모사사우루스
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
모사사우루스는 백악기 후기에 번성했던 멸종된 해양 파충류의 한 속으로, 1764년 네덜란드 마스트리흐트 근처에서 두개골 화석이 처음 발견되었다. 모사사우루스는 유선형 몸체, 지느러미, 아래로 굽은 꼬리를 가지고 있었으며, 최대 17m 이상까지 성장하는 대형 해양 포식자였다. 이들은 다양한 해양 환경에 서식하며 어류, 상어, 두족류, 다른 해양 파충류 등을 먹이로 삼았다. 모사사우루스는 턱의 강력한 물기 힘과 시각 능력을 활용하여 사냥했으며, 내온성이었을 가능성이 높다. 모사사우루스는 백악기-고생대 대량 절멸 사건으로 멸종되었다.
더 읽어볼만한 페이지
- 모사사우루스과 - 틸로사우루스
틸로사우루스는 백악기 후기에 서식한, 길쭉한 주둥이를 가진 최대 14m에 달하는 모사사우루스과의 해양 파충류로, 북아메리카와 유럽 등지에서 화석이 발견되며 해양 생물을 포식하는 최상위 포식자였다. - 모사사우루스과 - 프로그나토돈
프로그나토돈은 백악기 후기에 살았던 모사사우루스과의 멸종된 해양 파충류로, 강력한 턱과 튼튼한 이빨을 가졌으며, 앨버타의 베어포 지층 등에서 화석이 발견되었다. - 1822년 기재된 화석 분류군 - 렙토넥테스
렙토넥테스는 트라이아스기 후기부터 쥐라기 초기에 살았던 어룡의 한 속으로, 유럽에서 화석이 발견되었으며 긴 꼬리와 뾰족한 주둥이를 특징으로 한다. - 1822년 기재된 화석 분류군 - 키프로스난쟁이하마
키프로스난쟁이하마는 플라이스토세 후기 키프로스 섬에 서식했던 멸종된 하마의 일종으로, 섬 왜소화의 대표적인 사례이며 키프로스왜소코끼리와 함께 섬의 유일한 대형 포유류였으나 인간의 활동으로 멸종된 것으로 추정된다.

2. 연구사
모사사우루스 연구는 18세기 후반 네덜란드 마스트리흐트 근처에서 발견된 두개골 화석으로부터 시작되었다. 1764년 처음 발견된 두개골(TM 7424)은 처음에 고래의 것으로 오인되었으나,[7] 현재는 하를럼의 테일러스 박물관에 보관되어 있다.[8][16][13]
1780년경 발견된 두 번째 두개골은 요한 레오나르트 호프만에 의해 악어로 여겨졌으나, 페트루스 캄퍼의 연구를 통해 국제적인 주목을 받았다.[16][10][11] 프랑스 혁명 당시 프랑스 혁명가들은 이 화석을 약탈하여 1795년 프랑스 국립 자연사 박물관으로 옮겼다(MNHN AC 9648).[11][13] 이후 1800년 아드리안 질레스 캄퍼는 이 화석이 모니터 도마뱀과 유사하지만 다른 현대 동물과는 다른 해양 파충류에 속한다고 결론지었고,[14] 1808년 조르주 퀴비에는 이를 확인했다.[15][8][16]
이 두개골은 퀴비에가 멸종 개념을 추측하고, 격변설과 진화 이론의 전조가 되는 데 중요한 역할을 했다.[17] 1822년 윌리엄 다니엘 코니베어는 뫼즈강의 라틴어 이름 'Mosa'와 도마뱀을 뜻하는 고대 그리스어 '사우로스'를 합쳐 '모사사우루스'라는 속명을 만들었다.[18][8][58] 1829년 기드온 맨텔은 호프만을 기리기 위해 '호프만니(hoffmannii)'라는 종소명을 추가했다.[19]
1834년 리처드 할란은 미주리 강 근처에서 발견된 주둥이 조각을 바탕으로 'M. 미주리엔시스(M. missouriensis)'를 처음 기술했다.[21] 이후 여러 학자들이 다양한 지역에서 발견된 화석을 바탕으로 새로운 종을 기술했다.
초기에는 모사사우루스가 물갈퀴가 달린 발과 육상 보행이 가능한 사지를 가진 양서류로 묘사되었으나, 1854년 헤르만 슐레겔이 완전한 수생 지느러미를 가졌음을 증명했다.[40][41] 슐레겔의 가설은 처음에는 무시되었지만, 1870년대 오스니얼 찰스 마시와 에드워드 드링커 코프가 북아메리카에서 더 완전한 유해를 발견하면서 널리 받아들여졌다.[16][41]
벤자민 워터하우스 호킨스는 1854년 런던 수정궁 공원에 전시된 선사 시대 동물 조각 컬렉션의 일부로 모사사우루스의 실물 크기 콘크리트 조각을 제작했다.[42] 이 복원은 리처드 오웬의 해석과 왕도마뱀 해부학을 바탕으로 했으나, 당시 기준으로도 부정확한 부분이 있었다.[42]
2. 1. 발견
과학계에 최초로 알려진 모사사우루스 화석은 1764년 네덜란드 마스트리흐트 근처 백악 채석장에서 발견된 두개골(TM 7424)이다. 이 화석은 처음에는 고래로 오인되었다.[7] 이 표본은 현재 하를럼의 테일러스 박물관에 전시되어 있다.[8][16][13]1780년경, 채석장에서 두 번째 두개골이 발견되었다.[13] 이 두개골은 의사 요한 레오나르트 호프만의 관심을 끌었는데, 그는 이것이 악어라고 생각했다. 호프만은 저명한 생물학자 페트루스 캄퍼에게 연락했고, 캄퍼는 연구 결과를 발표하여 두개골이 국제적인 주목을 받게 되었다.[16][10][11]
프랑스 혁명 당시 프랑스 혁명가들은 이 화석에 관심을 가졌고, 1794년 마스트리흐트 포위전 중 약탈했다.[11][13] 1795년 이 두개골은 프랑스 국립 자연사 박물관으로 보내졌고(MNHN AC 9648),[13] 1800년까지 캄퍼의 아들 아드리안 질레스 캄퍼는 이 화석이 모니터 도마뱀과 유사하지만 다른 현대 동물과는 다른 해양 파충류에 속한다고 결론지었다.[14] 1808년 조르주 퀴비에는 캄퍼 주니어의 관찰을 확인했다.[15][8][16]
이 두개골은 퀴비에가 멸종 개념을 추측하고, 격변설 이론과 진화 이론의 전조가 되는 데 중요한 역할을 했다.[17] 당시에는 종의 멸종을 믿지 않았고, 화석은 현존하는 종으로 해석되었다.[17] 퀴비에는 1812년에 "마스트리흐트의 유명한 동물을 정확하게 결정하는 것은 지구의 역사뿐만 아니라 동물학적 법칙의 이론에도 중요하다"고 선언했다.[13]
1822년 윌리엄 다니엘 코니베어는 뫼즈강의 라틴어 이름 'Mosa'와 도마뱀을 뜻하는 고대 그리스어 '사우로스'를 합쳐 '모사사우루스'라는 속명을 만들었다.[18][8][58] 1829년 기드온 맨텔은 호프만을 기리기 위해 종소명 '호프만니(hoffmannii)'를 추가했다.[19] 이후 두 번째 두개골은 새로운 종의 기준 표본(정의 예)으로 지정되었다.[48][58]
1834년 리처드 할란은 미주리 강 근처에서 발견된 주둥이 조각을 바탕으로 'M. 미주리엔시스(M. missouriensis)'를 처음 기술했다.[21] 1881년 에드워드 드링커 코프는 뉴저지주에서 발견된 화석을 바탕으로 'M. 코노돈(M. conodon)'을 기술했다.[31]
1889년 루이 돌로는 벨기에에서 발견된 두개골을 바탕으로 'M. 레모니에리(M. lemonnieri)'를 기술했다.[34] 1952년 카미유 아람부르는 모로코에서 발견된 이빨을 바탕으로 'M. 보게이(M. beaugei)'를 기술했다.[38]
2. 2. 초기 복원
1800년대 초중반 과학자들은 모사사우루스를 물갈퀴가 달린 발과 육상 보행이 가능한 사지를 가진 양서류 해양 파충류로 묘사했다. 이는 1845년 골드푸스(Goldfuss)가 육상 보행 능력의 증거로 탄력적인 척추를 제시하고 일부 발가락뼈를 발톱으로 해석한 ''M. missouriensis'' 모식표본에 기반한 것이었다.[28] 그러나 1854년, 헤르만 슐레겔은 모사사우루스가 실제로는 완전한 수생 지느러미를 가졌음을 증명했다. 그는 발톱에 대한 이전 해석이 잘못되었고, 발가락뼈가 근육이나 힘줄 부착 흔적이 없어 보행이 불가능하며, 넓고 평평하여 노를 형성한다는 것을 밝혀냈다.[40][41] 슐레겔의 가설은 당시에는 무시되었지만, 1870년대 오스니얼 찰스 마시와 코프가 북아메리카에서 더 완전한 모사사우루스 유해를 발견하면서 널리 받아들여졌다.[16][41]
초기 복원 중 하나는 벤자민 워터하우스 호킨스가 1852년에서 1854년 사이에[43] 런던 수정궁 공원에 전시된 선사 시대 동물 조각 컬렉션의 일부로 제작한 실물 크기 콘크리트 조각이다.[42] 이 복원은 리처드 오웬의 ''M. hoffmannii'' 모식표본 해석과 왕도마뱀 해부학을 바탕으로, 호킨스는 모사사우루스를 물에서 생활하는 왕도마뱀처럼 묘사했다. 조각은 네모난 머리, 두개골 옆 콧구멍, 눈 주변 연조직, 왕도마뱀을 닮은 입술, 코모도왕도마뱀 같은 대형 도마뱀 비늘과 일치하는 비늘, 지느러미를 가지도록 만들어졌다. 이 모델은 의도적으로 불완전하게 조각되었는데, 마크 위튼은 시간과 비용 절약을 위한 것으로 추정했다. 조각의 많은 부분은 당시에도 부정확하다고 여겨질 수 있었다. 1845년 골드푸스의 ''M. missouriensis'' 연구를 고려하지 않아 더 좁은 두개골, 두개골 상단 콧구멍, 양서류 육상 사지(현대 기준으로는 부정확)가 필요했다.[42][28]
3. 형태
모사사우루스는 완전한 수생 생활에 적응한 파생된 모사사우루스류로, 유선형 몸체와 두 쌍의 지느러미, 아래쪽으로 굽은 꼬리를 가졌다. 과거에는 파생된 모사사우루스류가 거대한 물갈퀴 바다뱀과 유사한 것으로 묘사되었지만, 현재는 수렴 진화를 통해 어룡, 탈라토수쿠스아목, 고대고래아목과 같은 다른 대형 해양 척추동물과 더 유사한 체격을 가진 것으로 이해되고 있다.[170][171][172]
모식종인 *M. 호프만니*는 최대 몸길이가 15m에 달하는 거대한 종으로, 백악기 바다의 최상위 포식자였다.[168] *M. 미주리엔시스*와 *M. 레모니에리*는 *M. 호프만니*보다 작지만, 더 완전한 화석으로 알려져 있다. *M. 보게이*는 *M. 호프만니*와 비슷하거나 약간 더 큰 크기였을 것으로 추정된다. *M. 코노돈*은 속 내에서 비교적 작은 크기였다.
모사사우루스의 몸길이는 종에 따라 차이가 있는데, 대략적인 크기는 다음과 같다.
3. 1. 두개골
모사사우루스의 두개골은 원뿔형이며 짧은 주둥이로 가늘어진다. ''M. hoffmannii''의 주둥이는 무딘 반면,[58] ''M. lemonnieri''의 주둥이는 뾰족하다.[82] 두 턱에는 강력한 원뿔형 이빨이 늘어서 있다.[58][48] 턱뼈 윗부분에는 구멍이라고 하는 작은 구멍들이 턱선과 평행하게 한 줄로 늘어서 있는데, 턱 신경의 말단 가지를 고정하는 데 사용된다.[48] 종에 따라 주둥이 형태, 턱뼈와 이빨의 특징이 다르다.[57][82] ''M. hoffmannii''의 이빨뼈 상단 가장자리는 약간 위로 휘어져 있지만,[58] ''M. lemonnieri''의 두개골은 거의 완벽하게 곧은 턱선을 가지고 있다.[82]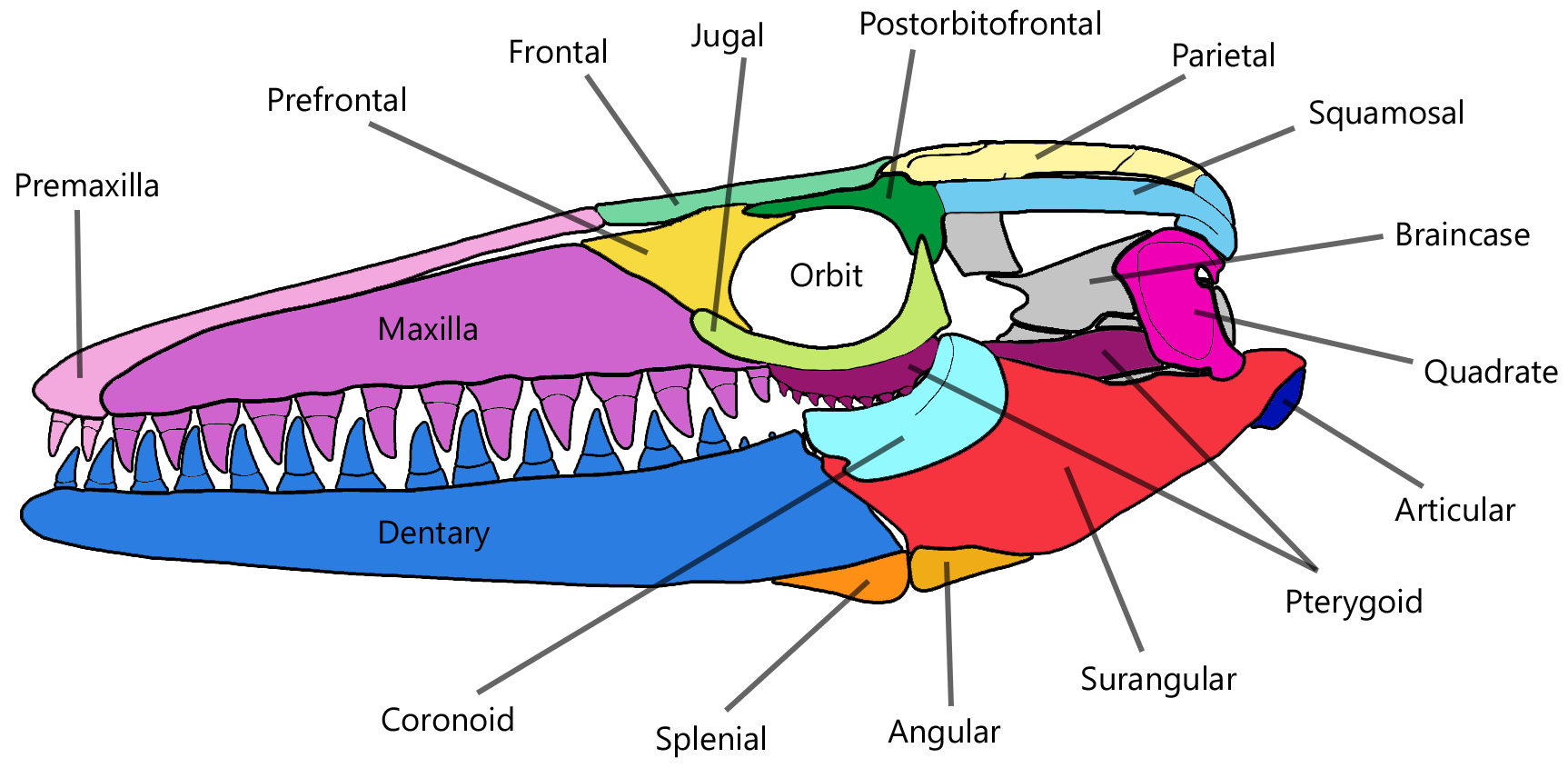
외부 콧구멍은 적당한 크기이며 ''M. hoffmannii''에서 두개골 길이의 약 21–24%를 차지한다. 콧구멍은 거의 모든 다른 모사사우루스보다 두개골 뒤쪽으로 더 멀리 위치하며, 넷째 또는 다섯 번째 상악 치아 위에서 시작한다.[48]
뇌두개는 다른 모사사우루스에 비해 좁고 비교적 작다. 두정골 구멍은 두정안과 관련이 있으며, 모사사우루스과에서 가장 작다.[48] 아래턱을 두개골에 연결하는 사각형 뼈는 키가 크고 직사각형 모양이며, 청각 구조를 수용한다. 고막은 고막 날개라고 하는 오목한 우울증 안에 위치한다.[59]
3. 2. 치아
''모사사우루스'' 이빨의 특징은 종에 따라 다르지만, 먹이를 자르는 데 특화된 디자인, 매우 각주형 표면(프리즘이라고 불리는 평평한 면으로 형성된 에나멜 둘레), 두 개의 반대 절단면이 특징이다.[57][60][61][62] ''모사사우루스'' 이빨은 ''M. conodon''과 ''M. lemonnieri''의 이빨을 제외하고 크고 튼튼하며, 이 종들은 더 가늘어 보인다.[57][60]
''모사사우루스''의 절단면은 종에 따라 다르다. ''M. hoffmannii''와 ''M. missouriensis''의 절단면은 섬세하게 톱니 모양을 이루고 있는 반면,[58][63] ''M. conodon''과 ''M. lemonnieri''에서는 톱니 모양이 존재하지 않는다.[37] ''M. beaugei''의 절단면은 톱니 모양도 부드럽지도 않고, 대신 크레눌레이션으로 알려진 미세한 주름이 있다.[60] ''모사사우루스'' 이빨의 프리즘 수는 이빨 유형에 따라 약간 다를 수 있으며 일반적인 패턴은 종에 따라 다르다—''M. hoffmannii''는 순측(바깥쪽을 향하는 면)에 2~3개의 프리즘이 있고 설측(혀를 향하는 면)에는 프리즘이 없었으며, ''M. missouriensis''는 4~6개의 순측 프리즘과 8개의 설측 프리즘, ''M. lemonnieri''는 8~10개의 순측 프리즘, ''M. beaugei''는 3~5개의 순측 프리즘과 8~9개의 설측 프리즘을 가지고 있었다.[60]
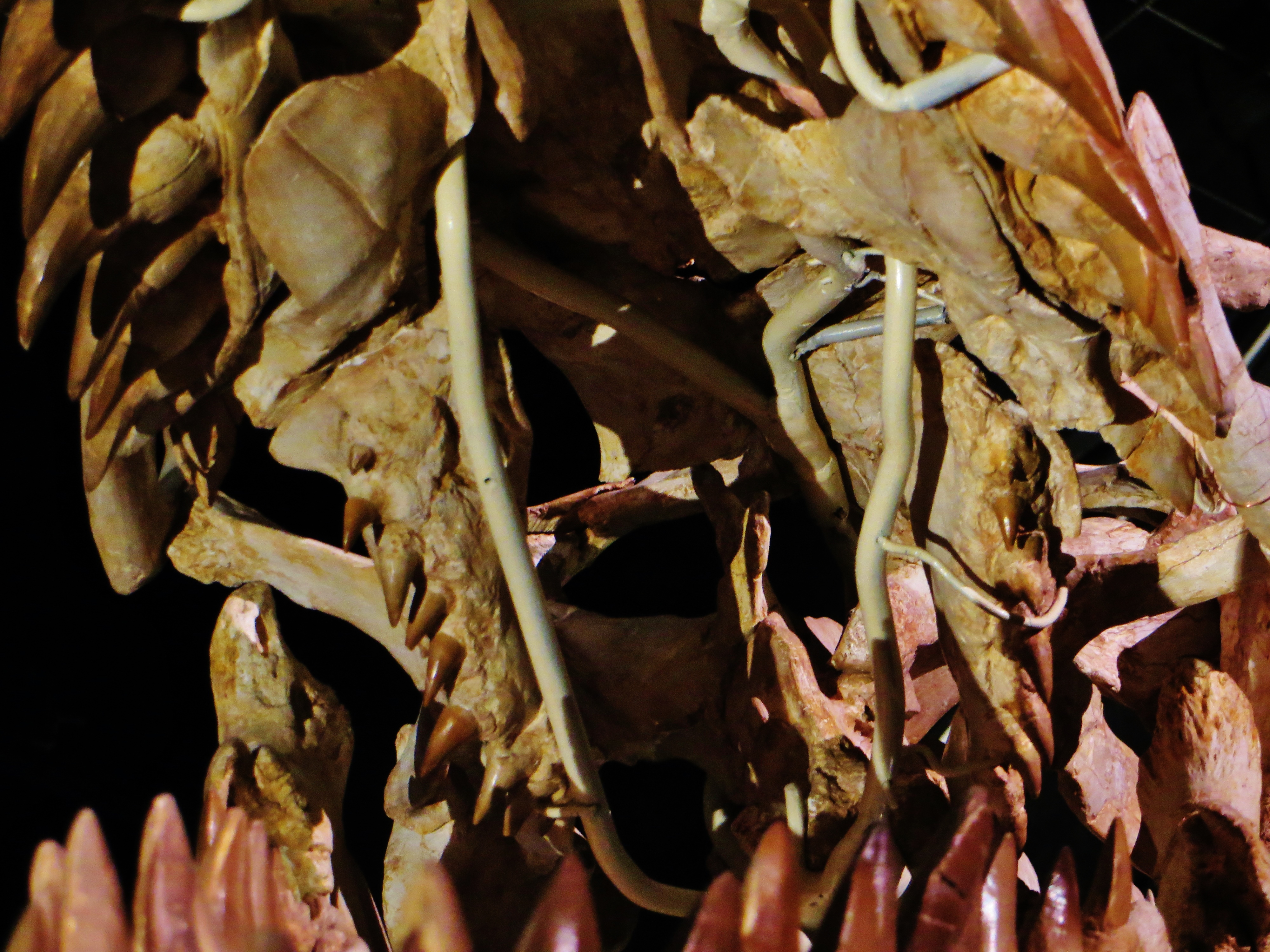
모든 모사사우루스처럼, ''모사사우루스''는 이빨이 위치한 턱뼈에 따라 분류된 네 가지 유형의 이빨을 가지고 있었다. 위턱에는 전상악 치아, 상악 치아, 익상골 치아의 세 가지 유형이 있었다. 아래턱에는 치아의 한 종류인 치아만 존재했다. 각 턱 줄에서 앞쪽에서 뒤쪽으로, ''모사사우루스''는 위턱에 두 개의 전상악 치아, 12~16개의 상악 치아, 8~16개의 익상골 치아와 아래턱에 14~17개의 치아를 가지고 있었다. 이빨은 작은 익상골 치아를 제외하고 턱 전체에서 미미한 차이만 있을 뿐 크기와 모양이 거의 일치했다(동형치).[20][57][60][64] 상악골, 익상골, 치아의 이빨 수는 종에 따라 다르며 때로는 개체에 따라서도 다르다—''M. hoffmannii''는 14~16개의 상악 치아, 14~15개의 치아, 8개의 익상골 치아를 가지고 있었고;[57][47][48] ''M. missouriensis''는 14~15개의 상악 치아, 14~15개의 치아, 8~9개의 익상골 치아를 가지고 있었으며;[20][60][65] ''M. conodon''은 14~15개의 상악 치아, 16~17개의 치아, 8개의 익상골 치아를 가지고 있었고;[57][60] ''M. lemonnieri''는 15개의 상악 치아, 14~17개의 치아, 11~12개의 익상골 치아를 가지고 있었고;[82][57][60] ''M. beaugei''는 12~13개의 상악 치아, 14~16개의 치아, 6개 이상의 익상골 치아를 가지고 있었다.[60] 노스다코타의 펨비나 협곡 주립 휴양지에서 발견된 ''Mosasaurus''의 한 미정 표본은 ''M. conodon''과 유사하며, 알려진 종보다 훨씬 많은 16개의 익상골 치아가 있는 것으로 밝혀졌다.[64]
치열은 치조치였다(이빨 뿌리가 턱뼈 안에 깊숙이 시멘트됨). 이빨은 교체 이빨이 원래 이빨의 뿌리 안에서 발달한 다음 턱에서 밀려나는 과정을 통해 끊임없이 탈락했다.[66] ''M. hoffmannii'' 상악 치아에 대해 수행된 화학 연구에서는 상아모세포의 평균 증착 속도를 측정했는데, 이 세포는 상아질을 형성하는 데 책임이 있으며, 하루에 10.9um였다. 이는 매일 형성되는 상아질의 증가 표지인 von Ebner선을 관찰하여 얻은 결과였다. 상아모세포가 이빨에서 관찰된 정도로 발달하는 데 511일, 상아질이 발달하는 데 233일이 걸리는 것으로 추정되었다.[67]
3. 3. 골격
모사사우루스는 7개의 경추, 32~40개의 등뼈(척추), 9~22개의 미추, 68~90개의 꼬리 척추를 가졌으며, 종에 따라 척추의 수가 다르다.[140] 늑골은 깊고 반원형으로 통 모양의 가슴을 형성하며, 광범위한 연골이 늑골을 흉골과 연결하여 호흡 운동과 압축을 용이하게 했다.[140] 뼈의 질감은 현대 고래와 유사하여 고도의 수생 적응 능력과 중성 부력을 가졌음을 나타낸다.[140]꼬리는 이엽 꼬리 지느러미를 지지하며, 중앙 근처에서 아래로 굽어 강력한 추진력을 제공했다.[140] 앞다리는 넓고 튼튼하며, 노 모양의 지느러미는 조종 기능을 담당했다.[140]
4. 분류
모사사우루스는 모사사우루스과 모사사우루스아과의 모식 속이며, 뱀목(도마뱀과 뱀을 포함)에 속한다. 척추 표현 측면에서 가장 완전한 ''모사사우루스'' 골격 중 하나는 7개의 경추(목) 척추, 38개의 등 척추 (흉추 및 요추)를 가지고 있으며, 8개의 미추 (혈관궁이 없는 앞쪽 꼬리 척추)와 꼬리에 68개의 미추가 있다.[57] 모든 ''모사사우루스'' 종은 7개의 경추를 가지고 있지만, 다른 척추 수는 종에 따라 다르다.
모사사우루스는 에레미아사우루스, 플로토사우루스,[75] 모아나사우루스와 함께[76] 모사사우루스족 또는 플로토사우루스니라고 불리는 모사사우루스아과 내의 한 부족을 형성한다.[46][77][75] 일반적으로 5종(모사사우루스 호프마니(타입종), 모사사우루스 코노돈, 모사사우루스 레모니에리, 모사사우루스 보게이, 모사사우루스 미주리엔시스)이 유효한 것으로 여겨진다.[156]
''모사사우루스''의 늑골은 다른 모사사우루스와 비교하여 특이하게 깊고 거의 완벽한 반원을 형성하여 통 모양의 가슴을 가지고 있다. 늑골은 함께 융합되기보다는 광범위한 연골을 통해 흉골과 연결되어, 더 깊은 물속에서 호흡 운동과 압축을 용이하게 했을 것이다.[48] 뼈의 질감은 현대 고래와 거의 동일하여, ''모사사우루스''가 고도의 수생 적응 능력과 중성 부력을 가졌음을 나타낸다.
''모사사우루스''의 꼬리 구조는 연조직 증거로 이엽 꼬리가 알려진 ''Prognathodon''과 유사하다.[69] 꼬리 척추는 꼬리 중앙 주위에서 점차 짧아지고 중앙 뒤에서 길어져, 꼬리 중앙 주위에 강성을 보이고 그 뒤로 뛰어난 유연성을 보였을 것으로 추정된다. 대부분의 진보된 모사사우루스와 마찬가지로, 꼬리는 중앙에 가까워지면서 약간 아래로 굽지만, 이 굽힘은 등쪽 평면에서 약간 벗어난다. ''모사사우루스''는 또한 꼬리 중간 근처에서 굽는 각 미추 하단에 큰 혈관궁을 가지고 있는데, 이는 어룡과 같은 다른 해양 파충류에서 혈관궁의 감소와 대조된다. 이러한 특징과 다른 특징들은 ''모사사우루스''에서 크고 강력한 노 모양 꼬리지느러미를 뒷받침한다.[68]
''모사사우루스''의 앞다리는 넓고 튼튼하다.[57][48] 견갑골과 상완골은 부채꼴 모양이며 높이보다 넓다. 요골과 척골은 짧지만, 요골이 척골보다 더 키가 크고 크다.[57] 장골은 막대 모양이고 가늘다. 5세트의 중수골과 지골(손가락 뼈)은 노를 감싸고 지지하며, 다섯 번째 세트는 더 짧고 나머지와 오프셋되어 있었다. 노의 전체적인 구조는 압축되어 ''Plotosaurus''와 유사하며, 더 빠른 수영에 적합했다.[57][48] 뒷다리에서, 노는 4세트의 손가락에 의해 지지된다.[20]
4. 1. 분류의 역사
19세기 과학자들은 최초 기술에서 ''모사사우루스''에 적절한 진단을 내리지 못해 속(屬)이 어떻게 정의되는지에 대한 모호성을 초래했다. 이 때문에 ''모사사우루스''는 최대 50개의 서로 다른 종을 포함하는 쓰레기통 분류군이 되었다.[58]
1764년 네덜란드 마스트리흐트 근처 백악 채석장에서 과학계에 알려진 최초의 ''모사사우루스'' 화석이 두개골 형태로 발견되었으며, 처음에는 고래로 확인되었다.[7] TM 7424로 분류된 이 표본은 현재 하를럼의 테일러스 박물관에 전시되어 있다.[8][16][13] 이후 1780년경 채석장에서 두 번째 두개골이 발견되었고, 이는 의사 요한 레오나르트 호프만의 관심을 끌었는데, 그는 이것이 악어라고 생각했다.[16][10][11] 그는 저명한 생물학자 페트루스 캄퍼에게 연락했고, 캄퍼가 고래로 확인하는 연구를 발표한 후 두개골은 국제적인 주목을 받았다.
1794년 마스트리흐트 포위전 중 프랑스 혁명가들이 이 화석을 약탈했다.[11][13] 1795년 두 번째 두개골은 프랑스 국립 자연사 박물관으로 보내졌고, 나중에 MNHN AC 9648로 분류되었다.[13] 1800년, 캄퍼의 아들 아드리안 질레스 캄퍼는 이 화석이 모니터 도마뱀과 유사성을 공유하지만 다른 현대 동물과는 다른 해양 파충류에 속한다고 결론지었다.[14] 조르주 퀴비에는 1808년에 발표된 연구에서 캄퍼 주니어의 관찰을 확인했다.[15][8][16]
1822년 윌리엄 다니엘 코니베어는 라틴어 ''Mosa'' "뫼즈강"과 고대 그리스어 σαῦρος(''saûros'', "도마뱀")에서 ''모사사우루스''(Mosasaurus)라는 속명을 만들었다.[18][8][58] 1829년, 기드온 맨텔은 호프만을 기리기 위해 종 소명 ''호프만니''(hoffmannii)를 추가했다.[19] 이후 두 번째 두개골은 새로운 종의 기준 표본으로 지정되었다.[48][58]
2017년 Hallie Street와 Michael Caldwell의 연구는 ''M. hoffmannii'' 홀로타입에 대한 최초의 적절한 진단 및 설명을 수행하여 ''M. hoffmannii'', ''M. missouriensis'', ''M. conodon'', ''M. lemonnieri'' 및 ''M. beaugei'' 5종을 유효한 것으로 확인했다.[58]
4. 2. 계통 및 진화
모사사우루스는 모사사우루스과 모사사우루스아과의 모식 속이며, 뱀목(도마뱀과 뱀을 포함)에 속한다. 모사사우루스와 현존하는 뱀목 동물 사이의 관계는 과학자들이 모사사우루스의 가장 가까운 현존하는 친척이 왕도마뱀인지 뱀인지에 대해 격렬하게 논쟁을 벌이고 있기 때문에 여전히 논쟁의 여지가 있다.[45][74] ''모사사우루스''는 에레미아사우루스, 플로토사우루스[75], 모아나사우루스와 함께[76] 모사사우루스족 또는 플로토사우루스니라고 불리는 모사사우루스아과 내의 한 부족을 형성한다.[46][77][75]모사사우루스의 진화 연구에 관한 초기 시도는 1967년 러셀(Russell)에 의해 이루어졌다.[46][77] 그는 모사사우루스가 클리다스테스와 유사한 모사사우르에서 진화하여 두 계통으로 분화되었으며, 한 계통은 ''M. conodon''을, 다른 계통은 ''M. ivoensis'', ''M. missouriensis'', ''M. maximus-hoffmanni'' 순으로 이어지는 연대종 계열을 낳았다고 제안했다.[46] 그러나 러셀은 초기의 계통발생학적 방법을 사용했고, 분지학을 사용하지 않았다.[77]
1997년, 벨(Bell)은 북미 모사사우르에 대한 최초의 분지학적 연구를 발표했다. ''M. missouriensis'', ''M. conodon'', ''M. maximus'', 불확정적인 표본(UNSM 77040)을 포함한 그의 연구 결과 중 일부는 러셀(1967)의 연구와 일치했는데, 예를 들어 모사사우루스가 ''Clidastes''를 포함하는 조상 그룹에서 유래하고, ''M. conodon''이 속의 가장 기저적인 종이라는 점이다. 러셀(1967)과 달리, 벨은 ''Mosasaurus''가 ''Globidens''와 ''Prognathodon''을 포함하는 다른 그룹과 자매 관계에 있으며, ''M. maximus''가 ''Plotosaurus''의 자매 종이라는 것을 밝혀냈다. 후자는 ''Mosasaurus''를 측계통군으로 만들었지만, 벨(1997)은 ''Plotosaurus''를 별개의 속으로 인식했다.[77]
벨의 연구는 이후 연구의 선례가 되었으며, 대부분 ''Mosasaurus''의 계통 분류를 변경하지 않았다.[20][73] 하지만, 몇몇 후속 연구에서는 ''Mosasaurus''와 ''Plotosaurus''의 자매 그룹이 사용된 데이터 해석 방법에 따라 ''Eremiasaurus'' 또는 ''Plesiotylosaurus''라고 밝혀졌으며,[75][78][79] 적어도 한 연구에서는 ''M. missouriensis''가 ''M. conodon'' 대신 속의 가장 기저적인 종으로 밝혀졌다.[80] 2014년, 코니시(Konishi)와 동료들은 벨의 연구에 대한 의존성에 대해 여러 가지 우려를 표명했다. 첫째, 세 종의 북미 종인 ''M. hoffmannii/M. maximus'', ''M. missouriensis'', ''M. conodon''만을 포함함으로써 속이 심각하게 과소 대표되었으며, 그 결과, 속에서 가장 완벽하게 알려진 종 중 하나인 ''M. lemonnieri''와 같은 종들이 무시되어 계통발생학적 결과에 영향을 미쳤다. 둘째, 명확한 홀형 진단 부재로 인해 연구는 ''Mosasaurus'' 속의 불확실하고 흔들리는 분류법에 의존했으며, 이는 속의 측계통군 지위의 원인이 되었을 수 있다.[73][20] 셋째, 당시 대형 모사사우린의 골격 해부학에 대한 비교 연구가 여전히 부족했다.[20] 이러한 문제들은 스트리트(Street)의 2016년 논문에서 업데이트된 계통발생학적 분석을 통해 해결되었다.[73]
콘래드(Conrad)는 2008년 계통발생학적 분석에서 독특하게 ''M. hoffmannii''와 ''M. lemonnieri''만을 사용했으며, 그 결과 ''M. hoffmannii''가 (가장 기저적인 종에서 가장 덜 기저적인 종 순으로) ''Globidens'', ''M. lemonnieri'', ''Goronyosaurus'', ''Plotosaurus''를 포함하는 수많은 자손 분류군의 기저가 된다는 것을 밝혀냈다. 이 결과는 ''M. hoffmannii''와 ''M. lemonnieri''가 동일한 속에 속하지 않는다는 것을 시사한다.[81] 그러나, 이 연구는 모사사우르 분류가 아닌 전체 유린목 그룹의 관계에 초점을 맞추었기 때문에, 모사사우르 종에 대한 전통적인 계통발생학적 연구에 비정통적인 방법을 사용했다. 그 결과, 일부 고생물학자들은 콘래드의 2008년 연구에서 ''Mosasaurus''의 특정 배치와 같은 하위 분류 결과에 기술적인 문제가 있을 수 있으며, 부정확할 수 있다고 경고한다.[79]
다음 계통수 왼쪽 (토폴로지 A)은 마디아 & 코(Cau) (2017)의 모사사우린 아과에 대한 가장 최근의 주요 계통발생학적 분석에서 베이지안 분석에 의해 추론된 최대 클레이드 신뢰도 트리를 수정한 것이며, 이는 시모이스(Simões) 등(2017)의 더 큰 연구를 개선한 것으로 설명되었다.[78] 오른쪽의 계통수 (토폴로지 B)는 모사사우린에 대한 수정을 제안하는 스트리트(Street)의 2016년 박사 학위 논문에서 수정되었으며, 제안된 새로운 분류군과 이름 변경이 작은 따옴표로 표시되었다.[73]
{{clade gallery |width=450px |height=600px
|caption1=토폴로지 A:
|header1=마디아 & 코(2017)의 최대 클레이드 신뢰도 트리[78]
|cladogram1=
{{clade| style=font-size:85%;line-height:85%
|label1x=Mosasaurinae
|1={{clade
|1='''''Dallasaurus turneri'''''
|2={{clade
|1='''''Clidastes liodontus'''''
|2={{clade
|1='''''Clidastes moorevillensis'''''
|2={{clade
|1='''''Clidastes propython'''''
|2={{clade
|label1=Mosasaurini
|1={{clade
|1={{clade
|1={{clade
|1={{clade
|1='''''Prognathodon overtoni'''''
|2='''''Prognathodon rapax'''''
}}
|2={{clade
|1={{clade
|1={{clade
|1='''''Prognathodon saturator'''''
|2={{clade
|1='''''Prognathodon currii'''''
|2='''''Prognathodon solvayi'''''
}}
}}
}}
|2={{clade
|1='''''Prognathodon waiparaensis'''''
|2={{clade
|1='''''Prognathodon kianda'''''
|2='''''Eremiasaurus heterodontus'''''
}}
}}
}}
}}
}}
|2={{clade|style2=background-color:#eeccFF;
|1='''''Plesiotylosaurus crassidens'''''
|2={{clade
|1='''''Mosasaurus conodon'''''|style1=background-color:#ffa1a1;
|2={{clade
|1='''''Mosasaurus missouriensis'''''
|2={{clade
|1='''''Mosasaurus hoffmannii'''''
|2='''''Plotosaurus bennisoni'''''|style2=background-color:#bbedFF;
}}
}}
}}
}}
|2={{clade
|1='''''Globidens alabamaensis'''''
|2='''''Globidens dakotensis'''''
}}
}}}}}}}}}}}}
'''그룹의 위치'''
'''개별 분류군의 위치'''
|caption2=토폴로지 B:
|header2=스트리트(2016)의 제안된 수정[73]
|cladogram2=
{{clade| style=font-size:85%;line-height:85%
|label1x=Mosasaurinae
|1={{clade
|1='''''Prognathodon solvayi'''''
|2={{clade
|1='''''Clidastes propython'''''
|2={{clade
|1={{clade
|1='''''Clidastes liodontus'''''
|2='''''Clidastes moorevillensis'''''
}}
|2={{clade
|1={{clade
|1='''''Globidens alabamaensis'''''
|2='''''Globidens dakotensis'''''
}}
|2={{clade
|1='''''Prognathodon kianda'''''
|2={{clade
|1={{clade
|1='''''Eremiasaurus heterodontus'''''
|2={{clade
|1={{clade
|1='''''Prognathodon overtoni'''''
|2={{clade
|1='''''Prognathodon saturator'''''
|2={{clade
|1='''''Prognathodon currii'''''
|2='''''Prognathodon rapax'''''
}}
}}
}}
|2={{clade
|1='''''Plesiotylosaurus crassidens'''''
|2={{clade
|1='''''Marichimaera waiparaensis'''''
|label2=Mosasaurini
|2={{clade
|1={{clade
|1='''''Amblyrhynchosaurus wiffeni'''''
|2={{clade
|1='''''Moanasaurus hobetsuensis'''''|style1=background-color:#bbFFbb;
|2={{clade
|1='''''Moanasaurus mangahouange'''''
|2='''''Moanasaurus longirostis'''''
}}}}}}
|2={{clade|style1=background-color:#eeccFF;
|1={{clade
|1='''''Mosasaurus missouriensis'''''
|2={{clade
|1='''''Mosasaurus lemonnieri'''''
|2={{clade
|1='''''Mosasaurus hoffmannii'''''
|2='''''Mosasaurus glycys'''''
}}}}}}
|2={{clade
|1='''''Antipodinectes mokoroa'''''|style1=background-color:#bbFFbb;
|2={{clade
|1='''''Umikosaurus prismaticus'''''|style1=background-color:#bbFFbb;
|2={{clade
|1='''''Aktisaurus conodon'''''|style1=background-color:#ffa1a1;
|2='''''Plotosaurus bennisoni'''''|style2=background-color:#bbedFF;
}}}}}}}}}}
}}
}}
}}
}}
}}
}}
}}
}}
}}
}}
모사사우루스과는 여러 아과로 나뉘며, 모사사우루스는 모사사우루스아과에 속한다. 모사사우루스아과는 더 세분화되어 족으로 나뉘며, 모사사우루스는 클리다스테스, 리오돈, 모아나사우루스와 함께 모사사우루스족으로 분류된다.
본 속이 명명된 것이 19세기 전반이기 때문에, 북아메리카와 유럽에서 산출된 수많은 종이 모사사우루스족에 할당되었다. 불완전한 표본이 많이 포함되어 있지만, 유럽의 종은 실제로는 미국의 종의 시노님이라고 많은 연구자가 주장하고 있으며, 어떤 종을 시노님으로 하고 어떤 종을 독립종으로 할지에 대해서는 견해가 갈리고 있다. 예를 들어, 미국에서 산출된 거대한 종 모사사우루스 막시무스는 대부분의 연구자가 모사사우루스 호프마니의 주니어 시노님으로 간주하지만, 두개골의 특징에 의해 양자를 구별할 수 있다고 주장하는 연구자도 있다[156][157]。
일반적으로 5종이 유효한 것으로 여겨지며, 그 5종은 모사사우루스 호프마니(타입종), 모사사우루스 코노돈, 모사사우루스 레모니에리, 모사사우루스 보게이, 모사사우루스 미주리엔시스이다[156]。 또한 일본에서 산출된 모사사우루스 호베츠엔시스와 모사사우루스 프리즈마티쿠스 2종, 뉴질랜드에서 산출된 모사사우루스 모코로아 1종도 2016년에 Street와 Caldwell가 발표한 태평양에서의 모사사우루스아과의 분류학적 재검토에서 유효하다고 인정받았다[158]。
다음 모사사우루스류와 근연한 분류군의 클래도그램은 D.V. Grigoriev(2013)에 근거한다[159]。
{{clade| style=font-size:85%;line-height:85%
|label1=모사사우루스아과
|1={{clade
|1=''Dallasaurus turneri''
|2={{clade
|1={{clade
|1=''Clidastes liodontus''
|2=''Clidastes moorevillensis''
|3=''Clidastes propython'' }}
|2={{clade
|1=''"Prognathodon" kianda''
|2={{clade
|label1=글로비덴스
|1={{clade
|1=''Globidens alabamaensis''
|2=''Globidens dakotensis'' }}
|2={{clade
|1=''"Prognathodon" overtoni''
|2=''"Prognathodon" rapax''
|3=''"Prognathodon" waiparaensis''
|label4=프로그나토돈
|4={{clade
|1=''Prognathodon saturator''
|2={{clade
|1=''Prognathodon currii''
|2={{clade
|1=''Prognathodon solvayi''
|2=''Prognathodon lutugini'' }} }} }}
|label5=모사사우루스족
|5={{clade
|1=''Plesiotylosaurus crassidens''
|2={{clade
|1=''Eremiasaurus heterodontus''
|2={{clade
|1=''Plotosaurus bennisoni''
|label2='''''모사사우루스'''''
|2={{clade
|1='''''Mosasaurus conodon'''''
|2='''''Mosasaurus hoffmanni'''''
|3='''''Mosasaurus missouriensis''''' }} }} }} }} }} }} }} }} }}
5. 고생물학
모사사우루스는 뱀목(도마뱀과 뱀 포함)에 속하는 동물로, 왕도마뱀과 뱀과 같은 부류이지만, 과학자들은 어떤 쪽이 가장 가까운 현존하는 친척인지에 대해 여전히 논쟁을 벌이고 있다.[45][74] 모사사우루스는 에레미아사우루스, 플로토사우루스[75], 모아나사우루스와 함께 모사사우루스아과 내에서 모사사우루스족 또는 플로토사우루스족으로 불리는 부족을 형성한다.[46][77][75]
모사사우루스의 진화 연구는 1967년 러셀(Russell)에 의해 처음 시도되었다.[46][77] 그는 모사사우루스가 클리다스테스와 유사한 모사사우르에서 진화하여 두 계통으로 분화되었으며, 한 계통은 ''M. conodon''을, 다른 계통은 ''M. ivoensis'', ''M. missouriensis'', ''M. maximus-hoffmanni'' 순으로 이어지는 연대종 계열을 낳았다고 제안했다.[46] 그러나 러셀은 초기의 계통발생학적 방법을 사용했고, 분지학을 사용하지 않았다.[77]
1997년 벨(Bell)은 북미 모사사우르에 대한 최초의 분지학적 연구를 발표했다.[77] 그의 연구 결과는 러셀의 연구와 일부 일치했는데, 모사사우루스가 ''Clidastes''를 포함하는 조상 그룹에서 유래하고, ''M. conodon''이 속의 가장 기저적인 종이라는 점 등이다. 그러나 벨은 ''Mosasaurus''가 ''Globidens''와 ''Prognathodon''을 포함하는 다른 그룹과 자매 관계에 있으며, ''M. maximus''가 ''Plotosaurus''의 자매 종이라는 것도 밝혀냈다. 이는 ''Mosasaurus''를 측계통군으로 만들었지만, 벨은 ''Plotosaurus''를 별개의 속으로 인식했다.[77]
벨의 연구는 이후 연구의 선례가 되었지만,[20][73] 일부 후속 연구에서는 ''Mosasaurus''와 ''Plotosaurus''의 자매 그룹이 ''Eremiasaurus'' 또는 ''Plesiotylosaurus''라고 밝혀졌으며,[75][78][79] 한 연구에서는 ''M. missouriensis''가 ''M. conodon'' 대신 속의 가장 기저적인 종으로 밝혀졌다.[80] 2014년 코니시(Konishi) 등은 벨의 연구에 대한 의존성에 대해, 북미 세 종만을 포함하여 속이 과소 대표되었고, ''M. lemonnieri''와 같은 종들이 무시되어 결과에 영향을 미쳤으며, 명확한 홀형 진단 부재로 속의 분류법이 불확실하여 측계통군 지위의 원인이 되었을 수 있다는 우려를 표명했다.[73][20] 스트리트(Street)는 2016년 논문에서 업데이트된 계통발생학적 분석을 통해 이러한 문제를 해결했다.[73]
콘래드(Conrad)는 2008년 분석에서 ''M. hoffmannii''와 ''M. lemonnieri''만을 사용하여 ''M. hoffmannii''가 ''Globidens'', ''M. lemonnieri'', ''Goronyosaurus'', ''Plotosaurus''를 포함하는 자손 분류군의 기저가 된다는 것을 밝혔다.[81] 그러나 이 연구는 유린목 그룹의 관계에 초점을 맞추어 비정통적인 방법을 사용했기에, 일부 고생물학자들은 ''Mosasaurus''의 배치 결과에 기술적 문제가 있을 수 있다고 경고한다.[79]
마디아 & 코(Cau) (2017)는 베이지안 분석을 통해 모사사우린 아과에 대한 계통발생학적 분석을 수행하여 최대 클레이드 신뢰도 트리를 수정했다.[78] 스트리트(Street)는 2016년 박사 학위 논문에서 모사사우린에 대한 수정을 제안했다.[73]
모사사우루스는 꼬리를 사용하여 헤엄쳤으며, 수영 방식은 오늘날 고등어와 같은 반가사리형일 가능성이 높다.[68][84] 길쭉한 노와 같은 지느러미는 양력 날개처럼 동물을 조종하는 데 사용되었다. 지느러미는 상완골 바깥쪽 면에서 요골과 척골까지 이어지는 큰 근육 부착과 수정된 관절 덕분에 회전이 가능하여 조종 능력이 향상되었다.[48]
모사사우루스의 뼈는 현대 유린목보다 훨씬 높은 신진대사율을 가졌음을 시사하며, 휴식 대사율은 장수거북과 어룡 및 플레시오사우루스의 중간 정도였다.[85] 모사사우루스는 내온성 동물일 가능성이 높으며, 외부 환경에 관계없이 일정한 체온을 유지했다. 직접적인 증거는 없지만, ''클리다스테스''와 같은 관련 속들에 대한 생화학 연구는 모든 모사사우루스류에서 내온성이 존재했을 가능성을 시사한다. 이는 유린목 중에서 독특하며, 알려진 유일한 예외는 부분적인 내온성을 유지할 수 있는 아르헨티나 검은색 및 흰색 테구이다.[87] 이러한 적응은 넓은 지역에서 먹이를 찾고 사냥할 때 지구력을 높이는 등 여러 이점을 제공했을 것이며,[88] 남극과 같이 더 추운 기후에서 번성할 수 있게 한 요인이었을 수도 있다.[88][89][90][91]
''모사사우루스''는 상대적으로 큰 안와[48]와 큰 공막륜을 가지고 있어 좋은 시력을 가졌음을 시사한다.[82] 안와는 두개골 측면에 위치하여 약 28.5°의 좁은 양안시를 가졌지만,[48][92] 얕은 수면과 같은 2차원 환경을 처리하는 데는 충분했다.[48]
''모사사우루스'' 화석의 뇌 주형은 냄새를 담당하는 후구와 서골코 기관이 제대로 발달하지 않았음을 보여준다. ''M. hoffmannii''에서는 일부 구조가 결여되어 후각이 좋지 않았음을 알 수 있다. ''M. lemonnieri''의 경우 후각 기관이 작지만, ''M. hoffmannii''에 비해 더 발달되어 있고 일부 구성 요소도 갖추고 있다. 이는 ''모사사우루스''에게 후각이 중요하지 않았고, 시각과 같은 다른 감각이 더 유용했을 수 있음을 시사한다.[48]
''모사사우루스''는 다양한 해양 동물을 먹이로 삼는 활동적인 포식자였다.[48][61] 먹이로는 경골어류, 상어, 두족류, 조류, 다른 모사사우루스류,[61] 거북과 같은 해양 파충류가 포함되었을 것으로 추정된다.[48] 후각이 좋지 않아 청소동물이었을 가능성은 낮다. 크고 튼튼하며 날카로운 이빨을 가진 큰 개체들은 거의 모든 동물을 사냥할 수 있었을 것이다.[61]
모사사우루스는 동종 간의 공격적이고 치명적인 전투를 벌였다는 화석 증거가 있다. ''M. conodon'' 골격에는 다른 ''M. conodon''의 이빨이 관통한 상처가 있고 치유 흔적이 없어 치명적인 공격을 받았음을 시사한다.[96] ''M. missouriensis'' 골격에도 이빨이 박혀 있는 경우가 있는데, 치유 흔적이 있어 생존했음을 알 수 있다.[97]
모사사우루스 화석에서는 골절, 감염, 골수염, 무혈관성 괴사, 척추 융합 등 다양한 질병의 증거가 발견되었다.
''모사사우루스''는 태생(새끼를 낳는)이었을 가능성이 높다. ''모사사우루스'' 자체의 태생 증거는 없지만, 다른 모사사우루스류에서 알려져 있다.[104]
5. 1. 두개골 근육 구조 및 역학

1995년, 링엄-솔리아르는 ''M. hoffmannii''의 머리 근육 구조를 연구했다. 근육과 같은 연조직은 쉽게 화석화되지 않기 때문에 근육 구조의 재구성은 주로 두개골의 구조, 두개골의 근육 흔적, 현존하는 왕도마뱀의 근육 구조를 기반으로 했다.[48]
현대 도마뱀의 두개골은 턱의 유연한 움직임을 가능하게 하는 두개골의 4점식 기하학적 구조로 특징지어진다. 이는 동물이 먹이를 더 잘 위치시키고 사냥 시 먹이의 탈출을 방지하기 위한 것으로 보인다. 반면, 현대 도마뱀에서 유연한 회전축을 형성하기 위해 연결되는 전두골과 두정골은 ''M. hoffmannii''의 두개골에서 겹쳐져 있어 3점식 기하학적 두개 구조를 만든다. 이러한 두개 구조는 아래턱 근육의 아래쪽 힘이나 먹이의 위쪽 힘에 의해 발생하는 압축 및 전단력에 저항하기 위해 형성된 강한 맞물림 봉합으로 결합된다. 이처럼 단단하지만 충격 흡수력이 뛰어난 두개골 구조는 강력한 물기 힘을 가능하게 했을 것이다.[48]
모든 모사사우루스처럼, ''모사사우루스''(Mosasaurus)의 아래턱은 앞뒤로 움직일 수 있었다. ''프로그나토돈''과 ''M. lemonnieri''와 같은 많은 모사사우루스에서 이 기능은 주로 톱니 먹이 섭취를 가능하게 하는 데 사용되었는데, 여기서 익상골과 턱은 컨베이어 벨트처럼 포획된 먹이를 입 안으로 "이동"시켰다. 그러나 특히 ''M. lemonnieri''의 턱에 비해 ''M. hoffmannii''의 익상골 치아는 비교적 작아서 톱니 먹이 섭취가 사냥과 먹이 섭취에 비교적 중요하지 않았음을 나타낸다.[48][82] 오히려, ''M. hoffmannii''는 관성 먹이 섭취(동물이 머리와 목을 뒤로 밀어 잡고 있던 먹이를 놓은 다음 즉시 머리와 목을 앞으로 밀어 먹이를 턱으로 물어 닫는 방식[83])를 사용했을 가능성이 높으며, 먹이를 잡는 동안 물기를 돕기 위해 턱 내전을 사용했다. 턱과 두개골에 부착되어 물기 기능에 중요한 역할을 하는 ''대내전근''은 거대하여 ''M. hoffmannii''가 엄청난 물기 힘을 낼 수 있었음을 나타낸다. 아래턱의 길고 좁고 무거운 특성과 관상 돌기에 힘줄이 부착되어 있어 물속에서 적은 에너지 투입으로 입을 빠르게 열고 닫을 수 있었으며, 이는 또한 ''M. hoffmannii''의 강력한 물기 힘에 기여했고 일부 플레시오사우루스에서 볼 수 있는 강력한 ''대하악근''(입을 여는 근육)이 필요하지 않았을 것임을 시사한다.[48]
5. 2. 이동과 체온 조절
모사사우루스는 꼬리를 사용하여 헤엄쳤다. 수영 방식은 오늘날 고등어가 보여주는 것과 같은 반가사리형일 가능성이 높다.[68][84] 길쭉한 노와 같은 지느러미는 동물을 조종하기 위한 양력 날개로 기능했다. 지느러미의 조종 기능은 상완골의 바깥쪽 면에서 요골과 척골까지 이어지는 큰 근육 부착과 수정된 관절로 가능해졌으며, 이는 지느러미 회전 능력을 향상시켰다. 지느러미 사용으로 인한 강력한 힘은 뼈 손상을 초래했을 수 있는데, 이는 관절 부위에서 빈번한 전단력으로 인해 뼈의 나머지 부분에서 머리가 현저하게 분리된 ''M. hoffmannii''의 장골에서 알 수 있다.[48]모사사우루스 뼈의 조직 구조는 현대의 유린목보다 훨씬 높은 신진대사율을 가졌음을 시사하며, 휴식 대사율은 장수거북과 어룡 및 플레시오사우루스의 중간 정도였다.[85] 모사사우루스는 내온성 동물일 가능성이 높으며, 외부 환경과 관계없이 일정한 체온을 유지했다. 속에 특정한 직접적인 증거는 없지만, 관련 모사사우루스 속인 ''클리다스테스''와 같은 속의 생화학 연구는 모든 모사사우루스에서 내온성이 존재했을 가능성을 시사한다. 이러한 특징은 유린목 중에서는 독특하며, 알려진 유일한 예외는 부분적인 내온성을 유지할 수 있는 아르헨티나 검은색 및 흰색 테구이다.[87] 이러한 적응은 모사사우루스에게 더 넓은 지역에서 먹이를 찾고 사냥할 때 증가된 지구력을 포함한 여러 가지 이점을 제공했을 것이다.[88] 또한 남극과 같은 지역의 더 추운 기후에서 모사사우루스가 번성할 수 있도록 한 요인이었을 수도 있다.[88][89][90][91]
5. 3. 감각 기능
모사사우루스는 상대적으로 큰 안와와 공막륜을 가지고 있어 시력이 좋았음을 알 수 있다.[48][82] 안와는 두개골 측면에 위치해 28.5° 정도의 좁은 양안시 시야를 가졌지만,[48][92] 얕은 수면과 같은 2차원 환경을 훌륭하게 처리할 수 있게 해주었다.[48]뇌 주형은 냄새 기능을 제어하는 후구와 서골코 기관이 제대로 발달하지 않았음을 보여준다. ''M. hoffmannii''에서는 일부 구조가 부족하여 후각이 좋지 않았음을 나타낸다.[48] ''M. lemonnieri''에서는 이 후각 기관이 여전히 작지만, 더 잘 발달되어 있고 ''M. hoffmannii''에서는 부족한 일부 구성 요소를 가지고 있다. 강력한 후각의 부재는 후각이 ''모사사우루스''에게 특히 중요하지 않았음을 시사한다. 대신 시각과 같은 다른 감각이 더 유용했을 수 있다.[48]
5. 4. 섭식
Mosasaurusla는 활동적인 포식자로, 다양한 해양 동물을 잡아먹었을 것으로 추정된다.[48][61] 먹이로는 경골어류, 상어, 두족류, 조류, 그리고 다른 모사사우루스[61]와 바다거북 같은 해양 파충류가 포함되었을 것으로 보인다.[48] Mosasaurusla는 후각이 좋지 않았기 때문에 청소동물이었을 가능성은 낮다.[48]
Mosasaurusla는 당시 가장 큰 해양 동물 중 하나였으며,[48] 크고 튼튼하며 날카로운 이빨을 가지고 있어, 더 큰 개체들은 거의 모든 동물을 사냥할 수 있었을 것으로 보인다.[61] 링엄-솔리아(1995)는 거대한 바다거북 ''Allopleuron hoffmanni''의 갑피에서 발견된 큰 이빨 자국과 ''M. hoffmannii''의 부러진 턱이 다시 아문 화석을 증거로 Mosasaurusla가 다소 "사나운" 먹이 섭식 행동을 보였다고 제안했다.[48] 이 종은 대개 양안시를 통해 먹이를 효과적으로 발견하고 잡기 위해 큰, 이차원적으로 적응된 눈을 사용하여 매복 포식자로서 바다 표면 근처에서 사냥했을 가능성이 높다.[48] ''M. lemonnieri''와 ''M. conodon''의 화석에서 나온 화학적, 구조적 데이터는 그들이 더 깊은 물에서도 사냥했을 수 있음을 시사한다.[93]
여러 ''M. hoffmannii'' 개체의 화석에 대한 탄소 동위 원소 연구에서는 가장 큰 개체의 경우 모든 모사사우루스에서 가장 낮은 δ13C인 매우 낮은 δ13C 값이 발견되었다. δ13C 값이 낮은 모사사우루스는 영양 단계가 더 높은 경향이 있었는데, 이에 대한 한 가지 요인은 식단이었다. 바다거북이나 다른 큰 해양 파충류와 같이 지질이 풍부한 먹이를 먹으면 δ13C 값이 낮아질 수 있다. ''M. hoffmannii''의 낮은 δ13C 수치는 그들이 최상위 포식자였을 가능성을 보여준다.[61]
현재 위 내용물이 보존된 Mosasaurusla의 예는 단 두 가지이다. 첫 번째는 약 7,500만 년 전에 살았던 작은 ''M. missouriensis''의 잘 보존된 부분 골격으로, 창자에 1m 길이의 물고기 잔해가 절단되고 뚫린 채 들어있다. 이 물고기는 길이가 66cm인 모사사우루스의 두개골 길이보다 훨씬 길어, ''M. missouriensis''가 부위를 절단하여 한 번에 조금씩 먹는 방식으로 머리보다 큰 먹이를 섭취했음을 보여준다. ''Prognathodon''과 같이 튼튼한 먹이를 전문적으로 섭취하는 다른 큰 모사사우루스와의 공존으로 인해 ''M. missouriensis''는 생태적 지위 분화의 한 예로 절단에 적응된 이빨을 사용하여 섭취하기에 가장 적합한 먹이에 더 전문화되었을 가능성이 높다.[20] 두 번째는 마스트리흐트 근처의 종의 모식표본과 동일한 지역에서 발견된 아성체 ''M. hoffmannii'' 부분 골격으로, "Lars"라는 별명을 가지고 있다. 이 표본은 2024년 컨퍼런스에서 어린 Ctenochelys와 비슷한 거북의 부식된 뼈와 창자 부분에서 다른 구별되지만 확인되지 않은 동물의 뼈를 포함하고 있는 것으로 보고되었다. 불확정적인 조류 또는 새와 같은 공룡의 뼈도 모사사우루스와 관련하여 발견되었으며, 이는 창자 내용물을 나타낼 수도 있다.[94]
Mosasaurusla는 화석 앵무고둥 ''Argonautilus catarinae''에서 두 개의 동종 모사사우루스, 즉 어린 개체와 성체의 이빨 자국이 발견된 것으로 보아 새끼에게 사냥하는 법을 가르쳤을 수 있다. 카우프만(2004)의 연구에서 이빨 자국 분석 결과, 모사사우루스는 ''모사사우루스'' 또는 ''Platecarpus''일 가능성이 높다고 결론지었다. 두 이빨 자국의 위치는 앵무고둥의 머리가 향했을 방향에 있으며, 이는 앵무고둥이 도망칠 수 없었고, 따라서 공격 당시 이미 병들었거나 죽었음을 나타낸다. 이 현상은 부모 모사사우루스가 새끼에게 먹이의 대체원으로서 두족류에 대해, 그리고 두족류를 사냥하는 방법에 대해 가르치는 것일 가능성이 있다. 대안적 설명은 이빨 자국이 한 개체의 모사사우루스가 처음에는 앵무고둥을 가볍게 물고, 그 다음에는 더 큰 힘으로 다시 물었던 것이라고 추정한다. 그러나 두 번의 물림 사이의 이빨 간격에는 차이가 있어 다른 턱 크기를 나타낸다.[95]
5. 5. 행동과 고병리학
모사사우루스는 꼬리를 사용하여 헤엄쳤으며, 수영 방식은 오늘날 고등어와 같은 반가사리형일 가능성이 높다.[68][84] 길쭉한 노와 같은 지느러미는 양력 날개처럼 동물을 조종하는 데 사용되었다. 지느러미는 상완골 바깥쪽 면에서 요골과 척골까지 이어지는 큰 근육 부착과 수정된 관절 덕분에 회전이 가능하여 조종 능력이 향상되었다. 지느러미를 사용하면서 발생하는 강력한 힘은 뼈 손상을 유발할 수 있었는데, 이는 ''M. hoffmannii''의 장골에서 관절 부위의 빈번한 전단력으로 인해 뼈의 나머지 부분에서 머리가 분리된 것으로 확인된다.[48]
모사사우루스의 뼈는 현대 유린목보다 훨씬 높은 신진대사율을 가졌음을 시사하며, 휴식 대사율은 장수거북과 어룡 및 플레시오사우루스의 중간 정도였다.[85] 모사사우루스는 내온성 동물일 가능성이 높으며, 외부 환경에 관계없이 일정한 체온을 유지했다. 모사사우루스 속에 대한 직접적인 증거는 없지만, ''클리다스테스''와 같은 관련 속들에 대한 생화학 연구는 모든 모사사우루스류에서 내온성이 존재했을 가능성을 시사한다. 이는 유린목 중에서 독특한 특징이며, 알려진 유일한 예외는 부분적인 내온성을 유지할 수 있는 아르헨티나 검은색 및 흰색 테구이다.[87] 이러한 적응은 모사사우루스가 더 넓은 지역에서 먹이를 찾고 사냥할 때 지구력을 높이는 등 여러 이점을 제공했을 것이다.[88] 또한 남극과 같이 더 추운 기후에서 모사사우루스가 번성할 수 있게 한 요인이었을 수도 있다.[88][89][90][91]

''모사사우루스''는 상대적으로 큰 안와[48]를 가졌고, 안와 지름의 대부분을 차지하는 큰 공막륜을 가지고 있었다.[82] 공막륜은 눈의 크기와 상관관계가 있어 좋은 시력을 가졌음을 시사한다. 안와는 두개골 측면에 위치하여 약 28.5°의 좁은 양안시를 가졌지만,[48][92] ''모사사우루스''가 서식했던 얕은 수면과 같은 2차원 환경을 처리하는 데는 충분했다.[48]
''모사사우루스'' 화석에서 만들어진 뇌 주형은 냄새를 담당하는 후구와 서골코 기관이 제대로 발달하지 않았음을 보여준다. 특히 ''M. hoffmannii''에서는 일부 구조가 아예 결여되어 있어 후각이 좋지 않았음을 알 수 있다. ''M. lemonnieri''의 경우 후각 기관이 여전히 작지만, ''M. hoffmannii''에 비해 더 발달되어 있고 일부 구성 요소도 갖추고 있다. 이러한 후각 능력의 부족은 ''모사사우루스''에게 후각이 그다지 중요하지 않았음을 시사하며, 시각과 같은 다른 감각이 더 유용했을 수 있다.[48]
고생물학자들은 ''모사사우루스''가 다양한 해양 동물을 먹이로 삼는 활동적인 포식자였다는 데 일반적으로 동의한다.[48][61] 먹이로는 경골어류, 상어, 두족류, 조류, 다른 모사사우루스류,[61] 그리고 거북과 같은 해양 파충류가 포함되었을 것으로 추정된다.[48] ''모사사우루스''는 후각이 좋지 않았기 때문에 청소동물이었을 가능성은 낮다. ''모사사우루스''는 당시 가장 큰 해양 동물 중 하나였고,[48] 크고 튼튼하며 날카로운 이빨을 가지고 있어, 과학자들은 이 속의 더 큰 개체들이 거의 모든 동물을 사냥할 수 있었을 것이라고 생각한다.[61]
5. 5. 1. 종내 경쟁
모사사우루스는 동종 간의 공격적이고 치명적인 전투를 벌였다는 화석 증거가 있다. ''M. conodon''의 부분 골격에는 다른 ''M. conodon''의 이빨이 사각형 뼈를 관통하는 등 여러 상처가 있으며, 치유 흔적이 없어 치명적인 공격을 받았음을 시사한다.[96] ''M. missouriensis'' 골격에도 다른 ''M. missouriensis''의 이빨이 박혀 있는 경우가 있는데, 이 경우에는 치유 흔적이 있어 생존했음을 알 수 있다.[97]코니시 타쿠야는 이러한 머리 물기 행동이 구애 중에 발생했을 가능성을 제시했다.[97][98] 다른 ''모사사우루스''의 공격 외에도, 단단한 거북 등껍질을 물려는 시도 등 다른 사건으로 인한 부상일 가능성도 제기된다. 링엄-솔리아르는 동종 간 공격이라면 두개골 부위에 집중되는 경향이 있다고 보았다. 현대 악어처럼 ''모사사우루스''도 머리 잡기 행동을 했을 것으로 추정된다.[99] 동종 간 전투로 추정되는 부상을 입은 화석은 어린 개체가 많지만, 공격자는 피해자와 비슷한 크기였을 가능성이 높다.[97][96] 2006년, 슐프와 그의 동료들은 ''모사사우루스''가 동종 간의 공격으로 인해 때때로 식인 행위를 했을 수 있다고 추측했다.[100]

5. 5. 2. 질병
모사사우루스 화석에서는 골절, 감염, 골수염, 무혈관성 괴사, 척추 융합 등 다양한 질병의 증거가 발견되었다.''M. hoffmannii''의 아래턱뼈 화석에서는 물리적 부상으로 인한 감염성 질환의 증거가 발견되었다. IRSNB R25와 IRSNB R27 표본은 하악골에 골절과 더불어 심각한 세균 감염의 징후를 보인다. 뼈 가골에는 치유 과정의 일부로 추정되는 미세한 궤양성 긁힘이 있고, IRSNB R27에는 부러진 이빨과 얕은 긁힌 자국, 농양 관에 연결된 큰 구멍이 있는 불유합 형태의 뼈 가골이 관찰된다. 이는 종종 동종 간의 공격으로 인한 치아 자국과 유사하다고 묘사되기도 한다. 이러한 감염은 인접한 손상된 치아로 퍼져 충치를 유발했을 가능성이 있으며, 외상 후 또는 이차 감염으로 인해 더 깊은 조직에 침투했을 수 있다. 그러나 하악골 앞쪽은 양호한 상태를 유지하여 동맥과 삼차 신경이 손상되지 않았음을 시사하는데, 이는 모사사우루스가 치유 동안 골절을 고정하는 효율적인 과정을 통해 주요 혈관과 신경의 손상을 막았을 수 있음을 보여준다.[99]
2006년에는 ''M. hoffmannii''의 사각형 뼈에서 심각한 골수염의 흔적이 발견되었다. 이는 패혈성 관절염에서 시작되어 뼈의 상당 부분이 농양으로 축소될 정도로 진행되었을 가능성이 높다. 많은 양의 뼈 복구 조직은 감염과 치유 과정이 수개월 동안 지속되었음을 시사한다. 이 정도의 뼈 감염은 극심한 고통과 턱 사용 능력 저하, 호흡 곤란을 야기했을 것으로 추정된다. 연구자들은 이 개체가 생존을 위해 오징어와 같은 부드러운 먹이를 통째로 삼키는 방식으로 식단을 변경했을 것이라고 추측한다. 감염의 원인은 불분명하지만, 종내 공격으로 인한 것일 가능성도 제기된다.[100]
무혈관성 괴사는 ''M. lemonnieri''와 ''M. conodon''의 많은 표본에서 발견된다.[61][101][102] 2005년 연구에서는 모사사우루스 척추의 3-17%가 이 질환에 영향을 받았다고 보고되었다.[101] 이는 감압병의 일반적인 결과로, 잦은 잠수 중 질소 기포 형성이나 반복적인 잠수와 짧은 호흡 간격으로 인한 뼈 손상 때문일 수 있다. 이는 해당 종들이 습관적인 심해 잠수부였거나 반복적인 잠수를 했을 가능성을 시사한다. 그러나 급격한 압력 변화로부터 잘 보호된 고막과 같은 적응을 보인 화석도 존재한다.[102]
꼬리 척추의 부자연스러운 융합은 외상이나 질병으로 인한 손상 후 뼈가 골화될 때 발생한다. 2015년 연구에서는 조사된 15개의 ''모사사우루스'' 표본 중 3개에서 꼬리 척추 융합 사례가 발견되었다. 이 중 두 사례는 뼈 감염의 징후를 보였으며, 이는 상어나 다른 포식자의 공격으로 인한 것일 수 있다. 나머지 한 사례는 관절염의 한 형태로 확인되었다.[103]
5. 6. 생활사
모사사우루스는 꼬리를 사용하여 헤엄쳤다. 수영 방식은 오늘날 고등어가 보여주는 것과 같은 반가사리형일 가능성이 높다.[68][84] 길쭉한 노와 같은 지느러미는 동물을 조종하기 위한 양력 날개로 기능했다. 지느러미의 조종 기능은 상완골의 바깥쪽 면에서 요골과 척골까지 이어지는 큰 근육 부착과 수정된 관절로 가능해졌으며, 이는 지느러미 회전 능력을 향상시켰다. 지느러미 사용으로 인한 강력한 힘은 뼈 손상을 초래했을 수 있는데, 이는 관절 부위에서 빈번한 전단력으로 인해 뼈의 나머지 부분에서 머리가 현저하게 분리된 ''M. hoffmannii''의 장골에서 알 수 있다.[48]모사사우루스 뼈의 조직 구조는 현대의 유린목보다 훨씬 높은 신진대사율을 가졌음을 시사하며, 휴식 대사율은 장수거북과 어룡 및 플레시오사우루스의 중간 정도였다.[85] 모사사우루스는 내온성 동물일 가능성이 높으며, 외부 환경과 관계없이 일정한 체온을 유지했다. 속(genus)에 특정한 직접적인 증거는 없지만, 관련 모사사우루스 속인 클리다스테스와 같은 속의 생화학 연구는 모든 모사사우루스에서 내온성이 존재했을 가능성을 시사한다. 이러한 특징은 유린목 중에서는 독특하며, 알려진 유일한 예외는 부분적인 내온성을 유지할 수 있는 아르헨티나 검은색 및 흰색 테구이다.[87] 이러한 적응은 모사사우루스에게 더 넓은 지역에서 먹이를 찾고 사냥할 때 증가된 지구력을 포함한 여러 가지 이점을 제공했을 것이다.[88] 또한 남극과 같은 지역의 더 추운 기후에서 모사사우루스가 번성할 수 있도록 한 요인이었을 수도 있다.[88][89][90][91]
고생물학자들은 일반적으로 ''모사사우루스''가 다양한 해양 동물을 잡아먹는 활동적인 포식자였을 것이라고 동의한다.[48][61] 이 속이 잡아먹었을 가능성이 있는 동물군에는 경골어류, 상어, 두족류, 조류, 그리고 다른 모사사우루스와 거북과 같은 해양 파충류가 포함된다.[48] ''모사사우루스''는 후각이 좋지 않아 청소동물이었을 가능성은 낮다. ''모사사우루스''는 당시 가장 큰 해양 동물 중 하나였으며,[48] 크고 튼튼한 날카로운 이빨을 가지고 있어 과학자들은 이 속의 더 큰 개체들이 거의 모든 동물을 처리할 수 있었을 것이라고 믿는다.[61] 링엄-솔리아(1995)는 거대한 바다거북 ''Allopleuron hoffmanni''의 갑피에서 큰 이빨 자국과 ''M. hoffmannii''의 부러진 턱이 다시 아문 화석을 통해 ''모사사우루스''가 다소 "사나운" 먹이 행동을 보였다고 제안했다.[48] 이 종은 대개 양안시를 통해 먹이를 효과적으로 발견하고 잡기 위해 큰 이차원적으로 적응된 눈을 사용하여 매복 포식자로서 바다 표면 근처에서 사냥했을 가능성이 높다.[48] ''M. lemonnieri''와 ''M. conodon''의 화석에서 화학적, 구조적 데이터는 그들이 더 깊은 물에서도 사냥했을 수 있음을 시사한다.[93]
여러 ''M. hoffmannii'' 개체의 화석에 대한 탄소 동위 원소 연구에서 가장 큰 개체의 경우 모든 모사사우루스에서 가장 낮은 δ13C인 매우 낮은 δ13C 값이 발견되었다. δ13C 값이 낮은 모사사우루스는 영양 단계가 더 높은 경향이 있었고, 이에 대한 한 가지 요인은 식단이었다. 바다거북이나 다른 큰 해양 파충류와 같이 지질이 풍부한 먹이를 먹으면 δ13C 값이 낮아질 수 있다. ''M. hoffmannii''의 낮은 δ13C 수치는 그들이 최상위 포식자였을 가능성을 강화한다.[61]
현재 위 내용물이 보존된 ''모사사우루스''의 예는 단 두 가지이다. 첫 번째는 약 7,500만 년 전에 살았던 작은 ''M. missouriensis''의 잘 보존된 부분 골격으로, 창자가 1m 길이의 물고기 잔해가 절단되고 뚫린 채 들어있다. 이 물고기는 길이가 66cm인 모사사우루스의 두개골 길이보다 훨씬 길어, ''M. missouriensis''가 부위를 절단하여 한 번에 조금씩 먹는 방식으로 머리보다 큰 먹이를 섭취했음을 확인시켜준다. 프로그나토돈과 같이 튼튼한 먹이를 전문적으로 섭취하는 다른 큰 모사사우루스와의 공존으로 인해 ''M. missouriensis''는 생태적 지위 분화의 한 예로 절단에 적응된 이빨을 사용하여 섭취하기에 가장 적합한 먹이에 더 전문화되었을 가능성이 높다.[20] 두 번째는 마스트리흐트 근처의 종의 모식표본과 동일한 지역에서 발견된 아성체 ''M. hoffmannii'' 부분 골격으로, "Lars"라는 별명을 가지고 있다. 이 표본은 2024년 컨퍼런스에서 어린 Ctenochelys와 비슷한 거북의 부식된 뼈와 창자 부분에서 다른 구별되지만 확인되지 않은 동물의 뼈를 포함하고 있는 것으로 보고되었다. 불확정적인 조류 또는 새와 같은 공룡의 뼈도 모사사우루스와 관련하여 발견되었으며, 이는 창자 내용물을 나타낼 수도 있다.[94]
''모사사우루스''는 화석 앵무고둥 ''Argonautilus catarinae''에서 두 개의 동종 모사사우루스, 즉 어린 개체와 성체의 이빨 자국이 발견된 것으로 보아 새끼에게 사냥하는 법을 가르쳤을 수 있다. 카우프만(2004)의 연구에서 이빨 자국 분석 결과, 모사사우루스는 ''모사사우루스'' 또는 ''Platecarpus''일 가능성이 높다고 결론지었다. 두 이빨 자국의 위치는 앵무고둥의 머리가 향했을 방향에 있으며, 이는 앵무고둥이 도망칠 수 없었고, 따라서 공격 당시 이미 병들었거나 죽었음을 나타낸다. 이 현상은 부모 모사사우루스가 새끼에게 먹이의 대체원으로서 두족류에 대해, 그리고 두족류를 사냥하는 방법에 대해 가르치는 것일 가능성이 있다. 대안적 설명은 이빨 자국이 한 개체의 모사사우루스가 처음에는 앵무고둥을 가볍게 물고, 그 다음에는 더 큰 힘으로 다시 물었던 것이라고 추정한다. 그러나 두 번의 물림 사이의 이빨 간격에는 차이가 있어 다른 턱 크기를 나타낸다.[95]
모사사우루스는 동종 간의 공격적이고 치명적인 전투를 벌였다는 화석 증거가 있다. ''M. conodon''의 부분 골격에는 다양한 뼈, 특히 두개골과 목의 뒷부분에 여러 개의 절단, 골절 및 구멍이 있으며, 다른 ''M. conodon''의 이빨이 사각형 뼈를 관통하고 있다. 화석의 부상에서 치유의 징후는 보이지 않아, 모사사우루스가 공격자에 의해 두개골에 치명적인 일격을 받아 죽었음을 시사한다.[96] 마찬가지로, ''M. missouriensis'' 골격에는 다른 ''M. missouriensis''의 이빨이 눈 아래 아래턱에 박혀 있다. 이 경우 상처 주변에 치유의 징후가 있어 사건 생존을 암시한다.[97] 코니시 타쿠야는 현대 도마뱀에서 볼 수 있는 머리 물기 행동이 구애 중에 발생했을 가능성을 제시했다.[97][98] 다른 ''모사사우루스''의 공격은 다른 두개골에서 나타나는 신체적 병리학의 가능한 원인이지만, 단단한 거북 등껍질을 물려고 시도하는 것과 같은 다른 사건에서 발생했을 수도 있다. 2004년, 링검-솔리아르는 이러한 부상이 실제로 동종 간의 공격으로 인한 것이라면 두개골 부위에 집중되는 패턴이 있다는 것을 관찰했다. 현대의 악어는 흔히 턱을 사용하여 상대방의 머리를 잡고 서로 공격하며, 링검-솔리아르는 ''모사사우루스''가 동종 간의 전투 중에 유사한 머리 잡기 행동을 했을 것이라고 가설을 세웠다. 동종 간의 전투로 추정되는 부상을 입은 화석 중 다수는 어린 또는 아성체 ''모사사우루스''이며, 더 작고 약한 개체에 대한 공격이 더 흔했을 수 있다.[99] 그러나 ''M. conodon''과 ''M. missouriensis'' 표본을 공격한 모사사우루스는 피해자와 크기가 비슷했을 가능성이 높다.[97][96] 2006년, 슐프와 그의 동료들은 ''모사사우루스''가 동종 간의 공격으로 인해 때때로 식인 행위를 했을 수 있다고 추측했다.[100]

''모사사우루스''는 오늘날 대부분의 현대 포유류처럼 태생(새끼를 낳는)이었을 가능성이 높다. ''모사사우루스'' 자체의 태생에 대한 증거는 없지만, 다른 여러 모사사우루스류에서 알려져 있다.[104] 예를 들어 임신한 ''카르소사우루스''의 골격[104], 두 개의 모사사우루스 배아 화석과 관련된 ''플리오플라테카르푸스'' 화석,[105] 그리고 원양 (대양) 퇴적층에서 발견된 신생 ''클리다스테스''의 화석이 있다.[104] 이러한 화석 기록과 외부 알 기반 번식에 대한 증거가 전혀 없다는 사실은 ''모사사우루스''의 태생 가능성을 시사한다.[104][105] 어린 ''모사사우루스'' 및 관련 속의 뼈에 대한 미세 해부학적 연구에 따르면 그들의 뼈 구조는 성인과 유사하다. 그들은 얕은 물에서의 생활 방식과 관련된 부력을 지원하기 위해 어린 원시 모사사우루스류에서 발견되는 뼈 질량 증가를 보이지 않으며, 이는 ''모사사우루스''가 조숙성이었다는 것을 의미한다. 즉, 그들은 이미 효율적인 수영 선수였으며 매우 어린 나이에 개방된 물에서 완전히 기능적인 생활 방식을 살았고 새끼를 키울 보육 공간이 필요하지 않았다.[106][104] 유럽과 사우스다코타의 일부 지역에서는 어린 ''M. hoffmannii'', ''M. missouriensis'' 및/또는 ''M. lemonnieri''의 농축된 집합체가 발견되었다. 이 지역은 모두 얕은 해양 퇴적층으로, 어린 ''모사사우루스''가 여전히 얕은 물에서 살았을 수 있음을 시사한다.[107]
''모사사우루스''는 타일로사우루스아과와 프로그나토돈과 같이 최상위 포식자로 여겨지는 다른 대형 포식성 모사사우루스들과 함께 살았다.[48][61] 마스트리히트 절에 유일하게 생존한 속인 Tylosaurus bernardi는 길이가 12.2m에 달했고[134], ''P. saturator''와 같이 공존했던 프로그나토돈의 가장 큰 종은 12m를 넘었다.[61] 이 세 종류의 모사사우루스는 해양 파충류와 같은 비슷한 동물을 잡아먹었다.[20][48][61]
2013년 Schulp와 동료들이 발표한 연구에 따르면, ''M. hoffmannii''와 ''P. saturator''와 같은 모사사우루스가 δ13C 분석을 통해 동일 지역에서 어떻게 공존할 수 있었는지를 구체적으로 조사했다. 과학자들은 동위원소 값의 차이가 생활 방식, 식단, 서식지 선호도와 같은 여러 환경적 요인의 영향을 받기 때문에 자원 분할 수준을 설명하는 데 도움이 될 수 있다는 해석을 활용했다. 마스트리히트 지층의 마스트리히트 절 시대 ''M. hoffmannii''와 ''P. saturator''의 여러 치아에서 δ13C 수준을 비교한 결과, 특정 표본 간에 약간의 수렴이 있었지만, 두 종 간의 평균 δ13C 값은 평균적으로 달랐다. 이는 두 모사사우루스 속이 직접적인 경쟁 갈등 없이 공존하기 위해 서로 다른 서식지에서 먹이를 구하거나 서로 다른 특정 식단을 가졌을 가능성이 높은 생태적 지위 분화의 한 징후이다. ''P. saturator''의 치아는 ''M. hoffmannii''의 치아보다 훨씬 튼튼하며, 특히 거북과 같은 튼튼한 먹이를 잡아먹는 데 적합했다. ''M. hoffmannii'' 또한 거북을 잡아먹었지만, 이빨은 ''P. saturator''에게는 적합하지 않은 더 넓은 범위의 먹이를 처리하도록 제작되었다.[61]
앨버타의 베어포 지층에서 ''모사사우루스''와 프로그나토돈 간의 생태적 지위 분화의 또 다른 사례가 Konishi와 동료들의 2014년 연구에서 문서화되었다. 이 연구는 위 내용물을 바탕으로 ''M. missouriensis''와 ''P. overtoni'' 사이의 식단 분할을 발견했다. ''P. overtoni''의 위 내용물에는 거북과 암모나이트가 포함되어 있어, 더 단단한 먹이에 특화된 식단의 또 다른 예시를 제공했다. 반대로, ''M. missouriensis''의 위 내용물은 물고기로 구성되어 있어, 더 부드러운 먹이에 특화된 식단을 나타냈다. 이러한 적응은 두 모사사우루스 사이의 자원 분할을 유지하는 데 도움이 되었을 것이라고 가설이 세워졌다.[20]
그럼에도 불구하고, 경쟁적 교류는 완전히 피할 수 없었던 것으로 보인다. 또한 ''모사사우루스''와 다른 대형 모사사우루스 종 간의 공격적인 종간 전투의 증거도 있다. 이는 아성체 ''M. hoffmannii''의 화석 두개골에서 거대한 집중적인 타격으로 뇌 케이스에 골절이 발생한 것으로 나타난다. Lingham-Soliar (1998)는 이러한 골절의 형성이 조율된 공격의 특징을 보였고(사고나 화석화 손상이 아닌), 그러한 손상을 일으킬 수 있는 유일하게 알려진 공존 동물은 Tylosaurus bernardi였으며, 튼튼한 화살 모양의 길쭉한 주둥이를 사용했기 때문에 이 타격은 T. bernardi에 의한 돌진 공격으로 인해 발생했다고 주장했다. 이러한 종류의 공격은 병코돌고래가 부리를 사용하여 레몬상어를 죽이거나 격퇴하는 방어 행동과 비교되었으며, T. bernardi가 방심한 ''모사사우루스''를 매복하여 공격했을 것으로 추정된다.[135]
6. 고생태학
모사사우루스는 동족 간의 공격적이고 치명적인 전투를 벌였다는 화석 증거가 있다. ''M. conodon''의 부분 골격에는 두개골과 목 뒷부분에 여러 개의 절단, 골절 및 구멍이 있으며, 다른 ''M. conodon''의 이빨이 사각형 뼈를 관통하고 있다. 화석에서는 부상에서 치유된 흔적은 보이지 않아, 모사사우루스가 공격자의 두개골에 치명적인 타격을 받아 죽었음을 시사한다.[96] ''M. missouriensis'' 골격에는 다른 ''M. missouriensis''의 이빨이 눈 아래 아래턱에 박혀 있는 경우도 있는데, 이 경우에는 상처 주변에 치유된 흔적이 있어 생존했음을 알 수 있다.[97] 코니시 타쿠야는 현대 도마뱀에서 볼 수 있는 머리 물기 행동이 구애 중에 발생했을 가능성을 제시했다.[97][98]
다른 ''모사사우루스''의 공격은 두개골 부상으로 나타나는 신체적 병리학의 가능한 원인이지만, 단단한 거북 등껍질을 물려고 시도하는 것과 같은 다른 사건에서 발생했을 수도 있다. 2004년 링검-솔리아르는 이러한 부상이 동종 간의 공격으로 인한 것이라면 두개골 부위에 집중되는 패턴이 있다는 것을 관찰했다. 현대 악어는 턱을 사용하여 상대방의 머리를 잡고 서로 공격하며, 링검-솔리아르는 ''모사사우루스''가 동종 간의 전투 중에 유사한 머리 잡기 행동을 했을 것이라고 추측했다. 동종 간의 전투로 추정되는 부상을 입은 화석 중 다수는 어린 또는 아성체 ''모사사우루스''이며, 더 작고 약한 개체에 대한 공격이 더 흔했을 수 있다.[99] 그러나 ''M. conodon''과 ''M. missouriensis''를 공격한 모사사우루스는 피해자와 크기가 비슷했을 가능성이 높다.[97][96] 2006년 슐프와 동료들은 ''모사사우루스''가 동종 간의 공격으로 인해 때때로 식인 행위를 했을 수 있다고 추측했다.[100]
6. 1. 분포, 생태계 및 생태적 영향
모사사우루스는 대서양을 가로질러 분포했으며, 미국 중서부와 동해안, 캐나다, 유럽, 튀르키예, 러시아, 레반트, 모로코에서[173] 남아프리카 공화국까지 이어지는 아프리카 해안선, 브라질, 아르헨티나, 남극 등 다양한 지역에서 화석이 발견되었다.[174][175][176]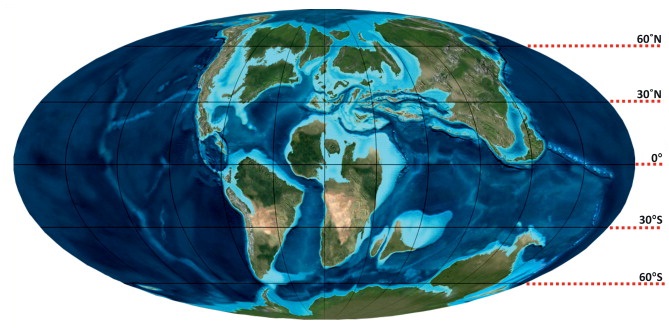 백악기 지구 지도">
백악기 지구 지도">백악기 후기에 이 지역들은 모사사우루스가 서식하는 세 개의 해로, 즉 대서양, 서부 내륙 해로, 테티스 해를 구성했다.[176] 이 해로들은 열대, 아열대, 온대, 아한대 기후를 포함한 여러 해양 기후대를 아울렀다.[176][177][178] 이렇게 다양한 해양 기후는 모사사우루스와 공존하는 다양한 동물군을 형성하는 데 영향을 주었다.
모사사우루스는 꼬리를 사용하여 헤엄쳤으며, 수영 방식은 오늘날 고등어와 같은 반가사리형일 가능성이 높다.[68][84] 길쭉한 노와 같은 지느러미는 동물을 조종하기 위한 양력 날개로 기능했다. 지느러미의 조종 기능은 상완골의 바깥쪽 면에서 요골과 척골까지 이어지는 큰 근육 부착과 수정된 관절로 가능해졌으며, 이는 지느러미 회전 능력을 향상시켰다.[48]
모사사우루스 뼈의 조직 구조는 현대의 유린목보다 훨씬 높은 신진대사율을 가졌음을 시사하며, 휴식 대사율은 장수거북과 어룡 및 플레시오사우루스의 중간 정도였다.[85] 모사사우루스는 내온성 동물일 가능성이 높으며, 외부 환경과 관계없이 일정한 체온을 유지했다. 직접적인 증거는 없지만, 관련 모사사우루스 속인 ''클리다스테스''에 대한 연구는 모든 모사사우루스에서 내온성이 존재했을 가능성을 시사한다. 이러한 특징은 유린목 중에서는 독특하며, 알려진 유일한 예외는 부분적인 내온성을 유지할 수 있는 아르헨티나 검은색 및 흰색 테구이다.[87] 이러한 적응은 모사사우루스에게 더 넓은 지역에서 먹이를 찾고 사냥할 때 증가된 지구력을 포함한 여러 가지 이점을 제공했을 것이며,[88] 남극과 같은 추운 기후에서 모사사우루스가 번성할 수 있도록 한 요인이었을 수도 있다.[88][89][90][91]

물리적 부상으로 인한 감염성 질환의 증거가 있는 ''M. hoffmannii''의 하악골이 몇 개 있는데, IRSNB R25와 IRSNB R27는 하악골에 골절 및 기타 병리가 있다. IRSNB R25는 여섯 번째 치아 소켓 근처의 완전 골절을 보존하며, 골절 주변에는 다량의 뼈 가골이 거의 치아 소켓을 과성장하고 있고, 다양한 골용해 공동, 농양 관, 삼차 신경 손상, 그리고 심각한 세균 감염을 나타내는 염증성 침식이 존재한다. IRSNB R27에는 두 개의 골절이 있는데, 하나는 거의 완전히 치유되었고 다른 하나는 인접한 치아가 부러진 열린 골절이다. 두 표본 모두 골절과 함께 심각한 세균 감염의 징후를 보이며, 일부 박테리아는 인접한 손상된 치아로 퍼져 충치를 유발했을 수 있다.[99]
2006년, 슐프(Schulp)와 동료들은 ''M. hoffmannii''의 사각형 뼈에 대한 연구를 발표했는데, 여기에는 여러 개의 부자연스러운 개구부와 조직 파괴가 있었다. 이는 패혈성 관절염으로 시작되어 심각한 골수염으로 진행되었을 가능성이 높다. 감염과 그에 따른 치유 과정은 몇 달 동안 진행되었을 수 있으며, 이로 인해 모사사우루스는 턱 사용 능력이 심각하게 저하되고 호흡에도 영향을 받았을 수 있다. 슐프와 동료들은 이 개체가 턱 사용을 최소화하기 위해 오징어와 같은 부드러운 먹이를 통째로 삼키는 방식으로 먹이를 찾았을 것이라고 추측했다.[100]
무혈관성 괴사는 ''M. lemonnieri''와 ''M. conodon''의 모든 검사 표본에서 존재하는 것으로 보고되었다.[61][101][102] 2005년 연구에서 로스차일드(Rothschild)와 마틴(Martin)은 모사사우루스 척추의 3-17%가 이 질환에 영향을 받았다고 관찰했다.[101] 무혈관성 괴사는 감압병의 일반적인 결과이며, 잦은 잠수 여행 중에 흡입된 공기의 감압으로 인한 질소 기포의 형성이나 반복적인 잠수와 짧은 호흡 간격으로 인한 뼈 손상을 포함한다. 이는 두 ''모사사우루스'' 종이 습관적인 심해 잠수부이거나 반복적인 잠수부였을 수 있음을 나타낸다.[102]
꼬리 척추의 부자연스러운 융합은 ''모사사우루스''에서 기록되었으며, 이는 외상이나 질병으로 인한 손상 후 뼈가 골화될 때 발생한다. 2015년 연구에서는 15개의 ''모사사우루스'' 표본을 조사한 결과, 그 중 세 개에서 꼬리 척추 융합 사례가 발견되었다.[103] 이 사례 중 두 개는 융합 부위 주변에 불규칙한 표면 변형을 보였는데, 이는 척추 정맥동의 배출로 인해 발생했으며, 뼈 감염의 징후이다. 세 번째 사례는 관절염의 한 형태로 결정되었다.[103]
''모사사우루스''는 오늘날 대부분의 현대 포유류처럼 태생(새끼를 낳는)이었을 가능성이 높다. ''모사사우루스'' 자체의 태생에 대한 증거는 없지만, 다른 여러 모사사우루스류에서 알려져 있다.[104] 어린 ''모사사우루스'' 및 관련 속의 뼈에 대한 미세 해부학적 연구에 따르면 그들의 뼈 구조는 성인과 유사하며, 이는 ''모사사우루스''가 조숙성이었다는 것을 의미한다. 즉, 그들은 이미 효율적인 수영 선수였으며 매우 어린 나이에 개방된 물에서 완전히 기능적인 생활 방식을 살았고 새끼를 키울 보육 공간이 필요하지 않았다.[106][104] 유럽과 사우스다코타의 일부 지역에서는 어린 ''M. hoffmannii'', ''M. missouriensis'' 및/또는 ''M. lemonnieri''의 농축된 집합체가 발견되었는데, 이 지역은 모두 얕은 해양 퇴적층으로, 어린 ''모사사우루스''가 여전히 얕은 물에서 살았을 수 있음을 시사한다.[107]
''모사사우루스''는 다양한 깊이의 원해 해양 서식지에 서식했다. 알려진 ''모사사우루스'' 화석은 전형적으로 백악기 동안 연안 서식지를 나타내는 퇴적층에서 발견되었으며, 일부 화석은 더 깊은 물의 퇴적층에서 나왔다.[93][133]
δ13C는 해양 동물의 먹이 서식지와 상관관계가 있으며, 동위 원소 수준은 서식지가 해안선에서 멀어질수록 고갈되므로 일부 과학자들은 동위 원소 수준을 서식지 선호도의 지표로 해석했다. 여러 ''모사사우루스'' 표본을 포함하는 별도의 연구에서 치아 에나멜의 일관되게 낮은 δ13C 수준이 나타났으며, 이는 ''모사사우루스''가 더 먼 해상 또는 개방 해역에서 먹이를 먹었음을 나타낸다.[93][133] 2014년 연구에서는 앨라배마의 ''M. hoffmannii'' 및 ''모사사우루스'' sp. 화석의 네오디뮴, 가돌리늄, 이터븀의 농도 비율을 조사했다. 희토류 원소 비율은 대부분의 조사된 ''모사사우루스'' 화석에서 매우 일관되게 나타났으며, 보다 깊은 해양 깊이의 해상 서식지를 나타내는 비율로 클러스터화되었다.[133]
6. 1. 1. 테티스 해
마스트리흐트절 동안 테티스 해는 현재의 유럽, 아프리카, 중동에 위치했으며, 북부와 남부 테티스 해역으로 나뉘었다. 지리적으로 북부 테티스 해역은 30–40°N의 고위도 주변에 위치하여 온대 기후를 보였고, M. 호프만니(M. hoffmannii)와 M. 레모니에리(M. lemonnieri)가 지배적인 종이었다. 특히 벨기에와 같은 특정 지역에서는 M. 레모니에리가 다른 대형 모사사우루스보다 훨씬 많이 발견되었다.[82] 그 외에도 할리사우루스, 플리오플라테카르푸스, 플라테카르푸스와 같은 소형 모사사우루스, 껍질을 부수는 카리노덴스, 타일로사우루스 베르나르디 및 4종의 프로그나토돈 등이 서식했다. 알로플레우로돈 호프만니 및 글립토켈로네 수이케르부이키와 같은 바다거북과 불확실한 엘라스모사우루스를 포함한 다른 해양 파충류도 가끔 발견되었다.[109]북부 테티스 해역의 뉴저지 지역은 유럽과 매우 유사한 해양 파충류 군집을 보였지만, M. 레모니에리, 카리노덴스, 타일로사우루스 및 특정 종의 할리사우루스와 프로그나토돈은 발견되지 않았다. 대신 M. 코노돈(M. conodon), 할리사우루스 플라티스폰딜루스 및 프로그나토돈 라팍스가 독점적으로 나타났다.[109] 스쿠알리코락스, 크레탈람나, 세라톨람나 및 모래상어와 같은 다양한 상어,[112] 키몰리크티스, 칼날 이빨 청어 엔코두스, 황새치와 같은 프로토스피라에나와 같은 경골어류도 서식했다.[109][113]

남부 테티스 해역은 20°N과 20°S 사이의 적도를 따라 위치하여 더 따뜻한 열대 기후를 보였다. 아프리카와 아라비아의 크레이톤과 접하고 레반트와 브라질까지 뻗어 있는 해저는 광대한 얕은 해양 환경을 제공했으며, 모사사우루스와 바다 목 옆 거북이 지배했다. 이 중 글로비덴스 포스파티쿠스가 특징적인 종이었다. 아프리카 및 아라비아 지역에서는 할리사우루스 아람보르기와 플라테카르푸스 프티코돈[109]이 글로비덴스와 함께 흔한 모사사우루스였다.[109] M. 베아우게이(M. beaugei)는 모로코와 브라질에 분포했으며, 시리아에서 발견된 이빨은 M. 레모니에리의 존재 가능성을 시사했지만, M. 호프만니도 해당 구역 전체에 어느 정도 존재했다.[58][109] 그 외 수수께끼의 고로니오사우루스, 껍질을 부수는 이그다마노사우루스 및 카리노덴스, 에레미아사우루스, 4종의 프로그나토돈, 다양한 종의 할리사우루스, 해양 모니터 도마뱀 파키바라누스 및 바다뱀 팔라에오피스 등이 서식했다. 모로코의 자라파사우라를 제외하면, 플레시오사우루스는 희귀했다. 엔코두스 및 스트라토두스와 같은 경골어류와 다양한 상어가 남부 테티스 해역 전체에서 흔하게 발견되었다.[109]
6. 1. 2. 서부 내륙 해로

''모사사우루스''의 가장 초기 화석 중 다수는 캄파니아절 북아메리카에서 발견되었으며, 여기에는 현재 미국 중부와 캐나다를 관통하여 북극해와 현대의 멕시코만을 연결했던 서부 내륙 해로도 포함된다.[115] 이 해로는 얕아서 최대 깊이가 약 800m에 달했다.[115] 인접한 아팔라치아와 라라미디아 대륙에서 유입된 막대한 양의 퇴적물, 북극해의 담수, 남쪽의 따뜻한 염분 테티스 해의 혼합은 영양분이 풍부한 심층수를 형성하여 풍부한 해양 생물을 지원했다.[116][117][118]
이 지역은 기후와 동물상 구조가 다른 두 개의 내륙 아(亞)주(Subprovince)로 나뉘었으며, 경계는 현대의 캔자스주에 있었다. 북부 내륙 아주는 서늘한 온대 기후였고, 남부 내륙 아주는 따뜻한 온대에서 아열대 기후였다.[110] ''M. missouriensis''와 ''M. conodon''이 7950만 년 전에 나타나면서 서부 내륙 해로의 해양 생태계는 근본적으로 재구성되었다.[119] 니오브라라 시대로 알려진 동물상 단계에서, 두 아주의 동물상 구조는 네이싱크 시대보다 훨씬 다양했다.[119][110][120]
앨라배마를 포함하는 남부 내륙 아주에서는 ''크레톡시리나(Cretoxyrhina)''와 같은 상어와 ''클리다스테스''(Clidastes), ''틸로사우루스''(Tylosaurus), ''글로비덴스''(Globidens), ''할리사우루스''(Halisaurus), ''플라테카르푸스''(Platecarpus)를 포함한 대부분의 주요 속들이 ''모사사우루스''(Mosasaurus)로 대체되었다.[119][121] 네이싱크 시대 동안 ''모사사우루스''(Mosasaurus)는 전체 지역을 지배하며, 전체 모사사우루스 다양성의 약 2/3를 차지했고 ''플리오플라테카르푸스''(Plioplatecarpus)와 ''프로그나토돈''(Prognathodon)이 나머지 1/3을 차지했다. 북부 내륙 아주에서도 ''플라테카르푸스''(Platecarpus)와 같은 모사사우루스가 사라지고 ''모사사우루스''(Mosasaurus)와 ''플리오플라테카르푸스''(Plioplatecarpus)로 대체되는 등 재구성이 있었다.[119] ''틸로사우루스''(Tylosaurus),[122] ''크레톡시리나''(Cretoxyrhina),[123] 헤스페르오르니테스,[124] ''테르미노나토르(Terminonatator)''와 같은 엘라스모사우루스를 포함한 장경룡과 ''돌리코린코프스(Dolichorhynchops)''와 같은 폴리코틸리드는[125][126] 캄파니아절 말까지 존재했으며, 이후 서부 내륙 해로는 북쪽에서부터 물러나기 시작했다.[116] ''모사사우루스''(Mosasaurus)는 백악기 말 네이싱크 시대가 끝날 때까지 해로에서 지배적인 속으로 남아있었다.[119]
당시 동물상에는 ''프로토스테가(Protostega)''[121]와 ''아르켈론(Archelon)''과 같은 바다거북;[127] ''밥토르니스(Baptornis)'',[124] ''익티오르니스(Ichthyornis)'', ''할리모르니스(Halimornis)''를 포함한 다양한 바닷새; 청상아리 상어인 ''크레탈람나''(Cretalamna), ''스콰리코락스''(Squalicorax), ''프세우도코락스(Pseudocorax)'', ''세라톨람나''(Serratolamna), 고블린 상어인 ''스카파노린쿠스(Scapanorhynchus)'', 백상아리인 ''오돈타스피스(Odontaspis)'', 톱상어류인 ''이시리자(Ischyrhiza)''와 같은 상어; ''엔코두스''(Enchodus), ''프로토스피라에나''(Protosphyraena), ''스트라토두스''(Stratodus), 익티오데크테스과의 ''크시팍티누스(Xiphactinus)'' 및 ''사우로돈(Saurodon)''과 같은 경골어류가 있었다.[121][128]
6. 1. 3. 남극
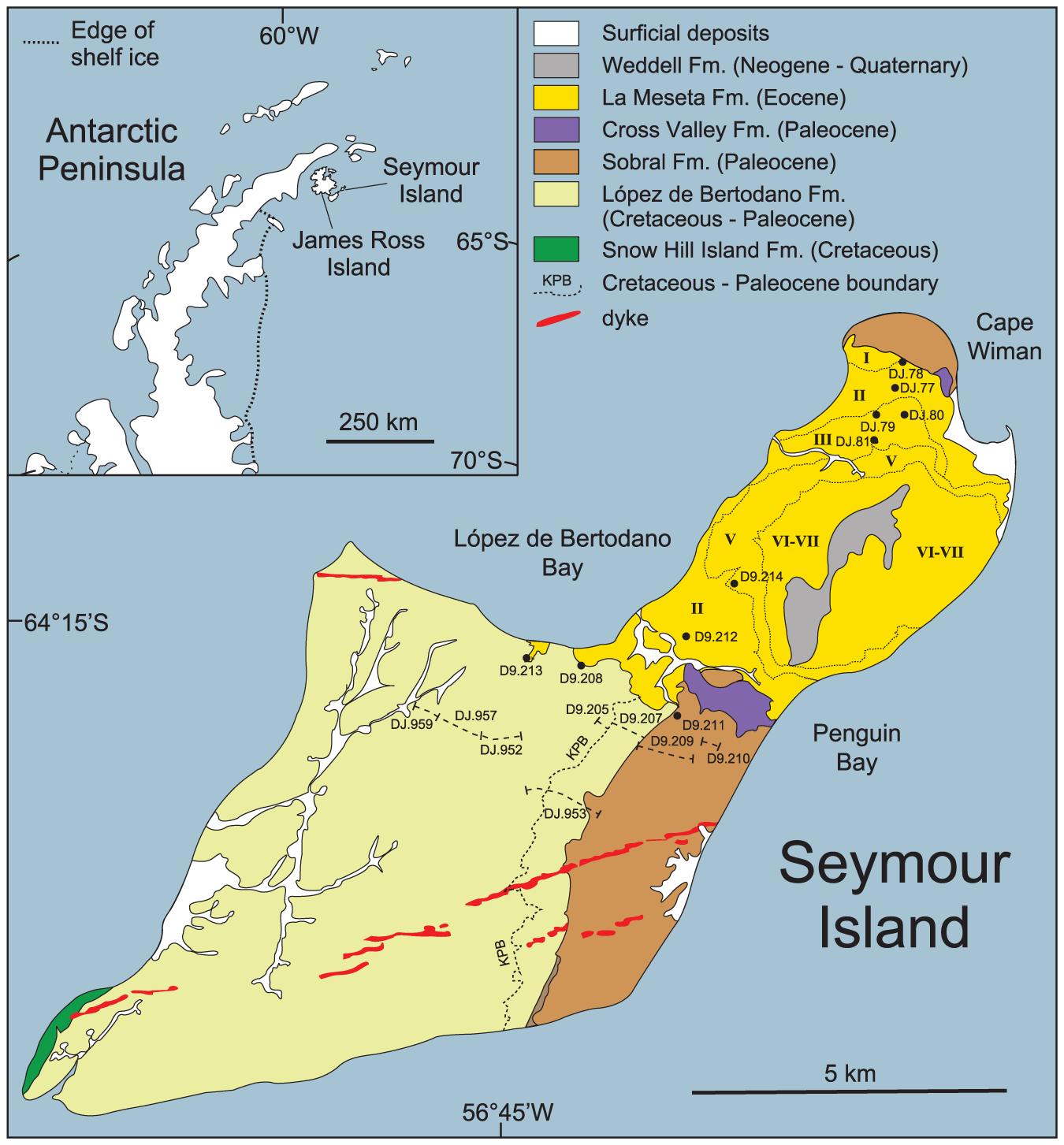
''모사사우루스''는 남극 반도의 후기 마스트리흐트절 지층, 특히 시모어 섬의 로페스 데 베르토다노 층에서 알려져 있다.[90] 이 지역은 극권에서 남쪽으로 약 65° 떨어진 곳에 위치했으며,[111] 중간 깊이의 수온은 평균 6°C 정도였고, 해수면 온도는 어는점 아래로 내려가 때때로 해빙이 형성되기도 했다.[89][129]
마스트리흐트절 남극 대륙에서 ''모사사우루스''는 가장 다양한 모사사우루스였던 것으로 보인다. 최소 두 종의 ''모사사우루스''가 기술되었지만, 화석 파편이 많고 표본이 개방 명명법으로 설명되어 정확한 종 수는 알 수 없다. 이 종들에는 ''M. lemonnieri''와 유사한 종과 ''M. hoffmannii''와 밀접하게 관련된 것으로 보이는 종이 포함된다.[90] ''M. sp.''도 기술되었으나, 이 표본은 ''모아나사우루스''를 나타낼 가능성이 있으며, 이는 해당 속에 대한 개정 결과에 따라 달라진다.[35]
''플리오플라테카르푸스'', 모사사우루스아과의 ''모아나사우루스'' 및 ''리오돈'',[90] 그리고 ''카이카이필루''를 포함하여 최소 4개의 다른 모사사우루스 속이 남극 대륙에서 보고되었다. 하지만 이 속들 중 일부는 주로 이빨 화석에 기반하고 있어 유효성에 대한 논란이 있다.[130] ''프로그나토돈''과 ''글로비덴스''도 분포 경향에 따라 존재했을 것으로 예상되지만, 확실한 화석은 아직 발견되지 않았다.[90]
다른 남극 해양 파충류로는 ''아리스토넥테스''와 다른 불확정 엘라스모사우루스과를 포함한 엘라스모사우루스과 플레시오사우루스가 있었다.[131] 로페스 데 베르토다노 층의 어류 군집은 ''엔코두스''와 어룡류가 주를 이루었다.[132]
6. 2. 서식지 선호도
''모사사우루스'' 화석은 일반적으로 백악기 동안 연안 서식지를 나타내는 퇴적층에서 발견되었지만, 일부는 더 깊은 물의 퇴적층에서도 발견되었다.[93][133] 링엄솔리아(Lingham-Soliar, 1995)는 네덜란드의 마스트리히트기 퇴적층이 40m 깊이의 연안 해역을 나타내며, 기온 변화와 풍부한 해양 생물이 특징이었다고 설명했다. 그럼에도 ''M. hoffmannii''의 형태는 표해성 표면 생활 방식에 적응되어 있었다.[48]δ13C는 해양 동물의 먹이 서식지와 상관관계가 있으며, 해안선에서 멀어질수록 동위 원소 수준이 고갈된다. 일부 과학자들은 이를 서식지 선호도의 지표로 해석했다. 여러 ''모사사우루스'' 표본의 치아 에나멜에서 일관되게 낮은 δ13C 수준이 나타났는데, 이는 ''모사사우루스''가 더 먼 해상 또는 개방 해역에서 먹이를 먹었음을 시사한다.[93][133]
T. 린 해럴 주니어(T. Lynn Harrell Jr.)와 알베르토 페레스우에르타(Alberto Perez-Huerta)의 2014년 연구에서는 앨라배마의 ''M. hoffmannii'' 및 ''모사사우루스'' 종 화석, 데모폴리스 백악질(Demopolis Chalk) 및 호너스타운 층(Hornerstown Formation)의 네오디뮴, 가돌리늄, 이터븀 농도 비율을 조사했다. 희토류 원소 비율은 대부분 조사된 ''모사사우루스'' 화석에서 매우 일관되게 나타났으며, 50m보다 깊은 해양 깊이의 해상 서식지를 선호했음을 나타냈다.[133]
6. 3. 종간 경쟁
''모사사우루스''는 타일로사우루스아과와 ''프로그나토돈''과 같이 최상위 포식자로 여겨지는 다른 대형 포식성 모사사우루스들과 함께 살았다.[48][61] 마스트리흐트절에 유일하게 생존한 속인 ''틸로사우루스 베르나르디''는 길이가 12.2m에 달했고[134], ''P. saturator''와 같이 공존했던 ''프로그나토돈''의 가장 큰 종은 12m를 넘었다.[61] 이 세 종류의 모사사우루스는 해양 파충류와 같은 비슷한 동물을 잡아먹었다.[20][48][61]2013년 Schulp와 동료들이 발표한 연구에서는 ''M. hoffmannii''와 ''P. saturator''와 같은 모사사우루스가 δ13C 분석을 통해 동일 지역에서 어떻게 공존할 수 있었는지를 조사했다. 과학자들은 동위원소 값의 차이가 생활 방식, 식단, 서식지 선호도와 같은 여러 환경적 요인의 영향을 받기 때문에 자원 분할 수준을 설명하는 데 도움이 될 수 있다고 보았다. 마스트리히트 지층의 마스트리흐트절 시대 ''M. hoffmannii''와 ''P. saturator''의 여러 치아에서 δ13C 수준을 비교한 결과, 특정 표본 간에 약간의 수렴이 있었지만, 두 종 간의 평균 δ13C 값은 달랐다. 이는 두 모사사우루스 속이 직접적인 경쟁 없이 공존하기 위해 서로 다른 서식지에서 먹이를 구하거나 서로 다른 특정 식단을 가졌을 가능성이 높은, 생태적 지위 분화의 한 징후이다. ''P. saturator''의 치아는 ''M. hoffmannii''의 치아보다 훨씬 튼튼하며, 특히 거북과 같은 단단한 먹이를 잡아먹는 데 적합했다. ''M. hoffmannii'' 또한 거북을 잡아먹었지만, 이빨은 ''P. saturator''에게는 적합하지 않은 더 넓은 범위의 먹이를 처리하도록 만들어졌다.[61]
앨버타의 베어포 지층에서 ''모사사우루스''와 ''프로그나토돈'' 간의 생태적 지위 분화의 또 다른 사례가 Konishi와 동료들의 2014년 연구에서 나타났다. 이 연구는 위 내용물을 바탕으로 ''M. missouriensis''와 ''P. overtoni'' 사이의 식단 분할을 발견했다. ''P. overtoni''의 위 내용물에는 거북과 암모나이트가 포함되어 있어, 더 단단한 먹이에 특화된 식단을 보여주었다. 반대로, ''M. missouriensis''의 위 내용물은 물고기로 구성되어 있어, 더 부드러운 먹이에 특화된 식단을 나타냈다. 이러한 적응은 두 모사사우루스 사이의 자원 분할을 유지하는 데 도움이 되었을 것으로 추정된다.[20]
그럼에도 불구하고, 경쟁적 교류는 피할 수 없었던 것으로 보인다. ''모사사우루스''와 다른 대형 모사사우루스 종 간의 공격적인 종간 전투의 증거도 있다. 아성체 ''M. hoffmannii''의 화석 두개골에서 뇌 케이스에 골절이 발생한 거대한 집중적인 타격이 발견되었다. Lingham-Soliar (1998)는 이러한 골절의 형성이 조율된 공격의 특징을 보였고(사고나 화석화 손상이 아닌), 그러한 손상을 일으킬 수 있는 유일하게 알려진 공존 동물은 ''T. bernardi''였으며, 튼튼한 화살 모양의 길쭉한 주둥이를 사용했기 때문에 이 타격은 ''T. bernardi''에 의한 돌진 공격으로 인해 발생했다고 주장했다. 이러한 종류의 공격은 병코돌고래가 부리를 사용하여 레몬상어를 죽이거나 격퇴하는 방어 행동과 비교되었으며, ''T. bernardi''가 방심한 ''모사사우루스''를 매복하여 공격했을 것으로 추정된다.[135]
7. 멸종
백악기 말, 모사사우루스는 진화적 번성의 정점에 있었지만 갑작스럽게 멸종했다.[48] 후기 마스트리히트절 동안 전 세계 해수면이 낮아지면서 대륙붕 주변의 영양분이 풍부한 연안 서식지가 사라지고, 해류와 영양 순환 패턴이 변화하여 모사사우루스가 살 수 있는 환경이 줄어들었다.[136][137] 모사사우루스는 더 넓은 해역에서 새로운 서식지를 찾아 적응했다.[136][137]
''모사사우루스''의 마지막 화석은 백악기-고생대 경계 (K-Pg 경계)까지 발견되며, 여기에는 ''M. 호프만니''와 불확실한 종들이 포함된다. 이 속의 멸종은 비조류 공룡을 멸종시킨 백악기-고생대 대량 절멸의 결과일 가능성이 높다. ''모사사우루스'' 화석은 매스트리흐트 지층, 터키의 다부틀라르 지층, 아르헨티나의 자구엘 지층, 덴마크의 스테븐스 클린트, 시모어 섬, 미주리 등에서 경계선 아래 15m 미만에서 발견되었다.[138]
''M. 호프만니'' 화석은 미주리 남동부의 K-Pg 경계선 내, 고신세 클레이턴 지층과 백악기 올크릭 지층 사이에서 발견되었다. 이 지층의 척추뼈 화석은 사후 골절과 함께 발견되었다. 이 지층은 쓰나마이트로 퇴적되었을 가능성이 있으며, "백악기 칵테일 퇴적물"이라고도 불린다. 이는 칙술루브 소행성 충돌로 인한 대재앙적인 지진, 지질 교란, 메가 허리케인, 거대 쓰나미 등의 복합적인 결과로 형성되었으며, 이 충돌이 K-Pg 멸종 사건을 일으켰다.[136] 물리적 파괴 외에도, 충돌로 인해 햇빛이 차단되어[139] 해양 먹이 사슬이 붕괴되었다.[136] 깊은 물속으로 피신하여 즉각적인 대격변에서 살아남은 ''모사사우루스''는 먹이 부족으로 굶어 죽었을 것이다.[136]
''모사사우루스'' sp. 화석 중 일부는 호너스타운 지층에서 발견되는데, 이 지층은 일반적으로 마스트리히트절 직후인 고신세 다니안으로 연대가 측정된다. 이 화석은 메인 화석층으로 알려진 호너스타운 지층 기저부의 독특한 화석 밀집 지층에서 ''스쿠알리코락스'', ''엔코두스'', 다양한 암모나이트 화석과 함께 발견되었다. 이것이 ''모사사우루스''와 관련 동물군이 K-Pg 멸종에서 살아남았음을 의미하지는 않는다. 한 가지 가설은 이 화석들이 이전 백악기 퇴적물에서 유래하여 초기 퇴적 과정에서 고신세 지층으로 재가공되었다는 것이다. 재가공의 증거는 일반적으로 재퇴적 시기에 노출되어 침식된 화석에서 나타난다. 메인 화석층의 많은 ''모사사우루스'' 화석은 마모되고 닳은 고립된 뼈로 구성되어 있지만, 이 지층에서는 보존 상태가 더 좋은 ''모사사우루스'' 잔해도 발견되었다. 또 다른 설명은 메인 화석층이 백악기 퇴적물에서 기원하고 선별된 저퇴적 환경을 가진 마스트리히트기 시간 평균 재작업 퇴적물이라는 것이다. 세 번째 가설은 이 층이 쓰나미의 강력한 충격으로 밀려난 백악기 퇴적물의 지연 퇴적물이며, 이후 신생대 화석으로 채워졌다는 것이다.[2]
참조
[1]
서적
The Geologic Time Scale
Elsevier
[2]
논문
Recent mosasaur discoveries from New Jersey and Delaware, USA: stratigraphy, taphonomy and implications for mosasaur extinction
[3]
논문
Paleoecology of the Delaware Valley region, Part II: Cretaceous to Quaternary
https://docs.google.[...]
[4]
논문
Glauconite Composition and Morphology, Shocked Quartz, and the Origin of the Cretaceous(?) Main Fossiliferous Layer (MFL) in Southern New Jersey, U.S.A.
[5]
서적
Cretaceous Reptiles of the United States
https://books.google[...]
Smithsonian Contributions to Knowledge
[6]
논문
A mosasaur from the Maastrichtian Fox Hills Formation of the northern Western Interior Seaway of the United States and the synonymy of ''Mosasaurus maximus'' with ''Mosasaurus hoffmanni'' (Reptilia: Mosasauridae)
[7]
논문
Beschrijving der beenderen van den kop van eenen visch, gevonden in den St Pietersberg bij Maastricht, en geplaatst in Teylers Museum
https://www.delpher.[...]
[8]
웹사이트
"Mosasaurus hoffmanni''-The First Discovery of a Mosasaur?"
http://oceansofkansa[...]
2010-05-14
[9]
뉴스
Datum vondst mosasaurus ontdekt: in oktober 1778
https://www.1limburg[...]
2015-09-21
[10]
논문
Conjectures relative to the petrifactions found in St. Peter's Mountain near Maestricht
[11]
서적
Napoleon's Legacy: The Rise of National Museums in Europe, 1794-1830
G+H Verlag
[12]
서적
Histoire naturelle de la montagne de Saint-Pierre de Maestricht
https://www.biodiver[...]
H. J. Jansen
[13]
논문
A new look at Faujas de Saint-Fond's fantastic story on the provenance and acquisition of the type specimen of ''Mosasaurus hoffmanni'' Mantell, 1829
[14]
논문
Sur les ossements fossiles de la montagne de St. Pierre, à Maëstricht
https://www.biodiver[...]
[15]
논문
Sur le grand animal fossile des carrières de Maestricht
https://www.biodiver[...]
[16]
서적
Dutch Pioneers of the Earth Sciences
Royal Netherlands Academy of Arts and Sciences
[17]
논문
The roles played by museums, collections and collectors in the early history of reptile palaeontology
[18]
서적
An Introduction to the Study of Fossil Organic Remains
https://books.google[...]
Printed for the Author
[19]
논문
A tabular arrangement of the organic remains of the county of Sussex
https://upload.wikim[...]
[20]
논문
A small, exquisitely preserved specimen of ''Mosasaurus missouriensis'' (Squamata, Mosasauridae) from the upper Campanian of the Bearpaw Formation, western Canada, and the first stomach contents for the genus
[21]
웹사이트
The Goldfuss Mosasaur
http://oceansofkansa[...]
2013-10-21
[22]
서적
Sea Dragons: Predators of the Prehistoric Oceans
https://archive.org/[...]
University Press of Kansas
[23]
논문
The largest mosasaur (Squamata: Mosasauridae) from the Missouri River area (Late Cretaceous; Pierre Shale Group) of South Dakota and its relationship to Lewis and Clark
https://www.montclai[...]
[24]
서적
Lethaea Geognostica Oder Abbildungen und Beschreibungen Der für die Gebirgs-Formationen bezeichnendsten Versteinerungen
https://babel.hathit[...]
Stuttgart
[25]
논문
On the Remains of Extinct Reptiles of the genera ''Mosasaurus'' and ''Geosaurus'' found in the secondary formation of New-Jersey; and on the occurrence of the substance recently named Coprolite by Dr. Buckland, in the same locality
https://www.biodiver[...]
[26]
논문
Notice of the Discovery of the Remains of the ''Ichthyosaurus'' in Missouri, N. A.
[27]
논문
Notice of the discovery of ''Basilosaurus'' and ''Batrachiosaurus''
https://www.biodiver[...]
[28]
논문
Der Schädelbau des ''Mosasaurus'', durch Beschreibung einer neuen Art dieser Gattung erläutert
https://books.google[...]
[29]
논문
(Letter directed to Professor Bronn)
https://www.biodiver[...]
[30]
논문
List of extinct Vertebrata; the remains of which have been discovered in the region of the Missouri river: with remarks on their geological age
https://www.biodiver[...]
[31]
논문
A new ''Clidastes'' from New Jersey
https://www.biodiver[...]
[32]
논문
Rare marine reptiles from the Cretaceous of New Jersey
[33]
웹사이트
Mosasauridae Translation and Pronunciation Guide
http://www.dinosauri[...]
[34]
논문
Première note sur les mosasauriens de Mesvin
https://www.biodiver[...]
[35]
논문
A new Plotosaurini mosasaur skull from the upper Maastrichtian of Antarctica. Plotosaurini paleogeographic occurrences
https://rid.unrn.edu[...]
[36]
서적
First Mosasaur Meeting, Maastricht, 8-12 May 2004, Abstract book and Field guide
Natuurhistorisch Museum Maastricht
[37]
간행물
Dental variability and distinguishability in ''Mosasaurus lemonnieri'' (Mosasauridae) from the Campanian and Maastrichtian of Belgium, and implications for taxonomic assessments of mosasaurid dentitions
[38]
서적
Les vertébrés fossiles des gisements de phosphates (Maroc–Algérie–Tunisie)
http://mmtk.ginras.r[...]
Typographie Firmin-Didot
[39]
간행물
Reptilian assemblages from the latest Cretaceous – Palaeogene phosphates of Morocco: from Arambourg to present time
https://www.research[...]
[40]
간행물
Note sur le mosasaure
https://www.biodiver[...]
[41]
간행물
Hermann Schlegel's investigation of the Maastricht mosasaurs
https://www.research[...]
[42]
웹사이트
The science of the Crystal Palace Dinosaurs, part 2: ''Teleosaurus'', pterosaurs and ''Mosasaurus''
http://markwitton-co[...]
2019-05-17
[43]
웹사이트
The world's first dinosaur park: what the Victorians got right and wrong
https://www.nhm.ac.u[...]
[44]
간행물
Convergent Evolution in Aquatic Tetrapods: Insights from an Exceptional Fossil Mosasaur
[45]
간행물
A challenge to categories: "What, if anything, is a mosasaur?"
[46]
서적
Systematics and Morphology of American Mosasaurs
https://www.biodiver[...]
Bulletin of the Peabody Museum of Natural History
[47]
간행물
Giant ''Mosasaurus hoffmanni'' (Squamata, Mosasauridae) from the Late Cretaceous (Maastrichtian) of Penza, Russia
https://www.zin.ru/j[...]
[48]
간행물
Anatomy and functional morphology of the largest marine reptile known, ''Mosasaurus hoffmanni'' (Mosasauridae, Reptilia) from the Upper Cretaceous, Upper Maastrichtian of The Netherlands
https://royalsociety[...]
[49]
간행물
Lepidosaurian diversity in the Mesozoic–Palaeogene: the potential roles of sampling biases and environmental drivers
[50]
간행물
A giant mosasaur (Reptilia, Squamata) with an unusually twisted dentition from the Argille Scagliose Complex (late Campanian) of Northern Italy
https://www.disva.un[...]
[51]
간행물
Cautionary tales on the use of proxies to estimate body size and form of extinct animals
[52]
conference
Mosasaurs—how large did they really get?
https://www.research[...]
5th Triennial Mosasaur Meeting—A Global Perspective on Mesozoic Marine Amniotes
2016
[53]
간행물
Nouvelle note sur l'osteologie des mosasauriens
https://www.biodiver[...]
[54]
간행물
Physical drivers of mosasaur evolution
https://docentes.fct[...]
[55]
간행물
Unusual histology and morphology of the ribs of mosasaurs (Squamata)
[56]
간행물
Mosasaurids (Squamata) from the Maastrichtian Phosphates of Morocco: Biodiversity, palaeobiogeography and palaeoecology based on tooth morphoguilds
https://www.research[...]
[57]
간행물
Osteology and taxonomy of ''Mosasaurus conodon'' Cope 1881 from the Late Cretaceous of North America
https://www.research[...]
[58]
간행물
Rediagnosis and redescription of ''Mosasaurus hoffmannii'' (Squamata: Mosasauridae) and an assessment of species assigned to the genus ''Mosasaurus''
https://www.research[...]
[59]
웹사이트
Mosasaur brain
http://oceansofkansa[...]
2010-01-01
[60]
간행물
''Mosasaurus beaugei'' Arambourg, 1952 (Squamata, Mosasauridae) from the Late Cretaceous phosphates of Morocco
https://www.academia[...]
[61]
간행물
On diving and diet: resource partitioning in type-Maastrichtian mosasaurs
[62]
간행물
New mosasaur material from the Maastrichtian of Angola, with notes on the phylogeny, distribution, and paleoecology of the genus ''Prognathodon''
https://docentes.fct[...]
[63]
간행물
Of German princes and North American rivers: Harlan's lost mosasaur snout rediscovered
[64]
간행물
A New Addition to the Cretaceous Seaway of North Dakota
https://www.dmr.nd.g[...]
North Dakota Geological Society
[65]
웹사이트
Samuel Wilson's ''Mosasaurus horridus''
http://oceansofkansa[...]
2009-03-26
[66]
간행물
Ontogeny, anatomy and attachment of the dentition in mosasaurs (Mosasauridae: Squamata)
[67]
간행물
Dental microstructure and geochemistry of ''Mosasaurus hoffmanni'' (Squamata: Mosasauridae) from the Late Cretaceous of Turkey
[68]
간행물
Landlubbers to leviathans: evolution of swimming in mosasaurine mosasaurs
[69]
간행물
Soft tissue preservation in a fossil marine lizard with a bilobed tail fin
[70]
간행물
Transatlantic latest Cretaceous mosasaurs (Reptilia, Lacertilia) from the Maastrichtian type area and New Jersey
[71]
웹사이트
Article 8. What constitutes published work
https://code.iczn.or[...]
2021-07-16
[72]
웹사이트
"''Notes on Early Mesozoic Theropods'' and the future of zoological nomenclature"
https://svpow.com/20[...]
2010-06-08
[73]
PhD
A re-assessment of the genus Mosasaurus (Squamata: Mosasauridae)
https://era.library.[...]
University of Alberta
[74]
논문
Current State of the Problems in the Phylogeny of Squamate Reptiles (Squamata, Reptilia)
[75]
논문
A new mosasaurine from the Maastrichtian (Upper Cretaceous) phosphates of Morocco and its implications for mosasaurine systematics
https://www.research[...]
[76]
논문
Reassessing Mosasaurini based on a systematic revision of ''Mosasaurus''
https://www.research[...]
[77]
서적
Ancient Marine Reptiles
Academic Press
[78]
논문
Inferring 'weak spots' in phylogenetic trees: application to mosasauroid nomenclature
[79]
논문
Mosasauroid phylogeny under multiple phylogenetic methods provides new insights on the evolution of aquatic adaptations in the group
[80]
논문
Redescription of ''Prognathodon lutugini'' (Squamata, Mosasauridae)
http://www.zin.ru/Jo[...]
[81]
논문
Phylogeny And Systematics Of Squamata (Reptilia) Based On Morphology
[82]
논문
The mosasaur ''Mosasaurus lemonnieri'' (Lepidosauromorpha, Squamata) from the Upper Cretaceous of Belgium and The Netherlands
[83]
웹사이트
Inertial feeding in reptiles: the role of skull mass reduction
https://sicb.burkcli[...]
[84]
논문
Locomotion in mosasaurs
[85]
논문
Microanatomical and Histological Features in the Long Bones of Mosasaurine Mosasaurs (Reptilia, Squamata) – Implications for Aquatic Adaptation and Growth Rates
[86]
MS
Osteohistology And Skeletochronology Of an Ontogenetic Series Of ''Clidastes'' (Squamata: Mosasauridae): Growth And Metabolism In Basal Mosasaurids
https://scholars.fhs[...]
Fort Hays State University
[87]
논문
Seasonal reproductive endothermy in tegu lizards
[88]
논문
Endothermic mosasaurs? Possible thermoregulation of Late Cretaceous mosasaurs (Reptilia, Squamata) indicated by stable oxygen isotopes in fossil bioapatite in comparison with coeval marine fish and pelagic seabirds
[89]
논문
Late Cretaceous winter sea ice in Antarctica?
http://nora.nerc.ac.[...]
[90]
논문
Biostratigraphy of the Mosasauridae (Reptilia) from the Cretaceous of Antarctica
[91]
논문
Campanian and Maastrichtian mosasaurs from Antarctic Peninsula and Patagonia, Argentina
[92]
논문
A new halisaurine mosasaur (Squamata: Halisaurinae) from Japan: the first record in the western Pacific realm and the first documented insights into binocular vision in mosasaurs
https://www.research[...]
[93]
PhD
Investigating Holocene climate change on the northern Channel Islands and Cretaceous mosasaur ecology using stable isotopes
https://www.proquest[...]
Southern Methodist University
[94]
간행물
Gut contents of a subadult individual of ''Mosasaurus hoffmannii'' Mantell, 1829 from the Maastrichtian type area (the Netherlands) hint at the species’ dietary preferences
https://www.nhmmaast[...]
7th Triennial Mosasaur Meeting—A Global Perspective on Mesozoic Marine Amniotes
[95]
논문
Mosasaur Predation on Upper Cretaceous Nautiloids and Ammonites from the United States Pacific Coast
http://doc.rero.ch/r[...]
[96]
논문
Direct evidence of aggressive intraspecific competition in ''Mosasaurus conodon'' (Mosasauridae:Squamata)
[97]
뉴스
Ancient sea monster battle revealed in unusual fossil
https://www.science.[...]
2016-10-26
[98]
웹사이트
Anything Mosasaur
https://konishta.wix[...]
[99]
논문
Palaeopathology and injury in the extinct mosasaurs (Lepidosauromorpha, Squamata) and implications for modern reptiles
http://doc.rero.ch/r[...]
[100]
논문
Chronic bone infection in the jaw of ''Mosasaurus hoffmanni'' (Squamata)
http://www.dinosauri[...]
[101]
논문
Mosasaur ascending: the phytogeny of bends
[102]
논문
Frequency of decompression illness among recent and extinct mammals and "reptiles": a review
[103]
논문
Co-Ossification of Vertebrae in Mosasaurs (Squamata, Mosasauridae); Evidence of Habitat Interactions and Susceptibility to Bone Disease
https://www.research[...]
[104]
논문
Pelagic neonatal fossils support viviparity and precocial life history of Cretaceous mosasaurs
[105]
논문
The first direct evidence of live birth in Mosasauridae (Squamata): Exceptional preservation in Cretaceous Pierre Shale of South Dakota
[106]
논문
What vertebral microanatomy reveals about the ecology of juvenile mosasaurs (Reptilia, Squamata)
[107]
논문
Juvenile marine reptiles from the Late Cretaceous of the Antarctic peninsula and their relationships to other such occurrences in central South Dakota and Belgium
https://sdaos.org/wp[...]
[108]
논문
Occurrence of Mosasaurus hoffmannii Mantell, 1829 (Squamata, Mosasauridae) in the Maastrichtian Phosphates of Morocco
https://www.research[...]
[109]
논문
Maastrichtian marine reptiles of the Mediterranean Tethys: a palaeobiogeographical approach
[110]
논문
Paleobiogeography of the Cretaceous Western Interior Seaway of North America: the vertebrate evidence
[111]
논문
A cool temperate climate on the Antarctic Peninsula through the latest Cretaceous to early Paleogene
[112]
논문
Upper Maastrichtian shallow marine environments and neoselachian assemblages in North Iberian palaeomargin (Castilian Ramp, Spain)
[113]
논문
Ray-finned fishes (Osteichthyes, Actinopterygii) from the type Maastrichtian, the Netherlands and Belgium
https://www.research[...]
[114]
논문
A new species of longirostrine plioplatecarpine mosasaur (Squamata: Mosasauridae) from the Late Cretaceous of Morocco, with a re-evaluation of the problematic taxon'' 'Platecarpus' ptychodon''
[115]
서적
Earth System History
W.H. Freeman and Company
[116]
서적
Oceans of Kansas
Indiana University Press
[117]
논문
Paleoenvironment of the Western Interior Seaway inferred from δ18O and δ13C values of molluscs from the Cretaceous Bearpaw marine cyclothem
[118]
논문
Water mass characteristics in the Cenomanian US Western Interior seaway as indicated by stable isotopes of calcareous organisms
[119]
논문
Stratigraphic distribution and habitat segregation of mosasaurs in the Upper Cretaceous of western and central Alabama, with a historical review of Alabama mosasaur discoveries
https://www.research[...]
[120]
논문
A new species of ''Peritresius'' Leidy, 1856 (Testudines: Pan-Cheloniidae) from the Late Cretaceous (Campanian) of Alabama, USA, and the occurrence of the genus within the Mississippi Embayment of North America
[121]
논문
An Overview of Late Cretaceous Vertebrates from Alabama
https://www.research[...]
[122]
논문
A New Hypothesis of the Phylogenetic Relationships of the Tylosaurinae (Squamata: Mosasauroidea)
[123]
논문
A late Campanian euselachian assemblage from the Bearpaw Formation of Alberta, Canada: some notable range extensions
[124]
논문
''Baptornis'' sp. (Aves: Hesperornithiformes) from the Judith River Formation (Campanian) of Saskatchewan, Canada
[125]
논문
''Terminonatator ponteixensis'', a new elasmosaur (Reptilia; Sauropterygia) from the Upper Cretaceous of Saskatchewan
[126]
논문
A new polycotylid plesiosaur (Reptilia: Sauropterygia) from the Upper Cretaceous Bearpaw Formation in Saskatchewan, Canada
[127]
논문
Skeleton of the Rare Giant Sea Turtle, ''Archelon'', Recovered from the Cretaceous DeGrey Member of the Pierre Shale near Cooperstown, Griggs County, North Dakota
https://www.dmr.nd.g[...]
[128]
논문
Vertebrate Paleontology of the Pierre Shale and Fox Hills Formations (Late Campanian-Late Maastrichtian) of Badlands National Park, South Dakota
https://doc.rero.ch/[...]
[129]
논문
Extinction patterns, δ18 O trends, and magnetostratigraphy from a southern high-latitude Cretaceous–Paleogene section: Links with Deccan volcanism
https://authors.libr[...]
[130]
논문
''Kaikaifilu hervei'' gen. et sp. nov., a new large mosasaur (Squamata, Mosasauridae) from the upper Maastrichtian of Antarctica
[131]
논문
A new elasmosaurid from the upper Maastrichtian López de Bertodano Formation: new data on weddellonectian diversity
https://figshare.com[...]
[132]
논문
Before and after the K/Pg extinction in West Antarctica: New marine fish records from Marambio (Seymour) Island
[133]
논문
Habitat preference of mosasaurs indicated by rare earth element (REE) content of fossils from the Upper Cretaceous marine deposits of Alabama, New Jersey, and South Dakota (USA)
[134]
논문
The first record of ''Hainosaurus'' (Reptilia: Mosasauridae) from Sweden
http://doc.rero.ch/r[...]
[135]
논문
Unusual death of a Cretaceous giant
http://doc.rero.ch/r[...]
[136]
논문
Mosasaur (Reptilia, Squamata) material from the Cretaceous-Tertiary boundary interval in Missouri
[137]
웹사이트
GEOL 104 Lecture 38: The Cretaceous-Tertiary Extinction III: Not With a Bang, But a Whimper
https://www.geol.umd[...]
[138]
논문
The youngest ''in situ'' record to date of ''Mosasaurus hoffmannii'' (Squamata, Mosasauridae) from the Maastrichtian type area, The Netherlands
https://www.research[...]
[139]
논문
Global climate change driven by soot at the K-Pg boundary as the cause of the mass extinction
[140]
논문
Giant ''Mosasaurus hoffmanni'' (Squamata, Mosasauridae) from the Late Cretaceous (Maastrichtian) of Penza, Russia
https://www.zin.ru/j[...]
2016-06-26
[141]
논문
A giant mosasaur (Reptilia, Squamata) with an unusually twisted dentition from the Argille Scagliose Complex (late Campanian) of Northern Italy
https://www.scienced[...]
2014-05-01
[142]
서적
謎と不思議の生物史
"[[同文書院]]"
[143]
서적
恐竜博物図鑑
"[[新樹社]]"
[144]
간행물
"The roles played by museums, collections and collectors in the early history of reptile palaeontology."
Geological Society Special Publication 343
2010
[145]
간행물
"Beschrijving der beenderen van den kop van eenen visch, gevonden in den St Pietersberg bij Maastricht, en geplaatst in Teylers Museum"
1790
[146]
간행물
"Maastricht Cretaceous finds and Dutch pioneers in vertebrate palaeontology"
Royal Netherlands Academy of Arts and Sciences (KNAW), Amsterdam
2004
[147]
간행물
Histoire naturelle de la montagne de Saint-Pierre de Maëstricht
http://imgbase-scd-u[...]
1798
[148]
간행물
"Conjectures relative to the petrifactions found in St. Peter’s Mountain near Maestricht"
http://gallica.bnf.f[...]
1786
[149]
간행물
"Sur le grand animal fossile des carrières de Maestricht"
https://www.biodiver[...]
Parijs
1808
[150]
간행물
Fragmentum Maxillae superioris, lateris dextri capitis Physeteris incogniti ex Monte St. Petri, Traj. [ectum] ad Mosam
[151]
문서
A Faujas de Saint-Fond later (1798)
[152]
간행물
Mosasaurus hoffmanni: De lotgevallen van een type-exemplaar
1995
[153]
간행물
"Lettre de A.G. Camper à G. Cuvier sur les ossemens fossiles de la montagne de St. Pierre, à Maëstricht"
1800
[154]
웹사이트
ICZN Art. 33.3.1 plus example
http://www.nhm.ac.uk[...]
[155]
웹사이트
Mosasaur fossil found by teen boy on display in Maastricht
https://nltimes.nl/2[...]
2015-04-22
[156]
논문
Osteology and taxonomy of Mosasaurus conodon Cope 1881 from the Late Cretaceous of North America
[157]
논문
A mosasaur from the Maastrichtian Fox Hills Formation of the northern Western Interior Seaway of the United States and the synonymy of ''Mosasaurus maximus'' with ''Mosasaurus hoffmanni'' (Reptilia: Mosasauridae)
[158]
논문
Rediagnosis and redescription of ''Mosasaurus hoffmannii'' (Squamata: Mosasauridae) and an assessment of species assigned to the genus ''Mosasaurus''
[159]
논문
Redescription of ''Prognathodon lutugini'' (Squamata, Mosasauridae)
https://pdfs.semanti[...]
[160]
서적
The geology and fossils of the Tertiary and Cretaceous formations of Sussex
https://www.biodiver[...]
Longman, Brown, Green and Longmans
1850
[161]
서적
A monograph on the fossil Reptilia of the Cretaceous formations. Part I
https://www.biodiver[...]
The Palaeontographical Society
1851
[162]
논문
Reassessment of Turonian mosasaur material from the 'Middle Chalk' (England, U.K.), and the status of Mosasaurus gracilis Owen, 1849.
[163]
웹사이트
Bulletins de l'Académie royale des sciences, des lettres et des beaux-arts de Belgique
https://books.google[...]
2019-10-21
[164]
서적
Cretaceous
[165]
논문
Recent mosasaur discoveries from New Jersey and Delaware, USA: stratigraphy, taphonomy and implications for mosasaur extinction
[166]
논문
Paleoecology of the Delaware Valley region, Part II: Cretaceous to Quaternary
https://docs.google.[...]
[167]
논문
Glauconite Composition and Morphology, Shocked Quartz, and the Origin of the Cretaceous(?) Main Fossiliferous Layer (MFL) in Southern New Jersey, U.S.A.
[168]
서적
The Princeton Field Guide to Mesozoic Sea Reptiles
https://www.geokniga[...]
Princeton University Press
[169]
논문
Lepidosaurian diversity in the Mesozoic–Palaeogene: the potential roles of sampling biases and environmental drivers
https://royalsociety[...]
[170]
논문
Convergent Evolution in Aquatic Tetrapods: Insights from an Exceptional Fossil Mosasaur
[171]
논문
A challenge to categories: "What, if anything, is a mosasaur?"
[172]
논문
Landlubbers to leviathans: evolution of swimming in mosasaurine mosasaurs
[173]
논문
Occurrence of Mosasaurus hoffmannii Mantell, 1829 (Squamata, Mosasauridae) in the Maastrichtian Phosphates of Morocco
https://www.research[...]
[174]
논문
Rediagnosis and redescription of ''Mosasaurus hoffmannii'' (Squamata: Mosasauridae) and an assessment of species assigned to the genus ''Mosasaurus''
https://www.research[...]
[175]
논문
Biostratigraphy of the Mosasauridae (Reptilia) from the Cretaceous of Antarctica
[176]
논문
Maastrichtian marine reptiles of the Mediterranean Tethys: a palaeobiogeographical approach
[177]
논문
Paleobiogeography of the Cretaceous Western Interior Seaway of North America: the vertebrate evidence
[178]
논문
A cool temperate climate on the Antarctic Peninsula through the latest Cretaceous to early Paleogene
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com