이 허용 가능하게 되었고, 나무로 덮이게 되었다.[214]
종자는 산포에도 이점을 가져다주었다. 그것은 수정된 배우체의 성공률도 증대시켰다. 또한 배와 영양이 함께 들어있으므로 생존에 부적합한 환경에서도 스스로 영양을 획득할 수 있을 만큼 빠르게 성장할 수 있게 되었다.[213] 예를 들어 배젖이 없으면 묘목은 건조한 상태에서는 지하수면에 닿을 정도로 뿌리를 뻗을 여유가 없다.[213] 마찬가지로, 어두운 하부 관목에 떨어진 종자는 자체 유지를 할 수 있을 때까지 햇빛을 얻기 위해 빠르게 높이 자라야 하므로 여분의 에너지가 필요하다.[213] 종자 식물은 이러한 이점으로 인해 초기 숲의 주요 식물이었던 ''Archaeopteris''를 생태학적으로 압도했다.[213]
속새류의 계통적 위치한편, 속새류는 현생종이나 화석종 모두 씨가 형성될 기미를 보이지 않는다. 석탄기의 ''Calamites''에서 겨우 이형 포자성을 볼 수 있지만, 대포자의 수가 줄어들거나 주피가 형성되는 등의 종자로 발전하는 경향은 전혀 나타나지 않는다. 따라서 속새류와 현재의 종자식물 사이에는 직접적인 계통적 유연 관계가 없는 것으로 간주된다.
12. 2. 속씨식물의 출현
실루리아기에 육상으로 진출한 식물들은 데본기에 여러 갈래로 분화되었는데, 그중 한 갈래는 건조한 환경에 적응하여 씨라는 번식 방법을 만들어냈다. 이후 육상 식물 중 겉씨식물은 석탄기에서 페름기를 거치며 점차 늘어났다. 석탄기에는 원시겉씨강에서 진화했다고 여겨지는 코르다이테스목(Cordaitales)과 양치종자목(Pteridospermatophyta)이라는 겉씨식물의 두 주요 계통군이 발달했다. 페름기에서 트라이아스기를 거치는 동안 코르다이테스목에서는 침엽수목이, 양치종자목에서는 소철목과 벤네티테스목(Bennettitales)이 분화되어 발달하게 되었다. 페름기 말에서 트라이아스기 초에 걸쳐 지구상에는 두 번째 고온·건조 시대가 도래했다. 이 시기에 포자로 번식하던 거대한 목본성 양치식물(예: 인목, 봉인목 등)은 멸망하고, 씨로 번식하는 겉씨식물이 살아남게 되었다.이러한 환경 변화 속에서 씨앗은 건조한 환경으로부터 자신을 더욱 효과적으로 보호하는 방향으로 진화했다. 외부에 노출되어 있던 고사리류의 전엽체와 달리, 겉씨식물의 배우체는 대포자벽, 주심(nucellus), 주피(integument)라는 3중 보호 기관으로 덮여 있었다. 나아가 밑씨(ovule) 자체가 대포자엽(megasporophyll)에 싸이게 되었는데, 이렇게 대포자엽이 접혀서 밑씨를 감싼 구조가 씨방(ovary)의 기원이 되었다.

새로운 적색 사암층이 퇴적될 무렵, 그린란드에는 카이토니아(Caytonia)라는 식물이 자라고 있었다. 카이토니아는 겉보기에는 가지 끝에 씨가 하나씩 달린 것처럼 보이지만, 단면을 보면 10~20개의 밑씨를 가진 대포자엽 자체가 안쪽으로 말려 둥글게 변한 구조임을 알 수 있다. 이는 씨방의 조상 형태로 여겨지며, 이 작은 덩어리는 열매의 조상 형태일 수 있다. 카이토니아의 대포자엽은 밑부분 가까이에 입술 모양의 작은 돌기가 있어 입처럼 벌어져 있는데, 이는 마치 암술의 암술머리처럼 보인다. 그러나 실제 수분은 꽃가루가 대포자엽에 싸인 밑씨의 주공(micropyle)에 직접 도달하는 겉씨식물의 방식으로 이루어진다. 또한, 꽃가루는 소나무처럼 양쪽에 공기 주머니를 가지고 있어 겉씨식물의 특징을 보인다. 하지만 밑씨의 구조는 속씨식물(피자식물) 단계에 더 가깝다. 카이토니아는 소철목이나 벤네티테스목처럼 양치종자류에서 진화한 것으로 보이며, 양치종자류로부터 더 직접적인 영향을 받았기 때문에 '중생대의 양치종자류'라고도 불린다.
카이토니아는 포자엽이 밀집된 꽃 구조를 갖지 않았다. 현존하는 속씨식물의 꽃이 어떻게 발생했는지 이해하기 위해서는 고사리의 포자엽이 밀집된 구조를 살펴볼 필요가 있는데, 이러한 구조는 소철목이나 벤네티테스목에서 발견된다. 특히, 벤네티테스목의 키카디오이데아(Cycadeoidea)는 긴 꽃대 위에 아래쪽부터 포엽(bract, 화피 역할), 깃털 모양의 소포자엽(microsporophyll, 수술 역할), 대포자엽(megasporophyll, 1개의 밑씨를 가짐, 암술 역할)이 나선상으로 빽빽하게 배열되어 있어 현존하는 목련목(Magnoliales)의 꽃과 유사하다. 목련목과 벤네티테스목의 유연 관계는 내부 구조의 특징에서도 나타난다. 목련목 식물 중 일부는 속씨식물이면서도 물관이 없고 헛물관만 가지고 있는데, 이는 벤네티테스류와 동일하다. 이 두 그룹의 헛물관은 원래 계단 모양(계문)으로 두꺼워지는 특징을 보이는데, 이는 양치식물에서 흔히 볼 수 있는 원시적인 형태이다. 만약 벤네티테스형 꽃의 대포자엽이 카이토니아처럼 접혀 밑씨를 보호하게 된다면 목련형 꽃과 유사해질 것이다. 이러한 이유로 현재 목련목은 가장 원시적인 속씨식물 그룹 중 하나로 여겨진다.

초기 육상 식물은 양치식물처럼 포자를 통해 번식했다. 포자는 작은 배우체로 발아하여 난자와 정자를 만들었고, 정자는 습한 환경을 통해 이동하여 수정란을 형성하고 새로운 포자체로 자라났다.[100][213] 이러한 방식은 식물을 습한 환경에 제약했지만, 이형포자성(heterospory)의 발달은 이러한 제약에서 벗어나 밑씨와 종자로 이어지는 길을 열었다. 이형포자 식물은 크기가 다른 두 종류의 포자, 즉 소포자(microspore)와 대포자(megaspore)를 만든다. 이들은 각각 소배우체(microgametophyte)와 대배우체(megagametophyte)로 발달한다. 극단적인 경우, 대포자낭(megasporangium)은 단 하나의 대포자만을 가지게 되고, 이 대포자는 포자낭 안에서 발아하여 대배우체가 된다. 이 대배우체는 방수성 외피(주피) 안에 포함되어 밑씨를 형성한다. 소포자에서 발달한 꽃가루는 바람 등을 통해 운반되어 대배우체에 도달한 후 정자를 방출하여 수정한다.[1][178]
석송류와 속새류 일부는 종자와 유사한 구조를 발달시켰지만 완전한 종자에는 이르지 못했다. 예를 들어, 화석 석송류인 레피도카르폰(Lepidocarpon)의 대포자는 지름이 1cm에 달하고 영양 조직에 둘러싸여 포자체 내에서 발아했지만, 포자를 완전히 감싸는 주피가 없어 밑씨라고 하기에는 부족했다.[1][178] 벨기에에서 발견된 데본기 식물 화석인 런카리아(Runcaria)는 가장 오래된 종자 식물보다 약 2천만 년 앞선 전구체로 여겨진다. 런카리아는 컵 모양 구조(cupule)로 둘러싸인 대포자낭을 가지며, 종피와 꽃가루 유도 시스템을 제외한 종자 식물의 여러 특징을 보여준다.[101]
최초로 진정한 종자를 가진 식물은 '씨앗고사리(seed fern)' 또는 양치종자목(Pteridospermatophyta)으로 불린다. 이들은 잎 모양이 고사리와 비슷했지만 실제로는 밀접한 관련이 없었다. 가장 오래된 종자 식물 화석은 후기 데본기 지층에서 발견되며, 원시겉씨식물(progymnosperm) 그룹에서 진화한 것으로 보인다. 이 초기 종자 식물들은 나무에서 작은 관목까지 다양했으며, 대부분 양치식물과 유사한 잎을 가진 목본 식물이었다. 밑씨는 가지고 있었지만 구과(cone)나 열매는 없었다.[1][178] 종자 식물은 데본기 말 파메니안 시대에 첫 번째 주요 진화적 방사(diversification)를 겪었다.[102]
이러한 종자 구조는 기본적으로 모든 겉씨식물(gymnosperm)에서 공유된다. 겉씨식물은 대부분 씨앗을 목질의 구과나 육질 구조 안에 두지만, 씨앗을 완전히 감싸지는 않는다("나체 씨앗"이라는 의미). 반면 속씨식물(angiosperm, "씨앗이 용기 안에 있음")은 씨앗을 심피(carpel) 안에 완전히 감싸는 유일한 식물 그룹이다.
씨앗이 완전히 감싸지면서 종자 휴면(seed dormancy)이라는 새로운 전략이 가능해졌다. 외부 환경과 완전히 차단되어 건조로부터 보호받는 배아는 발아하기 전에 수년간의 가뭄도 견딜 수 있었다. 후기 석탄기 겉씨식물 씨앗에서 배아가 발견된 것은 수정과 발아 사이에 긴 간격이 있었음을 시사하며, 이는 건조한 기후 조건에 대한 적응이었을 가능성이 높다.[103][214] 이러한 진화적 돌파구는 식물이 건조한 산비탈과 같이 이전에는 살기 어려웠던 지역으로 서식지를 확장하는 데 기여했다.[103][214] 또한, 씨앗은 배아와 함께 영양 저장고(배젖 등)를 가지고 있어 불리한 환경에서도 빠르게 발아하고 초기 성장을 할 수 있게 해주었다. 이는 어두운 숲 바닥이나 건조한 환경에서 묘목의 생존율을 크게 높였다.[100][213] 이러한 이점들 덕분에 종자 식물은 데본기 숲의 주요 구성원이었던 아르카이옵테리스(Archaeopteris)와 같은 초기 식물들을 생태학적으로 압도하며 초기 숲의 생물 다양성을 증가시켰다.[100][213] 그러나 씨앗은 휴면 기간 동안 DNA 손상이 축적될 수 있다는 단점도 가진다.[105][106][107][108]
꽃은 화석 기록에서 비교적 늦게 나타나는 속씨식물 고유의 변형된 잎 구조이다. 속씨식물 그룹은 초기 백악기에 기원하여 다양화되었으며, 이후 생태계에서 중요한 위치를 차지하게 되었다.[109] 꽃과 유사한 구조는 약 1억 3천만 년 전 백악기 화석 기록에서 처음 나타난다.[110] (일부 연구는 더 이른 시기를 주장하기도 하지만 논란이 있다.[111][112]) 꽃의 주요 기능은 번식이며, 이는 꽃이 진화하기 전에는 소포자엽과 대포자엽이 담당했던 역할이다.
오랫동안 속씨식물은 겉씨식물 내에서, 특히 네토식물(Gnetophyta)과 가까운 관계에서 진화했다고 여겨져 왔다. 그러나 최근의 분자 계통학 연구 결과는 이러한 전통적인 형태학적 관점과 다르다.[94][95][199][200] 분자 증거에 따르면 네토식물은 속씨식물보다 다른 겉씨식물 그룹(예: 침엽수)과 더 가깝고,[93][198] 현존하는 겉씨식물과 속씨식물은 각각 별개의 분기군(clade)을 형성하며 약 3억 년 전에 갈라진 것으로 추정된다.[93][94][95][198][113][215]
꽃의 진화를 이해하기 위해서는 속씨식물의 줄기군(stem group)을 파악하는 것이 중요하다. 배우체를 보호하는 구조는 여러 식물 그룹에서 독립적으로 진화했을 수 있으므로(수렴 진화), 화석 기록 해석에 주의가 필요하다. 속씨식물의 꽃에서는 심피가 밑씨를 감싸 보호하며, 밑씨 자체도 이중의 주피로 보호된다. 이러한 다중 보호층을 뚫고 수정하기 위해 속씨식물은 독특한 생식 전략을 발달시켰다. 꽃가루는 단 3개의 세포로 구성되며, 그중 하나가 주피를 뚫는 꽃가루관을 형성하여 2개의 정자 세포를 운반한다. 대배우체는 단 7개의 세포로 이루어지는데, 하나의 정자는 난세포와 융합하여 배를 형성하고, 다른 하나의 정자는 중앙 세포와 융합하여 영양 조직인 배젖(endosperm)을 형성한다. 이 '중복 수정(double fertilization)' 과정은 속씨식물 고유의 특징이다.
화석 기록에는 꽃과 유사한 구조를 가진 몇몇 멸종 그룹이 존재한다. 페름기의 씨앗고사리 글로소프테리스(Glossopteris)는 이미 심피와 유사하게 구부러진 잎 구조를 가지고 있었다.[97] 트라이아스기의 카이토니아(Caytonia)는 더욱 꽃과 유사하며 밑씨가 외피(단일 주피)에 싸여 있었지만, 꽃가루와 수술의 세부 구조는 속씨식물과 다르다. 벤네티테스목(Bennettitales)은 꽃잎이나 꽃받침과 유사한 포엽으로 둘러싸인 놀랍도록 꽃과 비슷한 생식 기관을 가졌지만, 이들은 속씨식물보다 소철이나 은행나무와 더 가까운 관계이며 독립적으로 유사한 구조를 진화시킨 것으로 보인다.
진정한 꽃은 현존하는 속씨식물 그룹에서만 발견된다. 대부분의 형태학적 및 분자적 분석 결과, 암보렐라(Amborella), 수련목(Nymphaeales), 아우스트로바일레이아목(Austrobaileyales)이 속씨식물 계통수의 가장 기저에서 분기한 그룹("ANA 등급")으로 여겨진다. 이 그룹들은 초기 백악기 약 1억 3천만 년 전에 분화한 것으로 추정되는데, 이는 가장 오래된 속씨식물 화석(아르카이프룩투스(Archaefructus))과 최초의 속씨식물 유사 꽃가루(클라바티폴레니테스(Clavatipollenites))가 나타난 시기와 일치한다.[114][115][216][217][116][218] 이후 목련류(magnoliids)가 분기했고, 약 1억 2500만 년 전까지 빠른 속도로 진정쌍자엽식물(eudicots)과 외떡잎식물(monocots)이 출현했다.[116][218] 백악기 말(약 6600만 년 전)에는 현존하는 속씨식물 목(order)의 절반 이상이 진화했으며, 속씨식물은 전 세계 식물 종의 약 70%를 차지하게 되었다.[117][219] 이 시기에는 속씨식물 나무가 침엽수보다 우세해지기 시작했다.[1][220]
기저 그룹인 "ANA 등급" 식물의 특징은 속씨식물이 어둡고 습하며 자주 교란되는 환경에서 기원했을 가능성을 시사한다.[118][221] 속씨식물은 백악기 동안 이러한 환경에 주로 서식하며 작은 초본 식물의 생태적 지위를 차지했을 수 있다.[117][219] 이는 초기 속씨식물의 중요성을 제한했을 수 있지만, 동시에 다른 다양한 환경으로 빠르게 적응하고 퍼져나갈 수 있는 유연성을 제공했을 것으로 보인다.[118][221] 2017년에는 약 1억 4천만 년 전에 살았던 것으로 추정되는 최초의 꽃에 대한 상세한 3D 복원 연구 결과가 발표되기도 했다.[119][120]
13. 육상으로의 진출 (일본어 문서 추가 내용)
육상 식물은 약 5억 1000만 년 전에 녹조류로부터 진화했을 것으로 추정된다[158]. 현존하는 육상 식물과 가장 가까운 녹조류는 차축조 식물문이며, 특히 차축조류이다. 만약 차축조류의 생태가 당시와 크게 다르지 않다고 가정한다면, 육상 식물의 조상은 가지를 치는 실 모양의 반수체( haplontic영어 ) 조류였을 것이다. 이들은 얕은 민물[159], 아마도 계절적으로 마르는 작은 웅덩이 가장자리에서 서식했을 것으로 보인다[158]. 또한, 균류와의 공생 관계가 초기 식물이 육상 환경에 적응하는 데 도움을 주었을 가능성이 있다[158][160].
육상 식물이 육상 최초의 광합성 생물은 아니었다. 암석 풍화 속도 연구에 따르면, 생명체는 이미 12억 년 전에 육상에 존재했을 수 있으며[158], 10억 년 전의 담수호 퇴적물에서는 미생물 화석도 발견되었다[161]. 그러나 탄소 동위원소 기록은 이들 생물이 8억 5천만 년 전까지 대기 조성에 큰 영향을 미칠 정도는 아니었음을 시사한다[162]. 이 초기 육상 생물들은 아마도 작고 단순했을 것이며, 조류 피막(algal scum영어) 정도의 형태였을 것으로 추정된다[158].
육상 식물의 가장 오래된 증거는 약 4억 7천만 년 전 중기 오르도비스기의 암석에서 발견된 4분자 포자(tetrads)이다[163][164][165]. 4분자 포자는 동일한 4개의 포자가 입체적으로 연결된 형태로, 하나의 세포가 감수 분열을 거치면서 만들어진다. 이는 모든 육상 식물과 일부 조류에서 나타나는 특징이다[163]. 이 초기 포자들의 미세 구조는 현생 선태식물(특히 우산이끼류)의 포자와 매우 유사하여, 비슷한 수준의 조직 단계를 가졌음을 시사한다[166]. 포자 벽에는 스포로폴레닌이라는 매우 부패에 강한 물질이 포함되어 있어 화석으로 잘 보존될 수 있었으며, 이는 이들이 배아식물과 관련이 있음을 뒷받침한다[17].
관다발 식물과 유사한 삼렬 포자(trilete spores영어)는 그 직후인 약 4억 5500만 년 전 상부 오르도비스기 암석에서 나타난다[18][19][169]. 4분자 포자가 분리될 때, 각 포자에는 Y자 모양의 "삼렬 흔적"(trilete mark영어)이 생길 수 있는데, 이는 포자가 서로 맞닿아 눌렸던 자국이다[163][20]. 이러한 흔적이 생기려면 포자벽이 초기 단계부터 튼튼하고 저항력이 있어야 한다. 이는 건조에 대한 저항성과 밀접한 관련이 있으며, 물 밖 환경에서 생존하는 데 필수적인 특징이다. 실제로 수중 환경으로 되돌아간 일부 배아식물은 저항성 포자벽이 없어 삼렬 흔적을 가지지 않는다[20][163]. 조류 포자 중에서는 삼렬 흔적을 가진 것이 발견되지 않는데, 이는 포자벽이 충분히 튼튼하지 않거나, 흔적이 생기기 전에 흩어지거나, 정사면체 형태의 4분자를 만들지 않기 때문이다[20][163].
최초의 육상 식물 대형 화석은 실루리아기 초기 란도베리절의 엽상체 유기체로, 하천 습지에 서식하며 범람원의 넓은 지역을 덮었던 것으로 보인다. 이들은 땅이 물에 잠겼을 때만 생존할 수 있었다[21][170]. 당시 미생물 매트도 존재했다[22].
식물이 육상에 정착한 후, 건조 환경에 대처하는 두 가지 주요 전략이 나타났다. 현생 선태식물은 건조를 피하거나(습한 환경에 서식) 건조에 굴복하여(물이 다시 공급될 때까지 대사 활동 중단) 생존한다. 반면 관다발 식물은 물 손실을 조절하여 건조에 저항한다. 이들은 공기에 노출된 표면에 방수성 큐티클 층을 발달시켰고(일부 선태식물도 가짐), 가스 교환을 위해 여닫을 수 있는 기공을 진화시켰다. 또한 물 수송을 위한 관다발 조직을 발달시켜 체내 물 이동을 효율화하고, 이를 통해 물의 부력 없이도 수직 성장이 가능해져 더 큰 식물로 진화할 수 있는 기반을 마련했다. 생활사에서도 배우자체 우세에서 포자체 우세로 변화했다.
육상 식물 군락이 확산되면서 광합성 결과로 대기 중 산소 농도가 크게 증가했다. 산소 농도가 약 13%를 넘어서면서 산불 발생 가능성이 생겨났으며, 이는 실루리아기 전기의 탄화된 식물 화석을 통해 확인된다[171]. 데본기 말기의 일부 공백기를 제외하면, 탄화된 식물 화석은 이후 지질 시대에서도 계속 발견된다. 탄화 작용은 중요한 화석화 과정 중 하나로, 산불로 인해 식물의 연한 부분이 제거되고 단단한 탄소 껍질만 남아 보존되는 방식이다. 이 탄화된 화석은 초식동물이나 부패 미생물에게 이용되지 않고 압력에도 잘 견뎌 보존 상태가 좋으며, 때로는 세포 수준의 미세 구조까지 관찰할 수 있게 해준다.
14. 생체 내 수분 수송
광합성을 위해 식물은 대기 중의 CO2를 흡수해야 하지만, 이 과정에서 수분을 잃게 된다. 기공이 열려 CO2를 흡수하는 동안 수분이 증발하기 때문이다.[174] 수분은 CO2보다 훨씬 빠르게 손실되므로 식물은 이를 보충해야 했다. 이를 위해 식물은 습한 토양에서 광합성이 일어나는 부위까지 물을 운반하는 시스템을 발달시켰다.[174]
초기 육상 식물은 세포벽 사이의 공간을 통해 아포플라스트 방식으로 물을 흡수하고 수송했다.[44] 이후 식물은 수분 손실을 조절하기 위해 몇 가지 중요한 구조를 진화시켰다. 첫째, 물 손실을 줄이기 위해 방수성 외부 덮개인 큐티클을 발달시켰다. 둘째, 기공을 진화시켜 CO2 흡수와 수분 증발 속도를 조절할 수 있게 되었다.[174][178] 셋째, 광합성 조직 내 세포 간 공간을 발달시켜 CO2의 내부 확산을 효율적으로 만들었다.[45] 이러한 시스템은 식물이 체내 수분 함량을 일정하게 유지하는 데 도움을 주었으며(항상성), 특히 물 공급이 불규칙한 환경에서 유리했다.[45] 방수성 표피(큐티클)의 진화는 식물이 항상 수분층에 접촉하지 않고도 생존할 수 있게 했으며, 수분 상태에 따라 신진대사가 변하는 변온성(poikilohydry) 식물에서 체내 수분을 일정하게 유지하는 등온성(homoiohydry) 식물로의 변화는 새로운 서식지로 진출할 가능성을 열었다.[174]
식물이 처음 육지에 정착했던 실루리아기와 초기 데본기에는 대기 중 CO2 농도가 매우 높았기 때문에, 식물은 비교적 적은 양의 물 손실로도 충분한 CO2를 얻을 수 있었다.[44][174] 그러나 식물의 광합성 활동으로 대기 중 CO2 농도가 점차 감소하면서, 동일한 양의 CO2를 얻기 위해 더 많은 물을 증발시켜야 했고, 이는 더 정교한 물 획득 및 수송 메커니즘의 진화를 촉진했다.[44][174] CO2 농도가 현재 수준과 비슷해진 석탄기 말에는 CO2 흡수 단위당 약 17배 더 많은 물이 손실되었다.[44][174] CO2가 풍부했던 초기에도 물은 귀중했으며, 건조를 피하기 위해 젖은 토양에서 식물 전체로 물을 효율적으로 수송하는 것이 중요했다.[45]
초기 식물의 수분 수송은 물 분자 간의 응집력과 표면과의 부착력에 기반한 모세관 현상과 응집력-장력 메커니즘을 이용했다. 식물 표면에서 물이 증발하면, 물기둥 내의 물 분자들이 서로 끌어당기며 위로 딸려 올라간다. 따라서 증발만으로 식물에서 물 수송의 구동력을 제공했다.[44][174] 그러나 특수한 수송 혈관이 없으면 이 메커니즘은 물 수송 세포를 붕괴시킬 만큼 강한 음압(-)을 유발할 수 있어 물 수송 거리가 수 cm 이내로 제한되었고, 초기 식물의 크기도 작을 수밖에 없었다.[44][174] 건조 시 물기둥이 끊어지는 것을 막기 위해 식물은 방수성 큐티클을 발달시켰다. 초기 큐티클은 기공이 없었지만 식물 전체를 덮지 않아 가스 교환이 가능했다.[174] 건조한 시기에는 세포벽 안에 물을 저장하고 생명 활동을 일시적으로 "보류"하며 견뎌냈다.[174]
크기 제한과 습한 환경 의존성에서 벗어나기 위해 식물은 더 효율적인 수분 수송 시스템을 진화시켰다. 실루리아기 초기부터 리그닌과 같은 단단한 물질이 침착된 특수한 수송 세포가 발달했다.[174] 이 세포들은 성숙하면서 내부 물질이 사라지고 세포벽만 남아 속이 빈 관 형태가 되어 물이 쉽게 흐를 수 있게 했다. 이 과정에서 세포는 죽게 된다.[174] 이러한 세포들은 일반 세포 사이로 물이 이동하는 것보다 훨씬 효율적으로(약 100만 배) 물을 통과시켰으며, 먼 거리까지 물을 수송하고 높은 CO2 확산 능력을 가능하게 했다. 수분 수송을 위한 특수 조직은 하이드로이드(hydroid), 가도관(tracheid), 내피로 지지되는 2차 목부, 최종적으로 도관(vessel element) 순서로 진화했다.[174]
초기 데본기의 ''아글라오피톤''(Aglaophyton)과 ''호르네오피톤''(Horneophyton)과 같은 전관속 식물 화석에서는 현대 선태식물의 하이드로이드와 유사한 초기 수분 수송 세포 구조가 발견된다.[174] 이후 물의 흐름을 개선하기 위해 세포벽에 나선형 또는 고리 모양의 비후(thickening)가 발달한 관(띠 모양 관, banded tube)이 실루리아기 초기에 나타났다.[175][179] 이러한 관들은 벽에 작은 구멍(피트, pit) 무늬가 있었으며 리그닌화되었다.[176] 이들이 단세포로 구성된 물관, 즉 가도관을 형성하면서 진정한 '''관다발 식물'''이 등장한 것으로 여겨진다. 가도관은 하이드로이드보다 더 견고하여 높은 수압을 견딜 수 있었다.[174] 관다발 조직은 뿔이끼류에서 처음 발달하여 모든 관다발 식물로 이어졌을 가능성도 있지만, 여러 번 독립적으로 발생했을 수도 있다.[177][174]
수분 수송 조절에는 기공이 중요한 역할을 한다. 기공 개폐를 통해 가스 교환량과 함께 수분 증발량을 조절할 수 있다.[178] 기공은 관다발 조직보다 먼저 진화했으며, 관다발 식물이 아닌 뿔이끼류에도 존재한다.[174]
뿌리의 내피는 아마도 실루리아기-데본기 사이에 진화했을 것으로 추정되지만, 명확한 화석 증거는 석탄기 이후에 나타난다.[44][174] 내피는 뿌리의 중심부에 있는 수분 수송 조직(관다발)을 둘러싸며, 토양과 관다발 사이의 이온 교환을 조절하고 병원균 침입을 막는다. 또한 증산 작용이 약할 때 뿌리 압력을 발생시켜 물을 위로 밀어 올리는 역할을 하기도 한다.
이러한 수분 수송 및 조절 시스템이 발달하면서 식물은 표면의 습기에 의존하지 않고 뿌리를 통해 물을 흡수하여 체내 수분 상태를 일정하게 유지하는 등온성(homoiohydry)을 획득하게 되었다. 이는 식물이 더 건조한 환경으로 진출하고 큰 크기로 성장하는 것을 가능하게 했다.[174] 하지만 환경으로부터 독립성이 커진 대가로, 극심한 건조 상태에서 생존하는 능력(변온성, poikilohydry)은 점차 잃게 되었다.[174]
데본기 동안 가도관의 최대 직경은 증가했지만, 최소 직경은 크게 변하지 않았다.[179] 중기 데본기에는 일부 식물 계통에서 가도관 직경이 안정화되는 경향을 보였다.[179] 가도관 직경이 굵을수록 물은 더 빨리 흐르지만, 전체적인 수송률은 물관부 전체 단면적에 따라 달라진다.[179] 가도관 직경 증가는 식물 축의 굵기 및 높이 증가와 관련이 있으며, 수분 요구량을 늘리는 잎의 출현 및 기공 밀도 증가와도 밀접하게 연관되어 있다.[179][174]
굵고 튼튼한 벽을 가진 가도관은 더 높은 수압을 견딜 수 있게 해주었지만, 직경이 커지면서 공동 현상(cavitation)의 위험도 커졌다.[174] 공동 현상은 물관 내부에 공기 방울(색전, embolism)이 생겨 물 분자 기둥의 연속성이 끊어지는 현상으로, 응집력-장력 메커니즘에 의한 물 흡수를 방해한다. 가도관에 일단 공동 현상이 발생하면 일반적으로 그 기능을 회복하기 어렵다 (일부 피자식물은 회복 기작을 발달시켰다).[174] 따라서 식물에게 공동 현상을 피하는 것은 매우 중요하다. 이를 위해 가도관 벽의 피트는 공기가 유입되어 기포가 형성되는 것을 막기 위해 직경이 매우 작다.[174] 동결과 해동은 공동 현상의 주요 원인 중 하나이며, 가도관 벽 손상 역시 공기 유입을 유발할 수 있다. 따라서 여러 개의 가도관이 병렬로 작동하는 것이 안전성을 높이는 데 중요하다.[174]
공동 현상을 완전히 피하기는 어렵기 때문에, 식물은 발생한 공동 현상에 대처하는 몇 가지 메커니즘을 발달시켰다.[174] 작은 피트는 인접한 도관을 연결하며 액체는 통과시키지만 기체는 차단하여 공동 현상의 확산을 막는다. 역설적이게도, 색전 확산을 막는 피트 자체가 때로는 색전 발생의 원인이 되기도 한다.[174] 침엽수는 쥐라기에 이르러 공동 현상이 발생한 부분을 격리하는 밸브와 같은 정교한 구조(유연벽공, bordered pit)를 발달시켰다.[174] 참나무와 같은 일부 식물은 매년 봄에 새로운 굵은 도관을 만들어 겨울 동안 발생한 공동 현상 문제를 해결한다. 단풍나무는 뿌리 압력을 이용하여 수액을 위로 밀어 올려 기포를 짜낸다.[174]
초기 식물에서 가도관은 기계적 강도가 부족하여 식물체 중심부에 위치하고 주변의 튼튼한 후벽 조직에 의해 지지받았다.[174] 이후 리그닌화된 가도관과 도관이 발달하면서 수분 수송뿐 아니라 구조적 지지 역할도 수행하게 되었다.
가도관은 단일 세포로 구성되어 길이가 제한되고, 세포벽으로 막혀 있어 물의 흐름에 상당한 저항이 발생한다.[179] 유효 직경도 약 80 μm로 제한된다.[174] 도관은 여러 개의 세포(도관 요소)가 세로로 연결되어 형성되며, 세포 사이의 종단벽에 구멍(천공판, perforation plate)이 뚫려 있어 마치 하나의 긴 관처럼 작동한다.[179] 이는 가도관의 길이 및 직경 제한(최대 500 μm, 길이 10 m 이상 가능)을 극복하고 수분 수송 효율을 크게 높였다.[174]
도관이 처음 진화한 것은 건조하고 CO2 농도가 낮았던 페름기 말이었다. 속새류, 양치류, 부처손류에서 독립적으로 진화했으며, 이후 백악기 중기에 피자식물과 구네툼강(Gnetopsida)에서 다시 독립적으로 진화했다.[174] 도관은 동일한 단면적에서 가도관보다 약 100배 더 많은 물을 운반할 수 있다.[174] 이는 식물이 줄기에 더 많은 구조적 섬유를 포함시켜 더 튼튼하게 자라거나, 물 수송 효율을 바탕으로 덩굴식물과 같은 새로운 생태적 지위를 개척하는 것을 가능하게 했다.[174] 그러나 도관은 공동 현상을 피하기 위해 구조적으로 더 강화되어야 하며, 가도관으로 이루어진 목재보다 무겁고 형성하는 데 더 많은 자원이 필요하다는 단점도 있다.[174]
15. 잎의 진화

현재 식물의 잎은 대부분 광합성을 위해 태양 빛을 최대한 많이 포착하도록 적응된 기관이다.[53][62] 잎은 식물 진화 역사상 여러 번 독립적으로 진화했으며, 그 기원 중 하나는 초식동물로부터 자신을 보호하기 위한 바늘 모양의 돌기(enation)였을 가능성이 있다.
스코틀랜드의 라이니 처트에서 발견된 초기 육상 식물인 ''Rhyniophyta''는 장식 없는 가는 축으로만 이루어져 있었다. 이후 데본기 전기에서 후기에 걸쳐 나타난 ''Trimerophytina''에서 잎과 유사한 구조의 첫 증거가 나타난다. 이들은 가지 끝에 포자낭 덩어리가 달린 형태를 가졌다.[178] ''프실로피톤''과 같은 일부 초기 식물은 관다발이 없는 작은 가시 모양의 돌기를 가지고 있었다.
비슷한 시기에 ''Zosterophyllophyta''도 등장했다. 이들은 축에 가까운 짧은 곁가지에 콩팥 모양의 포자낭이 달리는 특징이 있으며, 때로는 H자 모양으로 분지하기도 했다.[178] 대부분 축에 뚜렷한 돌기를 가졌지만, 이 돌기에는 관다발이 없었다. 관다발이 있는 돌기의 가장 오래된 증거는 후기 실루리아기의 화석 석송류인 ''바라과나티아''(''Baragwanathia'')에서 발견된다.[181] 이 식물의 잎에는 잎맥이 중앙을 관통하고 있었다.[182] 라이니 처트에서 발견된 ''아스테록실론''(''Asteroxylon'')은 ''바라과나티아''보다 약 2천만 년 뒤의 식물이지만, 역시 원시적인 관다발(중심주에서 각 잎으로 뻗어 나가는 잎맥 흔적)을 가지고 있었다.[67] "돌기 이론"(enation theory)은 이러한 돌기에 관다발이 연결되면서 소엽이 발달했다고 설명한다. 다른 가설로는 축의 가지가 그물처럼 합쳐지면서 소엽이 진화했을 가능성도 제기된다.[178]
''아스테록실론''[183]과 ''바라과나티아''는 원시적인 석송류(lycopods)로 여겨지며,[178] 이들의 후예인 석송류는 물부추( ''Isoetes'' ), 부처손( ''Selaginella'' ), 석송( ''Lycopodium'' ) 등으로 현재까지 존재한다.[1] 석송류는 단 하나의 관다발(잎맥)을 가진 특징적인 소엽을 갖는다. 소엽은 크기가 다양하여, 멸종한 인목류(Lepidodendrales)의 경우 길이가 1m가 넘는 것도 있었지만, 거의 모두 단 하나의 잎맥만을 가진다. 일부 ''Selaginella'' 종에서 드물게 잎맥이 분지하는 것이 예외이다.
우리가 흔히 '잎'이라고 부르는 대엽(megaphyll)은 소엽과는 다른 기원을 가지며, 양치식물, 속새류, 전나자식물, 그리고 종자식물에서 독립적으로 네 번 이상 진화한 것으로 생각된다.[184][68] 대엽은 텔롬설에 따르면, 원래 3차원적으로 갈라지던 가지(텔롬)들이 몇 가지 단계를 거쳐 진화했다.[55][182] 주요 단계는 다음과 같다:
- 겹쳐지기 (Overtopping): 가지 중 하나가 다른 가지보다 우세하게 자라 주축을 형성하고 나머지는 곁가지가 된다.
- 편평화 (Planation): 3차원적인 가지들이 하나의 평면상에 배열된다.
- 그물화 또는 융합 (Webbing/Fusion): 평면화된 가지들 사이의 공간이 엽육 조직으로 채워져 넓은 잎 모양(엽신)을 형성한다.
이러한 과정은 오늘날 잎의 진화에서 여러 번 일어난 것으로 보인다.[56] 잎의 관다발이 줄기의 관다발에서 갈라져 나가는 지점인 엽극의 구조가 줄기에서 가지가 갈라져 나가는 모습과 유사하다는 점이 텔롬설의 증거로 제시된다.[182][66]
텔롬설은 화석 증거에 의해 널리 지지받지만, 볼프강 하게만(Wolfgang Hagemann) 등 일부 학자들은 형태학적, 생태학적 이유로 의문을 제기하며, 초기 육상 식물이 평평한 엽상체였고 축(줄기, 뿌리)이 나중에 진화했다는 대안 이론을 제시하기도 했다.[57][58] 롤프 자틀러(Rolf Sattler)는 두 이론을 통합하여, 평평한 구조와 원통형 구조 사이의 연속적인 변화 과정을 고려하는 관점을 제안했으며,[59][60] 이는 분자 유전학 연구(예: KNOX 유전자 발현 시점 차이)에 의해서도 뒷받침된다.[61]
대엽을 진화시킨 네 그룹은 모두 후기 데본기에서 전기 석탄기 사이에 처음으로 대엽을 발달시켰고, 석탄기 중기에 이르러 그 구조가 안정화될 때까지 빠르게 다양해졌다.[184][68]
잎이 진화하기까지 왜 오랜 시간이 걸렸는지에 대한 의문이 제기된다. 식물이 육상에 출현한 후 대엽이 보편화되기까지 약 5천만 년이 걸렸다. 하지만 전기 데본기의 ''에오필로피톤''(''Eophyllophyton'')에서 이미 작고 드문 중간 크기의 잎(mesophyll)이 발견되므로, 발달 과정 자체가 장벽은 아니었을 것이다.[185] 가장 유력한 설명은 이 기간 동안 대기 중 이산화 탄소(CO2) 농도가 급격히 감소했다는 것이다. 데본기 동안 CO2 농도는 약 90% 감소했으며,[186][70] 식물은 광합성 효율을 유지하기 위해 기공 밀도를 100배 가량 늘려야 했다.[53][62] 기공은 가스 교환에 필수적이지만, 물의 증발을 유발하여 잎을 냉각시키는 효과도 있다. 초기 데본기의 낮은 CO2 농도와 낮은 기공 밀도 하에서는 큰 잎이 태양열에 의해 쉽게 과열될 수 있었다. 당시 식물의 원시적인 중심주 구조와 제한적인 뿌리 시스템은 증가된 증산량을 감당할 만큼 충분한 물을 빠르게 공급하기 어려웠기 때문에, 기공 밀도를 높이고 잎의 크기를 키우는 데 제약이 있었을 것이다.[187][62][63]
잎이 항상 유리한 것만은 아니어서, 이차적으로 잎을 잃는 경우도 자주 발생한다. 대표적인 예로 선인장과 원시 양치류의 일종인 솔잎란( ''Psilotum'' )이 있다. 이러한 이차적 진화는 잎의 본래 기원을 파악하기 어렵게 만들기도 한다. 예를 들어, 속새과(Equisetaceae) 식물의 잎은 단 하나의 잎맥만 가지고 있어 소엽처럼 보이지만, 화석 기록과 분자 유전학적 증거에 따르면 이들의 조상은 복잡한 잎맥을 가진 대엽을 가졌으며 현재의 단순한 구조는 이차적으로 진화한 결과이다.[188][66]
낙엽수는 잎을 가짐으로써 발생하는 또 다른 불리함, 즉 기후 조건에 대응한다. 겨울철의 강한 바람이나 쌓이는 눈의 무게는 잎이 없을 때 식물이 견디기 더 수월하다. 일반적으로 식물이 낮 길이가 짧아지면 잎을 떨어뜨린다고 알려져 있지만, 이는 완전히 정확하지는 않다. 최근의 온실 지구 시기에는 북극권에서도 상록수가 번성했다.[189] 계절에 따라 잎을 떨어뜨리는 낙엽성은 은행나무, 일부 구과식물, 그리고 특정 피자식물 등 여러 식물 그룹에서 독립적으로 여러 번 진화했다.[190][72] 또한 잎을 떨어뜨리는 것은 곤충에 의한 피해 압력에 대한 대응책으로 진화했을 수도 있다. 손상된 잎을 계속 유지하고 수리하는 것보다 겨울이나 건기 동안 잎을 완전히 새로 만드는 것이 생존에 더 비용 효율적이었을 수 있다.[191][73]
16. 목질의 진화
초기 데본기의 풍경에는 사람 허리 높이보다 큰 식물이 없었다. 식물이 더 높이 자라기 위해서는 튼튼한 관다발 조직이 필수적이었다. 더 높은 키는 광합성을 위한 햇빛 확보, 다른 식물과의 경쟁, 그리고 포자나 씨앗을 더 멀리 퍼뜨리는 데 유리했기 때문에[192], 높이 성장에 대한 진화적 압력은 꾸준히 존재했다. 나무처럼 커지기 위해 초기 식물들은 지지 기능과 물 수송 기능을 모두 갖춘 목부 조직, 즉 목질을 발달시켜야 했다.
2차 생장(비대 생장)을 하는 식물의 줄기 중심부에는 관다발 형성층이라는 세포층이 고리 모양으로 존재한다. 이 형성층은 안쪽으로 물관부(목부)를, 바깥쪽으로 체관부(사부)를 만들어낸다. 목부 세포는 죽어서 단단한 리그닌 조직이 되는데, 해마다 새로운 목부 세포층이 기존 층에 겹겹이 쌓이면서 단단한 목질 줄기를 형성한다. 목질의 가장 단순한 형태는 약 4억 년 전, 모든 육상 식물이 작았던 초기 데본기에 처음 나타난 것으로 보인다.[86] 목질은 나무나 관목보다 훨씬 먼저 진화했기 때문에, 처음에는 물 수송이 주된 기능이었고 지지 기능은 나중에 추가되었을 가능성이 높다.[87]
처음으로 2차 생장 능력을 갖추고 나무처럼 자라기 시작한 식물은 대엽 양치식물로 보인다. 데본기 중기에는 클라도시론류(Cladoxylopsida)에 속하는 ''Wattieza''가 이미 8m 높이까지 자라는 나무 형태를 갖추었다.[88][193]
다른 식물 그룹들도 곧이어 나무와 같은 키를 갖추게 되었다. 데본기 후기에는 트리메로파이트에서 진화한 원시겉씨식물(progymnosperm)인 ''Archaeopteris''가 30m 높이까지 자랐다.[89][194] 이들은 양면 관다발 형성층에서 만들어진 진정한 목질을 발달시킨 최초의 식물 그룹이다. 그중 ''Rellimia''는 데본기 중기에 이미 등장했다.[90][195] 진정한 목질은 단 한 번만 진화한 것으로 여겨지며, 이 특징을 공유하는 식물들을 묶어 목질식물(Lignophyta)이라는 분기군으로 분류한다.
''Archaeopteris''가 만든 숲에는 곧이어 석송류(Lycopodiophyta) 나무들이 들어섰다. 특히 인목(Lepidodendrales)에 속하는 레피도덴드론(Lepidodendron) 등은 높이가 50m 이상, 줄기 밑동 지름이 2m를 넘기도 했다. 이 거대한 석송류는 데본기 후기와 석탄기의 숲을 지배하며 오늘날 석탄층 형성의 주요 원인이 되었다.[91][196] 하지만 이들 석송류는 현대의 나무와 달리 "유한 생장"을 했다. 즉, 어릴 때 영양분을 저장했다가 유전적으로 정해진 특정 높이까지 단번에 자란 뒤, 가지를 뻗어 포자를 퍼뜨리고 죽는 방식이었다.[92][197] 이들은 빠른 성장을 위해 상대적으로 가볍고 약한, 이른바 "값싼" 목질을 가졌는데, 줄기 내부의 상당 부분이 부드러운 조직으로 채워진 빈 공간이었다.[1][178] 또한 이들의 목질은 새로운 사부를 만들지 못하는 단면 형성층에서 만들어졌기 때문에, 줄기가 해마다 굵어지지는 못했다.
석탄기에는 속새류의 일종인 롭속새(Calamites)가 나타났다. 현대의 속새속(Equisetum)과 달리 롭속새는 관다발 형성층을 가지고 있어 목질을 만들고 10m가 넘는 높이까지 자랄 수 있었으며, 여러 번 가지를 쳤다.
이러한 초기 나무들의 겉모습은 오늘날의 나무와 비슷했지만, 현재 지구상 나무의 대부분을 차지하는 종자식물 그룹은 아직 진화하지 않은 상태였다. 오늘날 숲을 이루는 주요 나무 그룹은 겉씨식물(침엽수 등)과 속씨식물(꽃 피는 식물)이며, 이들은 모두 종자식물에 속한다. 오랫동안 속씨식물이 겉씨식물에서 진화했다고 여겨졌으나, 최근의 분자 유전학 연구 결과는 이 두 그룹이 실제로는 별개의 분기군이며, 아마도 페름기 초기에 공통 조상인 종자고사리류(Pteridospermatophyta) 내에서 갈라져 나왔을 가능성을 시사한다.[93][94][95][198][199][200][116][218] 다만, 이러한 분자 데이터 해석은 아직 형태학적 증거와 완전히 일치하지는 않으며[96][97][98][201][202][203], 기존의 형태학적 분석 결과 역시 두 그룹의 관계를 명확히 뒷받침하지는 못한다는 점이 지적된다.[116][218]
속씨식물과 그 조상 그룹은 백악기에 폭발적으로 다양해지기 전까지는 식물계에서 큰 비중을 차지하지 못했다. 초기에는 작고 습한 환경을 선호하는 식물이었으나, 백악기 중반 이후 급격히 다양화하여[99] 오늘날에는 타이가 지역을 제외한 전 세계 숲의 가장 중요한 구성원이 되었다.
17. 뿌리의 진화
| 석탄기 Lepidodendrales를 포함한 석송류의 근경(Stigmaria) (아래 이미지)은 다른 종의 표본에서 "잎 자국"과 "뿌리 자국"의 유사한 외관이 나타나듯이 줄기(위 이미지)와 발달적으로 동일한 것으로 생각된다.[85] |
뿌리는 식물에게 두 가지 중요한 기능을 수행한다. 첫째는 식물을 땅에 단단히 고정하는 것이고, 둘째는 토양으로부터 물과 영양분을 흡수하는 것이다. 뿌리 덕분에 식물은 더 높고 튼튼하게 자랄 수 있다.
뿌리의 출현은 지구적 규모로 영향을 미쳤다. 뿌리는 토양을 교란하고, 질산염이나 인산염 같은 영양분을 흡수하여 토양의 산성화를 촉진했다.[74] 이 과정에서 토양이 더 깊게 풍화되었고, 탄소 화합물이 토양 깊숙이 주입되었다.[75] 이는 결과적으로 기후에 큰 영향을 미쳤으며,[76] 심지어 후기 데본기 대멸종과 같은 대량 멸종의 원인이 되었을 수도 있다.[77]
뿌리가 언제 어떻게 처음 진화했는지는 불분명하다. 후기 실루리아기의 화석 토양에서 뿌리 모양의 흔적이 발견되기도 하지만,[78] 초기 식물의 몸체 화석들을 보면 뿌리가 없었다는 것을 알 수 있다. 많은 초기 식물은 땅 위나 아래로 뻗는 줄기를 가졌고, 일부는 광합성을 하지 않고 기공이 없는 지하 줄기를 가지기도 했다. 진정한 뿌리는 줄기와 발생 과정이 다르며, 분기하는 방식도 다르고 끝에 뿌리골무를 가지고 있다는 점에서 구분된다.[85] 실루리아기에서 데본기에 살았던 ''Rhynia''나 ''Horneophyton'' 같은 식물들은 생리적으로 뿌리와 같은 기능을 하는 구조를 가졌지만,[79][80] 뿌리골무가 있는 진정한 의미의 뿌리는 아니었다.[85] 뿌리는 화석으로 잘 보존되지 않기 때문에 그 기원과 진화에 대한 정보는 제한적이다.[85]
뿌리의 기능을 일부 수행하는 작은 구조인 가근(rhizoid)은 아마도 매우 일찍 진화했을 것으로 추정되며, 식물이 육지로 올라오기 전부터 존재했을 가능성도 있다. 육상 식물의 조상뻘인 차축조류(Characeae)에서도 가근이 발견된다.[85] 가근은 한 번만 진화한 것이 아니라 여러 차례 독립적으로 나타난 것으로 보인다. 예를 들어, 지의류의 가근(rhizine)이나 일부 동물(관벌레 ''Lamellibrachia'' 등)이 가진 뿌리 모양 구조도 비슷한 역할을 한다.[85] 라이니 차트(Rhynie chert) 화석에서는 가근이 뚜렷하게 관찰되며, 초기 관다발 식물 대부분에서 발견되어 진정한 뿌리의 진화를 예고한 것으로 여겨진다.[81]
더 발달된 뿌리 형태는 라이니 차트를 비롯한 초기 데본기 화석들에서 나타난다.[85] 리니아 식물은 미세한 가근을 가졌지만, 라이니 차트의 트리멜로피톤류나 초본 석송류는 토양에 몇 센티미터 파고드는 뿌리 같은 구조를 가지고 있었다.[100] 그러나 이들 역시 현생 식물의 뿌리가 가진 모든 특징을 갖추지는 못했다.[85] 최근 연구에 따르면 ''Asteroxylon''은 현존하는 관다발 식물의 뿌리와는 독립적으로 진화한 뿌리를 가진 것으로 밝혀졌다.[82] 데본기 동안 뿌리와 뿌리 유사 구조는 점점 보편화되었고 더 깊이 땅속으로 파고들었다. 석송류 나무는 중기 데본기(아이펠기, 지베티기)에 20cm 길이의 뿌리를 내렸고, 이후 후기 데본기(프라니기)에는 전(前)나자식물이 약 1m 깊이까지 뿌리를 뻗었다.[100] 진정한 나자식물과 양치류의 일종인 ''Zygopteridaceae''는 후기 데본기(파메니안기)에 얕은 뿌리 시스템을 형성했다.[100]
석송류의 근경(Stigmaria)은 뿌리와는 다른 방식으로 땅속에 자리 잡았다. 근경은 기본적으로 줄기와 동일한 구조이며, 잎과 상동 관계인 기관이 뿌리 역할을 했다.[85] 비슷한 구조가 현생 석송류인 물부추속(''Isoetes'')에서도 발견되는데, 이는 뿌리가 석송류와 다른 식물 그룹에서 적어도 두 번 이상 독립적으로 진화했다는 증거로 여겨진다.[85] 석송류와 유엽식물에서 뿌리가 시작되고 자라는 방식이 다른 메커니즘에 의해 조절된다는 연구 결과도 이를 뒷받침한다.[83]
관다발 조직은 뿌리를 가진 식물에게 필수적이다. 광합성을 하지 못하는 뿌리에 당분을 공급하고, 뿌리에서 흡수한 물과 영양분을 식물 전체로 운반해야 하기 때문이다.[159] 뿌리 시스템을 갖춘 초기 식물들은 뿌리가 없던 실루리아기 조상들보다 약간 더 진화했지만, 땅에 평평하게 누운 줄기에서 오늘날 선태식물의 가근과 유사한 구조가 발달하는 것을 볼 수 있다.[84]
중기 데본기에서 후기 데본기까지 대부분의 식물 그룹들은 각기 다른 방식으로 뿌리 조직을 독립적으로 발달시켰다.[84] 뿌리가 커지고 발달하면서 더 큰 나무를 지탱할 수 있게 되었고, 토양은 더욱 깊이 풍화되었다.[77] 이러한 깊은 토양 풍화는 앞서 언급된 이산화탄소 변화에 영향을 주었을 뿐만 아니라, 균류와 동물이 정착할 수 있는 새로운 서식지를 제공했다.[100]
현대 식물의 뿌리는 물리적 한계까지 발달했다. 뿌리는 지하수면에 도달할 때까지 땅속 수 미터를 뻗어 나간다. 가장 가는 뿌리의 직경은 약 40 μm인데, 이보다 더 가늘어지면 물리적으로 물을 운반할 수 없게 된다.[85] 반면, 지금까지 발견된 가장 오래된 뿌리 화석의 직경은 3mm에서 700 μm 미만 정도이다. 다만, 화석화 과정(유해화석학)의 영향으로 관찰 가능한 뿌리 두께에는 한계가 있다.[85]
18. 아르부스큘러 균근
많은 식물의 뿌리는 공생 관계를 통해 효율을 높인다. 가장 일반적인 형태는 아르부스큘러 균근(AM)이다. '아르부스큘러 균근'은 문자 그대로 "수목형(Arbuscular) 균근(mycorrhizae)"을 의미하며, 뿌리의 일부를 침범하는 균에 의해 형성된다. 이 균은 뿌리 세포막 안을 균사로 채운다. 균은 식물로부터 당을 얻는 대신, 토양에서 흡수하거나 합성한 영양분, 특히 인산염을 식물에게 제공한다. 식물은 균이 없으면 이러한 영양분을 얻기 어려울 수 있다.
이러한 공생 관계는 식물의 역사 초기에 빠르게 진화한 것으로 보인다. 아르부스큘러 균근은 선태식물부터 양치식물, 겉씨식물, 속씨식물에 이르기까지 모든 주요 육상 식물군에서 발견되며, 현존하는 관다발 식물의 80% 이상이 이 관계를 맺고 있다.[209][41] 이는 조상 식물과 균의 공생이 식물의 육상 진출 단계에서 중요한 역할을 했을 가능성을 시사한다.[210] DNA 염기 서열 분석 결과는 아르부스큘러 균근 상호작용이 육상 식물군의 공통 조상이 육지로 전환하는 동안 발생했으며,[39] 심지어 육상 환경에 정착하는 데 결정적인 단계였을 수 있음을 보여준다.[40]
실제로 데본기의 라이니 차트 화석에서도 아르부스큘러 균근이 풍부하게 발견된다.[211] 라이니 셰일에서 발견된 뿌리가 없는 초기 관다발 식물인 ''아글라오피톤''(Aglaophyton) 화석에서도[35] 줄기 피질 세포 내에 아르부스큘러 균근을 형성한 증거가 발견되었다.[36] ''아글라오피톤''과 같은 실루리아기 및 데본기 초기의 다른 뿌리 없는 육상 식물들은 토양에서 물과 영양분을 얻기 위해 수지상 균근 곰팡이에 의존했을 수 있다.[37]
아르부스큘러 균근을 형성하는 균은 주로 글로메로미코타문(Glomeromycota)에 속하며,[38] 이들은 약 10억 년 전에 처음 나타났을 것으로 추정된다.[41] 이러한 협력 관계는 식물에 진정한 뿌리가 생기기 전에도 일어났으며,[41] 심지어 뿌리가 균근균의 생육에 좋은 조건을 제공하기 위해 진화했다는 가설도 제기된다.[212] 뿌리가 발달하기 전, 이 균들은 식물이 스스로 합성할 수 없는 유기 화합물을 받는 대가로, 토양으로부터 물과 인과 같은 무기 영양분을 식물이 흡수하도록 도왔을 것이다.[41]
19. 종자의 진화
초기 육상 식물은 오늘날의 고사리류와 비슷한 방식으로 번식했다. 포자가 발아하여 배우체라는 작은 개체를 만들고, 여기서 생성된 정자는 주변의 물기를 이용해 헤엄쳐 자신의 또는 다른 개체의 조란기(난자를 만드는 기관)에 도달하여 난자와 수정한다. 수정된 난자는 배가 되고, 이 배는 다시 포자체(우리가 보통 보는 식물 형태)로 자라난다.[213]
이러한 번식 방식은 정자가 이동하기 위해 물이 꼭 필요했기 때문에, 초기 식물들은 물가나 습한 지역에서만 살아갈 수 있었다. 식물들이 이러한 환경적 제약에서 벗어나 더 다양한 환경으로 진출할 수 있게 된 것은 이형 포자성(heterospory)이라는 새로운 특성 덕분이었다.
이형 포자성 식물은 이름 그대로 크기가 다른 두 종류의 포자, 즉 작은 소포자와 큰 대포자를 만든다. 소포자는 수컷 배우체인 소배우체로, 대포자는 암컷 배우체인 대배우체로 각각 발달한다. 이 시스템은 궁극적으로 종자(씨앗)의 탄생으로 이어졌다. 진화 과정에서 대배우체는 점차 하나의 거대한 포자만을 포함하게 되었고(원래 4개의 포자 중 3개는 퇴화), 포자낭(포자를 담는 주머니) 안에는 단 하나의 대포자만 남게 되었다.
이 대포자는 더 이상 외부로 방출되지 않고 포자낭 안에서 발아하여 대배우체로 성장하는데, 이 구조가 바로 배주(ovule)의 원형이 된다. 대배우체는 주피(integument)라는 보호층(나중에 씨껍질이 됨)에 둘러싸이게 되며, 이 주피와 내부 구조가 종자 부피의 대부분을 차지하게 된다. 한편, 소포자에서 발아한 소배우체는 바람 등에 의해 운반되어(산포) 대배우체 근처에 도달한 뒤, 건조에 약한 정자를 방출하여 수분하는 역할만 담당하게 되었다.[178]
소엽 식물(석송류)과 스페노필룸류 같은 일부 고대 식물 그룹은 거의 종자에 가까운 구조를 만들었다. 예를 들어, 화석으로 발견된 ''Lepidocarpon''이나 ''Achlamydocarpon'' 같은 소엽 식물의 대포자는 지름이 1cm에 달했으며, 영양 조직에 둘러싸인 채 모체 식물 안에서 대배우체로 발달했다. 하지만 이 구조는 외부와 완전히 차단되지 않은 작은 틈이 있었고, 내부의 포자를 감싸는 주심(nucellus)이 포자 전체를 덮지 못했기 때문에 진정한 의미의 종자라고 보기는 어렵다. 이 때문에 건조에 대한 저항력이 부족했고, 정자가 수정하기 위해 조직을 뚫고 들어갈 필요도 없었다.[178]
최초로 진정한 종자를 가진 식물은 고사리 씨앗류(Pteridospermatophyta)로 알려져 있다. 이 이름은 잎 모양이 고사리와 비슷해서 붙여졌지만, 실제 대엽 고사리 식물과의 계통 관계가 반드시 가까운 것은 아니다. 가장 오래된 종자 식물 화석은 후기 데본기 지층에서 발견되며, 이들은 전나자식물(Progymnosperm)이라는 그룹에서 진화한 것으로 여겨진다. 이 초기 종자 식물들은 나무 형태부터 덩굴성 관목까지 다양했으며, 이들을 낳은 전나자식물 역시 나무처럼 자라며 고사리 같은 잎을 가지고 있어 외형적으로 매우 비슷했다. 이들은 배주는 가지고 있었지만, 오늘날의 소나무처럼 구과(솔방울)를 만들거나 사과나무처럼 과실을 만들지는 않았다. 초기 종자의 진화 과정을 정확히 추적하는 것은 어렵지만, 비교적 단순한 구조의 트리멜로피톤류에서 시작하여 전나자식물의 일종인 아네우로피톤류(Aneurophytales) 등으로 이어지는 계통을 생각해 볼 수 있다.[178]
이러한 초기 종자의 기본 형태는 오늘날의 나자식물(겉씨식물)에게 이어졌다. 나자식물의 종자는 대부분 목질이나 육질의 구과 구조 안에 들어 있지만, 완전히 밀폐되어 있지는 않다(주목처럼 씨가 노출된 경우도 있다). 반면, 피자식물(속씨식물)은 심피(씨방)가 발달하여 종자를 완전히 감싸는 유일한 식물 그룹이다.
종자가 외부 환경과 완전히 차단되면서 휴면(dormancy)이라는 새로운 능력이 나타났다. 배는 씨껍질 안에서 건조나 추위로부터 보호받으며, 환경 조건이 좋아질 때까지 길게는 수년 동안 발아하지 않고 기다릴 수 있게 되었다. 후기 석탄기의 나자식물 종자 화석에서 배가 들어있는 것이 발견되었는데, 이는 수정 후 발아까지 상당한 시간이 걸렸음을 시사한다.[214] 이 시기는 지구가 점차 따뜻해지고 건조해지던 때와 일치하는데, 이는 휴면 능력이 건조한 기후에 적응하기 위한 전략으로 진화했을 가능성을 보여준다. 즉, 비가 충분히 내리는 습윤기가 올 때까지 발아를 늦추는 것이 생존에 유리했을 것이다.[214] 이러한 진화적 혁신 덕분에 식물은 이전에는 살기 어려웠던 건조한 산지 등으로 서식지를 넓혀 나갈 수 있었다.[214]
종자는 번식과 생존에 여러 가지 이점을 제공했다. 우선, 포자에 비해 크고 영양분을 저장하고 있어 더 멀리 퍼져나갈 수 있고(산포), 수정된 배의 생존 성공률을 높였다. 또한, 종자 안의 배와 영양분(배젖 등) 덕분에 묘목은 싹이 튼 직후 불리한 환경에서도 스스로 영양분을 만들 수 있을 때까지 버틸 수 있게 되었다.[213] 예를 들어, 건조한 땅에 떨어진 종자는 저장된 양분으로 빠르게 뿌리를 내려 지하수면에 도달할 수 있고,[213] 숲 속 그늘진 곳에 떨어진 종자는 햇빛을 충분히 받을 수 있는 높이까지 자랄 때까지 저장된 에너지를 사용할 수 있다.[213] 이러한 강력한 이점들 덕분에 종자 식물은 초기 지구 숲의 주요 구성원이었던 아케오프테리스와 같은 원시적인 나무들을 점차 대체하며 생태계의 주역으로 자리 잡게 되었다.[213]
참조
[1]
서적
Paleobotany and the evolution of plants
Cambridge University Press
1993
[2]
논문
Earth's earliest non-marine eukaryotes
https://www.research[...]
2011
[3]
논문
The late Precambrian greening of the Earth
2009
[4]
논문
"Elkinsia gen. nov., a Late Devonian gymnosperm with cupulate ovules"
[5]
서적
The origin and early diversification of land plants. A cladistic study
Smithsonian Institution Press
[6]
논문
Xyloglucan evolution and the terrestrialization of green plants
[7]
논문
Molecular evidence for the early colonization of land by fungi and plants
2001-08
[8]
간행물
Walcott 2009
[9]
논문
A genomic timescale of prokaryote evolution: insights into the origin of methanogenesis, phototrophy, and the colonization of land
[10]
논문
No support for the emergence of lichens prior to the evolution of vascular plants
2019
[11]
논문
Massive release of hydrogen sulfide to the surface ocean and atmosphere during intervals of oceanic anoxia
[12]
논문
Oxygen, animals and oceanic ventilation: An alternative view
[13]
논문
The impacts of land plant evolution on Earth's climate and oxygenation state – An interdisciplinary review
2020
[14]
논문
New evidence for land plants from the lower Middle Ordovician of Saudi Arabia
1996
[15]
논문
Early Middle Ordovician evidence for land plants in Argentina (eastern Gondwana)
2010
[16]
논문
Fragments of the earliest land plants
http://eprints.white[...]
[17]
논문
FTIR characterisation of the chemical composition of Silurian miospores (cryptospores and trilete spores) from Gotland, Sweden
https://hal-insu.arc[...]
2010
[18]
논문
Origin and Radiation of the Earliest Vascular Land Plants
https://biblio.ugent[...]
2009-04
[19]
논문
Baltica cradle of early land plants? Oldest record of trilete spores and diverse cryptospore assemblages; evidence from Ordovician successions of Sweden
2019
[20]
논문
The Microfossil Record of Early Land Plants: Advances in Understanding of Early Terrestrialization, 1970–1984
[21]
서적
2014-05-28
[22]
논문
Earliest fossil record of bacterial–cyanobacterial mat consortia: the early Silurian Passage Creek biota (440 Ma, Virginia, USA)
[23]
웹사이트
First Land Plants and Fungi Changed Earth's Climate, Paving the Way for Explosive Evolution of Land Animals, New Gene Study Suggest
http://science.psu.e[...]
2018-04-09
[24]
논문
Cryogenian Glacial Habitats as a Plant Terrestrialisation Cradle – The Origin of the Anydrophytes and Zygnematophyceae Split
Frontiers Media
2022-01-27
[25]
논문
The rise of atmospheric oxygen
2008-01-16
[26]
논문
Charcoal in the Silurian as evidence for the earliest wildfire
2004
[27]
논문
Evolution of alluvial mudrock forced by early land plants
https://www.science.[...]
2018-03-02
[28]
논문
Evolution of sexual reproduction: Importance of DNA repair, complementation, and variation
[29]
논문
Masking of mutations and the evolution of sex
[30]
논문
Selection is no more efficient in haploid than in diploid life stages of an angiosperm and a moss
[31]
논문
How green was ''Cooksonia''? The importance of size in understanding the early evolution of physiology in the vascular plant lineage
[32]
논문
New gametophytes from the Early Devonian Rhynie chert
[33]
논문
Life history biology of early land plants: deciphering the gametophyte phase
[34]
논문
The gametophyte, embryo, and young rhizome of ''Psilotum triquetrum'' (Swartz)
1939
[35]
논문
On Old Red Sandstone plants showing structure, from the Rhynie Chert Bed, Aberdeenshire. Part II. Additional notes on ''Rhynia gwynne-vaughani'', Kidston and Lang; with descriptions of ''Rhynia major'', n.sp. and ''Hornea lignieri'', n.g., n.sp
https://zenodo.org/r[...]
[36]
논문
4 hundred million year old vesicular-arbuscular mycorrhizae
[37]
논문
Plants and arbuscular mycorrhizal fungi: an evolutionary-developmental perspective
https://doi.org/10.1[...]
2008-09
[38]
논문
A new fungal phylum, the ''Glomeromycota'': phylogeny and evolution
http://journals.camb[...]
[39]
논문
Origin and diversification of endomycorrhizal fungi and coincidence with vascular land plants
1993
[40]
논문
Origin and diversification of endomycorrhizal fungi and coincidence with vascular land plants
[41]
서적
Mycorrhizal symbiosis
Academic Press
2008
[42]
논문
Mutualistic mycorrhiza-like symbiosis in the most ancient group of land plants
[43]
논문
Coevolution of roots and mycorrhizas of land plants
[44]
논문
Evolution of Water Transport and Xylem Structure
[45]
논문
The evolution of vascular land plants in relation to supracellular transport processes
1977
[46]
논문
The deepest divergences in land plants inferred from phylogenomic evidence
[47]
논문
A vascular conducting strand in the early land plant ''Cooksonia''
[48]
논문
Evidence for a Conducting Strand in Early Silurian (Llandoverian) Plants: Implications for the Evolution of the Land Plants
[49]
논문
Evidence for lignin-like constituents in Early Silurian (Llandoverian) plant fossils
[50]
논문
The Evolution of Tracheid Diameter in Early Vascular Plants and Its Implications on the Hydraulic Conductance of the Primary Xylem Strand
[51]
논문
Torus-Margo Pits Help Conifers Compete with Angiosperms
2005-12
[52]
웹사이트
Why Christmas trees are not extinct
http://www.physorg.c[...]
[53]
논문
Turning over a new leaf
2001
[54]
논문
Diverted development of reproductive organs: A source of morphological innovation in land plants
[55]
서적
Die Phylogenie der Pflanzen
Gustav Fischer Verlag
1959
[56]
논문
Evolution of leaf developmental mechanisms
[57]
문서
Hagemann, W. 1976. Sind Farne Kormophyten? Eine Alternative zur Telomtheorie.
[58]
논문
Towards an organismic concept of land plants: the marginal blastozone and the development of the vegetation body of selected frondose gametophytes of liverworts and ferns
[59]
논문
Process morphology: structural dynamics in development and evolution
[60]
문서
Sattler, R. 1998. On the origin of symmetry, branching and phyllotaxis in land plants. In: R.V. Jean and D. Barabé (eds) Symmetry in Plants. World Scientific, Singapore, pp. 775-793.
[61]
논문
'Tree and Leaf': A different angle
[62]
논문
Evolution of leaf-form in land plants linked to atmospheric CO2 decline in the Late Palaeozoic era
http://eprints.white[...]
[63]
웹사이트
A perspective on the CO2 theory of early leaf evolution
http://www.corante.c[...]
2011-03-08
[64]
논문
Morphologically complex plant macrofossils from the late Silurian of arctic Canada
2002
[65]
논문
The age of the earliest club mosses: the Silurian Baragwanathia flora in Victoria, Australia
http://geolmag.geosc[...]
2007-10-25
[66]
논문
The Science of Plant Morphology: Definition, History, and Role in Modern Biology
[67]
논문
Perithecial ascomycetes from the 400 million year old Rhynie chert: an example of ancestral polymorphism
[68]
논문
Evolution of developmental potential and the multiple independent origins of leaves in Paleozoic vascular plants
[69]
논문
Structure of the Earliest Leaves: Adaptations to High Concentrations of Atmospheric CO2
[70]
논문
Geocarb III: A Revised Model of Atmospheric CO2 over Phanerozoic Time
http://ajsonline.org[...]
2008-04-07
[71]
논문
Reconstructing a lost Eocene paradise: Part I. Simulating the change in global floral distribution at the initial Eocene thermal maximum
[72]
논문
The advantages of being evergreen
[73]
논문
Ninety-seven million years of angiosperm-insect association: paleobiological insights into the meaning of coevolution
[74]
논문
Plant growth-promoting bacteria as inoculants in agricultural soils
2015-12
[75]
논문
Middle to Late Paleozoic Atmospheric {{#if:{{{link|}}}|[Carbon dioxide}}}}CO2{{#if:{{{link|}}}|}} Levels from Soil Carbonate and Organic Matter
[76]
논문
GEOCARB II: A revised model of atmospheric {{#if:{{{link|}}}|[Carbon dioxide}}}}CO2{{#if:{{{link|}}}|}} over Phanerozoic time
[77]
논문
Late Devonian Oceanic Anoxic Events and Biotic Crises: "Rooted" in the Evolution of Vascular Land Plants?
https://www.geosocie[...]
2019-08-29
[78]
서적
Paleosols: their Recognition and Interpretation
Blackwell
[79]
웹사이트
Tracheophyta - 2
http://palaeos.com/p[...]
[80]
논문
EARLY EVOLUTION OF LAND PLANTS: Phylogeny, Physiology, and Ecology of the Primary Terrestrial Radiation
1998
[81]
논문
Bilaterally symmetric axes with rhizoids composed the rooting structure of the common ancestor of vascular plants
[82]
논문
Stepwise and independent origins of roots among land plants
[83]
논문
ASL/LBD Phylogeny Suggests that Genetic Mechanisms of Root Initiation Downstream of Auxin Are Distinct in Lycophytes and Euphyllophytes
[84]
논문
The origin and early evolution of plants on land
[85]
논문
Roots: evolutionary origins and biogeochemical significance
[86]
논문
A simple type of wood in two early Devonian plants
2011
[87]
뉴스
'Early wood' samples reshape plant history
https://www.bbc.co.u[...]
2011-08-16
[88]
논문
Giant cladoxylopsid trees resolve the enigma of the Earth's earliest forest stumps at Gilboa
[89]
논문
Fossil Soils as Grounds for Interpreting the Advent of Large Plants and Animals on Land [and Discussion]
[90]
논문
''Rellimia thomsonii'' from the Givetian of New York: Secondary Growth in Three Orders of Branching
[91]
서적
Fossil Plants
Smithsonian Books, Washington D.C.
[92]
논문
Key innovations, convergence, and success: macroevolutionary lessons from plant phylogeny
http://paleobiol.geo[...]
[93]
논문
Phylogeny of seed plants based on all three genomic compartments: Extant gymnosperms are monophyletic and Gnetales' closest relatives are conifers
[94]
논문
Seed plant phylogeny inferred from all three plant genomes: Monophyly of extant gymnosperms and origin of Gnetales from conifers
[95]
논문
Phylogeny of seed plants based on evidence from eight genes
[96]
논문
Cretaceous angiosperm flowers: Innovation and evolution in plant reproduction
[97]
논문
Pteridosperms are the backbone of seed-plant phylogeny
[98]
논문
Morphological and molecular phylogenetic context of the angiosperms: contrasting the 'top-down' and 'bottom-up' approaches used to infer the likely characteristics of the first flowers
[99]
논문
Angiosperm diversification through time
https://bsapubs.onli[...]
2009-01-01
[100]
논문
Terrestrial-marine teleconnections in the Devonian: links between the evolution of land plants, weathering processes, and marine anoxic events
[101]
논문
Runcaria, A Middle Devonian Seed Plant Precursor
https://www.science.[...]
American Association for the Advancement of Science
[102]
논문
A Late Devonian Fertile Organ with Seed Plant Affinities from China
[103]
논문
Evolution of seed dormancy
[104]
논문
Flower, fruit and seed abortion in tropical forest trees. Implications for the evolution of paternal and maternal reproductive patterns
[105]
논문
DNA lesions occur with loss of viability in embryos of ageing rye seed
1978-04
[106]
논문
The alkaline single-cell gel electrophoresis/comet assay: a way to study DNA repair in radicle cells of germinating Vicia faba
[107]
논문
DNA repair mechanisms in plants: crucial sensors and effectors for the maintenance of genome integrity
2005-12
[108]
서적
Aging, Sex, and DNA Repair
Academic Press, San Diego
[109]
논문
Fossil evidence for Cretaceous escalation in angiosperm leaf vein evolution
[110]
논문
Molecular evolution of flower development
[111]
뉴스
Flowers originated 50 million years earlier than previously thought
https://www.eurekale[...]
2018-12-18
[112]
논문
How deep is the conflict between molecular and fossil evidence on the age of angiosperms?
2019-01-25
[113]
논문
Antiquity and Evolution of the MADS-Box Gene Family Controlling Flower Development in Plants
[114]
논문
Archaefructaceae, a New Basal Angiosperm Family
[115]
논문
Archaefructus—angiosperm precursor or specialized early angiosperm?
[116]
논문
After a dozen years of progress the origin of angiosperms is still a great mystery
[117]
논문
Ecological Aspects Of The Cretaceous Flowering Plant Radiation
[118]
논문
Dark and disturbed: a new image of early angiosperm ecology
http://paleobiol.geo[...]
[119]
뉴스
Did the first flower look like this?
https://www.bbc.com/[...]
2017-08-01
[120]
논문
The ancestral flower of angiosperms and its early diversification
2017-08-01
[121]
논문
The ''Amborella'' Genome and the Evolution of Flowering Plants
https://www.science.[...]
2013-12-20
[122]
논문
A physical map for the ''Amborella trichopoda'' genome sheds light on the evolution of angiosperm genome structure
[123]
논문
Genetic damage, mutation, and the evolution of sex.
1985-09-20
[124]
논문
Apomixis and the paradox of sex in plants.
2024-03-18
[125]
논문
Function and evolution of the plant MADS-box gene family
2001-03
[126]
논문
MADS-Box Genes in Ginkgo biloba and the Evolution of the AGAMOUS Family
[127]
논문
Translational Biology: From Arabidopsis Flowers to Grass Inflorescence Architecture
http://www.plantphys[...]
[128]
논문
Rose MADS-box genes 'MASAKO C1 and D1' homologous to class C floral identity genes
[129]
논문
Multiple AGAMOUS Homologs from Cucumber and Petunia Differ in Their Ability to Induce Reproductive Organ Fate
[130]
논문
The floral genome: an evolutionary history of gene duplication and shifting patterns of gene expression
[131]
논문
It's time to flower: the genetic control of flowering time
http://www3.intersci[...]
[132]
논문
Flowering on time: genes that regulate the floral transition
[133]
논문
The Mostly Male Theory of Flower Evolutionary Origins: from Genes to Fossils
[134]
논문
Evolution of plant microRNA gene families
[135]
논문
The genetics of maize evolution
https://zenodo.org/r[...]
[136]
논문
Variation and Selection at the CAULIFLOWER Floral Homeotic Gene Accompanying the Evolution of Domesticated Brassica olerace
[137]
논문
Phenotypic landscape inference reveals multiple evolutionary paths to C4 photosynthesis
eLife Sciences Publications
2013-09-28
[138]
논문
Anatomical enablers and the evolution of C4 photosynthesis in grasses
[139]
논문
Review. Nature's green revolution: the remarkable evolutionary rise of C4 plants
[140]
논문
A Fossil Grass (Gramineae: Chloridoideae) from the Miocene with Kranz Anatomy
[141]
논문
Carbon Isotopes in Photosynthesis
1988-05
[142]
논문
Neogene Expansion of the North American Prairie
1997-08-01
[143]
논문
Ecological selection pressures for C4 photosynthesis in the grasses
2009-02
[144]
논문
The global distribution of ecosystems in a world without fire
[145]
문서
[146]
논문
Marked Decline in Atmospheric Carbon Dioxide Concentrations During the Paleogene
[147]
논문
Dinosaurs Dined on Grass
[148]
논문
Dinosaur Coprolites and the Early Evolution of Grasses and Grazers
[149]
논문
Fire and the Miocene expansion of C4 grasslands
[150]
논문
Cenozoic Expansion of Grasslands and Climatic Cooling
[151]
논문
An Arabidopsis transcriptional regulatory map reveals distinct functional and evolutionary features of novel transcription factors
2015-07
[152]
논문
Genetics and biochemistry of secondary metabolites in plants: an evolutionary perspective
[153]
논문
The evolution of function in plant secondary metabolites
http://www.journals.[...]
[154]
논문
Plant terpenoid synthases: molecular and phylogenetic analysis
[155]
간행물
"Microbial interactions within the plant holobiont"
[156]
웹사이트
Transition of plants to land
http://www.clas.ufl.[...]
2011-03-06
[157]
간행물
Elkinsia gen. nov., a Late Devonian gymnosperm with cupulate ovules
http://www.jstor.org[...]
[158]
간행물
Roots: evolutionary origins and biogeochemical significance
http://jxb.oxfordjou[...]
[159]
서적
The origin and early diversification of land plants. A cladistic study.
Smithsonian Institution Press
[160]
논문
Molecular evidence for the early colonization of land by fungi and plants
http://www.sciencema[...]
2001-08
[161]
문서
[162]
논문
The late Precambrian greening of the Earth
[163]
간행물
The Microfossil Record of Early Land Plants: Advances in Understanding of Early Terrestrialization, 1970-1984
http://links.jstor.o[...]
2008-04-26
[164]
논문
The microfossil record of early land plants
[165]
논문
Early Middle Ordovician evidence for land plants in Argentina (eastern Gondwana)
[166]
논문
Fragments of the earliest land plants
[167]
논문
Massive release of hydrogen sulfide to the surface ocean and atmosphere during intervals of oceanic anoxia
[168]
논문
Oxygen, animals and oceanic ventilation: an alternative view
[169]
논문
Origin and Radiation of the Earliest Vascular Land Plants
[170]
논문
Wetlands before tracheophytes: Thalloid terrestrial communities of the Early Silurian Passage Creek biota (Virginia)
http://www.humboldt.[...]
2014-05-28
[171]
논문
The diversification of Paleozoic fire systems and fluctuations in atmospheric oxygen concentration
[172]
서적
Paleobotany and the evolution of plants
Cambridge University Press
[173]
간행물
How green was ''Cooksonia''? The importance of size in understanding the early evolution of physiology in the vascular plant lineage
[174]
논문
[175]
논문
[176]
논문
Evidence for lignin-like constituents in early silurian (llandoverian) plant fossils
http://www.sciencema[...]
1980-07
[177]
논문
The deepest divergences in land plants inferred from phylogenomic evidence
[178]
서적
Paleobiology and the evolution of plants
Cambridge University Press
[179]
논문
[180]
문서
Zosterophyllophytes
[181]
논문
The age of the earliest club mosses: the Silurian Baragwanathia flora in Victoria, Australia
http://geolmag.geosc[...]
2007-10-25
[182]
논문
The Science of Plant Morphology: Definition, History, and Role in Modern Biology
http://links.jstor.o[...]
2008-01-31
[183]
논문
Perithecial ascomycetes from the 400 million year old Rhynie chert: an example of ancestral polymorphism
http://www.mycologia[...]
2008-04-07
[184]
논문
Evolution of developmental potential and the multiple independent origins of leaves in Paleozoic vascular plants
[185]
논문
Structure of the Earliest Leaves: Adaptations to High Concentrations of Atmospheric {{co2}}
[186]
논문
Geocarb III: A Revised Model of Atmospheric {{co2}} over Phanerozoic Time
http://ajsonline.org[...]
2008-04-07
[187]
논문
Evolution of leaf-form in land plants linked to atmospheric CO2 decline in the Late Palaeozoic era
[188]
문서
The biology and evolution of fossil plants
[189]
논문
Reconstructing a lost Eocene paradise: Part I. Simulating the change in global floral distribution at the initial Eocene thermal maximum
http://linkinghub.el[...]
2008-04-08
[190]
논문
The advantages of being evergreen
[191]
논문
Ninety-seven million years of angiosperm-insect association: paleobiological insights into the meaning of coevolution
[192]
논문
Devonian landscape heterogeneity recorded by a giant fungus
http://geology.geosc[...]
2007-05
[193]
논문
Giant cladoxylopsid trees resolve the enigma of the Earth's earliest forest stumps at Gilboa.
[194]
논문
Fossil Soils as Grounds for Interpreting the Advent of Large Plants and Animals on Land [and Discussion]
http://links.jstor.o[...]
2008-04-07
[195]
논문
"Rellimia thomsonii" from the Givetian of New York: Secondary Growth in Three Orders of Branching
http://links.jstor.o[...]
2008-04-07
[196]
서적
Fossil Plants
Smithsonian Books, Washington D.C.
[197]
논문
Key innovations, convergence, and success: macroevolutionary lessons from plant phylogeny
http://paleobiol.geo[...]
2008-04-07
[198]
논문
Phylogeny of seed plants based on all three genomic compartments: Extant gymnosperms are monophyletic and Gnetales' closest relatives are conifers
[199]
논문
Seed plant phylogeny inferred from all three plant genomes: Monophyly of extant gymnosperms and origin of Gnetales from conifers
[200]
논문
Phylogeny of seed plants based on evidence from eight genes
http://amjbot.org/cg[...]
2008-04-08
[201]
논문
Cretaceous angiosperm flowers: Innovation and evolution in plant reproduction
[202]
논문
Pteridosperms are the backbone of seed-plant phylogeny
[203]
논문
Morphological and molecular phylogenetic context of the angiosperms: contrasting the 'top-down' and 'bottom-up' approaches used to infer the likely characteristics of the first flowers
[204]
논문
Middle to Late Paleozoic Atmospheric {{co2}} Levels from Soil Carbonate and Organic Matter
[205]
논문
GEOCARB II: A revised model of atmospheric {{co2}} over Phanerozoic time
[206]
논문
Late Devonian Oceanic Anoxic Events and Biotic Crises: "Rooted" in the Evolution of Vascular Land Plants?
https://rock.geosoci[...]
[207]
서적
Paleosols: their Recognition and Interpretation
Blackwell
[208]
논문
The origin and early evolution of plants on land
[209]
논문
A new fungal phlyum, the Glomeromycota'': phylogeny and evolution.
http://journals.camb[...]
[210]
논문
Origin and diversification of endomycorrhizal fungi and coincidence with vascular land plants
[211]
논문
Four hundred-million-year-old vesicular arbuscular mycorrhizae.
[212]
논문
Coevolution of roots and mycorrhizas of land plants
[213]
논문
Terrestrial-marine teleconnections in the Devonian: links between the evolution of land plants, weathering processes, and marine anoxic events
[214]
논문
Evolution of seed dormancy
[215]
논문
Antiquity and Evolution of the MADS-Box Gene Family Controlling Flower Development in Plants
http://mbe.oxfordjou[...]
[216]
논문
Archaefructaceae, a New Basal Angiosperm Family
[217]
논문
Archaefructus--angiosperm precursor or specialized early angiosperm?
[218]
논문
After a dozen years of progress the origin of angiosperms is still a great mystery.
[219]
논문
Ecological Aspects Of The Cretaceous Flowering Plant Radiation
[220]
서적
Paleobotany and the evolution of plants
Cambridge Univ. Press
[221]
논문
Dark and disturbed: a new image of early angiosperm ecology
http://paleobiol.geo[...]
2008-04-08
[222]
논문
Review. Nature's green revolution: the remarkable evolutionary rise of C4 plants
http://www.journals.[...]
2008-02-11
[223]
논문
A Fossil Grass (Gramineae: Chloridoideae) from the Miocene with Kranz Anatomy
[224]
논문
[225]
문서
Functions of phytoliths in vascular plants: an evolutionary perspective
[226]
논문
Mass extinction events and the plant fossil record
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com