스페나코돈과
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
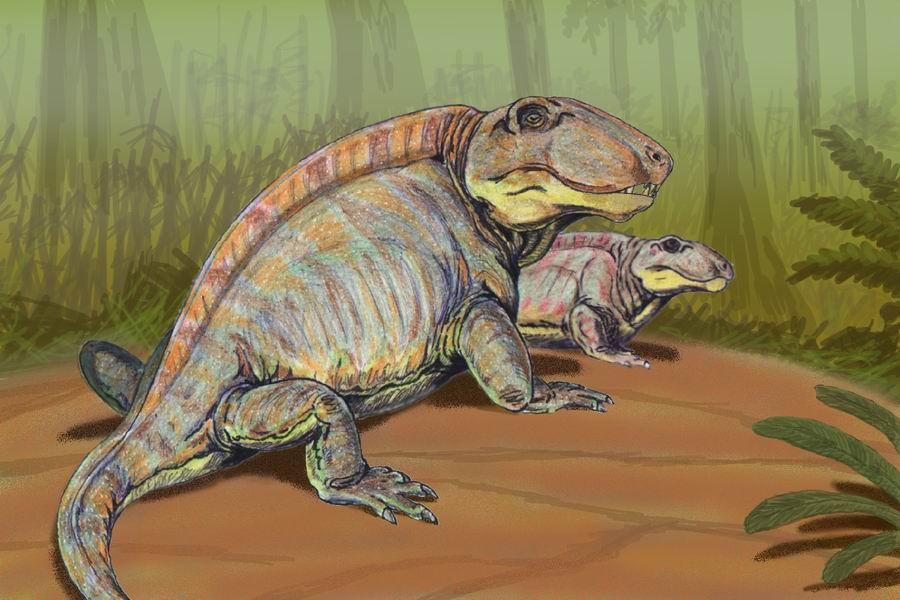
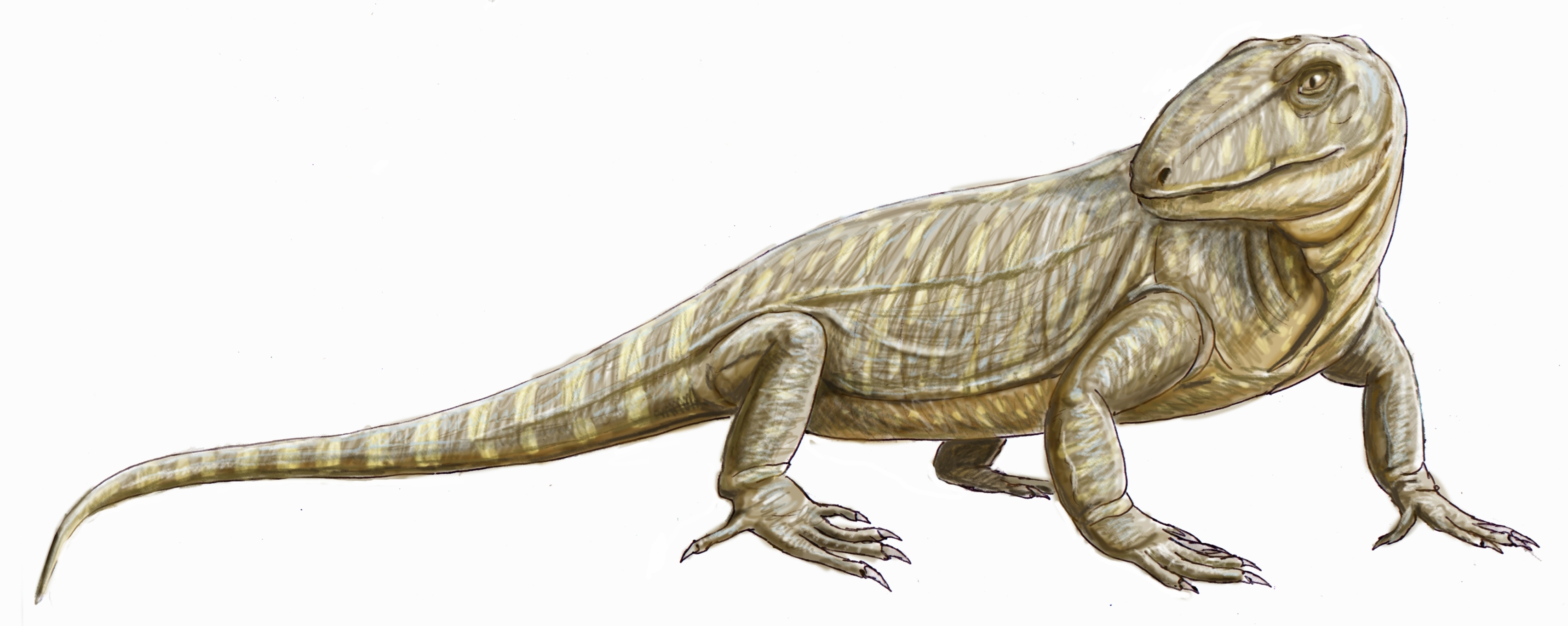
스페나코돈과는 페름기 전기 쇠퇴해가는 오피아코돈과를 대신하여 등장한 육식 단궁류의 한 무리이다. 이들은 강력한 턱과 이빨을 가지고 있었으며, 일부는 등줄기를 따라 솟아있는 돛을 가지고 있었다. 스페나코돈과는 북아메리카와 유럽에서 화석이 발견되었으며, 척추 신경극이 신장된 특징을 보인다. 스페나코돈과는 좁은 의미로 재정의되어 수궁류로 이어지는 계통과 분기된 이후의 단궁류만을 포함하며, 디메트로돈, 스페나코돈, 스테페사우루스 등이 주요 속에 속한다.

더 읽어볼만한 페이지
- 스페나코돈과 - 켄마그나투스
켄마그나투스는 석탄기에 살았던 초기 단궁류로, 높은 코와 뭉툭한 이빨을 가지고 이매패류를 주식으로 삼았을 것으로 추정되며, 화석은 미국 캔자스주에서 발견되었다. - 스페나코돈과 - 마크로메리온
마크로메리온은 석탄기 후기에 유럽 초원과 산림 지대에 서식했던 스페나코돈티드과의 육식성 파충류로, 등에 돛 모양의 구조가 발달했고 튼튼한 다리와 긴 꼬리를 가졌으며, 1875년 화석이 처음 발견되었다. - 오스니얼 찰스 마시가 명명한 분류군 - 트리케라톱스
트리케라톱스는 북아메리카에서 발견된 각룡류 공룡의 한 속으로, 거대한 두개골과 뿔이 특징이며 백악기 말기에 번성했고, 현재는 두 종으로 분류된다. - 오스니얼 찰스 마시가 명명한 분류군 - 프테라노돈
프테라노돈은 백악기 후기 북아메리카에서 서식한 이빨 없는 익룡 속으로, 날개폭이 최대 9미터에 달하며 수컷은 큰 머리 볏을 가지고 있고 주로 물고기를 잡아먹었을 것으로 추정된다.

2. 특징
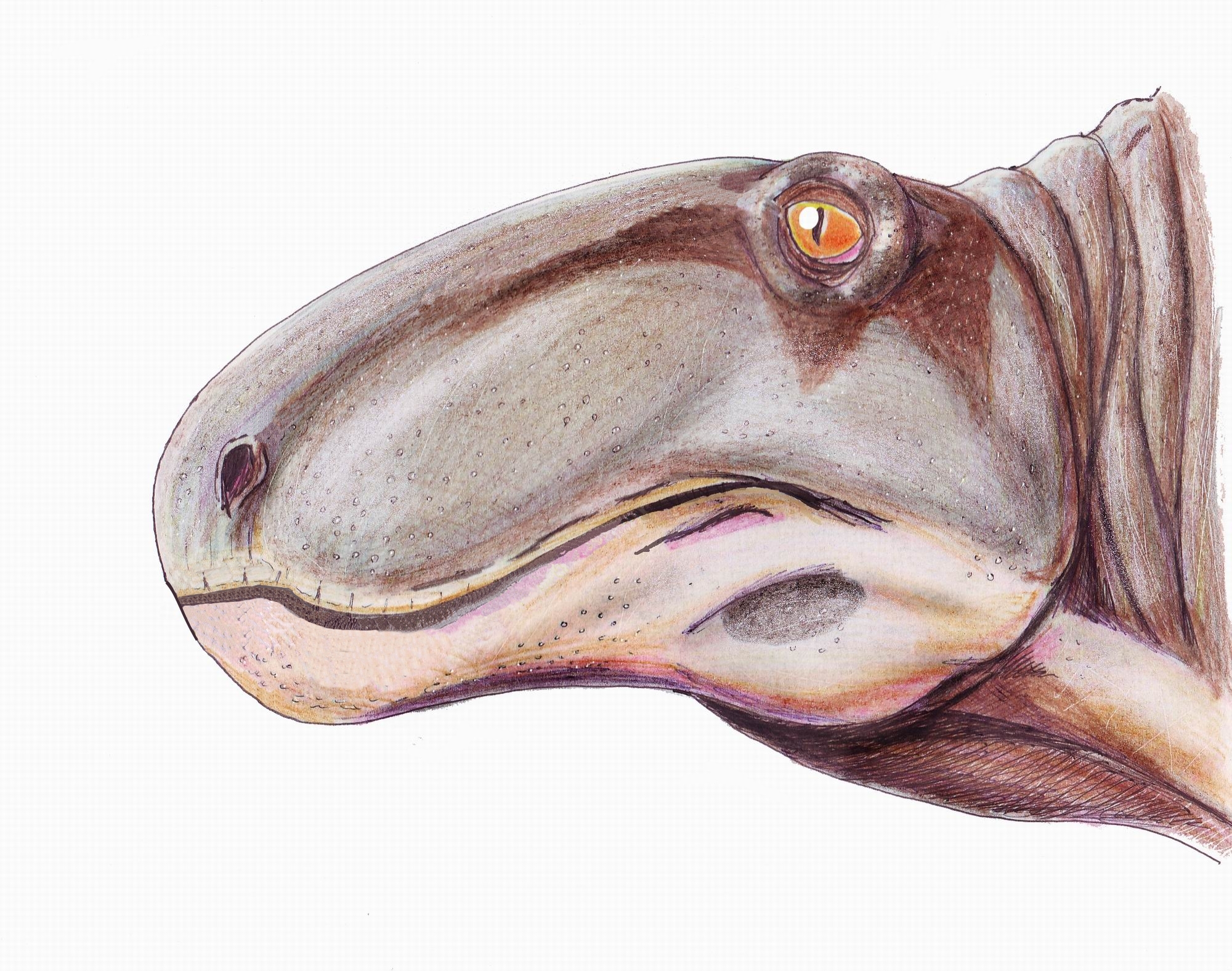
두개골은 길고 깊으며 좁은 형태를 띠는데, 이는 강력한 턱 근육에 적응한 것이다. 앞니는 크고 칼날과 같은 형태인 반면, 턱의 측면과 뒤쪽의 이빨은 훨씬 작다 (유명한 속인 ''디메트로돈''의 이름이 "두 개의 크기의 이빨"인 이유도 여기에 기인하며, 이는 이 과의 모든 구성원들이 갖는 특징이다).[9]
이 그룹의 몇몇 크고 (~3미터) 진보된 구성원들 (''크테노스폰딜루스'', ''스페나코돈'', ''세코돈토사우루스'' 및 ''디메트로돈'')은 등줄기를 따라 솟아있는 높은 돛으로 구별되는데, 이는 길어진 척추의 신경 배돌기로 이루어져 있으며, 살아있을 때는 피부와 혈관으로 덮여 있었을 것이고, 아마도 체온 조절 장치로 기능했을 것이다.[8] 그러나 돛을 갖는 것이 이러한 동물들에게 필수적인 것은 아니었던 것으로 보인다. 예를 들어, 한 속 (''스페나코돈'' - 뉴멕시코에서 화석이 발견됨)은 돛이 없는 반면, 매우 유사하고 밀접하게 관련된 속 (''디메트로돈'' - 텍사스에서 화석이 발견됨)은 돛을 가지고 있는 경우가 있다. 페름기 동안, 이 두 지역은 좁은 해로로 분리되어 있었지만, 한 지리적 고립된 집단은 왜 돛을 진화시키고 다른 집단은 그렇지 않았는지는 불분명하다.
;두개골
: 크고 높이가 있으며 튼튼하다. 이는 턱을 크게 벌리고 또한 큰 힘으로 닫기 위해 저작근이 발달한 것에 대응하는 적응 형태라고 여겨진다.[5] 이를 통해 그들은 자신과 동등한 크기의 먹이를 사냥할 수 있었다고 생각된다.[5] 전상악골은 발달한 이빨 열을 수용하기 위해 복측으로 폭이 넓어졌다. 위턱에 송곳니 모양의 이빨이 발달하고, 치근이 내비공 측으로 튀어나와 있다. 또한, 아래턱 선단부의 발달한 이빨은 상악골의 틈새에 들어맞도록 되어 있다. 측두창[6]은 확대되어 하악내전근의 부착부가 확대되어 저작이 효율적으로 이루어졌음을 알 수 있다. 또한, 아래턱에는 얇은 판 모양의 뼈인 반전판이 각골[7]에 부착되어 있다. 이것은 지면이나 공기 중의 소리를 감지하기 위한 픽업으로 사용되었다고 여겨진다.[8]
;치열
: 절치는 길고 날카롭다. 또한, 송곳니도 크게 발달해 있다. 한편, 후부의 이빨은 축소되어 어금니에 가까운 형태가 된다. "2종류의 긴 이빨" - 디메트로돈의 이름에 나타나듯이, '''이형성'''(이빨이 나는 위치에 따라 그 형태가 다른 것)이 발달했다. 그들은 먹이를 통째로 삼키지 않고, 이 이빨로 고기를 찢어 먹고, 잘게 썰어 먹는다는 포유류의 포식자와 공통되는 저작 형태를 얻은 최초의 생물이었다.[9]
;중이
: 등골[10]에 판 모양의 배측 돌기가 발달해 있다.[11]
;척추
: 전체적으로 척추의 신경극이 신장하는 경향이 있다. 디메트로돈, 크테노스폰딜루스, 세코돈토사우루스 등 대형 속에서 현저하지만, 스페나코돈 등에서도 돌기의 길이가 척추골의 5배를 넘고 있다.[12]
;돛
: 길게 신장된 극돌기 사이에는 피부 막이 있어 돛을 형성하고 있었다고 여겨진다. 이 돛의 사용 목적은 오랫동안 불분명했다. 과거에 주장된 설 중 하나는, 돛이 있는 디메트로돈이 수컷, 돛이 없는 스페나코돈이 암컷이며, 성적 이형태를 나타내는 것이라고 여겨졌다. 그러나, 디메트로돈은 텍사스주 산, 스페나코돈은 뉴멕시코주 산이며, 당시 이 두 곳의 산지는 좁은 내해에 의해 격리되어 있었다. 또한 두개골의 형태에도 차이가 있는 등, 이 둘은 별개의 속으로 간주하여 이 설은 부정되었다.[13] 후에 이러한 돌기가 전후 방향으로 넓어져 있고, 뼈의 표면에 혈관의 흔적이 발견된 것 등으로 미루어보아, 아마 생존 시에는 이 돛에 혈액이 대량으로 보내지는 것처럼 되어 있었고, 열교환기로 기능했을 것으로 여겨진다. 체온 조절 기능을 갖지 못한 그들은, 태양광을 이 돛에 받음으로써 혈액을 따뜻하게 하고, 먹이가 제대로 움직이지 못하는 사이에 포식했다고 여겨진다. 또한, 체온이 너무 올라가는 낮에는, 이 돛에 바람을 받음으로써 열을 식혔다고 한다. 그러나, 돛을 갖지 않은 단궁류도 번성하고 있으며, 이러한 돛을 갖는 것이 생존에 유리했는지는 단정할 수 없다. 사실, 단궁류의 후계자인 수궁류는, 돛을 갖지 않은 그룹의 자손이기 때문이다.[8] 선행하는 에다포사우루스과도 이러한 돛을 가지고 있지만, 양자는 특히 근연 관계가 있는 것은 아니며, 모두 돛을 갖지 않은 조상으로부터 진화하여, 각각이 획득한 수렴 진화에 의한 것이다.[14] 스페나코돈과의 돛에는 에다포사우루스와 같은 횡돌기가 없다. 아마도 양자 모두, 체온 조절과 동시에 성적 과시용으로도 이 돛을 사용했다고 여겨진다.
;사지
: 사지는 가늘고 길어져서, 당시의 생물로서는 민첩한 동작이 가능했다.
2. 1. 두개골
크고 높이가 있으며 튼튼한 구조로, 저작근이 발달하여 턱을 크게 벌리고 큰 힘으로 닫을 수 있었다.[5] 이를 통해 자신과 비슷한 크기의 먹이를 사냥할 수 있었을 것으로 추정된다. 전상악골은 발달한 이빨 열을 수용하기 위해 복측으로 넓어졌으며, 위턱에는 송곳니 모양의 이빨이 발달하고 치근이 내비공 측으로 튀어나와 있다. 아래턱 선단부의 발달한 이빨은 상악골 틈새에 들어맞도록 되어 있다. 측두창[6]은 확대되어 하악내전근의 부착부가 확대되어 저작 효율을 높였다.[5] 또한, 아래턱에는 얇은 판 모양의 뼈인 반전판이 각골[7]에 부착되어, 지면이나 공기 중의 소리를 감지하는 데 사용되었을 것으로 추정된다.[8]2. 2. 치열
스페나코돈과의 절치는 길고 날카로우며, 송곳니도 크게 발달했다.[9] 한편, 후부 이빨은 축소되어 어금니에 가까운 형태를 보인다.[9] "2종류의 긴 이빨"이라는 디메트로돈의 이름에서 알 수 있듯이, '''이형성'''(이빨이 나는 위치에 따라 형태가 다른 것)이 발달했다.[9] 스페나코돈과는 이빨을 이용해 먹이를 통째로 삼키지 않고 고기를 찢고 잘게 썰어 먹는, 포유류의 포식자와 공통되는 저작 형태를 얻은 최초의 생물이었다.[9]2. 3. 중이
등골에 판 모양의 배측 돌기가 발달해 있다.2. 4. 척추
스페나코돈과는 척추의 신경 배돌기가 길어져 솟아있는 높은 돛을 가진 것이 특징이다. 이는 체온 조절 장치로 기능했을 것으로 추정된다. 크테노스폰딜루스, 스페나코돈, 세코돈토사우루스 및 디메트로돈 등이 이러한 특징을 보인다. 하지만, 뉴멕시코에서 화석이 발견된 스페나코돈처럼 돛이 없는 경우도 있고, 텍사스에서 화석이 발견된 디메트로돈과 같이 돛을 가지고 있는 경우도 있다. 페름기 동안 두 지역은 좁은 해로로 분리되어 있었는데, 한 지리적으로 고립된 집단에서 돛이 발달하고 다른 집단은 그렇지 않은 이유는 불분명하다.2. 5. 돛
스페나코돈과의 일부 대형(~3미터) 구성원들은 등줄기를 따라 솟아있는 높은 돛을 가지고 있다.[12] 돛은 길게 신장된 척추의 신경 배돌기로 이루어져 있으며, 살아있을 때는 피부와 혈관으로 덮여 있어 체온 조절 장치로 기능했을 것으로 추정된다.[8]
돛의 기능은 체온 조절 외에도 성적 과시 용도로도 사용되었을 것으로 추정되지만, 돛이 없는 단궁류도 번성하였고, 돛을 갖는 것이 생존에 필수적인 것은 아니었다.[8] 뉴멕시코에서 화석이 발견된 ''스페나코돈''은 돛이 없는 반면, 텍사스에서 화석이 발견된 매우 유사하고 밀접하게 관련된 속인 ''디메트로돈''은 돛을 가지고 있다.[13] 페름기 동안 두 지역은 좁은 해로로 분리되어 있었지만, 한 지리적 고립된 집단은 돛을 진화시키고 다른 집단은 그렇지 않은 이유는 불분명하다.
에다포사우루스과도 돛을 가지고 있지만, 스페나코돈과와는 수렴 진화의 결과로 돛을 획득한 것이며, 스페나코돈과의 돛에는 에다포사우루스와 같은 횡돌기가 없다.[14]
3. 진화사
스페나코돈과는 쇠퇴해 가는 오피아코돈과를 대신하여 페름기 전기에 대두한 그룹이다. 초기에는 하프토두스(Haptodus) 등 소형 곤충을 먹는 종류가 주를 이루었으나, 페름기 전기 후반에는 대형화되어 강력한 육식 동물로 진화했다. 이들은 같은 크기의 먹이를 사냥하는 육상 최초의 포식자였으며, 당시 육상 생태계의 정점에 서는 최상위 포식자였다.
페름기 전기 말까지 전성기를 누렸으나, 페름기 중기에 급격히 쇠퇴하여 화석이 거의 발견되지 않는다. 이는 당시 일어난 환경 격변의 영향으로 보이며, 다른 많은 단궁류, 양서류, 파충류와 함께 페름기 중기 초반에 모습을 감춘 것으로 보인다. 스페나코돈과의 쇠퇴 및 멸종과 함께, 근연 관계인 수궁류가 급속히 다양화되어 생태계의 틈새를 메우며 대두하게 된다.
4. 분포
5. 분류
'''스페나코돈과'''는 원래 설명된 대로 실제로는 측계통군으로, 공유된 단궁류 특징에 의해 정의된다. 이 동물들은 원시적인 단궁류에서 초기 수궁류에 이르는 진화적 단계를 구성한다.
분기도 '''스페나코돈류'''는 스페나코돈과와 모든 후손(포유류 포함)을 포함하는 단계통군을 지정하는 데 사용되는 반면, 엄밀한 의미의 스페나코돈과는 ''햘토두스'', ''Palaeohatteria'', ''Pantelosaurus'', ''Cutleria''와 같은 과의 이전의 원시 구성원을 제외하고 전문화된 페름기를 포함한다.
가장 최근의 공통 조상인 스페나코돈과와 수궁류 및 모든 후손을 지정하기 위해 '''스페나코돈상과'''가 사용되었으며, 두개골의 특정 특징으로 정의된다.[15]
전통적으로 정의된 "스페나코돈과"는 수궁류의 조상을 포함하기 때문에 측계통이 된다. 이 그룹 내에는 초기 단궁류부터 수궁류에 이르는 진화의 다양한 단계에 있는 생물을 포함했다. 분지학에서 정의된 스페나코돈류(Sphenacodontia)라고 불리는 클레이드는 스페나코돈류 및 그들의 자손(수궁류, 포유류)을 모두 포함한다. 스페나코돈과는 수궁류로 이어지는 계통과 분기된 이후의 단궁류만을 포함하는 그룹으로 엄밀하게 재정의되었다. 햘토두스 등 조상적인 형태를 유지한 속은 더 초기에 분기되었다고 하여 좁은 의미의 스페나코돈과에서는 제외되었다. 또한 스페나코돈상과(Sphenacodontoidea)는 스페나코돈과와 수궁류의 공통 조상 및 그 자손 모두를 포함하는 클레이드로, 라우린과 라이스(1997)에 의해 두개골의 형태에 따라 정의되었다.[15]
- 단궁류(Synapsid)
- * 진단궁아목(Eupelycosauria)
- ** 오피아코돈과(Ophiacodontidae)
- ** 에다포사우루스과(Edaphosauridae)
- ** 스페나코돈류(Sphenacodontia)
햘토두스(Haptodus) 속
스페나코돈상과(Sphenacodontoidea)
* '''스페나코돈과(Sphenacodontidae)'''
* 테트라케라톱스과(Tetraceratopsidae) : 테트라케라톱스
* 수궁류(Therapsida)
** 포유류(Mammalia)

디메트로돈, 스페나코돈, 스테페사우루스, 크테노스폰딜루스, 세코돈토사우루스 등이 스페나코돈과에 속한다.
5. 1. 주요 속
(Dimetrodon)(Sphenacodon)
(Steppesaurus)
디메트로돈(Dimetrodon), 스페나코돈(Sphenacodon), 스테페사우루스(Steppesaurus), 크테노스폰딜루스(Ctenospondylus), 세코돈토사우루스(Secodontosaurus) 등이 스페나코돈과에 속한다.
참조
[1]
논문
Late Permian terrestrial vertebrates, U.S.A. and U.S.S.R.
https://www.jstor.or[...]
1962
[2]
논문
The age of North America’s youngest Paleozoic continental vertebrates: a review of data from the Middle Permian Pease River (Texas) and El Reno (Oklahoma) Groups
https://doi.org/10.1[...]
2022
[3]
논문
Distributions of extinction times from fossil ages and tree topologies: the example of some mid-Permian synapsid extinctions
2021
[4]
논문
A new basal sphenacodontid synapsid from the Late Carboniferous of the Saar-Nahe Basin, Germany
https://bioone.org/j[...]
[5]
서적
『脊椎動物の進化』
[6]
문서
眼窩後方にある顎の筋肉を収める穴。
[7]
문서
下顎を構成していた骨の一つ。哺乳類では[[鼓胞]]の一部になる。
[8]
서적
『哺乳類型爬虫類 : ヒトの知られざる祖先』
[9]
서적
『翼竜の謎 : 翼竜・魚竜・首長竜の時代』
[10]
문서
哺乳類の[[鐙骨]]と[[相同]]。
[11]
서적
『哺乳類型爬虫類 : ヒトの知られざる祖先』
[12]
서적
『哺乳類型爬虫類 : ヒトの知られざる祖先』
[13]
서적
『脊椎動物の進化』
[14]
서적
『消えた竜 : 哺乳類の先祖についての新しい考え』
[15]
웹사이트
Synapsida: Sphenacodontia
http://www.palaeos.c[...]
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com