틸로사우루스
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
틸로사우루스는 백악기 후기에 살았던 멸종된 해양 파충류의 한 속으로, 모사사우루스과에 속한다. 1869년 에드워드 드링커 코프에 의해 처음 명명되었으며, 긴 주둥이와 강력한 턱을 가진 것이 특징이다. 북아메리카에서 발견된 화석을 바탕으로, 틸로사우루스는 12~15m까지 자라며, 다양한 해양 생물을 먹이로 삼는 최상위 포식자였던 것으로 추정된다. 틸로사우루스는 온혈 동물이었으며, 꼬리를 이용한 카랑기폼 운동을 통해 헤엄쳤을 것으로 보인다. 현재까지 8종이 인정되고 있다.

더 읽어볼만한 페이지
- 모사사우루스과 - 프로그나토돈
프로그나토돈은 백악기 후기에 살았던 모사사우루스과의 멸종된 해양 파충류로, 강력한 턱과 튼튼한 이빨을 가졌으며, 앨버타의 베어포 지층 등에서 화석이 발견되었다. - 모사사우루스과 - 플로토사우루스
플로토사우루스는 백악기 후기에 번성했던 모사사우루스과의 멸종된 해양 파충류 속으로, 좁은 지느러미와 큰 꼬리지느러미, 유선형 몸매를 통해 효율적으로 움직였으며, 캘리포니아에서 발견된 최초의 모사사우루스류이자 활동적인 포식자로 추정되는 '플로토사우루스 베니소니' 단일 종으로 분류된다. - 백악기의 도마뱀 - 모사사우루스
모사사우루스는 백악기 후기에 바다에서 살았던 몸길이 10m에서 15m에 이르는 대형 해양 파충류로, 완전한 수중 생활에 적응했으며 당시 바다의 최상위 포식자 중 하나였고, 전 세계에서 화석이 발견된다. - 백악기의 도마뱀 - 모사사우루스과
모사사우루스과는 백악기 후기에 번성한 해양 파충류로, 다양한 크기의 종이 존재하며 뛰어난 수영 능력과 특수한 턱 구조를 통해 최상위 포식자로 군림했고, 화석 발견 이후 활발한 고생물학적 연구가 진행되고 있다. - 1872년 기재된 화석 분류군 - 하스트수리
하스트수리는 뉴질랜드에 서식했던 멸종된 대형 맹금류로, 흰점어깨수리, 쇠독수리와 가까운 관계이며, 모아를 주로 사냥했으나 마오리족의 등장과 모아의 멸종으로 멸종되었다. - 1872년 기재된 화석 분류군 - 아가타우마스
아가타우마스는 1872년 와이오밍 주에서 발견된 각룡류 공룡의 한 속으로, 한때 육상 동물 중 가장 큰 것으로 여겨졌으나, 현재는 불확실한 분류로 트리케라톱스의 화석일 가능성이 제기된다.

2. 발견 및 명명
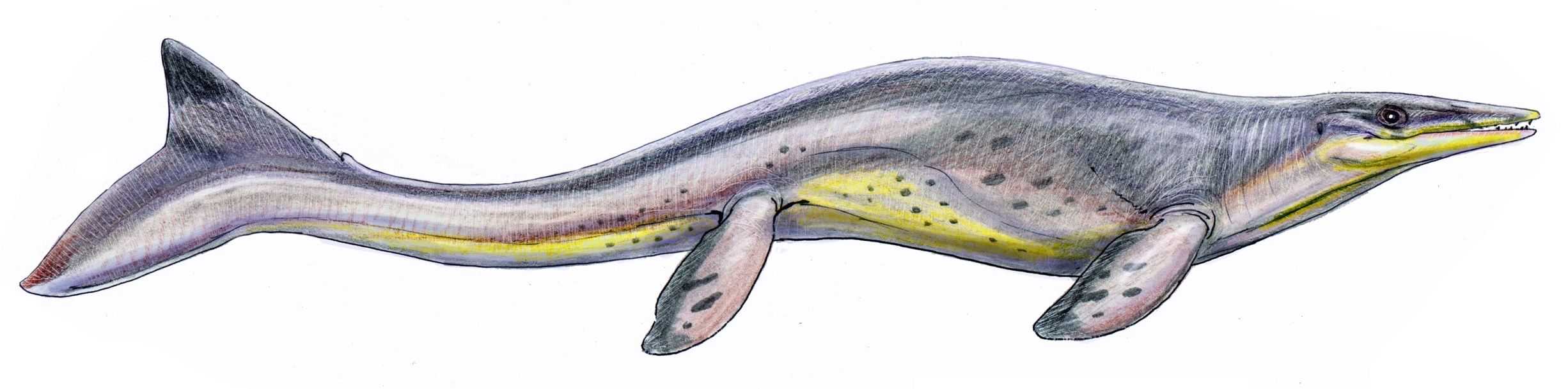
''틸로사우루스''는 클리다스테스와 플라테카르푸스에 이어 북아메리카에서 세 번째로 명명된 모사사우루스과 속이며, 캔자스주에서는 처음이었다.[9] 속명은 '혹 도마뱀'을 의미하며, 틸로사우루스의 특징인 길쭉한 부리(코)에서 유래했다.
틸로사우루스의 두드러진 특징은 원통형의 긴 전상악골(주둥이)이며, 다른 모사사우루스과 파충류와 달리 전상악골 전방에 이빨이 전혀 없었다.[100] 틸로사우루스는 상악에 24~26개, 구개에 20~22개, 하악에 26개의 이빨이 있었다.[84] 척추는 두개골에서 둔부 사이에 29~30개, 둔부에 6~7개, 골반과 함께 꼬리에 33~34개, 꼬리 끝에 56~58개가 있었다.[84]
초기 복원에서는 오동정된 기관 연골에 기초하여 등지느러미가 복원되기도 했다.[85][86]
1911년 캔자스주 월리스 군에서 C. D. 뱅커에 의해 발견된 ''T. proriger''의 거대한 표본은 캔자스 대학교 자연사 박물관에 전시되어 있다. 1918년 찰스 헤이젤리어스 스턴버그는 복부에 사경룡을 포함한 틸로사우루스 화석을 발견하여 스미소니언 박물관에 소장되었다.[91] 2009년에는 앨런 콤로스키에 의해 길이가 10미터인 틸로사우루스가 캔자스주에서 발견되어 위치타의 Museum of World Treasures에 전시되어 있다.[92] 틸로사우루스는 서스캐처원주와 사우스다코타주의 지층에서도 보고되었다.[93]
2. 1. 모식 표본
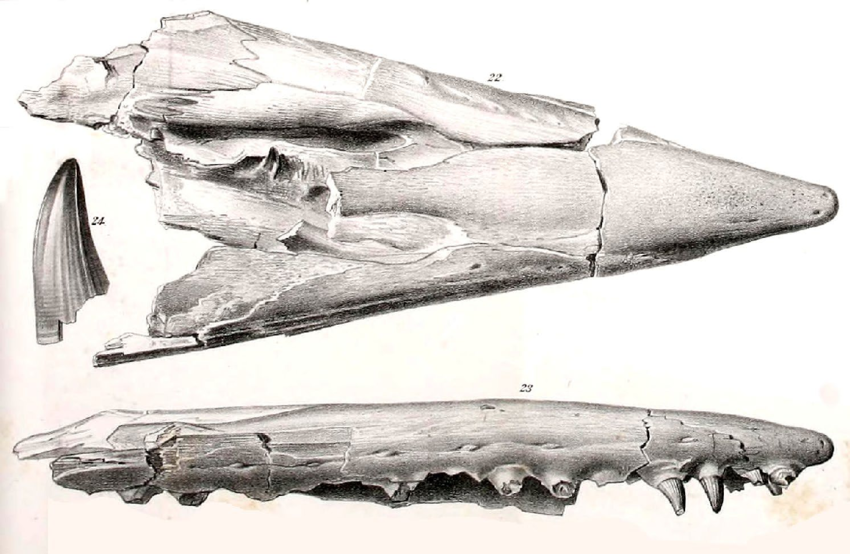
에드워드 드링커 코프는 1869년 캔자스주 니오브라라 지층에서 발견된 약 1.5m 길이의 파편적인 두개골과 13개의 척추뼈를 기반으로 틸로사우루스 모식 표본(MCZ 4374)을 처음 기술했다.[9][10][11] 코프는 처음에 이 화석을 '마크로사우루스 프로리게르(''Macrosaurus proriger'')'로 명명했다.[9] 종명인 ''proriger''는 "앞 부분을 가진"이라는 의미로, 표본의 독특한 뱃머리 모양의 길쭉한 부리를 지칭한다.[12] 1870년, 코프는 MCZ 4374에 대한 더 상세한 설명을 발표하면서 이 종을 다른 유럽 속인 ''Liodon''으로 옮겼다.[11]
2. 2. 속명의 확립
에드워드 드링커 코프와 오트니엘 찰스 마시의 경쟁으로 인해 틸로사우루스의 초기 분류 역사는 복잡하게 얽혀 있다.[90] 1869년 코프는 캔자스주에서 발견된 파편적인 두개골과 13개의 척추뼈를 바탕으로 ''Macrosaurus proriger''라는 이름을 처음 제안했다.[89] 1년 후, 코프는 이 표본을 리오돈으로 재분류했다.[11]1872년, 마시는 이 표본이 리오돈과는 다른 새로운 속(Genus)에 속한다고 주장하며 '리노사우루스(''Rhinosaurus'')'라는 이름을 제안했지만, 이 이름은 이미 다른 동물에게 사용되고 있었다.[90] 코프는 '람포사우루스(''Rhamphosaurus'')'라는 다른 이름을 제안했으나, 이 역시 이미 사용 중인 이름이었다.[15] 결국 마시는 1872년에 '틸로사우루스'라는 새로운 속명을 제안했고,[15] 1873년 조셉 레이디가 ''Rhinosaurus proriger''를 ''Tylosaurus proriger''로 재분류하면서 현재의 학명이 확립되었다.[16][17]
3. 형태 및 특징
틸로사우루스는 길고 원통 모양인 전상악골(주둥이)이 두드러진 특징이며, 속명은 여기에서 유래되었다. 다른 모사사우루스과 파충류와 달리, 틸로사우루스는 전상악골 앞쪽에 이빨이 전혀 없어 뼈 돌출부에는 이빨이 존재하지 않았다.[100] 상악에는 24~26개, 구개에는 20~22개, 하악에는 26개의 이빨이 나 있었다.[84] 두개골에서 둔부 사이에는 29~30개, 둔부에는 6~7개, 골반과 함께 꼬리에는 33~34개의 척추가 있었으며, 꼬리 끝에는 56~58개의 미추가 있었다.[84]
초기 복원에서는 오동정된 기관 연골 때문에 등지느러미가 있는 것으로 묘사되었는데, 당시에는 이러한 돌기를 가진 모사사우루스과를 그리는 것이 일반적이었다.[85][86]
1870년대 후반부터 틸로사우루스의 피부 화석이 발견되어 비늘 형태가 묘사되었다. 이 비늘은 작고 다이아몬드 모양이었으며, 방울뱀처럼 비스듬한 열로 배열되어 있었다. 각 비늘은 상어의 치아와 유사하게 용골이 있어 수중 저항[54]과 피부 반사를 줄이는 데 도움이 되었을 것이다.[56]
2014년 연구에서는 ''T. nepaeolicus'' 표본의 비늘에서 유멜라닌 흔적이 발견되어, 살아있는 바다거북처럼 어두운 색을 띠었을 가능성이 제기되었다. 어두운 색은 열 흡수를 늘려 추운 환경에서도 높은 체온을 유지하도록 돕고, 유아기에 빠른 성장을 촉진했을 수 있다. 또한 반사되지 않는 어두운 색은 위장에 유리했을 것이다.
3. 1. 크기

''틸로사우루스''는 알려진 모사사우루스과 중 가장 큰 종 중 하나였다. 캔자스 대학교 자연사 박물관에 보관된 ''T. proriger''의 골격(별명 "벙커", KUVP 5033)은 길이가 12m에서 15.8m 사이로 추정된다.[16][19] 스턴버그 자연사 박물관에 보관된 또 다른 ''T. proriger''의 부분 골격(FHSM VP-2496)은 더 큰 개체였을 수 있다. 에버하트는 이 표본이 벙커의 12m 추정치와 비교했을 때 14m 크기의 개체에서 나왔을 것으로 추정했다.
이 속은 코프의 법칙을 보여주는데, 지질학적 시간이 지남에 따라 신체 크기가 일반적으로 증가하는 경향을 보였다.[16] 북아메리카에서 투로니아[20] 및 코니아크 시대(90-86 mya)의 초기 ''틸로사우루스'' 대표종, 즉 초기 ''T. nepaeolicus''와 그 선구종은 일반적으로 길이가 5m에서 7m 사이였고[16] 무게는 200kg에서 500kg 사이였다.[21] 산토니아 시대(86-83 mya) 동안, ''T. nepaeolicus''와 새롭게 나타난 ''T. proriger''는 길이가 8m에서 9m 사이였으며[16] 무게는 약 1100kg였다.[22] 초기 캄파니아 시대까지 ''T. proriger''는 13m에서 14m의 길이에 도달했다.
에버하트는 모사사우루스과가 평생 동안 계속 성장했기 때문에, 매우 나이가 많은 일부 ''틸로사우루스'' 개체는 최대 길이가 20m에 달했을 수도 있다고 추정했다. 그러나 그는 그러한 크기를 시사하는 화석 증거가 부족하고, 그러한 크기의 개체가 보존될 가능성이 낮다고 강조했다.
다른 캄파니아-마스트리히트 시대 종들도 이와 비슷하게 컸다. ''T. bernardi''에 대한 가장 최근의 최대 추정치는 린드그렌(2005)에 의해 12.2m이다. 역사적으로 이 종은 15m에서 17m의 더 큰 크기로 잘못 추정되기도 했다.[23] 로열 서스캐처원 박물관의 ''T. saskatchewanensis'' 재구성은 전체 길이가 9.75m를 초과할 것으로 추정했다.[24] 캐나다 화석 발견 센터에 전시된 ''T. pembinensis''의 조립된 골격("브루스"라는 별명)은 길이가 13.05m이며, 2014년에 "전시된 가장 큰 모사사우루스"로 기네스 세계 기록에 등재되었다.[25] 그러나 그 골격은 불라드와 칼드웰(2010)이 종의 척추뼈 수를 과장된 것으로 재평가하기 전에 전시용으로 조립되었다.[26][27] ''T.'' "borealis"는 전체 길이가 6.5m에서 8m 사이로 추정된다.[28]
모사사우루스 호프마니와 함께 모사사우루스과에서 가장 큰 속 중 하나이며, 동속일 가능성이 있는[84]하이노사우루스 베르나르디는 전체 길이 12.2m, ''T. pembinensis''도 그에 필적하는 크기에 달했다[87].''T. proriger''는 틸로사우루스 속에서 가장 큰 종이며, 전체 길이는 14m에 달했다[88]。
3. 2. 두개골
틸로사우루스의 가장 두드러진 특징은 코에서 튀어나온 길쭉하고 이빨이 없는 주둥이인데, 속명은 여기서 유래되었다. 이것은 앞쪽 끝의 전상악골[41]과 치골[40]의 연장으로 형성된다. 주둥이는 태어날 때 작고 예각이었지만, 빠르게 둔하고 길쭉한 "혹"으로 발달했다. 주둥이는 튼튼하게 제작되었으며, 넓고 튼튼한 비간(premaxilla, 비골, 전두골의 앞쪽 과정으로 구성)으로 지지되어 효과적인 충격 흡수와 응력 전달을 제공했다.[41] 이 때문에 틸로사우린의 주둥이는 들이받기[41][31] 위해 사용되었을 것이라고 제안되었지만, 최근 ''Taniwhasaurus''에 대한 연구[32]에서는 주둥이에서 복잡한 신경혈관계를 발견하여, 주둥이가 극도로 민감했을 가능성이 높으며, 따라서 주둥이가 들이받는 무기로 사용되었을 가능성은 낮다는 것을 시사했다. 주둥이는 주둥이와 위턱의 복측 가장자리를 따라 무작위로 흩어져 있는 구멍을 통해 삼차 신경의 말단 가지를 담고 있다.
전상악골, 위턱, 전두골은 콧구멍 또는 몸의 콧구멍 개구부를 경계짓는다. 다른 모사사우루스와 달리, 전두정골은 위턱의 긴 후방 등쪽 돌기에 의해 콧구멍의 경계에서 제외된다.[6] 콧구멍은 ''T. proriger''와 ''T. pembinensis''의 네 번째 위턱 치아 앞쪽[27]에서 열리고, ''T. nepaeolicus''의 세 번째와 네 번째 치아 사이[27], ''T. bernardi''의 네 번째 치아 뒤에 열린다.[46] 두개골 길이에 대한 콧구멍 길이의 상대적 비율은 종에 따라 다르다. ''T. proriger'' (두개골 길이의 20-27%[38]), ''T. bernardi'' (두개골 길이의 24%[27]), ''T. gaudryi'' (두개골 길이의 25-27%)[38]에서 상대적으로 짧고, ''T. pembinensis'' (두개골 길이의 28-31%)[27]에서 길다. 비골은 자유롭게 떠 있거나 비간에 가볍게 연결되어 있었고,[41] 전두골과 접촉하지 않았으며, 현존하는 도마뱀에서처럼 서로 융합되지 않았다. 비골이 ''틸로사우루스''와 다른 모사사우루스에서 두개골의 나머지 부분과 느슨하게 연결되어 있다는 것은 뼈가 자주 손실되어 매우 희귀한 이유일 수 있다. ''틸로사우루스''는 비골이 명확하게 기록된 유일한 모사사우루스 중 하나이다;[41] 다른 하나는 ''Plotosaurus''의 모식 표본이지만, 뼈 하나가 없다.[34]
외부 콧구멍은 구개에서 코안 (내부 콧구멍)으로 연결되어 콧구멍에서 목구멍으로의 통로를 제공한다.[35] ''틸로사우루스''에서, 코안은 압축된 물방울 모양이며, 서골, 구개골, 위턱에 의해 경계지어진다. 코안 앞쪽에서, 각 서골은 혀를 기반으로 하는 후각과 관련된 야콥슨 기관을 위한 창을 경계짓는다. 이것은 ''틸로사우루스''의 네 번째 위턱 치아 맞은편에서 시작되며, ''T. bernardi''의 다섯 번째 위턱 치아 바로 뒤에서 끝난다.[41] 구개 내의 부비동으로 이어지는 정맥의 출구점은 서골과 위턱 사이의 야콥슨 기관 바로 앞에서 발생한다. 이것은 야콥슨 기관 뒤에서 출구가 발생하는 현존하는 도마뱀과와 다르다.

사두골 (포유류의 모루와 동계)은 두개골 뒤쪽에 위치하여 아래턱을 두개골에 연결하고 고막을 잡고 있다.[36] 뼈의 복잡한 해부학[37]은 종 수준에서도 매우 진단적이다.[6] 측면에서 볼 때, 사두골은 미성숙한 ''T. nepaeolicus''와 ''T. proriger'' 개체에서는 갈고리와 유사하지만, 두 종 모두의 성체 형태[29]와 ''T. bernardi''[6][46], ''T. pembinensis''[27], ''T. saskatchweanensis''에서는 튼튼한 타원형 모양을 띤다.[40] 고막 (고실)은 알라 조개라고 불리는 그릇 모양의 우울증 내에서 뼈의 측면 표면에 부착된다.[36] 조개는 ''T. nepaeolicus''[37], ''T. proriger'', ''T. bernardi''[6]에서 얕고, ''T. pembinensis''[37]와 ''T. saskatchewanensis''에서 깊다.[6] 알라 가장자리는 ''T. nepaeolicus'', ''T. proriger''[29], ''T. bernardi''[46][29]에서 얇고, ''T. bernardi'', ''T. pembinensis''[46], ''T. saskatchewanensis''에서 두껍다.[40] 상골돌기는 뼈의 갈고리 모양 확장으로, 줄기 꼭대기에서 후복측으로 구부러져 불완전한 루프를 이루며, 아래턱을 여는 저작근의 부착 지점으로 작용했을 가능성이 높다.[27][37] 이 돌기는 미성숙한 ''T. nepaeolicus''와 ''T. proriger''에서 가늘고 비례적으로 길며, 동물이 성숙해짐에 따라 두꺼워진다.[29] 이 돌기는 ''T. saskatchwanensis''[40]에서 ''T. proriger''와 비슷한 길이를 가지며 ''T. bernardi''에서 짧다.[46] ''T. pembinensis''에서는 45° 하향 각도로 갑자기 안쪽으로 회전한다.[27] 알라 조개의 후복측 가장자리에서 튀어나온 것은 하골돌기이다. 그 모양은 ''T. nepaeolicus''와 ''T. proriger''에서 발생학적으로 변하는 것으로 보이며, 전자의 경우 돌기가 미성숙한 개체에서는 없고 성체에서는 작은 혹으로 나타나지만, ''T. proriger''에서는 어린 개체에서는 미묘한 점으로 존재하고 성체에서는 뚜렷한 넓은 반원으로 변한다.[29] 이 돌기는 ''T. bernardi''[46]에서 작고, ''T. pembinensis''[27]와 ''T. saskatchewanensis''[40]에서는 둥글다. ''T. saskatchewanensis''에서 상골돌기는 하골돌기에 거의 닿는다.[40] 줄기의 바닥에는 하악 과두가 있는데, 이는 사두골과 아래턱 사이의 관절을 형성한다. 성체에서는 둥근 모양이다.[37][46][40][29]
2018년 코니시 타쿠야 등은 두개골 길이가 30cm로 추정되는 작은 유체 화석 FHSM VP-14845를 이빨의 배열, 기초접형골의 비율, 그리고 전상악골의 형태를 근거로 하여 ''틸로사우루스''에 속하는 것으로 분류했다. 하지만, 이 화석은 두개골 길이가 40~60cm인 ''T. nepaeolicus''와 ''T. proriger''의 유체에서 나타나는 다른 ''틸로사우루스''의 특징적인 긴 전상악골 부리가 없다. 이는 ''틸로사우루스''의 부리가 어린 시기에 빠르게 성장하며, 성 선택에 의해 발달하지 않았음을 시사한다. 코니시 등은 현대의 범고래가 사용하는 것처럼 먹이를 들이받는 기능을 제안했다.[31]
틸로사우루스의 두드러진 특징은 원통형의 긴 전상악골(주둥이)이며, 다른 모사사우루스과 파충류와 달리, 틸로사우루스는 전상악골 전방에 이빨이 전혀 없었다.[100] 틸로사우루스는 상악에 24~26개, 구개에 20~22개, 하악에 26개의 이빨이 나 있었다.[84]
3. 3. 턱과 치아
틸로사우루스는 길고 원통 모양인 전상악골(주둥이)이 두드러진 특징이며, 속명은 여기에서 유래되었다. 다른 모사사우루스과 파충류와 달리, 틸로사우루스는 전상악골 앞쪽에 이빨이 전혀 없어서 뼈 돌출부에는 이빨이 존재하지 않았다.[100] 틸로사우루스는 상악에 24~26개, 구개에 20~22개, 하악에 26개의 이빨을 가지고 있었다.[84]3. 4. 골격
1870년대 후반부터 비늘 형태의 ''틸로사우루스'' 피부 화석 증거가 묘사되었다. 이 비늘은 작고 다이아몬드 모양이었으며, 현대의 방울뱀과 다른 관련 파충류에서 발견되는 것과 유사하게 비스듬한 열로 배열되어 있었다. 그러나 모사사우루스의 비늘은 전체 몸에 비해 훨씬 작았다.[53][54] 전체 몸길이가 5m인 개체는 3.3x 크기의 피부 비늘을 가지고 있었고,[55] 모사사우루스의 아랫부분 1제곱인치(2.54 cm)에는 평균 90개의 비늘이 있었다.[53] 각 비늘은 상어의 치아와 유사한 형태로 용골이 있었다.[54] 이것은 수중 저항[54]과 피부 반사를 줄이는 데 도움이 되었을 것이다.[56]Lindgren 등이 2014년에 실시한 ''T. nepaeolicus'' 표본의 비늘에 대한 현미경 분석 결과, 유멜라닌의 높은 흔적이 발견되었으며, 이는 살아있는 바다거북과 유사한 어두운 색상을 나타낸다. 이는 많은 수생 동물에서 나타나는 대응음영으로 보완되었을 수 있지만, 이 종에서 어둡고 밝은 색소의 분포는 알려지지 않았다. 어두운 색상은 여러 진화적 이점을 제공했을 것이다. 어두운 색상은 열 흡수를 증가시켜 동물이 더 추운 환경에서도 높은 체온을 유지할 수 있도록 한다. 유아기에 이러한 특성을 가지면 빠른 성장 속도를 촉진한다. 반사되지 않는 어두운 색상과 대응음영은 모사사우루스에게 향상된 위장을 제공했을 것이다. 추가적인 추정 기능으로는 태양 자외선에 대한 내성 증가, 피부 강화 등이 있다. 이 연구는 특정 흑화증-코딩 유전자가 증가된 공격성에 대해 다면발현적이라고 언급했다.[56]
3. 5. 연조직
1870년대 후반부터 틸로사우루스의 피부 화석이 발견되어 비늘 형태가 묘사되었다. 이 비늘은 작고 다이아몬드 모양이었으며, 방울뱀과 같은 현대 파충류처럼 비스듬한 열로 배열되어 있었다. 하지만 틸로사우루스의 비늘은 몸 전체 크기에 비해 훨씬 작았다.[53][54] 몸길이가 5m인 개체는 3.3x 크기의 피부 비늘을 가지고 있었고,[55] 틸로사우루스 아랫부분 1제곱인치(2.54 cm)에는 평균 90개의 비늘이 있었다.[53] 각 비늘은 상어의 치아와 유사하게 용골이 있었다.[54] 이는 수중 저항[54]과 피부 반사를 줄이는 데 도움이 되었을 것이다.[56]2014년 Lindgren 등의 연구에서는 ''T. nepaeolicus'' 표본의 비늘에서 유멜라닌의 흔적이 발견되어, 살아있는 바다거북처럼 어두운 색상을 띠었을 가능성이 제기되었다. 이는 많은 수생 동물에서 나타나는 대응음영으로 보완되었을 수 있지만, 이 종에서 어둡고 밝은 색소의 분포는 알려지지 않았다. 어두운 색상은 열 흡수를 증가시켜 추운 환경에서도 높은 체온을 유지하도록 돕고, 유아기에 빠른 성장 속도를 촉진했을 수 있다. 또한 반사되지 않는 어두운 색상과 대응음영은 틸로사우루스에게 향상된 위장을 제공했을 것이다. 추가적으로 태양 자외선에 대한 내성 증가, 피부 강화 등의 기능도 추정된다. 이 연구는 특정 흑화증-코딩 유전자가 증가된 공격성에 대해 다면발현적이라고 언급했다.[56]
4. 분류
''틸로사우루스''는 모사사우루스과 틸로사우루스아과에 속하며, 이 아과의 기준 속이다. 틸로사우루스아과는 이빨이 없는 길쭉한 전상악골 주둥이를 가진 것이 특징이다.[6] 틸로사우루스아과의 근연 속으로는 타니와사우루스(''Taniwhasaurus'')와 카이카이필루(''Kaikaifilu'') 등이 있다.[6]
틸로사우루스는 플리오플라테카르푸스아과(Plioplatecarpinae)와 원시 아과인 테티사우루스아과(Tethysaurinae) 및 야구아라사우루스아과(Yaguarasaurinae)와 가까운 친척이다. 이들은 1993년에 처음 인식된 모사사우루스의 세 가지 주요 계통 중 하나에 속하며, 2005년 폴친과 벨에 의해 러셀로사우루스아강(Russellosaurina)으로 명명되었다.[61][62][63]
''틸로사우루스''의 가장 오래된 화석은 텍사스 주립 박물관(TMM) 40092-27에서 채취한 투로니아절(Middle Turonian) 퇴적층인 아르카디아 공원 셰일(Arcadia Park Shale)에서 발견된 전상악골[2]로, 9210만 년에서 9140만 년 사이로 추정된다.[1] 멕시코 치와와 주(Chihuahua, Mexico)의 오히나가 형성(Ojinaga Formation)에서 발견된 두개골(SGM-M1)은 약 9000만 년 전으로 추정된다.[1] 폴란드 나시우프(Nasiłów, Poland)에서 발견된 이빨 화석은 백악기-고생대 경계(Cretaceous–Paleogene boundary)에 가까운 시기로, ''하이나사우루스 sp.''(Hainosaurus sp.)로 추정되었으나,[4][8] ''하이나사우루스''가 ''틸로사우루스''의 동의어로 통합되면서, 이 속은 마지막 모사사우루스 중 하나가 되었다.[46][4]
4. 1. 유효한 종




현재 과학자들이 분류학적으로 유효하다고 인정하는 ''틸로사우루스'' 종은 다음과 같다.
오랜 세월 동안 틸로사우루스의 많은 종이 명명되었지만, 과학자들이 모두 유효하다고 판단하지는 않았다. 에반하트(Everhart)가 2005년에 명명한 ''Tylosaurus kansasensis''[95]는 캔자스 주의 후기 코니아시안에서 산출되었으며, ''T. nepaeolicus''의 어린 개체를 기반으로 하는 것으로 나타났다.[96]
다음 클레이도그램은 고생물학자 코니시 타쿠야와 Michael W. Caldwell에 의한 2011년 계통 분석을 따른다.[99]
{{clade| style=font-size:85%;line-height:80%
|1={{clade
|1=''Clidastes prophyton''
|2={{clade
|1=''Kourisodon puntledgensis''
|label2=러셀로사우리나
|2={{clade
|1={{clade
|1=''Yaguarasaurus columbianus''
|2={{clade
|1=''Russellosaurus coheni''
|2=''Tethysaurus nopcsai'' }} }}
|2={{clade
|1={{clade
|1=''Tylosaurus nepaeolicus''
|2=''Tylosaurus proriger'' }}
|label2='''플리오플라테카르푸스아과'''
|2={{clade
|1=''Ectenosaurus clidastoides''
|2={{clade
|1=''Angolasaurus bocagei''
|2={{clade
|1={{clade
|1=''Selmasaurus johnsoni''
|2=''Selmasaurus russelli'' }}
|2={{clade
|1=''Plesioplatecarpus planifrons''
|2={{clade
|1=''Platecarpus tympaniticus''
|2={{clade
|1=''Latoplatecarpus willistoni''
|2={{clade
|1={{clade
|1=''Latoplatecarpus nichollsae''
|2="''Platecarpus somenensis''" }}
|2={{clade
|1=''Plioplatecarpus primaevus''
|2={{clade
|1=''Plioplatecarpus houzeaui''
|2=''Plioplatecarpus marshi'' }} }} }} }} }} }} }} }} }} }} }} }} }}
5. 고생물학
틸로사우루스는 모사사우루스과에 속하는 해양 파충류로, 고생물학적으로 다양한 특징을 보인다.
이빨틸로사우루스의 두개골에는 전상악골, 상악골, 치골, 익상골 등 다양한 뼈들이 존재하며, 각 뼈에는 날카로운 이빨들이 박혀 있었다. 틸로사우루스는 동형치로, 모든 이빨의 크기와 모양이 거의 동일했다. 다만, 구개에 위치한 익상골 치아는 가장자리 치아보다 작고 더 굽어 있었다.[38]
대부분 종의 가장자리 치열은 큰 해양 척추동물을 절단하는 데 적합했지만,[28][42] ''T. ivoensis''와 ''T. gaudryi''는 먹이를 꿰뚫거나 부수는 데,[5] ''T.'' "borealis"는 꿰뚫는 것과 절단하는 데 모두 최적화된 것으로 보인다.[28]
이빨은 턱 뒤쪽으로 약간 굽은 삼각형 모양이며, 설측 면은 U자형 곡선을 이룬다. 위에서 보면 타원형 모양을 형성한다.[43][40] 용골(절단 모서리)에는 작은 치돌이 있는 미세한 톱니 모양이다.[6][46][43]
Bardet 등(2006)은 가장자리 치열을 기반으로 ''틸로사우루스'' 종을 두 개의 형태학적 그룹으로 분류했다.
- ''proriger'' 그룹: ''T. proriger''와 ''T. nepaeolicus''를 포함하며, 매끄럽거나 희미한 면, 덜 두드러진 용골, 팁 근처까지 뻗어 있는 원시적인 줄무늬의 정맥과 같은 네트워크가 있는 치아가 특징이다.[47]
- ''ivoensis'' 그룹: ''T. ivoensis'', ''T. gaudryi'', ''T. pembinensis''로 구성되며, 튼튼하고 두드러진 용골이 있으며, 설측과 때로는 순측면에 줄무늬가 있고 치아 끝까지 닿지 않으며, 순측면에 면이 있다.[47]
익상골 치아는 하악이 스트렙토스틸 턱 관절을 통해 앞뒤로 미끄러지면서 상악 치아가 먹이를 제자리에 고정하는 래칫 피딩을 가능하게 했을 수 있다.[49]
피부와 색깔틸로사우루스의 피부는 작고 다이아몬드 모양의 비늘로 덮여 있었으며, 비스듬한 열로 배열되어 있었다. 각 비늘에는 용골이 있어 수중 저항과 피부 반사를 줄이는 데 도움이 되었을 것이다.[54][56]
''T. nepaeolicus'' 표본의 비늘에 대한 현미경 분석 결과, 유멜라닌의 흔적이 발견되어 살아있는 바다거북과 유사한 어두운 색상을 띠었을 것으로 추정된다.[56] 어두운 색상은 열 흡수, 위장, 자외선 차단 등 다양한 이점을 제공했을 것이다.[56]
진화2020년, 마지아와 카우는 ''틸로사우루스''의 진화 속도를 조사하는 베이시안 분석을 수행했다. 연구 결과, 틸로사우루스아과는 약 9300만 년 전에 플리오플라테카르푸스아과로부터 분기되었으며, 이는 모든 모사사우루스류 계통에서 가장 높은 진화율을 보였다. 이는 플리오사우루스의 멸종과 폴리코틸리드 다양성의 감소와 일치하며, ''틸로사우루스''가 이들이 남긴 생태적 공백을 빠르게 채웠을 가능성을 시사한다.[64]
신진대사틸로사우루스는 온혈 동물이었을 가능성이 높다. 2016년 연구에서 ''틸로사우루스'' 뼈의 ''δ''18O 동위원소를 조사한 결과, 평균 34.3°C의 내부 체온이 계산되었다. 이는 냉혈 어류나 바다거북보다 높고, 온혈 바닷새와 유사한 수준이었다. 이는 틸로사우루스가 더 넓은 지역을 탐색하고 먹이를 쫓는 데 필요한 지구력 증가, 더 차가운 물에 접근할 수 있는 능력 등 여러 이점을 제공했을 것이다.[65]
운동틸로사우루스는 카랑기폼 운동을 사용했다. 즉, 상체는 덜 유연하고 꼬리를 주로 사용하여 움직였다.[68] 틸로사우루스는 매복 포식자로, 몸 질량과 밀도를 크게 줄이도록 설계된 형태를 가지고 있어 같은 크기의 모사사우루스보다 가벼웠다. 이러한 특성으로 인해 틸로사우루스는 에너지 요구 사항에 더 보수적일 수 있었으며, 공격 시 빠르게 가속하는 데 도움이 되었을 것이다.[41]
주디스 마사레의 1988년 연구에서는 틸로사우루스의 지속적인 수영 속도를 2.32m/s로 계산했지만, 실제 속도는 이보다 절반 정도였을 것으로 예측했다.[69]
식성틸로사우루스는 다양한 해양 동물을 먹이로 삼았던 최상위 포식자였다. 틸로사우루스의 위 내용물 화석에서는 다른 모사사우루스과, 플레시오사우루스, 거북, 헤스페로르니스와 같은 조류, 경골어류, 상어 등 다양한 먹이의 흔적이 발견되었다.[70]

알래스카의 마타누스카 지층에서 발견된 하드로사우루스과 공룡 골격에는 ''T. proriger''의 이빨 자국과 매우 유사한 물린 자국이 남아있었다. 이는 틸로사우루스가 육지 동물의 사체를 먹기도 했음을 시사한다.[77]
사회적 행동틸로사우루스는 동족 간에 공격적인 상호작용을 했을 가능성이 있다. 일부 화석에서는 동족에게 물린 것으로 추정되는 상처가 발견되었는데[21], 이는 영역 다툼이나 짝짓기 경쟁과 관련되었을 수 있다.
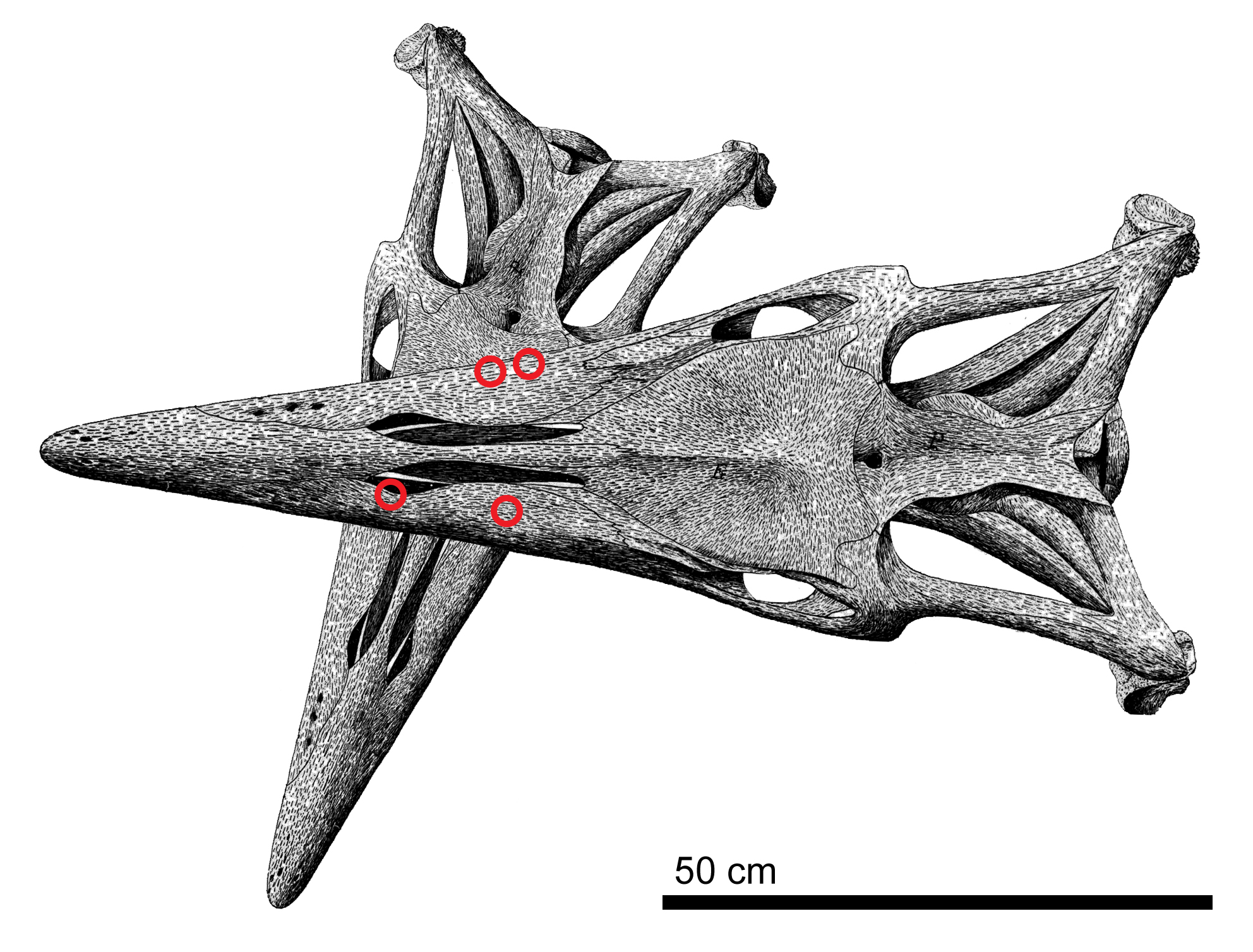
''T. kansasensis''의 모식 표본인 FHSM VP-2295에서는 최소 한 건의 치명적인 동족 간 전투가 기록되었다. 5m 크기의 이 개체는 더 큰 틸로사우루스에게 공격당해 죽은 것으로 보인다. 더 큰 틸로사우루스는 희생자의 머리 왼쪽 아래에서 공격했고, 이 충격으로 희생자의 두개골이 오른쪽으로 굴러가면서 공격자의 이빨이 두개골 지붕과 오른쪽 아래턱에 박혔다. 이로 인해 턱이 부러지고, 익상골 등 주변 뼈가 부러졌으며, 턱이 바깥쪽으로 비틀려 사각골이 분리되고 척수가 끊어져 즉사했을 것으로 추정된다.[21]
병리틸로사우루스 화석에서는 무혈관성 괴사, 꼬리 척추의 비정상적인 융합, 골절 등 다양한 병리학적 증거가 발견되었다. 무혈관성 괴사는 잦은 잠수나 반복적인 잠수와 짧은 호흡 간격으로 인해 발생했을 가능성이 높다.[79] 척추 융합은 외상이나 질병으로 인한 손상 후 뼈가 골화를 거쳐 재형성될 때 발생하며, 포식자의 공격으로 인한 감염에 의해 발생했을 수도 있다.[81]
5. 1. 성장
틸로사우루스는 성장하면서 두개골과 주둥이의 형태가 변하는 특징을 보였다. 어린 틸로사우루스는 주둥이가 짧고 뾰족했지만, 성장하면서 점차 길고 뭉툭해졌다. 이러한 성장 패턴은 성 선택과는 관련이 없는 것으로 보이며, 현생 범고래처럼 먹이를 들이받는 기능을 수행하기 위해 발달했을 가능성이 제기된다.[100]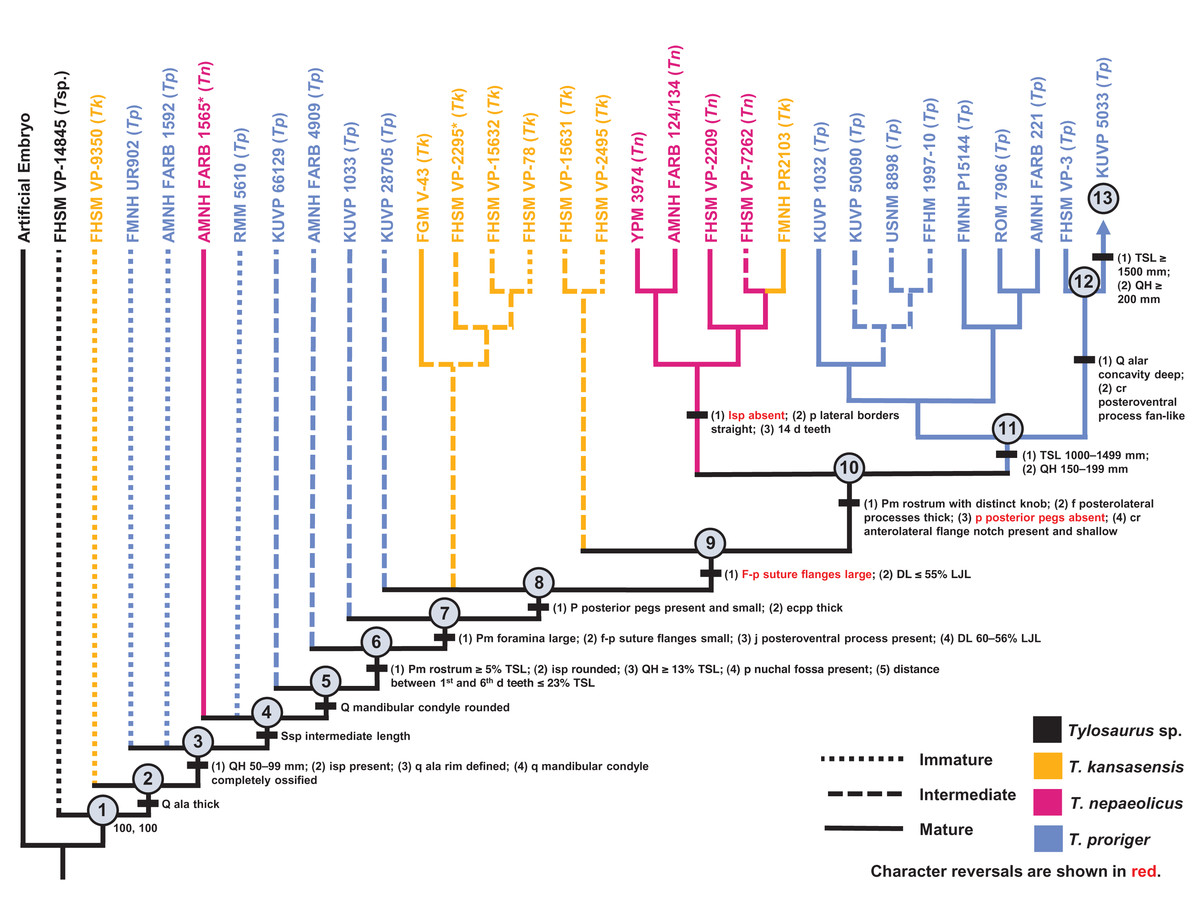
서부 내해에서 발견되는 ''T. nepaeolicus''와 ''T. proriger''는 시간종을 나타낼 수 있다. 이들은 성장발생이라는 과정을 통해 분기 없이 지속적으로 진화하는 단일 계통을 구성한다. 두 종은 층서적으로 겹치지 않고, 자매군이며, 사골의 점진적인 변화와 같은 작고 중간 정도의 형태학적 차이를 공유하며, 같은 지역에 서식했다.[33][29]
이 계통의 진화 방식은 발생학의 변화와 관련된 두 가지 진화 메커니즘 중 하나를 통해 이루어졌을 것으로 추정된다.
- 소아형태 발달: ''T. proriger''가 ''T. nepaeolicus''의 소아형태 발달로 진화했을 가능성이 있다. 즉, 후손이 조상의 어린 시절 특징을 성체까지 유지하면서 형태학적 변화가 나타났다는 것이다. 이는 어린 ''T. nepaeolicus''와 모든 ''T. proriger''에서 공유되지만 성체 ''T. nepaeolicus''에서는 사라지는 두개골의 전두엽 능선과 두정골의 볼록한 경계의 존재에 기반을 둔다.[43][29]
- 페라모르포시스: 페라모르포시스 가설에 따르면, ''T. proriger''가 미성숙한 상태에서 성숙한 ''T. nepaeolicus''에서 발견되는 형질을 발달시켜 진화했을 수 있다. 74개의 ''틸로사우루스'' 표본 데이터를 사용하여 개발된 분류학적 온토그램 결과에 따르면, 모든 ''T. proriger''와 성숙한 ''T. nepaeolicus''에 존재하지만 어린 ''T. nepaeolicus''에는 없는 수많은 형질이 확인되었다. 여기에는 두개골 크기와 깊이가 크고, 길쭉한 콧대의 길이가 총 두개골 길이의 5%를 초과하고, 사골 상부 봉합 돌기가 두껍고, 전반적인 사골 모양이 수렴하며, 후복측 돌기가 부채꼴인 점이 포함된다.[29]
2018년, 코니시 타쿠야 연구진은 두개골 길이가 30cm로 추정되는 작은 유체 화석 FHSM VP-14845를 틸로사우루스로 분류했다. 이 화석은 이빨의 배열, 기초접형골의 비율, 전상악골의 형태를 근거로 틸로사우루스에 속하는 것으로 판단되었다. 하지만, 이 화석은 두개골 길이가 40cm에서 60cm인 ''T. nepaeolicus''와 ''T. proriger''의 유체에서 나타나는 틸로사우루스의 특징적인 긴 전상악골 부리가 없다. 이는 틸로사우루스의 부리가 어린 시기에 빠르게 성장하며, 성 선택에 의해 발달하지 않았음을 시사한다.[31]
5. 2. 신진대사
거의 모든 유린목은 냉혈성 외온 동물적 신진대사를 특징으로 하지만, ''틸로사우루스''와 같은 모사사우루스과는 온혈, 즉 따뜻한 피를 가졌다는 점에서 독특하다.[65] 이러한 특징을 가진 다른 알려진 도마뱀은 아르헨티나 흑백 테구뿐이지만 부분적이다.[66] 2016년 Harrell, Pérez‐Huerta, 및 Suarez의 연구에서 ''틸로사우루스'' 뼈의 ''δ''18O 동위원소를 조사하여 ''틸로사우루스''의 내온성을 입증했다. ''δ''18O 수준은 동물의 내부 체온을 계산하는 데 사용할 수 있으며, 공존하는 냉혈 동물과 온혈 동물 간의 계산된 체온을 비교하여 신진대사 유형을 추론할 수 있다. 이 연구는 무어빌 백악에서 냉혈 어류 ''엔코두스''와 바다거북 ''톡소켈리스''(해양 온도와 상관관계)와 온혈 바닷새 ''이크티오르니스''의 체온을 대용물로 사용했다. 11개의 ''틸로사우루스'' 표본의 동위원소 수준을 분석한 결과 34.3°C의 평균 내부 체온이 계산되었다. 이는 ''엔코두스''와 ''톡소켈리스''(각각 28.3°C 및 27.2°C)의 체온보다 훨씬 높았으며, ''이크티오르니스''(38.6°C)의 체온과 유사했다. Harrell, Pérez‐Huerta, 및 Suarez는 또한 ''플라테카르푸스''와 ''클리다스테스''의 체온을 각각 36.3°C 및 33.1°C로 계산했다. 다른 모사사우루스가 ''틸로사우루스''보다 훨씬 작았음에도 불구하고 유사한 체온을 유지했다는 사실은 ''틸로사우루스''의 체온이 거대온혈과 같은 다른 신진대사 유형의 결과일 가능성이 낮다는 것을 보여주었다. 내온성은 ''틸로사우루스''에게 더 넓은 지역을 탐색하고 먹이를 쫓는 데 필요한 지구력 증가, 더 차가운 물에 접근할 수 있는 능력, 후기 백악기 동안의 지구 온도 점진적 냉각에 대한 더 나은 적응과 같은 몇 가지 이점을 제공했을 것이다.[65]5. 3. 운동
과거에는 틸로사우루스가 뱀장어처럼 몸 전체를 파동시켜 움직이는 수영을 하는 것으로 해석되었다. 하지만 현재는 틸로사우루스가 카랑기폼 운동을 사용했다는 것이 밝혀졌는데, 이는 상체가 덜 유연하고 고등어와 같이 꼬리를 주로 사용하여 움직였다는 것을 의미한다.[68] 2017년에 발표된 한 연구에서 ''T. proriger'' 척추의 이동성을 조사한 결과, 등쪽 척추는 비교적 뻣뻣했지만 목, 꼬리뼈, 꼬리 척추는 움직임이 더 자유로웠다. 이는 엉덩이까지 척추가 뻣뻣했던 ''플로토사우루스''와 같은 더 진화된 모사사우루스와 대조된다. 어린 ''T. proriger''을 검사한 결과 목과 등쪽 척추가 성체보다 훨씬 뻣뻣했는데, 이는 어린 개체들이 포식자를 피하거나 먹이를 잡는 데 더 유리했을 수 있다. 나이가 많은 개체는 생존에 포식자 회피가 덜 중요해짐에 따라 척추 유연성이 커진다.[68]틸로사우루스는 매복 포식자로 전문화되었을 가능성이 높다. 몸 질량과 밀도를 크게 줄이도록 설계된 형태를 가지고 있어 같은 크기의 모사사우루스보다 가볍다. 무게와 관련된 가슴 및 골반, 노는 비례적으로 작다. 뼈는 매우 해면골이었으며, 살아있을 때는 지방 세포로 채워져 부력을 증가시켰을 가능성이 높다. 이러한 특성으로 인해 틸로사우루스는 에너지 요구 사항에 더 보수적일 수 있었으며, 이는 매복 지점 사이를 장거리 또는 은밀하게 이동할 때 유용했을 것이다. 또한 신체 밀도 감소는 모사사우루스의 길고 강력한 꼬리와 함께 공격 시 틸로사우루스가 빠르게 가속하는 데 도움이 되었을 것이다.[41]
주디스 마사레의 1988년 연구에서는 수학적 모델을 통해 틸로사우루스의 지속적인 수영 속도(동물이 지치지 않고 움직이는 속도)를 계산했다. 6.46m 길이와 6.32m 길이의 두 ''T. proriger'' 표본을 사용하여 일관된 평균 최대 지속 수영 속도를 2.32m/s로 계산했다. 그러나 모델이 과장된 것으로 나타났는데, 이는 항력에 대한 변수가 과소평가되었을 수 있기 때문이었다. 마사레는 틸로사우루스의 실제 지속 수영 속도가 계산된 속도의 절반 정도일 것으로 예측했다.[69]
5. 4. 식성
틸로사우루스는 당시 가장 큰 해양 육식 동물 중 하나로, 다양한 해양 동물을 먹이로 삼았던 최상위 포식자였다. 틸로사우루스의 위 내용물 화석에서는 다른 모사사우루스과, 플레시오사우루스, 거북, 헤스페로르니스와 같은 조류, 경골어류, 상어 등 다양한 먹이의 흔적이 발견되었다.[70]1987년 사우스다코타의 피에르 셰일에서 발견된 모사사우루스과, 헤스페로르니스, 바나노그미우스, 상어의 화석이 단일 ''T. proriger'' 골격(SDSM 10439)의 위에서 발견된 것은 틸로사우루스의 다양한 식성을 보여주는 대표적인 사례이다.[41][70][75] 이 외에도 ''T. bernardi''와 유사한 종에서 발견된 바다거북,[70][41] 돌리코린코프스가 다른 ''T. proriger''에서 발견되었으며,[22] ''T. proriger''에서 부분적으로 소화된 키몰리크티스 뼈와 비늘, 클리다스테스의 척추뼈, ''T. nepaeolicus''에서 세 개의 플라테카르푸스 잔해, ''T. saskatchewanensis''에서 플리오플라테카르푸스 뼈가 발견되었다.[21][24][76]
암모나이트 화석의 구멍 자국,[72] 프로토스테가의 등딱지, 엔코테우티스의 검에서도 틸로사우루스에 의한 것으로 추정되는 흔적이 발견되었다.[71]
알래스카의 마타누스카 지층에서 발견된 하드로사우루스과 공룡 골격에는 ''T. proriger''의 이빨 자국과 매우 유사한 물린 자국이 남아있었다. 이는 틸로사우루스가 육지 동물의 사체를 먹기도 했음을 시사한다. 해당 공룡은 해안에서 표류하다가 틸로사우루스에게 뜯어먹혔을 것으로 추정된다.[77]
5. 5. 사회적 행동
틸로사우루스는 동족 간에 공격적인 상호작용을 했을 가능성이 있다. 일부 화석에서는 동족에게 물린 것으로 추정되는 상처가 발견되었는데[21], 이는 영역 다툼이나 짝짓기 경쟁과 관련되었을 수 있다.19세기 말과 20세기 초 화석 사냥꾼들은 틸로사우루스 화석에서 동족에게 공격당한 흔적을 자주 보고했지만, 과학적으로 수집된 표본은 드물다. 이 화석들은 대부분 머리 부위 주변에 집중된 치유된 물린 자국과 상처로 구성되어 있어, 치명적이지 않은 상호작용이었음을 시사한다. 하지만 이러한 접촉의 동기는 불분명하다.[21] 1993년 로스차일드와 마틴은 일부 현대 도마뱀이 구애 과정에서 짝의 머리를 물어 상처를 입히거나, 수컷 도마뱀들이 머리를 잡고 상대를 뒤집는 영토 행동을 보이는 것을 관찰했다. 틸로사우루스도 이와 유사한 행동을 했을 가능성이 있다.[21]
링검-솔리어는 틸로사우루스의 전투적인 길쭉한 부리가 사냥뿐만 아니라 수컷 간의 짝짓기 경쟁에도 사용되었을 수 있다고 제안했다.[41] 그러나 길쭉한 부리가 성별에 관계없이 모든 개체에서 발견된다는 점을 고려할 때,[41] 코니시 외(2018)와 지에트로우(2020)의 연구는[31][29] 성 선택이 아닌 다른 요인에 의해 부리가 발달했음을 시사한다.[31]
''T. kansasensis''의 모식 표본인 FHSM VP-2295에서는 최소 한 건의 치명적인 동족 간 전투가 기록되었다. 5m 크기의 이 개체는 더 큰 틸로사우루스에게 공격당해 죽은 것으로 보인다. 두개골 지붕과 주변에는 네 개의 큰 홈과 같은 외상 징후가 있고, 치골에는 최소 일곱 개의 찔린 상처와 홈이 있다. 이 상처들은 약 7m 크기의 더 큰 틸로사우루스의 물린 자국과 일치한다. 가장 큰 자국은 길이가 약 4cm이며, 큰 모사사우루스 이빨 크기와 턱이 수렴하는 각도(약 30°)와도 일치한다. 또한 FHSM VP-2295는 목에 손상을 입었는데, 경추가 두개골과 부자연스러운 각도(40°)로 연결되어 있었다. 이는 척추가 훼손된 것이 아니라, 생전에 목이 심하게 비틀렸음을 나타낸다. 더 큰 틸로사우루스는 희생자의 머리 왼쪽 아래에서 공격했고, 이 충격으로 희생자의 두개골이 오른쪽으로 굴러가면서 공격자의 이빨이 두개골 지붕과 오른쪽 아래턱에 박혔다. 이로 인해 턱이 부러지고, 익상골 등 주변 뼈가 부러졌으며, 턱이 바깥쪽으로 비틀려 사각골이 분리되고 척수가 끊어져 즉사했을 것으로 추정된다.[21]
5. 6. 병리
틸로사우루스 화석에서는 다양한 병리학적 증거가 발견되었다. 로스차일드와 마틴(2005)은 12구의 북미산 틸로사우루스 골격과 1구의 *T. bernardi* 골격을 조사한 결과, 모든 개체에서 무혈관성 괴사의 증거를 확인했다.[79] 무혈관성 괴사는 잦은 잠수나 반복적인 잠수와 짧은 호흡 간격으로 인해 뼈를 손상시키는 질소 기포가 축적되어 발생하는 감압병의 결과일 가능성이 높다. 연구된 틸로사우루스는 이러한 행동을 통해 무혈관성 괴사를 얻었을 가능성이 높으며, 틸로사우루스에서 변함없이 나타나는 것을 고려할 때, 깊은 잠수 또는 반복적인 잠수는 이 속의 일반적인 행동 특성이었을 가능성이 높다. 북미산 틸로사우루스의 척추 중 3~15%, *T. bernardi*의 척추 중 16%가 무혈관성 괴사의 영향을 받았다.[79]일부 틸로사우루스 골격에서는 꼬리 척추의 비정상적인 융합이 보고되었다. 이러한 융합은 꼬리 끝 근처에 집중되어 여러 융합된 척추의 단일 덩어리인 "클럽 테일"을 형성할 수 있다. 로스차일드와 에버하트(2015)는 23구의 북미산 틸로사우루스 골격과 1구의 *T. bernardi* 골격을 조사하여 5구의 북미 골격에서 융합된 꼬리 척추를 발견했다.[81] 척추 융합은 외상이나 질병으로 인한 손상 후 뼈가 골화를 거쳐 재형성될 때 발생한다. 클럽 테일 상태의 한 어린 개체에서 융합 부위에 상어 이빨이 박혀 있는 것이 발견되었는데, 이는 적어도 일부 사례가 포식자의 공격으로 인한 감염에 의해 발생했음을 확인해준다.[81] 틸로사우루스의 척추 융합 사례 대부분은 뼈 감염으로 인해 발생했지만, 일부 사례는 척추관절병증과 같은 관절 질환으로 인해 발생했을 수도 있다.[81]
다른 신체 부위의 골절에서도 재형성된 뼈의 축적이 나타난다. 한 *T. kansasensis* 표본은 완전히 치유된 두 개의 갈비뼈 골절을 가지고 있다.[82] 또 다른 *T. proriger* 두개골은 바위와 같은 단단한 물체에 부딪혀 발생한 것으로 보이는 부리 골절을 보여준다.[82]
참조
[1]
간행물
The Geologic Time Scale
Elsevier
[2]
학술지
The oldest North American mosasaurs (Squamata: Mosasauridae) from the Turonian (Upper Cretaceous) of Kansas and Texas with comments on the radiation of major mosasaur clades
https://www.research[...]
[3]
학술지
Occurrence of a tylosaurine mosasaur (Mosasauridae; Russellosaurina) from the Turonian of Chihuahua State, Mexico
http://boletinsgm.ig[...]
[4]
학술지
New records of the tylosaurine mosasaur ''Hainosaurus'' from the Campanian-Maastrichtian (Late Cretaceous) of central Poland
[5]
학술지
Tylosaurine mosasaurs (Squamata) from the Late Cretaceous of northern Germany
http://nbn-resolving[...]
[6]
학술지
A New Hypothesis of the Phylogenetic Relationships of the Tylosaurinae (Squamata: Mosasauroidea)
[7]
학술지
The occurrence and geological setting of Cretaceous dinosaurs, mosasaurs, plesiosaurs, and turtles from Angola
https://docentes.fct[...]
[8]
학술지
The late Maastrichtian Belemnella kazimiroviensis group (Cephalopoda, Coleoidea) in the Middle Vistula valley (Poland) and the Maastricht area (the Netherlands, Belgium) – taxonomy and palaeobiological implications
[9]
학술지
'[Remarks on ''Holops brevispinus'', ''Ornithotarsus immanis'', and ''Macrosaurus proriger'']'
https://www.biodiver[...]
[10]
웹사이트
Macrosaurus proriger
http://oceansofkansa[...]
[11]
학술지
Synopsis of the extinct Batrachia, Reptilia and Aves of North America
https://www.biodiver[...]
[12]
웹사이트
Sebastes proriger, Redstripe rockfish
https://www.fishbase[...]
[13]
학술지
On the structure of the skull and limbs in mosasaurid reptiles, with descriptions of new genera and species
https://archive.org/[...]
[14]
웹사이트
Tylosaurus proriger
http://oceansofkansa[...]
[15]
학술지
Note on ''Rhinosaurus''
https://books.google[...]
[16]
학술지
New Data on Cranial Measurements and Body Length of the Mosasaur, ''Tylosaurus nepaeolicus'' (Squamata; Mosasauridae), from the Niobrara Formation of Western Kansas
https://www.research[...]
[17]
서적
Contributions to the extinct vertebrate fauna of the western interior territories: Report of the United States Geological Survey of the Territories
https://www.biodiver[...]
U.S. Government Printing Office
[18]
학술지
Über die Pythonomorphen der Kansas-Kreide
https://www.biodiver[...]
[19]
학회자료
Mosasaurs—how large did they really get?
https://www.research[...]
5th Triennial Mosasaur Meeting—A Global Perspective on Mesozoic Marine Amniotes
2016
[20]
학술지
Rapid evolution, diversification and distribution of mosasaurs (Reptilia; Squamata) prior to the KT Boundary
https://www.research[...]
[21]
학술지
A bitten skull of ''Tylosaurus kansasensis'' (Squamata: Mosasauridae) and a review of mosasaur-on-mosasaur pathology in the fossil record
https://www.research[...]
[22]
학술지
Plesiosaurs as the food of mosasaurs; new data on the stomach contents of a ''Tylosaurus proriger'' (Squamata; Mosasauridae) from the Niobrara Formation of western Kansas
https://www.research[...]
[23]
학술지
The first record of ''Hainosaurus'' (Reptilia: Mosasauridae) from Sweden
[24]
웹사이트
Omācīw
https://royalsaskmus[...]
[25]
웹사이트
Largest mosasaur on display
https://www.guinness[...]
2020-06-03
[26]
뉴스
Manitoba dig uncovers 80-million-year-old sea creature
https://www.cbc.ca/n[...]
2008-08-27
[27]
학술지
Redescription and rediagnosis of the tylosaurine mosasaur ''Hainosaurus pembinensis'' Nicholls, 1988, as ''Tylosaurus pembinensis'' (Nicholls, 1988)
[28]
학위논문
A new high-latitude ''Tylosaurus'' (Squamata, Mosasauridae) from Canada with unique dentition
https://etd.ohiolink[...]
University of Alberta
[29]
학술지
Craniofacial ontogeny in Tylosaurinae
[30]
학술지
A new species of gigantic mosasaur from the Late Cretaceous of Israel
[31]
학술지
The Smallest-Known Neonate Individual of ''Tylosaurus'' (Mosasauridae, Tylosaurinae) Sheds New Light on the Tylosaurine Rostrum and Heterochrony
[32]
학술지
A rostral neurovascular system in the mosasaur Taniwhasaurus antarcticus
https://doi.org/10.1[...]
2020-04-24
[33]
학술지
''Tylosaurus kansasensis'', a new species of tylosaurine (Squamata, Mosasauridae) from the Niobrara Chalk of western Kansas, USA
https://www.research[...]
2005
[34]
학술지
California mosasaurs
https://catalog.hath[...]
[35]
학술지
Did mosasaurs have forked tongues?
[36]
논문
An halisaurine (Squamata: Mosasauridae) from the Late Cretaceous of Patagonia, with a preserved tympanic disc: Insights into the mosasaur middle ear
[37]
논문
A comprehensive review of the morphological diversity of the quadrate bone in mosasauroids (Squamata: Mosasauroidea), with comments on the homology of the infrastapedial process
[38]
논문
"''Tylosaurus ivoensis'': a giant mosasaur from the early Campanian of Sweden"
[39]
논문
The first record of the mosasaur ''Hainosaurus'' (Reptilia: Lacertilia) from North America
[40]
논문
A new species of tylosaurine mosasaur from the upper Campanian Bearpaw Formation of Saskatchewan, Canada
[41]
논문
The Tylosaurine Mosasaurs (Reptilia, Mosasauridae) from the Upper Cretaceous of Europe and Africa
https://core.ac.uk/r[...]
2020-12-24
[42]
논문
Charting the Late Cretaceous seas: mosasaur richness and morphological diversification
https://digitalcommo[...]
[43]
논문
Re-characterization of ''Tylosaurus nepaeolicus'' (Cope, 1874) and ''Tylosaurus kansasensis'' Everhart, 2005: Ontogeny or sympatry?
[44]
논문
Allometric growth in the skull of ''Tylosaurus proriger'' (Squamata: Mosasauridae) and its taxonomic implications
[45]
논문
Ecological and evolutionary implications of ontogenetic changes in the marginal dentition of ''Tylosaurus proriger'' (Squamata: Mosasauridae)
https://vertpaleo.or[...]
[46]
논문
Reassessment and reassignment of the early Maastrichtian mosasaur ''Hainosaurus bernardi'' Dollo, 1885, to ''Tylosaurus'' Marsh, 1872
[47]
논문
A Tylosaurine Mosasauridae (Squamata) from the Late Cretaceous of the Basque-Cantabrian Region
https://estudiosgeol[...]
[48]
논문
A new species of ''Taniwhasaurus'' (Mosasauridae, Tylosaurinae) from the upper Santonian-lower Campanian (Upper Cretaceous) of Hokkaido, Japan
[49]
논문
Anatomy and functional morphology of the largest marine reptile known, ''Mosasaurus hoffmanni'' (Mosasauridae, Reptilia) from the Upper Cretaceous, Upper Maastrichtian of The Netherlands
https://royalsociety[...]
[50]
논문
New exceptional specimens of Prognathodon overtoni (Squamata, Mosasauridae) from the upper Campanian of Alberta, Canada, and the systematics and ecology of the genus
http://www.tandfonli[...]
2011-09
[51]
논문
Ontogeny and phylogeny of the mesopodial skeleton in mosasauroid reptiles
https://academic.oup[...]
1996-04
[52]
논문
Soft tissue preservation in a fossil marine lizard with a bilobed tail fin
https://www.nature.c[...]
2013-09-10
[53]
논문
On the dermal covering of a mosasauroid reptile
http://oceansofkansa[...]
[54]
논문
Soft-tissue preservation in a 95 million year old marine lizard: form, function, and aquatic adaptation
[55]
논문
Three-Dimensionally Preserved Integument Reveals Hydrodynamic Adaptations in the Extinct Marine Lizard ''Ectenosaurus'' (Reptilia, Mosasauridae)
[56]
논문
Skin pigmentation provides evidence of convergent melanism in extinct marine reptiles
[57]
논문
A complete mosasaur skeleton, osseous and cartilaginous
http://digitallibrar[...]
[58]
논문
Convergent Evolution in Aquatic Tetrapods: Insights from an Exceptional Fossil Mosasaur
[59]
논문
A small, exquisitely preserved specimen of ''Mosasaurus missouriensis'' (Squamata, Mosasauridae) from the upper Campanian of the Bearpaw Formation, western Canada, and the first stomach contents for the genus
[60]
논문
"''Platecarpus tympaniticus'' (Squamata, Mosasauridae): osteology of an exceptionally preserved specimen and its insights into the acquisition of a streamlined body shape in mosasaurs"
[61]
논문
"''Dallasaurus turneri'', a new primitive mosasauroid from the Middle Turonian of Texas and comments on the phylogeny of the Mosasauridae (Squamata)"
https://www.research[...]
[62]
논문
A challenge to categories: "What, if anything, is a mosasaur?"
[63]
논문
Inferring 'weak spots' in phylogenetic trees: application to mosasauroid nomenclature
[64]
논문
Estimating the evolutionary rates in mosasauroids and plesiosaurs: discussion of niche occupation in Late Cretaceous seas
[65]
논문
Endothermic mosasaurs? Possible thermoregulation of Late Cretaceous mosasaurs (Reptilia, Squamata) indicated by stable oxygen isotopes in fossil bioapatite in comparison with coeval marine fish and pelagic seabirds
[66]
논문
Seasonal reproductive endothermy in tegu lizards
[67]
학위논문
Osteohistology And Skeletochronology Of An Ontogenetic Series Of ''Clidastes'' (Squamata: Mosasauridae): Growth And Metabolism In Basal Mosasaurids
https://scholars.fhs[...]
Fort Hays State University
[68]
학위논문
Locomotion and skeletal morphology of Late Cretaceous mosasaur, Tylosaurus proriger
https://digitalcommo[...]
Georgia Southern University
[69]
논문
Swimming capabilities of Mesozoic marine reptiles: Implications for method of predation
[70]
논문
A small, exquisitely preserved specimen of ''Mosasaurus missouriensis'' (Squamata, Mosasauridae) from the upper Campanian of the Bearpaw Formation, western Canada, and the first stomach contents for the genus
[71]
간행물
Enchoteuthididae, giant squids from the Upper Cretaceous of the Western Interior
https://www.research[...]
[72]
논문
Mosasaur Predation on Upper Cretaceous Nautiloids and Ammonites from the United States Pacific Coast
http://doc.rero.ch/r[...]
[73]
논문
An Elasmosaur with Stomach Contents and Gastroliths from the Pierre Shale (Late Cretaceous) of Kansas
[74]
웹사이트
Museum of Geology, South Dakota School of Mines
http://oceansofkansa[...]
[75]
논문
Gastric residues associated with a mosasaur from the Late Cretaceous (Campanian) Pierre Shale in South Dakota.
1987
[76]
Tweet
A map showing the position of Omācīw, the Tylosaurus skeleton, as it was found in the quarry near Lake Diefenbaker in 1995. Its bones were arranged almost as they were in life.
2020-03-06
[77]
서적
Mesozoic Vertebrate Life
Indiana University Press
2001
[78]
논문
Arctic mosasaurs (Squamata, Mosasauridae) from the Upper Cretaceous of Russia
https://www.research[...]
[79]
논문
Mosasaur ascending: the phytogeny of bends
[80]
논문
Frequency of decompression illness among recent and extinct mammals and "reptiles": a review
[81]
논문
Co-Ossification of Vertebrae in Mosasaurs (Squamata, Mosasauridae); Evidence of Habitat Interactions and Susceptibility to Bone Disease
[82]
웹사이트
Mosasaur Pathology
http://oceansofkansa[...]
[83]
문서
A new species of tylosaurine mosasaur from the upper Campanian Bearpaw Formation of Saskatchewan, Canada
2018
[84]
논문
Reassessment and reassignment of the early Maastrichtian mosasaur ''Hainosaurus bernardi'' Dollo, 1885, to ''Tylosaurus'' Marsh, 1872
[85]
웹사이트
H.F. Osborn 1899
http://www.oceansofk[...]
2008-04-18
[86]
웹사이트
Mosasaur Fringe
http://www.oceansofk[...]
2013-01-13
[87]
논문
The first record of ''Hainosaurus'' (Reptilia, Mosasauridae) from Sweden
http://jpaleontol.ge[...]
2005
[88]
웹사이트
Fact File: Tylosaurus Proriger from National Geographic
http://animals.natio[...]
2010-05-12
[89]
문서
Remarks on ''Macrosaurus proriger''.
1869
[90]
문서
Note on ''Rhinosaurus''.
1872
[91]
웹사이트
Tylosaur food
http://www.oceansofk[...]
Oceansofkansas.com
2013-01-28
[92]
웹사이트
Museum Of World Treasures
http://worldtreasure[...]
Worldtreasures.org
2013-01-28
[93]
웹사이트
'[dinosaur] Tylosaurus saskatchewanensis (new species) + African araripemydid turtle Taquetochelys decorata'
http://dml.cmnh.org/[...]
2019-02-04
[94]
문서
Review of the vertebrata of the Cretaceous period found west of the Mississippi River.
1874
[95]
논문
''Tylosaurus kansasensis'', a new species of tylosaurine (Squamata, Mosasauridae) from the Niobrara Chalk of western Kansas, USA
https://www.research[...]
2005
[96]
문서
Re-characterization of ''Tylosaurus nepaeolicus'' (Cope, 1874) and ''Tylosaurus kansasensis'' Everhart, 2005: Ontogeny or sympatry?
2016
[97]
문서
Mosasaurs.
1898
[98]
문서
A phylogenetic revision of North American and Adriatic Mosasauroidea.
1997
[99]
논문
Two new plioplatecarpine (Squamata, Mosasauridae) genera from the Upper Cretaceous of North America, and a global phylogenetic analysis of plioplatecarpines
[100]
논문
The Smallest-Known Neonate Individual of ''Tylosaurus'' (Mosasauridae, Tylosaurinae) Sheds New Light on the Tylosaurine Rostrum and Heterochrony
[101]
웹사이트
Mosasaurs ate plesiosaurs: New data on the gut contents of a Tylosaurus proriger (Squamata; Mosasauridae) from the Smoky Hill Chalk of western Kansas
http://oceansofkansa[...]
2019-10-20
[102]
서적
Sea Monsters Creatures of the Deep
https://books.google[...]
Mike Everhart
[103]
서적
Mesozoic Vertebrate Life
Indiana University Press
2001
[104]
서적
The Princeton Field Guide to Mesozoic Sea Reptiles
Princeton University Press
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com