기장아과
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
기장아과는 벼과의 아과로, 설상엽의 가장자리에 털이 나 있으며, 꽃차례는 공통 축을 중심으로 분지된다. 잎은 선형이며, 꽃차례는 원추꽃차례, 총상꽃차례 등이다. 소수는 2개의 작은 꽃을 포함하며, 억새형, 강아지풀형, 수수형의 3가지 유형으로 나뉜다. 남극을 제외한 전 세계에 분포하며, 특히 열대 지방에 많다. PACMAD 분지군에 속하며, 12개의 족으로 나뉜다. 옥수수, 기장, 조, 수수, 사탕수수 등 인류에게 중요한 식량 자원을 포함하며, 억새, 띠 등 문화적으로 중요한 식물도 포함한다.

더 읽어볼만한 페이지
- 1864년 기재된 식물 - 삼채
삼채(*Allium hookeri*)는 남아시아와 동남아시아 등에서 재배되는 식용 식물로, 대한민국에서는 건강식품으로 인기를 얻고 있다. - 1864년 기재된 식물 - 계수나무
계수나무는 동아시아에 분포하는 낙엽성 나무로, 가을에 캐러멜 향이 나는 잎과 건축재로 쓰이는 목재, 그리고 동아시아 문화권에서의 상징적인 의미와 관상수로서의 가치를 지닌다. - 기장아과 - 기네리움
기네리움은 벼과 기장아과에 속하는 식물 속으로, 잎의 구조와 특성을 통해 C3 및 C4 광합성 경로를 모두 사용하는 PACMAD 분지군에 속하며 다양한 종내 분류군을 가진다. - 기장아과 - 제우기테스족
제우기테스족은 벼과에 속하는 족으로 제우기테스속, 조릿대풀속 등 여러 속을 포함하며 PACMAD 분지군에 속한다.
2. 형태적 특징
기장아과에 속하는 식물은 매우 다양하지만, 몇 가지 공통적인 형태적 특징을 공유한다. 대부분 한해살이풀 또는 여러해살이풀의 초본이며,[2] 잎은 보통 선형으로 가늘고 긴 엽신을 가진다. 잎혀(엽설)는 가장자리에 털이 나 있는 경우가 많다.[1]
꽃차례는 원추꽃차례, 총상꽃차례 등 다양하며, 때로는 원추꽃차례의 가지가 짧아져 작은 이삭(소수)들이 빽빽하게 모여 밀집원추꽃차례를 이루기도 한다. 작은 이삭은 양성이거나 단성이며, 단성일 경우 암수딴그루 또는 암수한그루이다.[3] 작은 이삭은 하나씩 달리거나 때로는 쌍으로 달리는데, 쌍으로 달릴 경우 긴 자루를 가진 것과 짧은 자루(또는 자루가 없는)를 가진 것이 짝을 이룬다.[3]
기장아과의 가장 큰 특징은 작은 이삭의 구조에 있다. 겉모습이 매우 다른 억새, 강아지풀, 옥수수 등도 이 아과에 속하는데, 이들의 작은 이삭은 공통적으로 두 개의 작은 꽃(소화)만을 포함한다. 이 중 아래쪽에 위치한 첫 번째 작은 꽃은 퇴화하는 경향이 있으며, 위쪽의 두 번째 작은 꽃만이 결실성을 가진다. 첫 번째 작은 꽃은 수술만 가진 수꽃이 되기도 하지만, 꽃의 구조 자체가 퇴화하여 껍질 같은 영(穎)만 남는 경우가 많다. 이 때문에 작은 이삭은 비교적 단순하고 콤팩트한 형태를 띤다. 작은 이삭은 성숙하면 보통 포영(苞穎)에 싸인 채로 떨어져 퍼진다.[3]
작은 이삭을 구성하는 두 개의 작은 꽃은 모양이 다른 경우가 많으며, 이들을 붙이는 작은 축은 매우 짧아져 칼루스(callus)라고 불리는 구조가 된다. 포영이나 두 번째 작은 꽃의 호영(護穎, 바깥 꽃덮이) 중 어느 하나가 단단하게 발달하여 씨앗(열매)을 보호하는 역할을 한다. 작은 이삭 안에는 2개의 육질 인피(鱗被)와 보통 3개의 수술, 2개의 암술머리가 있으나 약간의 수 변화는 있을 수 있다. 열매인 영과는 큰 배를 가지는데, 배의 크기가 영과 전체 길이의 1/3에서 1/2에 달한다.[3] 이처럼 작은 이삭의 기본 구조는 공통적이지만, 포영과 호영의 발달 정도 등에 따라 세부 형태는 다양하게 나타난다. 이는 하위 분류군을 나누는 중요한 기준이 된다. (자세한 내용은 #작은 이삭의 구조 참조)
또한 기장아과 식물 중에는 C4 식물이 많아, 강한 햇빛이나 고온 환경에서도 효율적인 광합성을 할 수 있는 특징을 가진다. 이 아과에는 기장, 조, 옥수수, 사탕수수와 같이 인류에게 유용한 작물이 다수 포함되어 있을 뿐만 아니라, 바랭이나 돌피처럼 잡초로 분류되는 식물도 많아 우리 주변에서 흔히 볼 수 있는 식물들이 다수 속해 있다.
2. 1. 작은 이삭의 구조
벼과 식물의 작은 이삭은 기본적으로 여러 개의 작은 꽃이 축을 따라 배열되고, 각 작은 꽃은 바깥쪽의 호영(護穎)과 안쪽의 내영(內穎)이라는 두 장의 껍질 같은 구조물(영, 穎)로 싸여 있으며, 이 작은 꽃들의 맨 아래에는 두 장의 포영(苞穎)이 있는 구조를 가진다.[4] 이러한 기본 구조는 새포아풀속이나 갯그령속 등에서 잘 나타난다. 하지만 많은 벼과 식물에서는 이 구조의 일부가 퇴화하거나 변형된 모습을 보이며, 이는 분류에서 중요한 특징으로 여겨진다. 나가타(長田, 1993)는 이러한 변형을 몇 가지 유형으로 나누었는데, 기장아과에 속하는 식물들의 작은 이삭 구조는 주로 다음 세 가지 유형으로 구분할 수 있다.- 억새형 (Arundinella type): 포영 안쪽에 있는 작은 꽃 중 기부에 가까운 첫 번째와 두 번째 작은 꽃만 남고, 그 위쪽 부분은 퇴화하여 두 개의 작은 꽃만 남는다. 두 번째 작은 꽃은 암술과 수술을 모두 가진 양성(兩性)이며 그 밑부분에 털 뭉치가 있고, 첫 번째 작은 꽃은 수술만 가진 웅성(雄性)이거나 퇴화하여 털 뭉치가 없다. 억새가 이 유형에 속한다.
- 강아지풀형 (Setaria type): 억새형처럼 두 개의 작은 꽃만 남지만, 첫 번째 작은 꽃이 더욱 퇴화하여 거의 호영만 남은 형태가 된다. 두 번째 작은 꽃의 호영과 내영은 단단하게 각질화되어 씨앗(열매)을 보호하며, 첫 번째 작은 꽃의 호영이 이들을 다시 감싼다. 이 전체 구조의 바깥쪽에 첫 번째와 두 번째 포영이 있지만, 이들은 크게 발달하지 않고 작은 이삭의 밑부분만 감싸는 형태가 된다. 강아지풀이 대표적인 예이다.
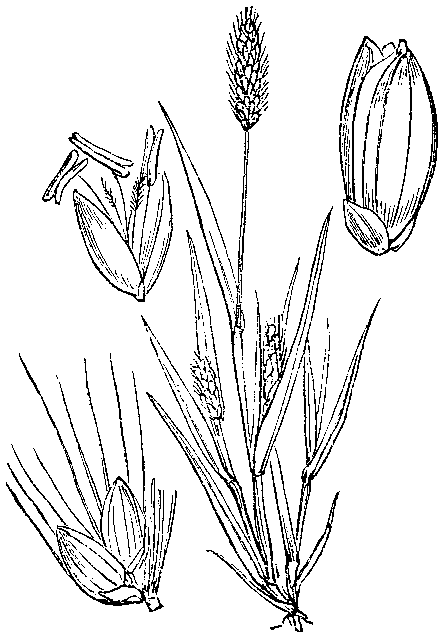
- 수수형 (Sorghum type): 강아지풀형과 비슷하게 첫 번째 작은 꽃은 호영만 남고, 두 번째 작은 꽃은 양성화로 호영과 내영을 모두 가진다. 하지만 이 유형에서는 두 번째 작은 꽃의 호영과 내영이 얇은 막질로 되어 단단하게 발달하지 않는다. 대신 이들을 감싸는 포영이 단단하고 두꺼워져 작은 이삭 전체를 감싸 보호하는 형태가 된다. 수수가 이 유형에 해당한다.
이처럼 기장아과의 작은 이삭은 '두 개의 작은 꽃만 포함하고, 첫 번째 작은 꽃이 퇴화하며 두 번째 작은 꽃이 양성화'라는 공통된 기본 구조를 가지면서도, 포영과 호영의 발달 정도와 역할에 따라 겉모습과 기능이 상당히 다양하게 나타난다.[5] 특히 수수형에서는 작은 이삭과 꽃차례의 겉모습이 매우 다양하여 구조를 파악하기 어려운 경우도 있다.[5] 그 대표적인 예가 옥수수인데, 옥수수는 작은 이삭에 암수의 구별이 있고, 수꽃 작은 이삭은 줄기 끝의 이삭(개꼬리)에, 암꽃 작은 이삭은 잎겨드랑이에서 나오는 이삭(옥수수 열매가 되는 부분)에 각각 모여 달리는 매우 특이한 구조를 보인다.
3. 생태 및 분포
세계에 널리 분포하며, 특히 열대 지방에 많은 종이 서식한다[6]. 분포 지역은 남극을 제외한 모든 대륙에 걸쳐 있다[7]. 전 세계적으로 200속 3,000종 이상이 알려져 있다[6]. 벼과 전체에는 약 700속 1만 종 이상이 있는데[8], 기장아과는 이 중 약 30%를 차지하며 벼과에서 가장 큰 아과 중 하나로 여겨진다[9].
4. 계통 분류
기장아과는 벼과(Poaceae) 내에서 PACMAD 분지군에 속하는 주요 아과 중 하나이다.[17][18] 벼과 내의 분류 체계는 역사적으로 많은 변화를 거쳐왔으나, 기장아과는 비교적 일찍부터 독자적인 그룹으로 인식되어 왔다. 1814년 로버트 브라운 (1773년)은 벼과를 기장족(Paniceceae)과 갯새족(Poaceae)으로 나누었는데, 이 당시 기장족의 범위는 현재의 기장아과와 상당히 유사했다.[12]
기장아과는 두 개의 작은 꽃으로 구성된 독특한 작은이삭 구조를 공통적으로 가지는 특징이 있다. 일반적으로 첫 번째 작은 꽃(제1소화)은 수술만 있거나 퇴화하여 포영(glume)만 남고, 두 번째 작은 꽃(제2소화)만이 양성화(암술과 수술을 모두 가짐)이다. 이러한 형태적 특징과 더불어 전분 입자의 성질 등 다른 형질들을 통해서도 기장아과는 전통적인 분류학 연구에서부터 단일계통군으로 인정받아 왔다.
20세기 후반 이후 분자계통학 연구가 활발해지면서 기장아과의 단일계통성은 더욱 명확하게 확인되었다.[17][18] 분자계통학적 분석 결과는 기장아과가 PACMAD 분지군 내에서 특정 아과들과 자매군 관계를 형성함을 보여준다. 또한, 분자 데이터는 기장아과 내부의 족 수준에서의 상세한 계통 관계를 밝히는 데 중요한 역할을 하고 있다. 현재 기장아과는 여러 족으로 세분되며, 이에 대한 자세한 내용은 하위 분류에서 다루어진다.
4. 1. 하위 분류
기장아과는 PACMAD 분지 내에서 Arundinoideae, Chloridoideae, Danthonioideae, Micrairoideae 4개 아과의 자매군이다. 현대의 계통분류학에서는 기장아과를 단일 계통 분지에 해당하는 12개의 족으로 나눈다.- 기네리움족 (Gynerieae Sánchez-Ken & L.G.Clark)
- 기장족 (Paniceae R.Br.)
- 수수족 (Andropogoneae Dumort.)
- 새족 (Arundinelleae Stapf)
- 스테이에르마르코클로아족 (Steyermarkochloeae Davidse & R.P.Ellis)
- 제우기테스족 (Zeugiteae Sánchez-Ken & L.G.Clark)
- 참새피족 (Paspaleae J.Presl)
- 카스만티움족 (Chasmanthieae W.V.Br. & B.N.Sm. ex Sánchez-Ken & L.G.Clark)
- 켄토테카족 (Centotheceae Ridl.)
- 키페로클로아족 (Cyperochloeae L.Watson & Dallwitz ex Sánchez-Ken & L.G.Clark)
- 트리스타키아족 (Tristachyideae Sánchez-Ken & L.G.Clark)
- 티사놀라이나족 (Thysanolaeneae C.E.Hubb.)
다음 두 속은 분류학적 위치가 명확히 정해지지 않았지만(''incertae sedis''), 트리스타키아족(Tristachyideae)에 속할 가능성이 있다.
- 얀세넬라속 (''Jansenella'' Bor)
- 칸드라세카라니아속 (''Chandrasekharania'' V.J.Nair, V.S.Ramach. & Sreek.)
주요 족인 기장족(Paniceae), 참새피족(Paspaleae), 수수족(Andropogoneae)은 새족(Arundinelleae)과 함께 아과의 핵심 그룹을 형성한다. 기네리움족(Gynerieae)은 과거 Arundinoideae 아과에 속했던 분류군이다. 또한, 7개의 족(카스만티움족, 제우기테스족, 스테이에르마르코클로아족, 트리스타키아족, 켄토테카족, 키페로클로아족, 티사놀라이나족)으로 구성된 기저 계통군인 "centothecoid clade"는 이전에는 Centothecoideae라는 별도의 아과로 간주되기도 했다.
엽록체 DNA 분석에 기반한 계통 발생 연구에 따르면, 아과 내 족들의 관계는 다음과 같이 추정된다. 기저 분지군("centothecoid clade") 내에서는 카스만티움족(Chasmanthieae)과 제우기테스족(Zeugiteae)이 자매군을 이루며 가장 먼저 분기한다. 나머지 centothecoid 족들 사이의 관계는 아직 완전히 명확하지 않으나, 스테이에르마르코클로아족(Steyermarkochloeae)과 트리스타키아족(Tristachyideae)(C4 광합성 수행)이, 그리고 켄토테카족(Centotheceae)과 키페로클로아족(Cyperochloeae)-티사놀라이나족(Thysanolaeneae) 그룹이 각각 자매군 관계일 가능성이 있다. 이 centothecoid clade 전체는 기네리움족(Gynerieae)과 자매군 관계이며, 이들은 핵심 기장아과 그룹(Andropogonodae)과 분기된다. 핵심 그룹 내에서는 기장족(Paniceae)(C3 및 C4 광합성 수행)이 가장 먼저 분기하고, 참새피족(Paspaleae)(C3 및 C4 광합성 수행)과 수수족(Andropogoneae)(C4)-새족(Arundinelleae)(C4) 그룹이 자매군을 이룬다. Lecomtelleae 족의 정확한 위치는 아직 불확실하다.
기장아과를 하위 분류하려는 시도는 1881년 벤담(Bentham)이 6개의 군(group)을 설정하면서 시작되었으나, 20세기를 거치면서 여러 분류 체계가 제안되고 폐기되기를 반복했다.[13] 과거에 비교적 널리 받아들여졌던 분류 체계 중 하나는 다음 7개의 군으로 나누는 방식이었다(괄호 안은 속의 수).[14]
상기 분류 중 가장 큰 두 군이었던 Andropogoneae와 Paniceae는 현재의 족 분류(수수족, 기장족)에서도 주요 그룹으로 인정받고 있으나, 분자 계통 정보에 기반한 세부적인 분류는 계속해서 연구 및 재검토가 이루어지고 있다.[15]
5. 인간과의 관계
기장아과는 인류에게 매우 중요한 식물군이다. 세계 3대 곡물 중 하나인 옥수수를 비롯하여 기장, 조, 수수 등 많은 작물이 이 아과에 속하며, 이들은 주요 식량 자원이자 잡곡으로 널리 재배된다. 특히 한국에서도 옥수수, 수수 등은 중요한 식량 및 사료 작물로 이용된다. 곡물 외에도 설탕의 주요 원료인 사탕수수 역시 기장아과에 속하며, 여러 종이 목초나 잔디로 활용되는 등 경제적으로 중요한 역할을 한다.
문화적으로도 기장아과 식물은 친숙하다. 가을의 7초 중 하나인 억새와 띠는 한국의 가을 풍경을 대표하는 식물로 여겨진다. 특히 띠는 과거 지붕을 이는 재료나 생활용품 제작에 사용되기도 했다.
하지만 모든 기장아과 식물이 인간에게 유익한 것은 아니다. 바랭이나 돌피와 같은 종들은 농경지에서 자라며 작물과 경쟁하는 잡초로 분류된다. 또한 털물괭이처럼 외래종으로 유입되어 생태계를 교란하고 생물 다양성을 위협하는 경우도 있다.
참조
[1]
서적
Guide to plant families of southern Africa
https://www.worldcat[...]
2014
[2]
문서
以下、主として大橋ほか編 (2016), p. 77
[3]
문서
Grass Phylogeny Working Group (2001), p.423
[4]
문서
以下、長田 (1993), pp. 23–26
[5]
문서
長田 (1993), p. 25
[6]
문서
大橋ほか編 (2016), p. 77
[7]
문서
Giussani et al. (2001), p. 1993
[8]
문서
Grass Phylogeny Working Group (2001), p. 374
[9]
문서
Teerawatananon (2011), p. 116
[10]
문서
Giussani et al. (2001), p. 1994
[11]
문서
Giussani et al. (2001), p. 2003
[12]
문서
以下もGrass Phylogeny Working Group (2001), pp. 374–375
[13]
문서
以下、Giussani et al. (2001), p. 1993
[14]
문서
大橋ほか編 (2016)、木場ほか (2011)、茨木ほか (2020) から
[15]
문서
Teerawatananon et al. (2009)
[16]
문서
Braun, Alexander Karl Heinrich. Flora der Provinz Brandenburg 1: 799. 1864.
[17]
저널
New grass phylogeny resolves deep evolutionary relationships and discovers C4 origins
2012
[18]
저널
A worldwide phylogenetic classification of the Poaceae (Gramineae)
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com