구인류와 현생인류의 혼혈
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
구인류와 현생인류의 혼혈은 네안데르탈인, 데니소바인, 그리고 아직 알려지지 않은 고대 인류와의 유전적 혼합을 포함한다. 네안데르탈인과의 혼혈은 유라시아인에게서 1~4%의 유전적 기여를 보이며, 동아시아인에게서 더 높은 비율로 나타난다. 데니소바인과의 혼혈은 멜라네시아인에게서 약 4~6%의 DNA를 기여하며, 동남아시아와 오스트레일리아 원주민에게서도 나타난다. 최근 연구에서는 아프리카에서도 알려지지 않은 고인류와의 혼혈 흔적이 발견되었으며, 유라시아의 네안데르탈인과 데니소바인 계통에서도 고대 인류와의 유전자 혼합이 확인되었다.
더 읽어볼만한 페이지
- 고인류유전사 - 현생 인류의 아프리카 기원
현생 인류의 아프리카 기원설은 호모 사피엔스가 약 30만 년에서 20만 년 전 아프리카에서 기원하여 전 세계로 이주했다는 가설로, 유전자 분석, 화석 증거, 고고학적 발견 등으로 뒷받침된다. - 중석기 시대 - 잔석기
잔석기는 작고 날카로운 석기로, 후기 구석기 시대에서 신석기 시대로의 전환기에 사용되었으며, 창, 작살, 화살 등에 사용되어 사냥 효율을 높이는 데 기여했다. - 중석기 시대 - 고대 북유라시아인
- 잡종 포유류 - 늑대개
늑대개는 늑대와 개의 교배로 태어난 동물로, 늑대 혈통 비율에 따라 분류되며, 일부는 특정 품종으로 인정받기도 하지만 여러 국가에서 법적으로 규제된다. - 잡종 포유류 - 홀핀
홀핀은 큰돌고래와 거짓 범고래 사이에서 태어난 잡종으로, 두 종의 특징을 가지며, 지능적이고 사교적인 행동을 보이는 중간 크기의 짙은 회색 톤의 몸 색깔을 가진 동물이다.
2. 네안데르탈인과의 혼혈
네안데르탈인 유해가 19세기에 발견된 이래 현생인류와의 혼혈 가능성이 제기되었다. 토머스 헉슬리는 유럽인에게서 네안데르탈인의 흔적이 나타난다고 주장했지만, 네안데르탈인의 특징을 원시적인 것으로 간주했다.[58] 20세기 초 한스 페데르 스틴스비는 현생인류가 혼합된 기원을 가지고 있다고 주장하며, 덴마크인에게서 네안데르탈인의 특징이 나타난다고 보았다.[61] 1962년 칼턴 스티븐스 쿤은 두개골 데이터와 물질 문화를 근거로 혼혈 가능성을 제시했다.[62]
2000년대 초 대다수 학자들은 아프리카 기원설을 지지하여 현생인류가 아프리카에서 기원하여 네안데르탈인을 대체했다고 보았다.[63][64] 그러나 에릭 트링커스는 포르투갈 Lagar Velho의 어린이 유골[66][67][68]과 루마니아 Peștera Muierii 유골[69] 등을 근거로 혼혈 가설을 계속 주장했다.
2. 1. 유전학적 증거

2010년, 빈디야 동굴 네안데르탈인 3명의 게놈 시퀀싱 결과, 네안데르탈인 게놈 초안이 발표되었다. 이 연구에 따르면 네안데르탈인은 사하라 이남 아프리카인(예: 요루바족, 산족)보다 유라시아인(예: 프랑스인, 한족, 파푸아뉴기니인)과 더 많은 대립유전자를 공유하는 것으로 나타났다.[49] 이는 네안데르탈인으로부터 현생 인류로의 유전자 흐름이 아프리카 밖으로 이주한 후에 발생했음을 시사한다.[49] 유라시아인 게놈의 1~4%가 네안데르탈인에게서 유래했다고 추정되었다.[49] 이후 다른 연구에서는 비아프리카인에게서 네안데르탈인 조상 비율을 1~6%[9], 1.5~2.1%[10], 3.4~7.3%[52], 1.8~2.6%[11] 등으로 추정했다.
이후 연구에 따르면 아프리카인(특히 1000 지놈 프로젝트 아프리카 인구)에게도 네안데르탈인 혼혈이 존재하며, 이는 게놈의 약 0.3%에 해당한다.[12][13] 이는 주로 조상 유럽인(동아시아인과 유럽인 사이의 분리 이후 분기)과 네안데르탈인 혼혈을 가진 현생 인류의 역이동으로 인해 발생했으며, 약 2만 년 전에 일어난 것으로 추정된다.[12][13] 그러나 일부 과학자들은 아프리카로의 DNA 흐름이 얼마나 광범위했을지에 대해 의문을 제기하며, 네안데르탈인 혼혈 신호가 "매우 약하다"고 언급했다.[14]
동아시아인은 유럽인보다 네안데르탈인 혼혈 비율이 더 높은 것으로 나타났는데,[25][19][20][22][21] 이는 약 20% 더 많은 유전자 유입으로 추정된다.[25][19][21] 이는 유럽인과 동아시아인이 분리된 후 동아시아 조상에게서 추가적인 혼혈이 발생했거나,[6][25][19][20][21] 네안데르탈인 조상이 낮은 개체군에서 유래한 인구에 의해 유럽인에게서 희석되었거나,[6][19][21] 동아시아로 이주하면서 유효 개체군 크기가 작아짐에 따라 동아시아 조상에서 정화 선택의 효율이 감소했기 때문일 수 있다.[6][22][21] 혼혈 모델을 시뮬레이션한 연구에서는 더 복잡한 모델이 선호되는데,[23][24] 조상 유라시아인에게 펄스, 분리, 그리고 조상 동아시아인에게 추가 펄스를 보여준다.[6] 유럽 인구 내에서는 네안데르탈인 혼혈 비율에 작지만 유의미한 변동이 관찰되지만, 동아시아 인구 내에서는 유의미한 변동이 없다.[25] 동아시아인이 서유라시아인보다 더 많은 네안데르탈인 DNA를 가지고 있다는 연구 결과도 있다.[11]
최근 연구에서는 동아시아인이 유럽인에 비해 8% 더 많은 네안데르탈인 조상을 가지고 있다고 수정되었는데,[12] 이는 아프리카인을 참조 표본으로 사용했기 때문에 아프리카인과 공유된 네안데르탈인 조상이 가려져 비아프리카인, 특히 유럽인에서 네안데르탈인 혼혈을 과소평가했기 때문이다.[12]
유전체 분석에 따르면 사하라 이남 아프리카 인구와 다른 현대 인류 집단(북아프리카인 포함) 간에는 네안데르탈인 유전자 유입에 대한 전반적인 구분이 존재한다.[26] 북아프리카 집단은 비아프리카 인구와 마찬가지로 네안데르탈인과 유사한 초과 유래 대립 유전자를 공유한다.[27] 북아프리카 인구 내 네안데르탈인 유전 신호는 북아프리카, 유럽, 근동 및 사하라 이남 조상의 상대적 양에 따라 다른 것으로 나타났다.[28] 마사이족에게서도 낮지만 유의미한 수준의 네안데르탈인 혼혈이 관찰되었는데, 이는 최근의 비아프리카 현대 인류(네안데르탈인 이후) 유전자 흐름이 기여한 것으로 추정된다.[20]

여성 알타이 네안데르탈인의 게놈 서열 분석 결과, 아프리카 외 현대 인류의 네안데르탈인 구성 요소는 메즈마이스카야 동굴(Mezmaiskaya cave) 네안데르탈인(북캅카스)과 데니소바 동굴(Denisova Cave) 네안데르탈인(시베리아) 또는 빈디야 동굴(Vindija Cave) 네안데르탈인(크로아티아)보다 더 관련이 있는 것으로 밝혀졌다.[10] 이후 빈디야 네안데르탈인 조각의 게놈 분석 결과, 빈디야 및 메즈마이스카야 네안데르탈인은 현대 인류와의 대립 유전자 공유 정도에서 차이가 없었다.[11]
염색체 21의 알타이, 엘 시드론 동굴(Sidrón Cave)(스페인), 빈디야 네안데르탈인 분석 결과, 엘 시드론 및 빈디야 네안데르탈인만이 현대 인류로의 유전자 흐름(0.3–2.6%)이 유의미하게 나타났다.[32] 반대로, 현대 인류의 유의미한 유전자 흐름은 알타이 네안데르탈인에서만 발생했으며(0.1–2.1%), 이는 현대 인류의 네안데르탈인으로의 유전자 흐름이 약 100,000년 전의 초기 현생 인류 집단에서 기원했음을 시사한다.[32]
현생 인류에게서 네안데르탈인 미토콘드리아 DNA의 증거는 발견되지 않았다.[33][34][35] 이는 네안데르탈인 남성과 현생 인류 여성의 짝짓기에서 성공적인 네안데르탈인 혼혈이 일어났음을 시사한다.[36][37] 네안데르탈인의 Y 염색체가 10만 년에서 37만 년 전에 현생 인류의 멸종된 Y 염색체 계통으로 대체되었다는 연구 결과도 있다.[38]
현생 인류와 네안데르탈인 사이의 매우 낮은 혼혈 비율(약 77세대마다 두 집단 사이에서 한 쌍의 개체가 교환)에 의해 혼혈이 발생했을 수 있다는 모델도 제시되었다.[39]
2. 2. 유입된 유전체의 영향
네안데르탈인으로부터 유입된 유전자는 현생인류의 다양한 형질에 영향을 미치는 것으로 나타났다. 케라틴 필라멘트, 당 대사, 근육 수축, 체지방 분포, 에나멜 두께, 난모세포 감수 분열, 뇌 크기 및 기능과 관련된 유전자가 대표적이다.[41] 특히 피부 색소 침착, 모발 형태, 면역 체계 관련 유전자에서 양성 선택의 증거가 발견되었다.[41]케라틴에 영향을 미치는 유전자는 네안데르탈인으로부터 현생인류, 특히 동아시아인과 유럽인에게 유입되어 피부와 모발 형태의 적응에 기여했을 것으로 추정된다.[25][22] 또한 전신성 홍반성 루푸스, 원발성 담즙성 간경변증, 크론병, 시신경 유두 크기, 흡연 행동, 인터루킨 18 수치, 제2형 당뇨병 등 다양한 질병과 관련된 유전자에서도 네안데르탈인 유래 변이가 발견되었다.[22]
동아시아인의 염색체 3p21.31 영역(HYAL 영역)에서는 UV 광선 적응과 관련된 18개의 네안데르탈인 유입 유전자가 발견되었다.[42] 이 유전자들은 동아시아인에게서만 높은 빈도로 나타나며, 약 45,000년 전부터 급격히 증가하여 5,000~3,500년 전 사이에 성장률이 정점에 달했다.[42] 이는 동아시아인과 아메리카 원주민의 공통 조상에서 발생한 것으로 추정된다.[42]
면역 체계에서는 HLA-A*02, A*26/*66, B*07, B*51, C*07:02, C*16:02 등 네안데르탈인 유래 유전자가 현생인류의 면역 다양성 회복과 지역 병원체 적응에 기여했다.[79]
유입된 네안데르탈인 유전자는 시스 조절 효과를 통해 현생인류의 유전체 복잡성과 표현형 변이에 기여한다.[45] 특히 뇌와 고환에서 네안데르탈인 대립 유전자의 발현이 낮게 나타났는데, 이는 소뇌와 기저핵에서 두드러졌다.[6][45]
고해상도 암컷 빈디야 네안데르탈인 유전체 연구에서는 LDL 콜레스테롤 및 비타민 D 수치, 식이 장애, 내장 지방 축적, 류마티스 관절염, 조현병, 항정신병 약물 반응 등에 영향을 미치는 네안데르탈인 유래 유전자 변이가 확인되었다.[11]
유럽 현생인류 대상 연구에서는 네안데르탈인 혼혈이 두개골 및 기저 뇌 형태 변화와 관련이 있으며, 신경학적 기능 변화를 시사하는 유전자 변이가 발견되었다.[46]
파푸아인의 경우, 네안데르탈인 유전자 변이는 뇌 발현 유전자에서, 데니소바인 DNA는 뼈 및 기타 조직 발현 유전자에서 높은 빈도로 나타났다.[47]
현생 인류가 상속받은 네안데르탈인 대립 유전자 SNP rs3917862는 과응고성과 관련이 있지만, 이 대립 유전자가 없는 여성은 출산 시 사망 확률이 더 높다는 연구 결과도 있다.[48]
2023년 12월, 과학자들은 네안데르탈인과 데니소바인으로부터 현생 인류에게 상속된 유전자가 현생 인류의 일상에 생물학적으로 영향을 미칠 수 있다고 보고했다.[8]
2. 3. 형태학적 증거
오리냐크 상부 구석기 시대의 아브리고 도 라가르 벨류 (포르투갈)에서 발견된 현대 인류 아이의 매장 유해는 이베리아로 확산된 현대 인류와 네안데르탈인의 이종 교배를 나타내는 특징을 보여준다.[53] 매장 유해는 24,500년 전으로 연대 측정되었으며, 이는 이베리아에서 네안데르탈인에서 현대 인류로의 전환기(28,000–30,000년 전) 이후 오랫동안 네안데르탈인의 특징이 지속되었음을 보여준다. 따라서 이 아이는 이미 혼혈이 심한 인구의 후손이었을 수 있다.[53]페스테라 무이에릴로 (루마니아)에서 발견된 35,000년 전의 초기 상부 구석기 시대 현대 인류 유해는 유럽 초기 현대 인류의 형태학적 패턴을 보여주지만, 아치형 또는 네안데르탈인의 특징도 가지고 있다. 이는 유럽 초기 현대 인류가 네안데르탈인과 이종 교배했음을 시사한다.[69] 이러한 특징으로는 넓은 안와 간격, 비교적 평평한 눈썹 활, 두드러진 후두 융기, 비대칭적이고 얕은 하악 절흔 모양, 높은 하악 관상 돌기, 절흔 능선 위치에 상대적으로 수직적인 하악 과두 돌기, 좁은 어깨뼈 관절 와 등이 있다.[69]
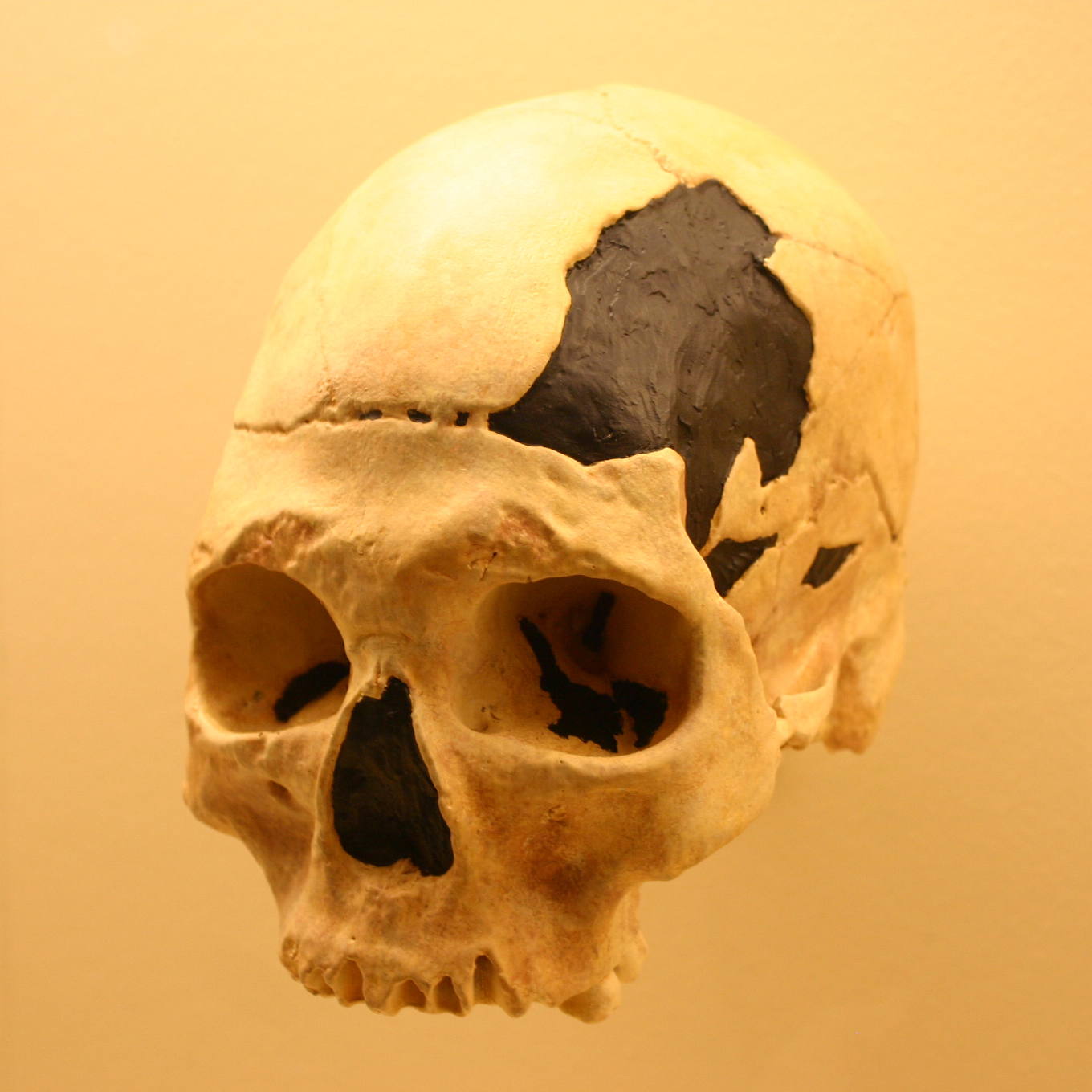
페스테라 쿠 오아세 (루마니아)에서 발견된 34,000–36,000년 전의 초기 현대 인류 오아세 1 하악골은 현대, 아치형, 그리고 네안데르탈인의 특징이 혼합된 모자이크 형태를 보인다.[55] 이 유해는 후기 중기 및 후기 플라이스토세의 네안데르탈인을 제외하고 이전 인류에게는 존재하지 않았던 하악공의 설측 가교 현상을 보여주는데, 이는 네안데르탈인과의 친화성을 시사한다.[55] 결론적으로, 오아세 1 하악골은 적어도 유럽의 초기 현대 인류가 네안데르탈인과의 어느 정도의 혼혈로 인해 상당한 두개안면 변화를 겪었을 가능성을 보여준다.[55]
가장 초기의 (약 33,000년 전) 유럽 현대 인류와 그 이후의 (중기 상부 구석기 시대) 그라베트 문화인들은 해부학적으로 초기의 (중기 구석기 시대) 아프리카 현대 인류와 거의 일치하지만, 네안데르탈인의 특징도 보여준다. 이는 유럽 초기 현대 인류를 중기 구석기 시대 현대 인류 조상만으로는 설명하기 어렵다는 것을 시사한다.[56]
최근 마노트 동굴 (이스라엘 서부 갈릴리)에서 발견되어 54.7±5.5 kyr BP로 연대 측정된 현대 인류의 부분적인 두개골인 마노트 1은 현대 인류가 아프리카에서 성공적으로 탈출하여 유라시아를 식민화한 시기의 최초의 화석 증거이다.[57] 이 화석은 또한 현대 인류가 중기-상부 구석기 시대의 경계 동안 네안데르탈인과 동시대에 남부 레반트에 거주했음을 보여주는 최초의 증거이며, 이는 이종 교배가 일어났을 가능성이 있는 시기와 가깝다.[57] 형태학적 특징은 마노트 개체군이 이후 유럽을 성공적으로 식민화하여 초기 상부 구석기 시대 개체군을 형성한 최초의 현대 인류와 밀접하게 관련되어 있거나 그들에게서 기원했을 수 있음을 시사한다.[57]
3. 데니소바인과의 혼혈
멜라네시아인, 오스트레일리아 원주민, 그리고 동남아시아 일부 섬 지역 인구 집단에서 높은 비율의 데니소바인 유래 DNA가 발견된다. 이는 데니소바 동굴에서 발견된 고인류인 데니소바인이 현생인류와 혼혈되었음을 보여주는 증거이다.
Reich et al. (2011)의 연구와 Skoglund와 Jakobsson (2011)의 연구는 공통적으로 오세아니아 집단에서 데니소바인 혼혈이 가장 높고, 많은 동남아시아 집단이 그 다음이며, 동아시아 집단에는 데니소바인 혼혈이 거의 없다는 결과를 제시했다.[71][73]
여러 연구에 따르면 데니소바인 유전자 유입은 필리핀 원주민, 호주 원주민, 뉴기니인의 공통 조상에게 일어났다.[71][74] 뉴기니인과 호주인은 유사한 데니소바인 혼혈 비율을 보이는데, 이는 적어도 44,000년 전 사훌 대륙붕 진입 이전에 혼혈이 발생했음을 시사한다.[71]
Browning et al. (2018)은 최소 두 번의 데니소바인 혼혈 사건을 제안했다. 동아시아인은 두 개의 별개 데니소바인 집단, 남아시아인과 오세아니아인은 하나의 데니소바인 집단에서 유입을 보인다.[75] Sankararaman et al. (2016)은 데니소바인 혼혈 시기를 44,000–54,000년 전으로 추정했다.[4]
2021년 연구에서는 필리핀 네그리토족, 특히 아이타 마그부콘에게서 세계 최고 수준의 데니소바인 조상이 발견되었다.[76] 이는 호주인과 파푸아인보다 ~30%–40% 더 높으며, 필리핀에 도착한 현대 인류와 혼혈된 별개 섬 데니소바인 집단이 존재했음을 시사한다.[76]
4만년 전 톈위안인 유해는 네안데르탈인 기여는 보였지만, 데니소바인 기여는 없었다.[78] 이는 데니소바인 유전적 기여가 대륙에서 희소했음을 시사한다.[10]
3. 1. 유전학적 증거
멜라네시아인의 유전체에는 약 4~6%의 데니소바인 DNA가 포함되어 있다.[77] 오스트레일리아 원주민에게서도 멜라네시아인과 유사한 수준의 데니소바인 혼혈이 나타난다.[70] 동아시아인과 아메리카 원주민에게서는 낮은 수준의 데니소바인 혼혈 흔적이 발견될 수 있다.[73] Prüfer et al. (2013)의 연구에 따르면, 대륙 아시아와 아메리카 원주민 집단은 0.2%의 데니소바인 유전자 기여를 보일 수 있으며, 이는 오세아니아 집단보다 약 25배 낮은 수치이다.[10] 그러나 Wall et al. (2013)은 동아시아인에게서 데니소바인 혼혈에 대한 증거를 찾지 못했다고 언급했다.[20]
티베트인은 데니소바인으로부터 고지대 생활에 적합한 EGLN1 및 EPAS1 유전자 변이를 획득했으며, 이는 헤모글로빈 농도 및 저산소증 반응과 관련이 있다.[41]
3. 2. 유입된 유전체의 영향
현생 인류의 변화에 대한 탐구에서 면역 체계의 HLA 대립 유전자를 탐구한 결과, HLA-B*73이 서아시아의 현생 인류로 데니소바인으로부터 유입되었다는 주장이 제기되었으며, 이는 HLA-B*73의 분포 패턴과 다른 HLA 대립 유전자와의 분기에서 비롯되었다.[79] HLA-B*73이 시퀀싱된 데니소바인 게놈에는 존재하지 않지만, HLA-B*73은 연관 불평형에서 데니소바인 유래의 HLA-C*15:05와 밀접하게 관련되어 있는 것으로 나타났다.[79] 그러나 계통 발생 분석 결과 HLA-B*73이 조상 유전자일 가능성이 매우 높다는 결론이 내려졌다.[41]데니소바인의 두 가지 HLA-A (A*02 및 A*11)와 두 가지 HLA-C (C*15 및 C*12:02) 동종 유전자는 현생 인류의 일반적인 대립 유전자와 일치하는 반면, 데니소바인의 HLA-B 동종 유전자 중 하나는 희귀한 재조합 대립 유전자와 일치하며 다른 하나는 현생 인류에게서 발견되지 않는다.[79] HLA 대립 유전자의 높은 돌연변이율로 인해 오랫동안 두 종에서 독립적으로 보존되었을 가능성이 낮기 때문에, 이러한 유전자들은 데니소바인으로부터 현생 인류에게 전달되었을 것으로 생각된다.[79]
티베트인은 데니소바인으로부터 고지대 생활에 적합한 EGLN1 및 EPAS1 유전자 변이를 획득했으며, 이는 헤모글로빈 농도 및 저산소증 반응과 관련이 있다.[41] EPAS1의 조상 유전자 변이는 고지대와 같이 산소 수치가 낮은 환경에서 헤모글로빈 수치를 증가시켜 이를 보상하지만, 이는 혈액 점도를 증가시키는 부적응을 초래한다.[80] 반면에 데니소바인 유래 변이는 이러한 헤모글로빈 수치의 증가를 제한하여 더 나은 고도 적응을 가능하게 한다.[80] 데니소바인 유래 EPAS1 유전자 변이는 티베트인에게서 흔하게 나타나며, 티베트 고원을 식민지화한 후 조상에게서 긍정적으로 선택되었다.[80]
4. 아프리카 고인류와의 혼혈
아프리카 내에서도 현생 인류와 멸종된 고인류 간의 혼혈 가능성이 제기되고 있다. 특히, 최근 연구들은 아프리카인들에게서도 네안데르탈인과의 혼혈 흔적이 발견된다고 보고 있다. 이는 현생 인류와 네안데르탈인의 혼혈이 아프리카 밖에서만 일어났다는 기존의 통념과는 다른 결과이다.
하지만, 사하라 사막 이남 아프리카 지역은 화석 보존이 어려워 고인류의 DNA를 확보하기 어렵기 때문에, 직접적인 비교 연구에는 한계가 있다. 그럼에도 불구하고, 일부 서아프리카 인구(예: 요루바족, 멘데족)에게서 고대 DNA의 흔적이 발견되었으며, 이는 이들이 농경 이전의 수렵 채집인이나 다른 고대 아프리카 인류와 유전적으로 연관되어 있을 가능성을 시사한다.
4. 1. 유전학적 증거
2010년 5월 7일, 빈디야 동굴 네안데르탈인 3명의 게놈 시퀀싱 결과, 네안데르탈인 게놈의 초안 염기서열이 발표되었다. 연구 결과에 따르면 네안데르탈인은 사하라 이남 아프리카인(예: 요루바족 및 산족)보다 유라시아 인구(예: 프랑스인, 한족, 파푸아뉴기니인)와 더 많은 대립유전자를 공유하는 것으로 나타났다.[49] 이는 아프리카 밖으로 이주한 후 네안데르탈인에서 현생 인류로 유전자 흐름이 발생했음을 보여준다.[49] 유라시아인 게놈에서 네안데르탈인 유래 조상의 비율은 1~4%로 추정되었다.[49] 이후 연구들에 따르면 비아프리카인의 네안데르탈인 조상 비율은 1.5~2.1%[10], 3.4~7.3%[52], 1.8~2.6%[11] 등으로 다양하게 추정되었다.2020년 연구에서는 아프리카인(특히, 1000 지놈 프로젝트 아프리카 인구)도 네안데르탈인 혼혈을 가지고 있다는 사실이 밝혀졌다.[12] 아프리카인 개인의 네안데르탈인 혼혈은 17 메가베이스(그들 게놈의 0.3%)를 차지한다.[12][13] 이는 주로 조상 유럽인(동아시아인과 유럽인 사이의 분리 이후)과 분기된 사람들(네안데르탈인 혼혈을 가진 현생 인류)의 역이동으로부터 얻어진 것으로, 약 2만 년 전에 일어났을 것으로 추정된다.[12][13] 그러나 데이비드 라이히와 같은 일부 과학자들은 아프리카로의 DNA 흐름이 얼마나 광범위했을지에 대해 의문을 제기하며, 네안데르탈인 혼혈 신호가 매우 약하다는 것을 발견했다.[14]
아프리카 사하라 사막 이남 환경에서는 화석의 빠른 부패로 인해, 현생 인류의 혼혈을 고대 사하라 사막 이남 아프리카 호미닌의 기준 표본과 비교하는 것은 현재 불가능하다.[6][84]
하지만, 약 4,500년 전 에티오피아 고지대 개인과 남부(약 2,300~1,300년 전), 동부 및 중남부 아프리카(약 8,100~400년 전)의 고대 DNA 데이터 분석 결과, 일부 서아프리카 인구는 농경 이전의 동아프리카 수렵 채집인, 남아프리카 수렵 채집 인구 또는 그들 사이의 유전적 점진성을 포함하지 않는 서아프리카인에서 고대 기원으로 가장 잘 설명되는 소량의 과도한 대립 유전자를 가지고 있음이 밝혀졌다. 이러한 고대 DNA를 보유한 서아프리카 그룹에는 나이지리아 해안의 요루바족과 시에라리온의 멘데족이 포함되며, 이는 고대 DNA가 농업 확산 훨씬 전, 홀로세(11,600년 전 시작) 훨씬 전에 획득되었음을 나타낸다. 이 고대 계통은 약 20만~30만 년 전에 시작된 것으로 추정되는 산족 조상의 분기 전에 분리되었어야 한다.[82][83]
이러한 가설은 Lachance et al.(2012),[84] Hammer et al., 2011,[85] 및 Plagnol and Wall(2006)을 포함한 다른 인간 해플로타입과 깊은 분기를 가진 긴 해플로타입을 기반으로 한 연구 결과와도 일치한다.[86]
Hammer 등의 연구에서 중앙 아프리카의 피그미족은 서아프리카의 요루바족과 대조적으로 남아프리카의 산족과 함께 그룹화된다. 카메룬의 슘 라카에서 발견된 4개의 화석(8,000~3,000년 전) DNA 분석 결과, 이들은 대부분의 DNA를 중앙 아프리카 수렵 채집인(피그미 조상)에게서 얻었으며 요루바족과 만데족에서 발견된 고대 DNA를 공유하지 않았다.[87] Lipson et al.(2020)의 연구에서도 동부, 중앙 및 남부 수렵 채집인과 서아프리카 그룹 간의 차이 패턴이 확인되었다.[88]
2020년 연구에서는 4개의 서아프리카 인구 DNA의 2%~19%가 36만 년에서 102만 년 전에 인간과 네안데르탈인의 조상으로부터 분리된 알려지지 않은 고대 호미닌에서 나왔을 수 있다는 징후가 발견되었다. 그러나 이 연구는 제안된 고대 혼혈의 적어도 일부가 유라시아인/비아프리카인에서도 존재하며, 혼혈 사건이 0~12만 4천 년 전(아프리카 외부 이주 이전 및 아프리카/유라시아 분리 이전)에 발생했을 가능성도 제시한다.[89][90][91] 최근 연구에서는 현대 인류보다 앞선 아프리카인에서 조상 유전적 변이가 발견되었으며, 이는 대부분의 비아프리카인에서 손실되었다.[92]
5. 유라시아 고인류와의 혼혈
유라시아 대륙에서 고인류와 현생인류 간의 혼혈은 여러 유전학적 증거를 통해 밝혀졌다. 2010년 빈디야 동굴 네안데르탈인의 게놈 염기서열 초안이 발표되면서, 네안데르탈인이 사하라 이남 아프리카인보다 유라시아인과 더 많은 대립유전자를 공유한다는 사실이 알려졌다.[49] 이는 아프리카에서 이주한 현생인류가 네안데르탈인과 유전자를 교환했음을 시사한다.[49]
이후 연구들을 통해 비아프리카인에게서 발견되는 네안데르탈인 유래 유전자의 비율은 다양하게 추정되었는데, 초기에는 1~4%[49], 이후 1.5~2.1%[10], 3.4~7.3%[52] 등으로 추정되었으며, 2017년에는 오세아니아를 제외한 비아프리카인에서 1.8~2.6%로 수정되었다.[11]
최근 연구에서는 아프리카인에게도 네안데르탈인 혼혈의 흔적이 발견되었다.[12] 1000 지놈 프로젝트의 아프리카인 유전체 분석 결과, 이들의 게놈 중 약 0.3%가 네안데르탈인으로부터 유래한 것으로 나타났다.[12][13] 이는 주로 유럽 조상으로부터의 역이동을 통해 약 2만 년 전에 발생한 것으로 추정된다.[12][13] 다만, 일부 과학자들은 이러한 DNA 흐름의 규모에 대해 의문을 제기하기도 한다.[14]
유라시아에는 적어도 200만 년 전부터 인류가 출현했으며, 이후 네안데르탈인과 데니소바인 계통이 유라시아로 확장되면서 이 지역에 살고 있던 고대 인류의 후손들과 유전적 혼합이 발생했다. 이러한 혼합은 네안데르탈인과 데니소바인, 그리고 현생인류의 게놈에 영향을 미쳤다.[93][94]
유전학 연구에 따르면, 초대 고대 인류로부터의 유전자 혼합은 크게 두 가지 사건으로 나타난다.[93] 첫 번째는 네안데르소반(네안데르탈인과 데니소바인의 공통 조상)이 유라시아로 확장된 직후, 200만 년 전 아프리카 호모 계통에서 분리된 초대 고대 인류 계통과의 혼혈 사건이다.[93] 두 번째는 약 35만 년 전, "에렉투스형" 생물의 유전자가 데니소바인 계통으로 유입된 사건이다.[93] 이 두 집단은 이전에 상호 교배하는 것으로 알려진 어떤 인간 집단보다 더 멀리 떨어져 있었다.[93][94]
5. 1. 유전학적 증거
2010년 5월 7일, 빈디야 동굴 네안데르탈인 3명의 게놈 시퀀싱을 통해 네안데르탈인 게놈 초안이 발표되었다. 연구 결과, 네안데르탈인은 사하라 이남 아프리카인보다 유라시아인(프랑스인, 한족, 파푸아뉴기니인 등)과 더 많은 대립유전자를 공유하는 것으로 나타났다.[49] 이는 네안데르탈인으로부터 현생 인류로 유전자 흐름이 있었음을 보여준다.[49] 유라시아인 게놈에서 네안데르탈인 유래 조상 비율은 1~4%로 추정되었다.[49] 이후 연구들에 따라 비아프리카인의 네안데르탈인 조상 비율 추정치는 1.5~2.1%[10], 3.4~7.3%[52] 등으로 다양하며, 2017년에는 오세아니아를 제외한 비아프리카인에서 1.8~2.6%로 수정되었다.[11]Chen 등(2020)의 연구에 따르면, 아프리카인(특히 1000 지놈 프로젝트 아프리카 인구)도 네안데르탈인 혼혈을 가지고 있으며, 이는 게놈의 0.3%에 해당한다.[12][13] 이 혼혈은 주로 조상 유럽인과의 역이동으로 인해 발생했으며, 약 2만 년 전에 일어난 것으로 추정된다.[12][13] 그러나 일부 과학자들은 아프리카로의 DNA 흐름이 얼마나 광범위했는지에 대해 의문을 제기한다.[14]
동아시아인은 유럽인보다 네안데르탈인 혼혈 비율이 더 높은 것으로 나타났다.[25][19][20][22][21] 이는 동아시아인에게 약 20% 더 많은 유전자 유입이 있었음을 의미한다.[25][19][21] 그 원인에 대해서는 여러 가설이 제시되었다.
- 유럽인과 동아시아인이 분리된 후 동아시아 조상에게서 추가적인 혼혈 발생[6][25][19][20][21]
- 네안데르탈인 조상이 낮은 개체군에서 유래한 인구에 의해 유럽인에게서 희석[6][19][21]
- 동아시아로 이주하면서 유효 개체군 크기가 작아짐에 따라 정화 선택의 효율 감소[6][22][21]
유전체 분석 결과, 사하라 이남 아프리카 인구와 다른 현대 인류 집단(북아프리카인 포함) 간에는 네안데르탈인 유전자 유입에 대한 뚜렷한 구분이 존재한다.[26] 북아프리카 집단은 비아프리카 인구와 유사하게 네안데르탈인과 초과 유래 대립 유전자를 공유한다.[27] 튀니지 베르베르족과 같이 북아프리카 조상이 많은 인구 집단에서 네안데르탈인 혼혈 추정치가 가장 높게 나타났다.[28]
르무스티에 네안데르탈인 두개골 재구성을 통해 알타이, 엘 시드론 동굴, 빈디야 네안데르탈인을 분석한 결과, 엘 시드론 및 빈디야 네안데르탈인만이 현대 인류로의 유전자 흐름(0.3–2.6%)이 유의미하게 나타났다.[32] 이는 엘 시드론 및 빈디야 네안데르탈인이 알타이 네안데르탈인보다 현대 인류와 더 밀접하게 관련되어 있음을 시사한다.[32]
유라시아에 인류가 출현한 것은 적어도 200만 년 전부터이다. 유전자 증거에 따르면, 네안데르탈인과 데니소바인의 계통이 유라시아로 확장되기 시작했을 때, 이 대륙에는 고대 인류의 후손들이 살고 있었고, 이들과의 유전적 혼합이 네안데르탈인과 데니소바인, 그리고 현생 인류의 게놈에 영향을 미쳤다.[93][94]
유전학 연구는 초대 고대 인류로부터의 유전자 혼합의 두 가지 주요 사건을 보여준다.[93]
6. 관련 연구
2019년, 과학자들은 인공지능을 활용한 유전학 연구를 통해, 현생 인류의 게놈에 네안데르탈인이나 데니소바인이 아닌, 알려지지 않은 인류 조상 종의 존재를 시사하는 증거를 발견했다.[95][96]
참조
[1]
논문
Midfacial Morphology and Neandertal–Modern Human Interbreeding
2022-08
[2]
뉴스
A handful of recent discoveries have shattered anthropologists' picture of where humans came from, and when
https://www.business[...]
2020-01-06
[3]
논문
Chad Genetic Diversity Reveals an African History Marked by Multiple Holocene Eurasian Migrations
2016
[4]
논문
The Combined Landscape of Denisovan and Neanderthal Ancestry in Present-Day Humans
2016
[5]
웹사이트
Ancient human relative interbred with ancestors of modern humans as recently as 50,000 years ago
https://newsroom.ucl[...]
2022-11-21
[6]
논문
Outstanding questions in the study of archaic hominin admixture
[7]
논문
The Hybrid Origin of "Modern" Humans
https://www.research[...]
[8]
뉴스
Morning Person? You Might Have Neanderthal Genes to Thank. - Hundreds of genetic variants carried by Neanderthals and Denisovans are shared by people who like to get up early.
https://www.nytimes.[...]
2023-12-14
[9]
논문
Diverse African genomes reveal selection on ancient modern human introgressions in Neanderthals
https://www.scienced[...]
2024-05-08
[10]
논문
The complete genome sequence of a Neanderthal from the Altai Mountains
2014
[11]
논문
A high-coverage Neandertal genome from Vindija Cave in Croatia
2017
[12]
논문
Identifying and Interpreting Apparent Neanderthal Ancestry in African Individuals
2020-01
[13]
논문
Africans, too, carry Neanderthal genetic legacy
2020-01-31
[14]
뉴스
Neanderthal Genes Hint at Much Earlier Human Migration From Africa
https://www.nytimes.[...]
2020-01-31
[15]
논문
50,000 years of Evolutionary History of India: Insights from ~2,700 Whole Genome Sequences
https://www.biorxiv.[...]
2024-02-20
[16]
논문
Genomes of modern Indian people include wide range of Neanderthal DNA
https://www.newscien[...]
2024-03-06
[17]
논문
Who are you? How the story of human origins is being rewritten
https://www.newscien[...]
2017-08-25
[18]
논문
Diverse African genomes reveal selection on ancient modern human introgressions in Neanderthals
https://www.scienced[...]
2024-05-08
[19]
논문
A High-Coverage Genome Sequence from an Archaic Denisovan Individual
http://repository.bi[...]
[20]
논문
Higher Levels of Neanderthal Ancestry in East Asians than in Europeans
[21]
논문
Tracing the peopling of the world through genomics
2017
[22]
논문
The genomic landscape of Neanderthal ancestry in present-day humans
2014
[23]
논문
Complex History of Admixture between Modern Humans and Neandertals
2015
[24]
논문
Selection and Reduced Population Size Cannot Explain Higher Amounts of Neandertal Ancestry in East Asian than in European Human Populations
2015
[25]
논문
Resurrecting Surviving Neandertal Lineages from Modern Human Genomes
2014
[26]
논문
North African Populations Carry the Signature of Admixture with Neandertals
[27]
논문
North African Populations Carry the Signature of Admixture with Neandertals
[28]
논문
North African Populations Carry the Signature of Admixture with Neandertals
[29]
논문
North African Populations Carry the Signature of Admixture with Neandertals
[30]
논문
Higher Levels of Neanderthal Ancestry in East Asians than in Europeans
[31]
웹사이트
Neues Museum in Berlin 1175
https://www.flickr.c[...]
2019-05-12
[32]
논문
Ancient gene flow from early modern humans into Eastern Neanderthals
2016
[33]
논문
Neandertal DNA Sequences and the Origin of Modern Humans
[34]
논문
No Evidence of Neandertal mtDNA Contribution to Early Modern Humans
[35]
논문
Archaic admixture in the human genome
[36]
논문
Neanderthal-human Hybrids
http://www.hypothesi[...]
2011
[37]
논문
Neanderthal DNA and modern human origins
[38]
논문
The evolutionary history of Neanderthal and Denisovan Y chromosomes
https://hal.archives[...]
2020-09-25
[39]
논문
Extremely Rare Interbreeding Events Can Explain Neanderthal DNA in Living Humans
2012
[40]
논문
40,000-Year-Old Individual from Asia Provides Insight into Early Population Structure in Eurasia
2017
[41]
논문
Evolutionary and Medical Consequences of Archaic Introgression into Modern Human Genomes
2018-07-18
[42]
논문
Neanderthal Introgression at Chromosome 3p21.31 was Under Positive Natural Selection in East Asians
[43]
논문
Evidence that the adaptive allele of the brain size gene microcephalin introgressed into Homo sapiens from an archaic Homo lineage
[44]
논문
The Microcephalin Ancestral Allele in a Neanderthal Individual
[45]
논문
Impacts of Neanderthal-Introgressed Sequences on the Landscape of Human Gene Expression
2017
[46]
논문
Neanderthal-Derived Genetic Variation Shapes Modern Human Cranium and Brain
2017
[47]
논문
Neanderthal and Denisovan ancestry in Papuans: A functional study
[48]
논문
The relative roles of maternal survival and inter-personal violence as selection pressures on the persistence of Neanderthal hypercoagulability alleles in modern Europeans
[49]
논문
A Draft Sequence of the Neandertal Genome
[50]
논문
Ancient Structure in Africa Unlikely to Explain Neanderthal and Non-African Genetic Similarity
2012
[51]
논문
The Date of Interbreeding between Neandertals and Modern Humans
2012
[52]
논문
Neandertal Admixture in Eurasia Confirmed by Maximum-Likelihood Analysis of Three Genomes
2014
[53]
논문
The early Upper Paleolithic human skeleton from the Abrigo do Lagar Velho (Portugal) and modern-human emergence in Iberia
1999
[54]
웹사이트
Oase 2
http://humanorigins.[...]
Smithsonian National Museum of Natural History
2010-01-23
[55]
논문
An early modern human from the Peştera cu Oase, Romania
2003
[56]
논문
European early modern humans and the fate of the Neandertals
2007
[57]
논문
Levantine cranium from Manot Cave (Israel) foreshadows the first European modern humans
2015-01-28
[58]
서적
Collected Essays: Volume VII, Man's Place in Nature
[59]
논문
L'homme fossile de La Chapelle-aux-Saints
1911–1913
[60]
논문
Neanderthal Man Not Our Ancestor
[61]
웹사이트
Racestudier i Danmark
http://img.kb.dk/tid[...]
Royal Library, Denmark
1907
[62]
논문
The Origin of races
https://archive.org/[...]
Knopf
1962
[63]
논문
A geographically explicit genetic model of worldwide human-settlement history
2006-08
[64]
논문
Human evolution: Out of Ethiopia
2003-06
[65]
뉴스
The Neanderthal within.
New Scientist
2007-03-03
[66]
웹사이트
The Lagar Velho 1 Skeleton
http://www.talkorigi[...]
TalkOrigins Archive
2000-07-31
[67]
웹사이트
Life on the edge: was a Gibraltar cave last outpost of the lost neanderthal?
https://www.theguard[...]
2006-09-13
[68]
웹사이트
Not a lasting last for the Neandertals
http://johnhawks.net[...]
2006-09-13
[69]
논문
Early modern humans from the Pestera Muierii, Baia de Fier, Romania
[70]
논문
An Aboriginal Australian Genome Reveals Separate Human Dispersals into Asia
[71]
논문
Denisova Admixture and the First Modern Human Dispersals into Southeast Asia and Oceania
[72]
논문
Did the Denisovans Cross Wallace's Line?
2013
[73]
논문
Archaic human ancestry in East Asia
2011
[74]
웹사이트
Meet Your Ancient Relatives: The Denisovans
https://www.npr.org/[...]
2012-08-31
[75]
논문
Analysis of Human Sequence Data Reveals Two Pulses of Archaic Denisovan Admixture
2018
[76]
논문
Philippine Ayta possess the highest level of Denisovan ancestry in the world
2021
[77]
논문
Genetic history of an archaic hominin group from Denisova Cave in Siberia
http://repositori.up[...]
2018-07-29
[78]
논문
DNA analysis of an early modern human from Tianyuan Cave, China
2013
[79]
논문
The Shaping of Modern Human Immune Systems by Multiregional Admixture with Archaic Humans
2011
[80]
논문
Altitude adaptation in Tibetans caused by introgression of Denisovan-like DNA
2014
[81]
논문
Ancient Ethiopian genome reveals extensive Eurasian admixture in Eastern Africa
[82]
논문
Reconstructing Prehistoric African Population Structure
2012
[83]
논문
Revising the human mutation rate: implications for understanding human evolution
2012
[84]
논문
Evolutionary History and Adaptation from High-Coverage Whole-Genome Sequences of Diverse African Hunter-Gatherers
2012
[85]
논문
Genetic evidence for archaic admixture in Africa
[86]
논문
Possible ancestral structure in human populations
2006
[87]
논문
Ancient West African foragers in the context of African population history
2020
[88]
논문
Ancient DNA and deep population structure in sub-Saharan African foragers
2022
[89]
논문
Recovering signals of ghost archaic introgression in African populations
[90]
Webarchive
Supplementary Materials for ''Recovering signals of ghost archaic introgression in African populations", section "S8.2" "We simulated data using the same priors in Section S5.2, but computed the spectrum for both YRI [West African Yoruba] and CEU [a population of European origin] . We found that the best fitting parameters were an archaic split time of 27,000 generations ago (95% HPD: 26,000–28,000), admixture fraction of 0.09 (95% HPD: 0.04–0.17), admixture time of 3,000 generations ago (95% HPD: 2,800–3,400), and an effective population size of 19,700 individuals (95% HPD: 19,300–20,200). We find that the lower bound of the admixture time is further back than the simulated split between CEU and YRI (2155 generations ago), providing some evidence in favor of a pre-Out-of-Africa event. This model suggests that many populations outside of Africa should also contain haplotypes from this introgression event, though detection is difficult because many methods use unadmixed outgroups to detect introgressed haplotypes [Browning et al., 2018, Skov et al., 2018, Durvasula and Sankararaman, 2019] (5, 53, 22). It is also possible that some of these haplotypes were lost during the Out-of-Africa bottleneck."
https://www.science.[...]
2020-12-07
[91]
논문
Recovering signals of ghost archaic introgression in African populations
[92]
논문
Insights into human genetic variation and population history from 929 diverse genomes
[93]
논문
Denisovan ancestors interbred with a distantly related hominin
2020
[94]
간행물
Happy New Year Homo erectus? More evidence for interbreeding with archaics predating the modern human/Neanderthal split
2013
[95]
논문
Approximate Bayesian computation with deep learning supports a third archaic introgression in Asia and Oceania
2019-01-16
[96]
뉴스
Artificial Intelligence Has Found an Unknown 'Ghost' Ancestor in The Human Genome
https://www.sciencea[...]
2019-02-11
[97]
저널
The Combined Landscape of Denisovan and Neanderthal Ancestry in Present-Day Humans
2016
[98]
저널
Outstanding questions in the study of archaic hominin admixture
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com