돌연변이
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
돌연변이는 유전 정보의 무작위적인 변화로, 작용 방식과 규모에 따라 여러 종류로 나뉜다. 1901년 휘호 더 프리스에 의해 처음 발견되었으며, 허먼 J. 뮬러의 실험을 통해 인위적인 유발이 가능하다는 것이 밝혀졌다. 돌연변이는 유전자의 기능, 표현형 변화, 적응도, 유전 여부 등에 따라 분류되며, DNA 복제 오류, 외부 요인(화학 물질, 방사선 등)에 의해 발생한다. 돌연변이는 질병, 항생제 내성, HIV 저항성 등 생물학적, 사회경제적 영향과 밀접하게 관련되어 있으며, 보상 돌연변이 연구를 통해 유전자 기능과 단백질 진화에 대한 이해를 높이고 있다.
더 읽어볼만한 페이지
- 분자 진화 - 중립 진화 이론
중립 진화 이론은 자연 선택 외에 중립적인 돌연변이와 유전적 부동이 생물 진화의 주요 원동력이라고 주장하며, 유전자 다형성, 분자 시계 등을 설명하고 자연 선택과 병립 가능하다. - 분자 진화 - 유전자 중복
유전자 중복은 DNA 서열 일부가 한 개체 내에서 두 번 이상 나타나는 현상으로, 진화의 원동력으로서 새로운 유전적 기능 획득, 종 분화, 질병 발생에 영향을 미치며, 복제 오류 등 다양한 기제를 통해 발생하고 신기능 획득, 부분기능화, 유전자 소실로 이어질 수 있으며, 유전체 연구 및 유전 공학에서 활용된다. - 진화 - 살아있는 화석
살아있는 화석은 화석 기록에서 오랜 기간 동안 형태적 변화가 거의 없이 나타나는 생물을 의미하며, 진화와 종분화를 이해하는 데 중요한 역할을 한다. - 진화 - 진화심리학
진화심리학은 다윈의 진화론에 기반하여 인간의 심리적 적응이 자연선택과 성선택을 통해 진화해왔다고 보고, 인류의 진화적 적응 환경에서 생존과 번식에 관련된 문제 해결에 특화된 심리적 메커니즘을 강조하며, 다양한 심리학 분야에 적용되어 인간 행동을 설명하지만 유전자 결정론, 검증 어려움, 윤리적 문제 등 비판도 존재하는 학문이다. - 돌연변이 - 조로증
조로증은 DNA 복구 과정 결함으로 아동기 또는 청소년기에 노화 징후를 보이는 희귀 유전 질환으로, RecQ 헬리카제, NER 단백질, 라민 등 DNA 복구 단백질 유전자 돌연변이가 주요 원인이며, 베르너 증후군, 블룸 증후군, 로스문드-톰슨 증후군, 콕케인 증후군, 색소피부건조증, 허친슨-길포드 프로제리아 증후군 등이 대표적이다. - 돌연변이 - 돌연변이원
돌연변이원은 DNA에 변화를 일으켜 유전독성을 유발하는 물질 또는 방사선으로, 에임스 검사로 검출이 가능하며 암을 비롯한 질병을 유발할 수 있고, 물리적, 화학적, 생물학적 요인 등 다양한 종류가 존재하여 위험성을 줄이기 위한 연구가 진행 중이다.
| 돌연변이 | |
|---|---|
| 돌연변이 | |
 | |
| |
| 정의 | |
| 설명 | 게놈의 뉴클레오타이드 서열의 변화 |
| 문화어 | 갑작변이 |
| 진화 과정 | |
| 요인 | 유전적 다양성 (유전정보) 유전적 부동 유전자 이동 돌연변이 자연선택 |
| 결과 | 적응 공진화 공생 종분화 절멸 |
| 연구 및 역사 | |
| 주요 분야 | 진화생물학 진화유전학 분류학 생태유전학 진화발생생물학 인류의 진화 분자 진화 계통학 집단유전학 고생물학 |
| 주요 학자 | |
| 학자 | 이래즈머스 다윈 라마르크 라이엘 찰스 다윈 월리스 헉슬리 멘델 바이스만 더프리스 피셔 기무라 리키 굴드 도킨스 로즈 콜린스 렌스키 |
| 관련 링크 | |
| 영어 | mutation |
2. 역사
돌연변이의 원인은 DNA 복제 과정에서 우연히 발생하는 자연적인 원인과 방사선이나 화학물질 등의 영향으로 인한 외부 요인이 있다. 자연 발생적 변이는 100만 번의 DNA 복제 중에서 한 번 정도의 비율로 일어나며, 방사선이나 약품을 처리하면 이보다 높은 빈도로 일어난다. 노랑초파리를 이용한 인위적인 돌연변이 실험 결과 돌연변이의 약 70%는 개체에게 해로운 방향으로 진행되며, 나머지는 중립적이거나 유리한 성향을 보인다.[160] 돌연변이가 개체에게 해로운 작용을 할 가능성이 높기 때문에 생물은 세포 수준에서 돌연변이를 무효화하는 DNA 수복 기제를 작동시키도록 진화하여 왔다.[161]
따라서, 생물 종에서 일어나는 실제 돌연변이 비율은 높은 돌연변이 비율을 가질 경우 발생할 수 있는 해로운 돌연변이로 인한 비용과 DNA 수리 효소 등의 생성에 필요한 물질대사를 이루기 위한 에너지 소모 비용 간의 균형을 통해 결정되게 된다.[162]
돌연변이 비율이 높은 것이 오히려 유리할 수도 있다. 바이러스의 경우 RNA를 유전 물질로 사용하며 돌연변이 비율이 매우 높은데,[163] 이는 인간 등의 면역 체계를 피해야 하는 바이러스에게 유리하게 작용한다.[164]
흔히 발견할 수 있는 돌연변이 가운데 하나는 유전자 재조합 과정에서 일어나는 유전자 중복이다.[165] 유전자 중복의 결과 나타난 새로운 유전자는 진화의 원인이 된다. 생물 종에서 유전자 중복 발현 정도는 매 백만 년 당 수십 회에서 수백 회 정도로 알려져 있다.[166] 생물에서 발견되는 상동성 역시 이러한 유전자 중복과 같은 돌연변이의 결과로 이해할 수 있다.[167] 즉, 유전자 재조합 등을 통해 다음 세대로 전달된 돌연변이는 기존의 유전자와는 다른 새로운 기능으로 발현되게 되는 것이다.[168][169]
유전자 수준뿐만 아니라 염색체 수준에서 발생하는 대규모 돌연변이도 있다. 예를 들면 염색체의 분리나 접합에 의한 염색체 수 변화가 그것이다. 사람속 염색체에서도 이러한 사실을 확인할 수 있다. 인간 염색체 2번은 원래 두 개의 염색체였으나 돌연변이에 의하여 하나의 염색체로 접합된 것이다. 때문에 다른 영장류가 24쌍의 염색체를 갖는 것에 비해 인간은 23쌍의 염색체를 갖게 되었다.[170]
잡종 발현과 같은 이유로 인해 발생할 수 있는 염색체 재배열은 집단 간 유전자 변이를 일으켜 결국 종분화가 이루어지는 원인이 되며, 그 결과 진화 진행을 가속화시킬 수 있다.[171]
게놈 안에서 위치를 바꿀 수 있는 유전자 군인 전이성 유전인자 역시 생물의 유전자 변화에 중요한 역할을 담당하고 있으며 게놈 진화에 기여한다.[172] 인간의 경우 전체 인간 게놈의 45%가 5종류의 전이성 유전인자로 이루어져 있다.
2. 1. 돌연변이 개념의 등장
돌연변이를 발견하고 이름을 붙인 것은 네덜란드의 생물학자 휘호 더 프리스였으며, 1901년의 일이었다. 휘호 더 프리스는 진화가 돌연변이에 의해 일어난다는 돌연변이설을 주장했다.돌연변이를 인위적으로 유발할 수 있다는 것을 실험적으로 증명한 것은 허먼 J. 뮬러이다. 뮬러는 초파리에 X선을 조사하여, 다음 세대의 치사율을 측정함으로써, 이론치를 통해 이를 추측했다. 이후, 생물학(유전학)에서는 인위적으로 돌연변이를 유도하는 변이 도입에 의해 돌연변이체를 얻고, 그 표현형을 관찰함으로써, 유전자의 기능을 해석해 왔다.
2. 2. 돌연변이 연구의 발전
돌연변이를 발견하고 이름을 붙인 사람은 네덜란드의 생물학자 휘호 더 프리스였으며, 1901년의 일이었다. 휘호 더 프리스는 여기서 진화가 돌연변이에 의해 일어난다는 돌연변이설을 주장했다.돌연변이를 인위적으로 일으킬 수 있다는 것을 실험적으로 증명한 사람은 허먼 J. 뮬러이다.(인위 돌연변이 항목 참조) 허먼 J. 뮬러는 초파리에 X선을 쬐어, 다음 세대의 치사율을 측정함으로써, 이론치에서 이를 추측했다. 이후, 생물학(유전학)에서는 인위적으로 돌연변이를 유도하는 변이 도입을 통해 돌연변이체를 얻고, 그 표현형을 관찰함으로써, 유전자의 기능을 해석해 왔다.
3. 종류
돌연변이는 자연 발생적이거나 외부 요인에 의해 발생한다. 자연 발생적 변이는 DNA 복제 과정에서 100만 번 중 한 번 정도의 비율로 나타나며, 방사선이나 화학물질은 돌연변이 발생 빈도를 높인다. 노랑초파리 실험에 따르면 돌연변이의 약 70%는 개체에 해로우며, 나머지는 중립적이거나 유리하다.[160]
생물은 DNA 수복 기제를 통해 돌연변이의 해로운 영향에 대응한다.[161] 그러나 바이러스처럼 돌연변이 비율이 높은 것이 면역 체계 회피에 유리한 경우도 있다.[163][164]
유전자 중복은 유전자 재조합 과정에서 흔히 발생하며, 진화와 상동성의 원인이 된다.[165][167] 염색체 수준의 대규모 돌연변이도 있는데, 사람속의 인간 염색체 2번은 두 염색체가 융합된 것이다.[170] 이러한 염색체 재배열은 종분화와 진화를 촉진한다.[171]
전이성 유전인자는 게놈 내에서 위치를 이동하며 유전자 변화와 게놈 진화에 기여한다.[172] 인간 게놈의 45%는 전이성 유전인자로 구성되며, LINEs에 포함된 ALU는 치명적인 돌연변이를 유발할 수 있다.[173]
돌연변이는 유전자 다양성을 증가시키고 자연 선택의 영향을 받는다. 유전자 수준의 돌연변이에는 점 돌연변이, 미스센스 돌연변이, 넌센스 돌연변이 등이 있다. 염색체 수준의 돌연변이에는 염색체 이상이 있으며, 염색체 구조나 수의 변화를 포함한다.
3. 1. 구조적 변화에 따른 분류
돌연변이는 유전자의 서열 변화로 인해 발생하며, 건강에 미치는 영향은 돌연변이의 위치와 단백질 기능 변화 여부에 따라 다양하다.[46] 구조적 변화에 따른 돌연변이는 크게 소규모 돌연변이와 대규모 돌연변이로 나뉜다.

3. 1. 1. 소규모 돌연변이
뉴클레오타이드 수준에서 일어나는 소규모 돌연변이는 다음과 같다.[174]- 점 돌연변이: 하나의 뉴클레오타이드가 변환되어 나타나는 돌연변이로 DNA 전사 단계에서 특정 단백질의 생성을 막거나 변형시킨다.[175] 대부분 같은 푸린 계열인 아데닌과 구아닌, 또는 같은 피리미딘 계열인 시토신과 티민의 잘못된 전사로 인해 일어난다. 점 돌연변이는 코돈의 정보를 파괴하거나 변형시켜 다음과 같은 돌연변이 효과를 나타낸다.
- * 불현성 돌연변이: 점 돌연변이로 인해 변이된 코돈이 기존의 코돈과 같은 아미노산 형성을 지시할 경우 결과적으로 돌연변이의 효과는 나타나지 않는다.
- * 미스센스 돌연변이: 변이된 코돈이 다른 아미노산 형성을 지시하는 경우.[153] 코돈 내 염기의 변화 또는 치환에 의해, 원래 들어가야 할 것과 다른 아미노산이 합성된 폴리펩티드 중에 들어가 비정상 단백질이 만들어진다.
- * 넌센스 돌연변이: 변이된 코돈으로 인해 아미노산 형성이 중단되거나 생략되는 경우.[154] 아미노산의 코돈을 종결 코돈으로 만드는 돌연변이이다.
- 삽입 돌연변이: 뉴클레오타이드의 일부가 원래의 염기 서열 사이로 삽입되어 일어나는 돌연 변이.
- 결실 돌연변이: 원래 있던 뉴클레오타이드의 일부가 사라져 버린 돌연변이.
- 프레임시프트 돌연변이: 염기의 삽입, 결손에 의해 오픈 리딩 프레임이 어긋나는 돌연변이.[155]
점 돌연변이 가운데 아미노산이 다른 아미노산으로 변하는 비동의적 돌연변이, 아미노산의 코돈이 종결 코돈으로 변하는 넌센스 돌연변이, 종결 코돈이 아미노산의 코돈으로 변하는 읽기 오류 돌연변이 등은 중립 진화설/분자 시계 기능에 영향을 미친다.
세 개의 뉴클레오티드가 하나의 아미노산을 코딩하기 때문에, 삽입 또는 결실된 뉴클레오티드가 3의 배수이면 아미노산의 삽입 또는 결실이 발생하고, 그렇지 않으면 코돈의 읽기틀이 어긋나 아미노산 배열이 크게 변하는 프레임시프트가 발생한다.
3. 1. 2. 대규모 돌연변이
염색체 수준에서 일어나는 대규모 돌연변이에는 다음과 같은 것들이 있다.[174]- 유전자 중복: 염색체의 파손된 조각이 상동 또는 비상동 염색체에 부착되어 일부 유전자가 두 개 이상의 복제량으로 존재하게 되면서 모든 염색체 영역의 복제본이 증가하고, 그 안에 위치한 유전자의 복제량도 증가한다.
- 유전자 결실: 큰 염색체 영역의 결손으로, 해당 영역 내 유전자의 손실을 초래한다.
- 염색체 역위: 염색체 부분의 방향을 반전시키는 것이다.
- 간질성 결실: 단일 염색체에서 DNA 부분을 제거하여 이전에 멀리 떨어져 있던 유전자를 서로 인접하게 만드는 염색체 내 결손이다. 예를 들어, 인간 성상세포종(일종의 뇌종양)에서 분리된 세포는 교아종에서 융합된 (FIG) 유전자와 수용체 티로신 키나아제(ROS) 사이의 서열을 제거하는 염색체 결손을 가지고 있는 것으로 밝혀졌으며, 이로 인해 융합 단백질(FIG-ROS)이 생성된다. 비정상적인 FIG-ROS 융합 단백질은 구성적으로 활성화된 키나아제 활성을 가지고 있어 종양 형성(정상 세포에서 암세포로의 변환)을 유발한다.
- 염색체 전위: 비상동 염색체에서 유전적 부분 교환이 일어난다.
- 이형접합 소실: 이전에 두 개의 다른 대립 유전자를 가지고 있던 유기체에서 결손이나 유전자 재조합 사건에 의해 하나의 대립 유전자가 손실되는 것이다.
- 다배체: 전체 염색체 세트의 중복으로, 잠재적으로 별도의 번식 집단과 종 분화를 초래할 수 있다.
염색체 이상은 염색체의 구조 이상이나, 그것에 따른 장애가 발생하는 변이이다. 염색체 이상에 의한 돌연변이에는 염색체 구조의 변화나 염색체 수의 변화 등이 있다.
- 결손: 염색체의 일부가 소실된다.
- 예: 흰 까마귀, 주황색 두더지, 검은색에서 흰색으로 변한 개(래브라도 레트리버종)
- 역위: 염색체의 일부가 정상적인 방향과 반대로 된다.
- 중복: 염색체의 일부가 중복된다.
- 전좌: 염색체의 일부가 끊어져 다른 염색체에 연결된다.
- 배수성: 염색체 수가 2배, 3배, 4배와 같이 정수배가 되는 것이다.
- 씨 없는 수박: 일반적인 수박이 게놈의 2배의 염색체를 가지는 데 반해, 3배의 염색체를 가진다.
- 이수성: 염색체 수가 1개 또는 수 개 증감하는 것이다.
- 다운증후군: 21번 염색체를 1개 더 가지는 것이다.
3. 2. 기능적 변화에 따른 분류
돌연변이는 유전자의 기능 변화에 따라 여러 유형으로 나눌 수 있다. 일반적으로 기능은 적응 정도에 따라 달라지며, 특정 기능이 사라지면서 다른 기능으로 이용되거나, 기능 증가로 인해 기존 기능을 사용하지 못하게 되는 경우도 있어, 여기서 설명하는 변이들은 서로 반대로 해석될 수도 있다.[46]- 기능 상실 돌연변이 (Loss-of-function mutation): 유전자의 기능이 없어지거나 감소하는 돌연변이이다.
- 예시: 유전자 접합에 의한 일부 변이들
- 기능 획득 돌연변이 (Gain-of-function mutation): 유전자에 새로운 기능이 추가되거나 기능이 강화되는 돌연변이이다.
- 예시:
- 호메오 박스 돌연변이(안테나피디아): 더듬이 부분에 새로운 다리가 생겨 다리 기능을 획득한다.
- 항생제 저항성 유전자 획득: 플라스미드를 통해 페니실린 분해 효소 생성 기능을 획득하여 세포벽을 보호한다.
- 대장균의 장기간 진화 실험: 시트르산 이용 기능 획득.
이 외에도 다음과 같은 돌연변이가 있다.
- 중립적 돌연변이: 자연선택에 유리하지도 불리하지도 않은 중립적인 돌연변이이다.(→중립진화설, 분자시계 항목 참조)[147]
- 비표현 돌연변이: 유전적 수준에서는 변이가 일어나지만 표현형에서는 알 수 없는 변이이다.[148]
- 억압 돌연변이: 억제 유전자 변이라고도 하며, tRNA의 안티코돈을 변화시켜 종결 코돈을 인식할 수 있게 되어 아미노산 사슬 합성이 종료되지 않는 변이이다.[150]
- 적응적 돌연변이: 무작위로 돌연변이가 일어나는 것이 아니라, 주변 환경에 적응하여 일어난다고 여겨졌던 돌연변이이나, 현재는 부정되고 있다.[151]
- 점 돌연변이: 하나의 뉴클레오티드의 치환, 결손, 또는 삽입 돌연변이이다.[152]
- 미스센스 돌연변이: 코돈 내 염기 변화 또는 치환으로 인해, 원래 들어가야 할 것과 다른 아미노산이 합성된 폴리펩티드 중에 들어가 비정상 단백질이 만들어지는 돌연변이이다.[153]
- 넌센스 돌연변이: 아미노산의 코돈을 종결 코돈으로 만드는 돌연변이이다.[154]
- 프레임시프트 돌연변이: 염기의 삽입, 결손에 의해 오픈 리딩 프레임이 어긋나는 돌연변이이다.[155]
3. 3. 표현형 변화에 따른 분류
일반적으로 변이는 유전자 다양성을 증가시키며, 각 세대마다 조합을 통해 증가하는 유전 정보 외에도 무작위적인 변화를 가능하게 한다. 이러한 돌연변이는 표현형에 다양한 영향을 미칠 수 있다.[47]- 기능 상실 돌연변이: 유전자의 기능이 사라지는 돌연변이이다.
- 기능 획득 돌연변이:
- 호메오 박스 돌연변이(안테나피디아): 더듬이 부분에 새로운 다리가 생겨 다리 기능을 획득한다.
- 항생제 저항성 유전자 획득: 플라스미드를 통해 페니실린 분해 효소 생성 기능을 획득하여 세포벽을 보호한다.
- 대장균의 장기간 진화 실험: 시트르산 이용 기능을 획득한 경우이다.
- 복귀 돌연변이: 원래 상태로 되돌아가는 돌연변이이다.
- 소규모 돌연변이:
- 삽입: DNA에 하나 이상의 뉴클레오티드가 추가된다. 전이인자 또는 반복 요소 복제 오류로 발생하며, 스플라이싱이나 읽기틀을 변화시켜(프레임시프트 돌연변이) 유전자 산물을 변경할 수 있다.
- 결손: DNA에서 하나 이상의 뉴클레오티드가 제거된다. 삽입과 마찬가지로 읽기틀을 변경할 수 있다.
- 치환 돌연변이: 한 뉴클레오티드가 다른 뉴클레오티드로 교환된다.[48] 퓨린 간(A ↔ G), 피리미딘 간(C ↔ T) 교환은 전이, 퓨린-피리미딘 교환(C/T ↔ A/G)은 트랜스버전이다.
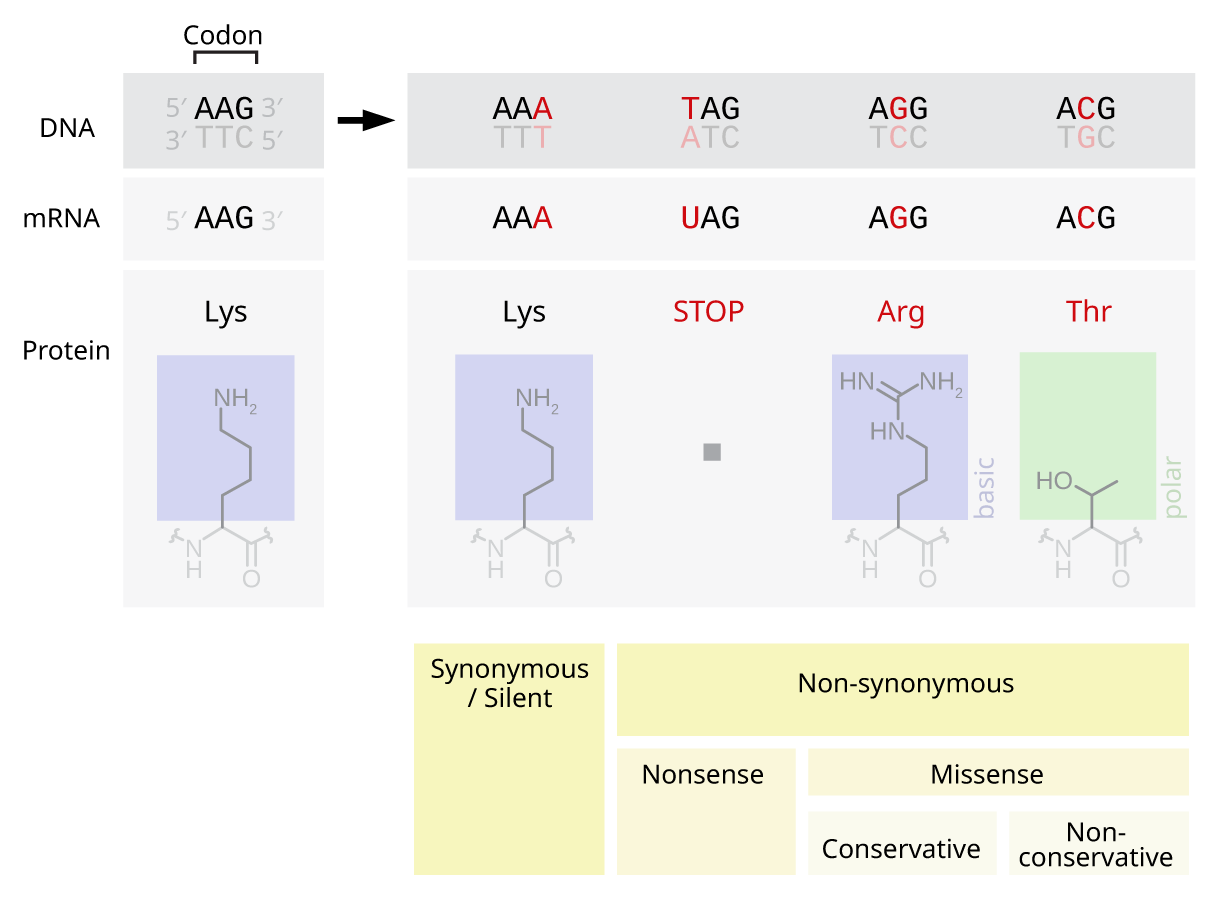
- 단백질 서열에 미치는 영향:
- 프레임시프트 돌연변이: 3의 배수가 아닌 뉴클레오티드 삽입/삭제로 읽기 프레임이 바뀌어 번역 결과가 달라진다.[50]
- 점 치환 돌연변이:
- 동의어 치환: 코돈이 동일한 아미노산을 코딩하는 다른 코돈으로 바뀌지만, 아미노산 서열은 변하지 않는다. 침묵 돌연변이라고도 하지만, 모든 동의어 치환이 침묵 돌연변이인 것은 아니다.
- 비동의어 치환: 코돈이 다른 아미노산을 코딩하는 코돈으로 바뀐다.
- 미스센스 돌연변이: 다른 아미노산 치환으로 단백질 기능 이상을 유발할 수 있다. 수포성 표피 박리증, 겸상 적혈구 빈혈, 근위축성 측삭 경화증 등이 예시이다.[51]
- 넌센스 돌연변이: 조기 종결 코돈을 생성하여 불완전한 단백질이 만들어진다. 선천성 부신 과형성증 등이 예시이다.
- 중립적 돌연변이:[147] 자연선택에 유리하지도 불리하지도 않은 중립적인 돌연변이이다. (→중립진화설, 분자시계 참조)
- 비표현 돌연변이:[148] 유전형에는 변이가 있지만 표현형에는 나타나지 않는 변이이다.
- 억압 돌연변이:[150] 억제 유전자 변이라고도 하며, tRNA의 안티코돈을 변화시켜 종결 코돈을 인식하고 아미노산 사슬 합성이 종료되지 않는 변이이다.
- 적응적 돌연변이:[151] 환경에 적응하여 일어난다고 여겨졌던 돌연변이이나, 현재는 부정되고 있다.
- 미스센스 돌연변이:[153] 코돈 내 염기 변화로 다른 아미노산이 합성되어 비정상 단백질이 만들어지는 돌연변이이다.
- 넌센스 돌연변이:[154] 아미노산 코돈을 종결 코돈으로 만드는 돌연변이이다.
- 프레임시프트 돌연변이:[155] 염기 삽입, 결손으로 오픈 리딩 프레임이 어긋나는 돌연변이이다.
3. 4. 적응도에 따른 분류
돌연변이는 생물의 적응도와 관련하여 해로운 돌연변이, 이로운 돌연변이, 중립적 돌연변이로 분류할 수 있다. 이 분류는 환경에 따른 진화 방향을 결정하며, 환경이 달라지면 기준도 바뀐다.- 해로운 돌연변이: 대부분의 환경에서 생존에 불리하게 작용하는 돌연변이. 예시: 겸형 적혈구 빈혈증
- 이로운 돌연변이: 특정 환경에서 생존에 유리하게 작용하는 돌연변이. 예시: 겸형 적혈구 빈혈증(말라리아 창궐 지역), 안테나 페디아(더듬이 부분에 농약이 있는 곳)
- 중립적 돌연변이: 자연선택(자연도태)에 유리하지도 불리하지도 않은 돌연변이. (중립진화설, 분자시계 항목 참조)[147]
겸형 적혈구 빈혈증은 대부분의 환경에서는 해롭지만, 말라리아가 창궐하는 지역에서는 이형 접합자 이점에 의해 이로운 변이로 작용한다. 안테나 페디아 역시 대부분의 환경에서는 불리하지만, 더듬이 부분에 작용하는 농약이 있는 곳에서는 유리하게 작용할 수 있다. 이처럼 많은 변이가 해롭게 작용하지만, 이로운 변이도 다수 존재하며, 이는 환경에 따라 달라지므로 해로움, 이로움, 중립을 절대적으로 정의할 수는 없다.[47]
일부 의사과학을 주장하는 집단(창조과학회 등)은 이로운 변이가 존재하지 않는다는 반지성주의적 주장을 한다. 그러나 이는 사실과 다르며, 이로운 변이는 다수 존재하고 환경에 따라 결정된다. 그들이 제시하는 "해로운 변이" 역시 특정 환경에서는 이로운 변이로 작용할 수 있다.
3. 5. 유전 여부에 따른 분류
변이는 유전 여부에 따라 다음과 같이 분류할 수 있다.- 중립적 돌연변이: 자연선택(자연도태)에 유리하지도 불리하지도 않은 중립적인 돌연변이이다. (중립진화설, 분자시계 항목 참조)[147]
- 비표현 돌연변이: 유전적 수준에서는 변이가 일어나지만, 표현형에서는 알 수 없는 변이이다.[148]
- 복귀 돌연변이: 돌연변이 유전자가 다시 변이를 일으켜 원래 유전자로 돌아가는 변이이다.[149]
- 억압 돌연변이: 억제 유전자 변이라고도 한다. tRNA의 안티코돈을 변화시켜 종결 코돈을 인식할 수 있게 되어 아미노산 사슬 합성이 종료되지 않는 변이이다.[150]
- 적응적 돌연변이: 무작위로 돌연변이가 일어나는 것이 아니라, 주변 환경에 적응하여 일어난다고 여겨졌던 돌연변이이다. 현재는 부정되고 있다.[151]
4. 발생 원인
돌연변이의 발생 원인은 크게 자연적인 요인과 외부적인 요인으로 나눌 수 있다. 자연적인 요인으로는 유전 물질 복제 과정에서 무작위로 발생하는 오류가 있으며, 외부적인 요인으로는 방사선이나 화학 물질 등이 있다.
자연 발생적인 변이는 DNA 복제 과정에서 드물게 발생하며, 100만 번의 복제 중 한 번 정도의 비율로 나타난다. 노랑초파리를 이용한 실험에 따르면, 이러한 돌연변이의 약 70%는 개체에게 해로운 영향을 미치지만, 나머지는 중립적이거나 유리하게 작용하기도 한다.[160] 생물은 돌연변이의 부정적인 영향을 줄이기 위해 DNA 수복과 같은 방어 기제를 진화시켜 왔다.[161] 바이러스와 같이 RNA를 유전 물질로 사용하는 경우에는 돌연변이 비율이 매우 높은데,[163] 이는 면역 체계를 회피하는 데 유리하게 작용하기 때문이다.[164]
유전자 중복은 유전자 재조합 과정에서 흔히 발생하는 돌연변이 중 하나로,[165] 새로운 유전자를 생성하여 진화의 원동력이 된다.[166] 상동성 역시 유전자 중복과 같은 돌연변이의 결과로 이해할 수 있다.[167] 염색체 수준에서도 대규모 돌연변이가 발생할 수 있는데, 예를 들어 사람속의 인간 염색체 2번은 원래 두 개의 염색체가 하나로 융합된 것이다.[170] 이러한 염색체 재배열은 종분화를 유발하여 진화를 가속화할 수 있다.[171]
게놈 내에서 위치를 이동할 수 있는 전이성 유전인자 또한 유전자 변화와 게놈 진화에 중요한 역할을 한다.[172] 인간 게놈의 상당 부분은 전이성 유전인자로 구성되어 있으며, 이들은 때때로 치명적인 돌연변이를 유발하기도 한다.[173]
돌연변이는 유전자나 염색체의 손상으로 인해 발생하며, 점 돌연변이, 삽입, 결실 등 다양한 형태로 나타날 수 있다. 점 돌연변이 중에서도 아미노산 변화를 유발하지 않는 침묵 돌연변이는 유전될 가능성이 높다. 아미노산이 변하는 비동의적 돌연변이는 생존에 불리할 수 있지만, 유리한 경우도 있어 진화의 요인이 되기도 한다. 약물, 방사선, 고에너지 입자 등도 유전자를 손상시켜 돌연변이를 유발할 수 있으며,[156][157][158] 식물에서는 영양 장애도 돌연변이의 원인이 될 수 있다.[159]
4. 1. 자연적 발생
자발적 돌연변이는 건강하고 오염되지 않은 세포에서도 0이 아닌 확률로 발생한다. 자연적으로 발생하는 산화적 DNA 손상은 인간의 경우 세포당 하루 10,000번, 쥐의 경우 세포당 하루 100,000번 발생하는 것으로 추정된다.[33] 자발적 돌연변이는 특정 변화에 따라 특징지어질 수 있다.[34]- 토토머화(Tautomerism) – 염기의 수소 원자의 위치가 바뀌어 해당 염기의 수소 결합 패턴이 변하고, 결과적으로 복제 중 잘못된 염기쌍 형성이 발생한다.[35] 이론적 결과는 양성자 터널링(proton tunnelling)이 GC 토토머(tautomer)의 자발적 생성에 중요한 요인임을 시사한다.[36]
- 탈퓨린화(Depurination) – 퓨린 염기(A 또는 G)의 손실로 아퓨린성 부위(AP 부위)가 형성된다.
- 탈아미노화(Deamination) – 가수분해에 의해 정상적인 염기가 원래의 아민기 대신 케톤기를 포함하는 비정형 염기로 바뀐다. 예를 들어 C → U 및 A → HX(히포크산틴)이 있는데, 이는 DNA 복구 메커니즘에 의해 수정될 수 있다. 그리고 5MeC(5-메틸시토신) → T는 티민이 정상적인 DNA 염기이기 때문에 돌연변이로 감지될 가능성이 적다.
- 미스페어링 슬립(Slipped strand mispairing) – 복제 중 주형으로부터 새로운 가닥의 변성이 일어나고, 다른 지점에서 재결합("미끄러짐")이 발생한다. 이는 삽입 또는 결손으로 이어질 수 있다.
자발적으로 발생하는 돌연변이의 대부분은 주형 가닥의 DNA 손상을 지나치는 오류가 발생하기 쉬운 복제(translesion synthesis) 때문이라는 증거가 늘고 있다. 쥐의 경우, 대부분의 돌연변이는 복제후합성에 의해 발생한다.[37] 마찬가지로, 효모에서 Kunz 등[38]은 자발적인 단일 염기쌍 치환과 결실의 60% 이상이 복제후합성에 의해 발생한다는 것을 발견했다.
DNA에서 자연적으로 발생하는 이중 가닥 절단은 비교적 낮은 빈도로 발생하지만, 이를 복구하는 과정에서 종종 돌연변이가 발생한다. 비상동 말단 결합(Non-homologous end joining, NHEJ)은 이중 가닥 절단을 복구하는 주요 경로이다. NHEJ는 두 말단의 다소 부정확한 정렬을 위한 재결합을 허용하기 위해 몇몇 뉴클레오타이드를 제거하고, 그 후 틈을 메우기 위해 뉴클레오타이드를 추가하는 과정을 포함한다. 결과적으로 NHEJ는 종종 돌연변이를 유발한다.[39]
4. 2. 외부 요인에 의한 유발
유도 돌연변이는 유전자가 돌연변이원 및 환경적 요인과 접촉한 후 발생하는 유전자 변화이다.분자 수준에서 유도 돌연변이는 다음과 같은 원인으로 발생할 수 있다.
- 화학 물질:
- 하이드록실아민
- 염기 유사체 (예: 브로모데옥시우리딘)
- 알킬화제 (예: N-에틸-N-니트로소요소). 이들은 DNA 복제 여부와 관계없이 돌연변이를 일으킬 수 있다. 반면, 염기 유사체는 DNA 복제 시에만 돌연변이를 유발한다. 이러한 화학적 돌연변이원은 전이, 트랜스버전, 결실 등을 유발한다.
- DNA 부가체 형성 물질 (예: 옥살라토신 A)[41]
- DNA 삽입제 (예: 에티디움 브로마이드)
- DNA 가교 결합제
- 산화적 손상
- 아질산: A와 C의 아민기를 다이아조기로 바꿔 수소 결합 패턴을 변경, 복제 중 잘못된 염기쌍을 형성하게 한다.
- 방사선:
- 자외선 (UV) (비전리 방사선 포함): DNA의 시토신, 티민은 방사선에 취약하다. UV는 DNA 가닥에서 인접한 피리미딘 염기가 피리미딘 이합체로 공유 결합하게 한다. 장파장 UVA는 DNA의 산화적 손상을 유발할 수 있다.[42]
- 전리 방사선 (예: 감마선): 돌연변이를 유발하여 암이나 사망으로 이어질 수 있다.
약물 자극[156], 전리방사선, 이온빔[157] 등 고에너지 입자[158]도 유전자를 손상시켜 돌연변이를 유발한다. 식물에서는 영양 장애에 의해서도 유발되는 것으로 보고되었다.[159]
5. 영향
돌연변이는 생물체에 다양한 방식으로 영향을 미친다. 돌연변이의 영향은 크게 생물학적 영향, 사회경제적 영향, 긍정적 영향과 부정적 영향으로 나눌 수 있다.
돌연변이의 약 70%는 개체에게 해로운 방향으로 작용하며,[160] 생물은 세포 수준에서 돌연변이를 무효화하는 DNA 수복 기제를 작동시키도록 진화해 왔다.[161] 그러나, 바이러스처럼 돌연변이 비율이 높은 것이 오히려 유리한 경우도 존재한다.
유전자 돌연변이는 DNA 복제 과정에서의 실수, 화학물질, 방사선, 전이성 유전인자 등에 의해 발생한다. 점돌연변이의 경우, 아미노산 변화를 유발하지 않는 침묵 돌연변이가 더 많이 유전될 수 있다. 아미노산이 변화된 단백질은 기능을 잃는 경우가 많지만, 생존에 유리한 경우도 있어 진화의 요인이 되기도 한다.
염색체 이상은 염색체의 구조나 수에 변화를 일으키는 돌연변이이다. 체세포 돌연변이는 종양을 유발할 수 있으며, 생식세포 돌연변이는 유전되어 진화에 영향을 줄 수 있다. 세균이나 바이러스는 돌연변이를 통해 백신이나 치료제에 대한 내성을 획득하기도 한다.
5. 1. 생물학적 영향
돌연변이는 생물에게 여러 영향을 준다. 돌연변이의 원인으로는 DNA 복제 과정에서 무작위로 발생하는 자연적인 변이와 방사선, 화학 물질 등에 의한 외부 요인이 있다. 자연적인 변이는 100만 번의 DNA 복제 중 한 번 정도의 비율로 발생하며, 외부 요인에 노출되면 이보다 더 자주 발생한다. 노랑초파리를 이용한 실험에 따르면, 돌연변이의 약 70%는 개체에게 해로운 영향을 주며, 나머지는 중립적이거나 유리한 영향을 준다.[160]생물은 돌연변이의 해로운 영향을 줄이기 위해 DNA 수복 기제를 진화시켜 왔다.[161] 따라서 실제 돌연변이 비율은 해로운 돌연변이 발생 비용과 DNA 수리 효소 생성에 필요한 에너지 소모 비용 사이의 균형에 의해 결정된다.[162]
바이러스와 같이 돌연변이 비율이 높은 것이 유리한 경우도 있다. RNA를 유전 물질로 사용하는 바이러스는 돌연변이 비율이 매우 높은데,[163] 이는 면역 체계를 회피하는 데 유리하게 작용한다.[164]
유전자 재조합 과정에서 일어나는 유전자 중복은 흔히 발견되는 돌연변이 중 하나이다.[165] 유전자 중복으로 생긴 새로운 유전자는 진화의 원인이 된다. 생물 종에서 유전자 중복은 매 백만 년 당 수십 회에서 수백 회 정도 발생한다.[166] 상동성 역시 유전자 중복과 같은 돌연변이의 결과이다.[167] 즉, 유전자 재조합 등을 통해 다음 세대로 전달된 돌연변이는 기존 유전자와 다른 새로운 기능으로 발현된다.[168][169]
염색체 수준에서 발생하는 대규모 돌연변이도 있다. 예를 들어, 염색체 분리나 접합에 의한 염색체 수 변화가 있다. 사람속의 인간 염색체 2번은 원래 두 개의 염색체였으나 돌연변이로 인해 하나로 합쳐진 것이다. 이 때문에 다른 영장류는 24쌍의 염색체를 갖는 반면, 인간은 23쌍의 염색체를 갖는다.[170]
잡종 형성 등으로 인해 발생하는 염색체 재배열은 집단 간 유전자 변이를 일으켜 종분화를 유발하고, 진화를 가속화할 수 있다.[171]
게놈 내에서 위치를 이동할 수 있는 전이성 유전인자 또한 생물의 유전자 변화와 게놈 진화에 중요한 역할을 한다.[172] 인간의 경우, 인간 게놈의 45%가 5종류의 전이성 유전인자로 구성되어 있다. 이 중 20%를 차지하는 LINEs는 매우 안정적인 전이성 유전인자로, 크기가 약 6,000개의 염기쌍(6 Kb)에 달한다. LINEs에는 약 300개의 염기쌍으로 구성된 ALU라는 기생 유전자 코드가 수십만 번 반복되는데, ALU 자체는 이동 능력이 없지만 LINEs가 이동할 때 함께 이동한다. ALU가 LINEs의 효소를 이용하여 다른 염색체로 이동하면 치명적인 돌연변이를 유발할 수 있다. 인간 게놈에는 LINEs 외에도 RNA 유래 전이인자(8%), DNA 유래 전이인자(3%) 등이 있으며, 나머지 4%는 전이 능력을 상실한 비활성 전이인자이다.[173]
5. 2. 사회경제적 영향
Human Genome Variation Society영어 (HGVS, 인간 게놈 변이 학회)는 표준 인간 서열 변이 명명법[100]을 개발했으며, 연구원과 DNA 진단 센터에서 모호하지 않은 돌연변이 설명을 생성하는 데 사용한다. 명명법은 돌연변이의 유형과 염기 또는 아미노산 변화를 지정한다.- 염기 치환 (예: 76A>T): 숫자는 5' 말단에서 염기의 위치를 나타낸다. 첫 번째 문자는 야생형 염기를 나타내고, 두 번째 문자는 야생형을 대체한 염기를 나타낸다. 주어진 예에서 76번째 위치의 아데닌은 티민으로 대체되었다.
- 게놈 DNA, 미토콘드리아 DNA, RNA의 돌연변이를 구별해야 할 필요가 있는 경우 간단한 관례를 사용한다. 예를 들어, 염기 서열의 100번째 염기가 G에서 C로 돌연변이된 경우, 게놈 DNA에서 돌연변이가 발생하면 g.100G>C, 미토콘드리아 DNA에서 돌연변이가 발생하면 m.100G>C, RNA에서 돌연변이가 발생하면 r.100g>c로 쓴다. RNA의 돌연변이의 경우 염기 코드는 소문자로 작성한다.
- 아미노산 치환 (예: D111E): 첫 번째 문자는 야생형 아미노산의 한 문자 코드이며, 숫자는 N-말단에서 아미노산의 위치이고, 두 번째 문자는 돌연변이에 존재하는 아미노산의 한 문자 코드이다. 넌센스 돌연변이는 두 번째 아미노산에 X로 표시한다(예: D111X).
- 아미노산 결손 (예: ΔF508): 그리스 문자 Δ(델타)는 결손을 나타낸다. 문자는 야생형에 존재하는 아미노산을 나타내고, 숫자는 야생형과 같이 존재하는 경우 아미노산의 N 말단에서의 위치를 나타낸다.
세균이나 바이러스는 돌연변이에 의해 백신의 변이 또는 치료제에 대한 내성을 획득하는 경우가 있으며, 치료와 예방을 어렵게 한다.
5. 3. 긍정적 영향
돌연변이는 생물체에게 해로울 수 있지만, 특정 환경에서는 긍정적인 영향을 미칠 수도 있다. 이러한 경우 돌연변이는 돌연변이 생물체가 야생형 생물체보다 특정 환경 스트레스에 더 잘 견디거나 더 빨리 번식할 수 있게 한다.[118] 이러한 경우 돌연변이는 자연 선택을 통해 개체군에서 더 흔해지는 경향이 있다. 그러나 같은 돌연변이라도 한 조건에서는 유익하지만 다른 조건에서는 불리할 수 있다.[118]- '''HIV 저항성''': 사람의 CCR5-Δ32에서 특정 32개 염기쌍 결실은 동형접합자에게 HIV 저항성을 부여하고 이형접합자에서 AIDS 발병을 지연시킨다.[114] 유럽 인구에서 CCR5-Δ32의 상대적으로 높은 빈도의 원인에 대한 한 가지 설명은 14세기 중반 유럽에서 흑사병에 대한 저항성을 부여했기 때문이다.[115] 이 돌연변이를 가진 사람들은 감염에서 살아남을 가능성이 더 높았고, 따라서 개체군에서의 빈도가 증가했다.[115] 이 이론은 이 돌연변이가 흑사병의 영향을 받지 않은 남아프리카에서는 발견되지 않는 이유를 설명할 수 있다.[115] 최근의 이론은 CCR5 델타 32 돌연변이에 대한 선택적 압력이 흑사병이 아닌 천연두에 의해 발생했다는 것이다.[116]
- '''말라리아 저항성''': 낫적혈구병은 신체가 적혈구에서 산소 운반 물질인 헤모글로빈의 비정상적인 유형을 생성하는 혈액 장애이다. 사하라 사막 이남 아프리카의 모든 원주민의 3분의 1이 이 대립유전자를 가지고 있는데, 말라리아가 흔한 지역에서는 단 하나의 낫적혈구 대립유전자(낫적혈구 형질)만 가지고 있어도 생존 가치가 있기 때문이다.[117] 낫적혈구병의 두 대립유전자 중 하나만 가진 사람들은 말라리아에 대한 저항성이 더 강하다. 말라리아 원충인 ''플라스모디움''의 감염이 침입한 세포의 낫 모양 변형으로 막히기 때문이다.
- '''항생제 내성''': 사실상 모든 세균은 항생제에 노출되면 항생제 내성을 나타낸다. 실제로 박테리아 개체군은 이미 항생제 선택 하에서 선택되는 그러한 돌연변이를 가지고 있다.[118] 분명히 이러한 돌연변이는 박테리아에게만 유익하지 감염된 사람에게는 유익하지 않다.
- '''락테이스 지속성''': 돌연변이는 인간이 모유 수유 후에도 락테이스 효소를 발현할 수 있게 하여 성인이 락토스를 소화할 수 있게 했는데, 이는 최근 인류 진화에서 가장 유익한 돌연변이 중 하나일 가능성이 높다.[119]
5. 4. 부정적 영향
노랑초파리를 이용한 인위적인 돌연변이 실험 결과, 돌연변이의 약 70%는 개체에게 해로운 방향으로 진행된다.[160] 돌연변이가 개체에게 해로운 작용을 할 가능성이 높기 때문에 생물은 세포 수준에서 돌연변이를 무효화하는 DNA 수복 기제를 작동시키도록 진화하여 왔다.[161]기능에 대한 영향으로 분류되는 많은 돌연변이가 있지만, 변화의 특이성에 따라 아래와 같은 돌연변이가 발생한다.[52]
- '''기능 상실 돌연변이'''(비활성화 돌연변이라고도 함): 유전자 생성물의 기능이 감소하거나 없어지는(부분적으로 또는 완전히 비활성화됨) 결과를 초래한다. 이러한 돌연변이와 관련된 표현형은 대부분 열성이다. 기능 상실 돌연변이로 인해 발생하는 질병으로는 기텔만 증후군과 낭포성 섬유증이 있다.[53]
- '''우성 음성 돌연변이'''(반형질 돌연변이라고도 함): 야생형 대립 유전자에 길항적으로 작용하는 변경된 유전자 생성물을 가진다. 이러한 돌연변이는 일반적으로 변경된 분자 기능(종종 비활성)을 초래하며, 우성 또는 반우성 표현형으로 특징지어진다. 인간에게서 우성 음성 돌연변이는 암과 관련이 있다(예: 유전자 p53, ATM, CEBPA, 및 PPAR감마의 돌연변이). 마르팡 증후군은 FBN1 유전자(15번 염색체에 위치)의 돌연변이에 의해 발생하며, 이 유전자는 세포외 기질의 당단백질 성분인 피브릴린-1을 암호화한다. 마르팡 증후군은 또한 우성 음성 돌연변이와 반수불충분성의 예이다.
- '''치사 돌연변이''': 발생 중에 발생할 때 유기체의 빠른 사망을 초래하며, 발달된 유기체의 수명을 크게 단축시킨다. 우성 치사 돌연변이에 의해 발생하는 질병의 예로는 헌팅턴병이 있다.
- '''무형질 돌연변이'''(널 돌연변이): 유전자의 기능을 완전히 금지하는 기능 상실 돌연변이의 한 형태이다. 아토피성 습진과 피부염 증후군은 필라그린을 활성화하는 유전자의 널 돌연변이에 의해 발생하는 일반적인 질병이다.
- '''억제 돌연변이''': 이중 돌연변이가 정상적으로 나타나게 하는 돌연변이의 한 유형이다. 억제 돌연변이에는 유전자 내 및 유전자 외 억제 돌연변이 두 가지 유형이 있다. 이러한 유형의 돌연변이로 인해 발생하는 일반적인 질병은 알츠하이머병이다.[54]
사람의 CCR5(CCR5-Δ32)에서 특정 32개 염기쌍 결실은 동형접합자에게 HIV 저항성을 부여하고 이형접합자에서 AIDS 발병을 지연시킨다.[114] 해로운 돌연변이의 예로는 낫적혈구병이 있는데, 이는 신체가 적혈구에서 산소 운반 물질인 헤모글로빈의 비정상적인 유형을 생성하는 혈액 장애이다. 낫적혈구병의 두 대립유전자 중 하나만 가진 사람들은 말라리아에 대한 저항성이 더 강하다. 말라리아 원충인 ''플라스모디움''의 감염이 침입한 세포의 낫 모양 변형으로 막히기 때문이다.[117]
유전자 돌연변이는 DNA 복제 과정에서의 실수, 화학물질에 의한 DNA 손상 및 복제 오류, 방사선 조사에 의한 DNA 또는 염색체 손상, 트랜스포존(transposon)의 이동에 의한 유전자 파괴 등으로 인해 발생한다.
점돌연변이에서 아미노산이 변화된 단백질은 변이 전의 기능을 유지할 수 없는 경우가 많으며, 이러한 변이체는 생존에 불리한 경우가 많다.
염색체 이상은 염색체의 구조 이상이나, 그에 따른 장애가 발생하는 변이이다. 체세포의 돌연변이는 종양 발생으로 이어질 수 있다.
세균이나 바이러스는 돌연변이에 의해 백신의 변이 또는 치료제에 대한 내성을 획득하는 경우가 있으며, 치료 및 예방을 어렵게 한다.
6. 보상 돌연변이
DNA 코딩 영역의 돌연변이로 인한 DNA 변화는 단백질 서열에 오류를 일으켜 부분적 또는 완전히 기능하지 않는 단백질을 생성할 수 있다. 각 세포는 올바르게 기능하기 위해 수천 개의 단백질이 적절한 시간에 적절한 위치에서 기능해야 한다. 돌연변이가 신체에서 중요한 역할을 하는 단백질을 변화시키면 질병이 발생할 수 있다. 다양한 초파리(*Drosophila*) 종 간의 유전자 비교에 대한 한 연구에 따르면, 돌연변이가 단백질을 변화시키면 그 돌연변이는 아미노산 다형성의 약 70%가 해로운 영향을 미치는 것으로 추정되며, 나머지는 중립적이거나 약간 유익한 것으로 보아 대부분 해로울 가능성이 높다.[8] 일부 돌연변이는 유전자의 DNA 염기 서열을 변경하지만 유전자에 의해 생성되는 단백질은 변경하지 않는다. 연구에 따르면 효모의 비코딩 DNA에서 점 돌연변이의 7%만이 해롭고, 코딩 DNA에서는 12%만이 해롭다는 것을 보여준다. 나머지 돌연변이는 중립적이거나 약간 유익하다.[111]
7. 돌연변이 연구의 미래
생식 세포(생식 세포)에 돌연변이가 존재하면, 모든 세포에 돌연변이를 지닌 자손을 낳을 수 있는데, 이는 유전 질환의 경우에 해당한다. 특히, 생식 세포 내 DNA 복구 유전자에 돌연변이가 있는 경우, 이러한 생식세포 돌연변이를 가진 사람들은 암 위험이 증가할 수 있다. 이러한 생식세포 돌연변이 34가지 목록은 DNA 복구 결핍 질환 문서에서 확인할 수 있다. 한 가지 예로 ''OCA1'' 또는 ''OCA2'' 유전자에서 발생하는 돌연변이인 알비니즘이 있다. 이 질환을 가진 사람들은 여러 유형의 암, 다른 질환에 더 취약하며 시력이 손상될 수 있다.[1]
DNA 손상은 DNA가 복제될 때 오류를 일으킬 수 있으며, 이 복제 오류는 유전자 돌연변이를 일으켜 유전 질환을 유발할 수 있다. DNA 손상은 세포의 DNA 복구 시스템에 의해 복구된다. 각 세포는 효소가 DNA의 손상을 인식하고 복구하는 여러 경로를 가지고 있다. DNA는 여러 가지 방식으로 손상될 수 있기 때문에 DNA 복구 과정은 신체가 질병으로부터 자신을 보호하는 중요한 방법이다. DNA 손상이 돌연변이를 일으킨 후에는 돌연변이를 복구할 수 없다.[1]
8. 한국의 돌연변이 연구 현황 및 정책 방향
한국에서는 돌연변이와 관련된 유전 질환 연구가 활발하게 진행되고 있다. 특히, 생식 세포 돌연변이로 인한 유전 질환과 암 발생 위험 증가에 대한 연구가 이루어지고 있다. 예를 들어, ''OCA1'' 또는 ''OCA2'' 유전자 돌연변이로 발생하는 알비니즘은 여러 유형의 암, 다른 질환, 시력 손상 등을 유발할 수 있어 이에 대한 연구가 중요하다.[112]
DNA 손상은 DNA 복제 오류를 일으켜 유전자 돌연변이와 유전 질환을 유발할 수 있다. DNA 손상은 세포의 DNA 복구 시스템에 의해 복구되지만, DNA 손상이 돌연변이를 일으킨 후에는 복구할 수 없다. 따라서 DNA 복구 과정은 신체가 질병으로부터 자신을 보호하는 중요한 방법이다.[113]
체세포 돌연변이는 생물체의 체세포에서 발생하며, 동일 생물체 내에서 해당 세포의 모든 자손에게 나타난다. 이러한 체세포 돌연변이의 축적은 정상 세포가 암세포로 악성화되는 원인 중 하나이다. 이형접합 기능 상실 돌연변이가 있는 세포는 정상 복제본이 자발적으로 체세포 돌연변이를 일으킬 때까지 정상적으로 기능할 수 있지만, 이러한 돌연변이 비율을 측정하기는 어렵다. 이 비율을 측정하는 것은 암 발생 예측에 중요하다.[113]
점 돌연변이는 DNA 복제 중 발생하는 자발적인 돌연변이에서 발생할 수 있으며, 돌연변이율은 UV선, X선 등의 물리적 요인이나 화학적 요인에 의해 증가될 수 있다. 암과 관련된 돌연변이원은 암 예방 연구에 활용된다.
참조
[1]
웹사이트
mutation Learn Science at Scitable
https://www.nature.c[...]
Nature Education
2018-09-24
[2]
논문
Microhomology-Mediated End Joining: A Back-up Survival Mechanism or Dedicated Pathway?
2015-11-01
[3]
논문
Repair of naturally occurring mismatches can induce mutations in flanking DNA
2014-04-01
[4]
논문
Error-Prone Repair of DNA Double-Strand Breaks
2016-01-01
[5]
논문
The molecular biology of cancer
2000-12-01
[6]
논문
Pesticide resistance via transposition-mediated adaptive gene truncation in Drosophila
2005-07-01
[7]
논문
Shaping bacterial genomes with integrative and conjugative elements
2004-06-01
[8]
논문
Prevalence of positive selection among nearly neutral amino acid replacements in Drosophila
2007-04-01
[9]
논문
Genetic damage, mutation, and the evolution of sex
1985-09-01
[10]
논문
Mechanisms of change in gene copy number
2009-08-01
[11]
서적
From DNA to Diversity: Molecular Genetics and the Evolution of Animal Design
Wiley-Blackwell
[12]
논문
Studying genomes through the aeons: protein families, pseudogenes and proteome evolution
2002-05-01
[13]
논문
Protein families and their evolution-a structural perspective
2005-07-01
[14]
논문
The origin of new genes: glimpses from the young and old
2003-11-01
[15]
논문
The evolutionary mechanics of domain organization in proteomes and the rise of modularity in the protein world
2009-01-01
[16]
논문
Evolution of colour vision in vertebrates
1998-05-01
[17]
논문
The modulation of DNA content: proximate causes and ultimate consequences
1999-04-01
[18]
논문
Gene duplication: the genomic trade in spare parts
2004-07-01
[19]
논문
The evolution and functional diversification of animal microRNA genes
2008-10-01
[20]
논문
Darwinian alchemy: Human genes from noncoding DNA
2009-10-01
[21]
논문
Testing the chromosomal speciation hypothesis for humans and chimpanzees
2004-05-01
[22]
논문
Chromosome speciation: humans, Drosophila, and mosquitoes
2005-05-01
[23]
논문
The role of selfish genetic elements in eukaryotic evolution
2001-08-01
[24]
논문
Alu elements as regulators of gene expression
2006-11-01
[25]
논문
The distribution of fitness effects of new mutations
http://www.lifesci.s[...]
2010-09-06
[26]
서적
Fundamentals of Polymer Physics and Molecular Biophysics
https://books.google[...]
Cambridge University Press
2015-01-01
[27]
웹사이트
What Is Apoptosis? Memorial Sloan Kettering Cancer Center
https://www.mskcc.or[...]
2014-05-16
[28]
서적
Dear Mr. Darwin: Letters on the Evolution of Life and Human Nature
https://books.google[...]
University of California Press
2000-01-01
[29]
서적
Genetics and Evolution of Infectious Diseases
https://books.google[...]
Elsevier
2017-01-12
[30]
웹사이트
Molecular Biology of the Cell: Studying Gene Expression and Function
https://www.ncbi.nlm[...]
2002-01-01
[31]
뉴스
Cancer Is Partly Caused By Bad Luck, Study Finds
https://www.npr.org/[...]
[32]
뉴스
Older fathers pass on more genetic mutations, study shows
https://www.theguard[...]
2012-08-22
[33]
논문
Oxidants, antioxidants, and the degenerative diseases of aging
1993-09-01
[34]
웹사이트
Mutation, Mutagens, and DNA Repair
http://www-personal.[...]
[35]
논문
Quantum and classical effects in DNA point mutations: Watson-Crick tautomerism in AT and GC base pairs
2021-02-01
[36]
논문
An open quantum systems approach to proton tunnelling in DNA
https://www.nature.c[...]
2022-05-05
[37]
논문
Mutation frequency and specificity with age in liver, bladder and brain of lacI transgenic mice
2000-03-01
[38]
논문
DNA sequence analysis of spontaneous mutagenesis in Saccharomyces cerevisiae
1998-04-01
[39]
논문
The mechanism of double-strand DNA break repair by the nonhomologous DNA end-joining pathway
2010-07-01
[40]
웹사이트
PDB 1JDG
http://www.rcsb.org/[...]
2015-12-31
[41]
논문
Ochratoxin A: An overview on toxicity and carcinogenicity in animals and humans
2007-01-01
[42]
논문
UVA radiation is highly mutagenic in cells that are unable to repair 7,8-dihydro-8-oxoguanine in Saccharomyces cerevisiae
2005-09-01
[43]
논문
What is mutation? A chapter in the series: How microbes "jeopardize" the modern synthesis
2019-04-01
[44]
논문
Mutation as a stress response and the regulation of evolvability
2007-01-01
[45]
논문
Yeasts acquire resistance secondary to antifungal drug treatment by adaptive mutagenesis
2012-07-31
[46]
웹사이트
The clinical impact of DNA sequence changes
http://www.thetgmi.o[...]
2017-06-27
[47]
논문
The Difference Between Spontaneous and Base-Analogue Induced Mutations of Phage T4
1959-04-01
[48]
논문
The specific mutagenic effect of base analogues on Phage T4
1959-06-01
[49]
문서
References for the image are found in Wikimedia Commons page at: Commons:File:Notable mutations.svg#References
[50]
백과사전
Mutation
http://www.eoearth.o[...]
Environmental Information Coalition, National Council for Science and the Environment
2010-10-12
[51]
논문
ALS: a disease of motor neurons and their nonneuronal neighbors
2006-10-01
[52]
논문
Predicting the functional impact of protein mutations: application to cancer genomics
2011-09-01
[53]
논문
Loss-of-function genetic tools for animal models: cross-species and cross-platform differences
2017-01-01
[54]
논문
Map positions and specificities of suppressor mutations in Escherichia coli K-12
1965-08-01
[55]
논문
Neomorphic mutations create therapeutic challenges in cancer
2017-03-01
[56]
논문
Back mutation can produce phenotype reversion in Bloom syndrome somatic cells
2001-02-01
[57]
논문
On causal roles and selected effects: our genome is mostly junk
2017-12-01
[58]
논문
Phenotypic landscape of a bacterial cell
2011-01-01
[59]
논문
Tn-seq: high-throughput parallel sequencing for fitness and genetic interaction studies in microorganisms
2009-10-01
[60]
논문
Hundreds of variants clustered in genomic loci and biological pathways affect human height
2010-10-01
[61]
논문
The pattern of neutral molecular variation under the background selection model
1995-12-01
[62]
논문
Quantifying the genomic decay paradox due to Muller's ratchet in human mitochondrial DNA
2006-04-01
[63]
서적
Molecular Genetics of Development
[64]
논문
Imperfect genes, Fisherian mutation and the evolution of sex
1997-04-01
[65]
논문
Refinement of evolutionary medicine predictions based on clinical evidence for the manifestations of Mendelian diseases
2019-12-01
[66]
논문
Toward a realistic model of mutations affecting fitness
2003-03-01
[67]
논문
Understanding quantitative genetic variation
2002-01-01
[68]
논문
The distribution of fitness effects caused by single-nucleotide substitutions in an RNA virus
2004-06-01
[69]
논문
Distribution of fitness and virulence effects caused by single-nucleotide substitutions in Tobacco Etch virus
2007-12-01
[70]
논문
Mutational fitness effects in RNA and single-stranded DNA viruses: common patterns revealed by site-directed mutagenesis studies
2010-06-01
[71]
논문
Distribution of fitness effects caused by single-nucleotide substitutions in bacteriophage f1
2010-06-01
[72]
논문
Distribution of fitness effects caused by random insertion mutations in Escherichia coli
1998-03-01
[73]
논문
Experimental illumination of a fitness landscape
2011-05-01
[74]
논문
High frequency of cryptic deleterious mutations in Caenorhabditis elegans
1999-09-01
[75]
논문
Inferring the distribution of mutational effects on fitness in Drosophila
2006-09-01
[76]
논문
The distribution of fitness effects of new deleterious amino acid mutations in humans
2006-06-01
[77]
논문
Bayesian analysis suggests that most amino acid replacements in Drosophila are driven by positive selection
2003-08-01
[78]
논문
Estimating the distribution of fitness effects from DNA sequence data: implications for the molecular clock
2003-09-01
[79]
서적
The Neutral Theory of Molecular Evolution
Cambridge University Press
[80]
논문
Evolutionary rate at the molecular level
1968-02-01
[81]
논문
Within- and between-species DNA sequence variation and the 'footprint' of natural selection
1999-09-01
[82]
논문
The genomic rate of adaptive evolution
2006-10-01
[83]
논문
Molecular Evolution Over the Mutational Landscape
1984-09-01
[84]
논문
The distribution of fitness effects among beneficial mutations
2003-04-01
[85]
논문
Distribution of fitness effects among beneficial mutations before selection in experimental populations of bacteria
2006-04-01
[86]
논문
An empirical test of the mutational landscape model of adaptation using a single-stranded DNA virus
2005-04-01
[87]
논문
Fitness effects of advantageous mutations in evolving Escherichia coli populations
2001-01-01
[88]
백과사전
Somatic cell genetic mutation
https://theodora.com[...]
Information Technology Associates
2007-06-30
[89]
백과사전
Compound heterozygote
http://www.medicinen[...]
WebMD
2012-06-14
[90]
웹사이트
'RB1' Genetics
http://www.daisyfund[...]
[91]
백과사전
somatic mutation {{!}} genetics
https://www.britanni[...]
2017-03-31
[92]
서적
Genetics Principles and Analysis
https://archive.org/[...]
Jones and Bartlett Publishers
[93]
논문
Differences between germline and somatic mutation rates in humans and mice
2017-05-01
[94]
서적
Molecular Biology of the Cell
Garland Science
[95]
논문
Conditional mutations in Drosophila melanogaster: On the occasion of the 150th anniversary of G. Mendel's report in Brünn
2015-07-01
[96]
논문
High-frequency generation of conditional mutations affecting Drosophila melanogaster development and life span
http://www.genetics.[...]
2017-03-21
[97]
논문
Strategies to achieve conditional gene mutation in mice
2014-04-01
[98]
논문
Creating conditional mutations in mammals
1996-05-01
[99]
논문
Temperature-sensitive mutations made easy: generating conditional mutations by using temperature-sensitive inteins that function within different temperature ranges
2009-09-01
[100]
논문
Mutation nomenclature extensions and suggestions to describe complex mutations: a discussion
2000-01-01
[101]
논문
Parental influence on human germline de novo mutations in 1,548 trios from Iceland
2017-09-01
[102]
논문
WHy do species very in their rate of molecular evolution?
2009-01-01
[103]
논문
The population genetics of mutations: good, bad, and indifferent
Philosophical transactions of the Royal Society of London
2010-01-01
[104]
논문
DE novo mutations, genetic mosaicism, and genetic disease
2022-01-01
[105]
논문
De novo mutations, genetic mosaicism, and genetic disease
2022-01-01
[106]
논문
New insights into the generation and role of de novo mutations in health and disease
2016
[107]
논문
The mechanisms of UV mutagenesis
https://pubmed.ncbi.[...]
J Radiat Res
2023-12-09
[108]
뉴스
Study challenges evolutionary theory that DNA mutations are random
https://phys.org/new[...]
2022-02-12
[109]
논문
Mutation bias reflects natural selection in Arabidopsis thaliana
2022-02-01
[110]
논문
Random with Respect to Fitness or External Selection? An Important but Often Overlooked Distinction
2023
[111]
논문
A catalog of neutral and deleterious polymorphism in yeast
2008-08-01
[112]
논문
Ubiquitous somatic mutations in simple repeated sequences reveal a new mechanism for colonic carcinogenesis
1993-06-01
[113]
논문
A quantitative measurement of the human somatic mutation rate
2005-09-01
[114]
논문
The coreceptor mutation CCR5Delta32 influences the dynamics of HIV epidemics and is selected for by HIV
2001-08-01
[115]
에피소드
Mystery of the Black Death
https://www.pbs.org/[...]
2015-10-10
[116]
논문
Evaluating plague and smallpox as historical selective pressures for the CCR5-Delta 32 HIV-resistance allele
2003-12-01
[117]
웹사이트
Frequently Asked Questions [FAQ's]
https://web.archive.[...]
2010-04-16
[118]
논문
Evolutionary Trajectories to Antibiotic Resistance
2017-09-01
[119]
논문
On the Evolution of Lactase Persistence in Humans
2017-08-01
[120]
논문
Molecular signatures of resource competition: Clonal interference favors ecological diversification and can lead to incipient speciation
International Journal of Organic Evolution
2021
[121]
논문
Compensated pathogenic deviations
2011-08-01
[122]
논문
Compensating for the meltdown: The critical effective size of a population with deleterious and compensatory mutations
https://www.jstor.or[...]
2003
[123]
논문
Population size and the rate of evolution
https://www.scienced[...]
2014-01-01
[124]
논문
Structural genomics of RNA
http://www.nature.co[...]
2000-11-01
[125]
논문
From Bad to Good: Fitness Reversals and the Ascent of Deleterious Mutations
2006-10-20
[126]
논문
How Mutational Networks Shape Evolution: Lessons from RNA Models
https://www.annualre[...]
2007-12-01
[127]
논문
Intragenic compensation through the lens of deep mutational scanning
https://doi.org/10.1[...]
2022-10-01
[128]
논문
Missense meanderings in sequence space: a biophysical view of protein evolution
https://pubmed.ncbi.[...]
2005-09-01
[129]
논문
Characterization of Compensated Mutations in Terms of Structural and Physico-Chemical Properties
https://www.scienced[...]
2007-01-05
[130]
논문
Pervasive Cryptic Epistasis in Molecular Evolution
2010-10-21
[131]
논문
c-di-AMP Is a New Second Messenger in Staphylococcus aureus with a Role in Controlling Cell Size and Envelope Stress
2011-09-01
[132]
논문
Whole-genome sequencing of rifampicin-resistant Mycobacterium tuberculosis strains identifies compensatory mutations in RNA polymerase genes
2012-01-01
[133]
논문
The competitive cost of antibiotic resistance in Mycobacterium tuberculosis
https://pubmed.ncbi.[...]
2006-06-30
[134]
논문
Compensatory evolution in rifampin-resistant Escherichia coli
2000-12-01
[135]
논문
Stability-mediated epistasis constrains the evolution of an influenza protein
2013-05-14
[136]
논문
Compensatory mutations are repeatable and clustered within proteins
2009-05-22
[137]
논문
Intragenic compensation through the lens of deep mutational scanning
https://doi.org/10.1[...]
2022-10-01
[138]
논문
Evolution by Small Steps and Rugged Landscapes in the RNA Virus ϕ6
https://academic.oup[...]
1999-03-01
[139]
논문
Full restoration of viral fitness by multiple compensatory co-mutations in the nucleoprotein of influenza A virus cytotoxic T-lymphocyte escape mutants
2005
[140]
논문
The role of compensatory neutral mutations in molecular evolution
https://doi.org/10.1[...]
1985-07-01
[141]
논문
A global reference for human genetic variation
2015
[142]
논문
Clan Genomics and the Complex Architecture of Human Disease
2011
[143]
논문
De novo mutations in human genetic Disease
https://www.nature.c[...]
2023-12-09
[144]
문서
mutagen
[145]
문서
mutation
[146]
문서
Mutation
[147]
문서
neutral mutation
[148]
문서
silent mutation
[149]
문서
back mutation
[150]
문서
suppressor mutation
[151]
문서
adaptive mutation
[152]
문서
point mutation
[153]
문서
missense mutation
[154]
문서
nonsense mutation
[155]
문서
frameshift mutation
[156]
논문
卵浸漬法によるN-メチル-N-ニトロソウレアのカイコの発生に及ぼす影響と突然変異誘発
https://doi.org/10.1[...]
일본 잠사학회
[157]
논문
イオンビームによる植物の突然変異誘発
https://doi.org/10.3[...]
日本アイソトープ協会
[158]
논문
4. ライサイエンスへの利用 4.3炭素およびネオンイオンビームによって誘発された細胞死と突然変異
https://search.jamas[...]
日本アイソト-プ協会
[159]
논문
水稲における燐欠乏による突然変異の誘起
日本育種学会
[160]
논문
Prevalence of positive selection among nearly neutral amino acid replacements in Drosophila
http://www.pubmedcen[...]
2007
[161]
논문
The molecular biology of cancer
2000
[162]
논문
The evolution of mutation rates: separating causes from consequences
2000
[163]
논문
Mutation rates among RNA viruses
http://www.pnas.org/[...]
1999
[164]
논문
Rapid evolution of RNA genomes
1982
[165]
논문
Mechanisms of change in gene copy number
2009
[166]
서적
From DNA to Diversity: Molecular Genetics and the Evolution of Animal Design
Oxford: Blackwell Publishing
[167]
논문
Studying genomes through the aeons: protein families, pseudogenes and proteome evolution
2002
[168]
논문
Protein families and their evolution-a structural perspective
2005
[169]
논문
The origin of new genes: glimpses from the young and old
2003-11
[170]
논문
Testing the chromosomal speciation hypothesis for humans and chimpanzees
http://www.pubmedcen[...]
2004
[171]
논문
Chromosome speciation: humans, Drosophila, and mosquitoes
http://www.pnas.org/[...]
2005
[172]
논문
The role of selfish genetic elements in eukaryotic evolution
2001
[173]
서적
생명과학
동화기술
[174]
논문
The Difference between Spontaneous and Base-Analogue Induced Mutations of Phage T4
http://www.pubmedcen[...]
1959-04
[175]
논문
The Specific Mutagenic Effect of Base Analogues on Phage T4
1959
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com