필석
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
필석은 린네가 명명한 멸종된 해양 생물 그룹으로, 고생대 시대에 번성했다. 필석은 군집을 이루는 형태로 살았으며, 다양한 형태의 껍데기 화석으로 발견된다. 필석 화석은 표준 화석으로, 지질 시대와 퇴적 환경을 파악하는 데 중요한 정보를 제공한다. 필석은 수지상필석목과 필석목으로 분류되며, 1970년대 이후 익새류와 밀접한 관련이 있는 것으로 여겨진다. 필석의 생태, 이동 방식, 생명 주기, 그리고 진화 발달 생물학에서의 중요성에 대한 연구가 이루어지고 있으며, 한국에서도 캄브리아기-오르도비스기 지층에서 화석이 발견된다.

더 읽어볼만한 페이지

2. 역사
필석은 오래전부터 그 존재가 알려졌지만, 처음에는 생물이 아닌 돌의 한 형태로 생각되었다. 이 때문에 "그래프톨라이트"와 같이 광물과 비슷한 이름이 붙었다. 1735년, 린네는 필석을 무생물일 가능성이 높다고 보면서도, Graptolithusla라는 학명을 붙여 속으로 취급했다. 1751년 린네는 『스카네 기행』에서 "기묘한 화석, 혹은 필석"에 대해 언급했으며, 1768년에는 저서 『자연의 체계』 제12권에서 G. sagittariusla와 G. scalarisla 두 종을 언급하며 "식물 같기도 하고, 필석(무생물) 같기도 하다"라고 설명했다.
이후 연구자들은 필석과 유사한 생물군을 Graptolithusla로 부르기도 했다. Graptolithusla가 정식으로 폐지된 것은 1954년 국제 동물 명명 규약에서였으며, 이는 Graptolithusla가 원래 무생물을 지칭하는 용어였기 때문이다.
1970년대 전자 현미경의 발달로 필석은 익새강(Pterobranchiala)에 가까운 반삭동물의 일종으로 여겨지게 되었다. 익새강에는 현존하는 종도 있으며, 두개원반충속(Cephalodiscusla) 20종과 가시원반충속(Rhabdopleurala) 8종이 알려져 있다. 두개원반충속은 1882년에 처음 발견되었다.
2. 1. 어원
"필석"이라는 이름은 1735년 린네가 실제 화석과 유사한 무기적 광물화 및 부착물을 지칭하기 위해 사용한 속인 ''Graptolithus''("바위에 쓴 글자")에서 유래되었다.[1] 1768년, ''자연의 체계'' 12권에서 그는 각각 식물 화석일 가능성이 있는 ''G. sagittarius''와 필석일 가능성이 있는 ''G. scalaris''를 포함시켰다.[1] 1751년 저서 ''Skånska Resa''에서 그는 현재 ''Climacograptus''(이중렬 필석 속)의 한 종류로 여겨지는 "이상한 종류의 화석 또는 필석"의 그림을 포함시켰다.[1]필석 화석은 이후 이끼벌레와 히드로충과 같은 다른 분지형 군체 동물들을 포함한 다양한 그룹으로 언급되었다.[1] Graptolithina이라는 용어는 1849년 Bronn에 의해 설립되었으며, 그는 이들을 정원뿔 두족류를 나타내는 것으로 간주했다.[1] 20세기 중반까지, 필석은 19세기 후반에 기술된 속인 ''Rhabdopleura''와 ''Cephalodiscus''에 속하는 살아있는 익새류와 밀접한 관련이 있는 독특한 그룹으로 인식되었다.[1] 속으로서의 ''Graptolithus''는 1954년 ICZN에 의해 공식적으로 폐기되었다.[1]
필석의 존재는 오래전부터 알려졌지만, 처음에는 생물이 아닌 돌의 한 형태라고 생각되었다.[1] 이 때문에 "그래프톨라이트"와 같이 광석과 같은 이름이 붙었다.[1] 1735년, 린네는 이를 무생물일 가능성이 높다고 하면서도, Graptolithusla 라는 학명을 붙여 속으로 취급했다.[1] 린네는 1768년, 저서 『자연의 체계』(''Systema Naturae|Systema Naturae영어'')의 제12권에서 G. sagittariusla, G. scalarisla의 2종을 채록하고, "식물과 같기도 하고, 필석(무생물)과 같기도 하다"라고 해설했다.[1] 린네는 1751년에 출판한 『스카네 기행』(''Skånska Resasv'')에서도 "기묘한 화석, 혹은 필석"에 대해 언급하고 있다(린네가 발견한 것은 현재 biserial graptolites영어에 속하는 필석으로 여겨진다).[1] 이후의 연구자들은 이것과 유사한 생물 군을 부르는 데 Graptolithusla의 이름을 사용했다.[1] Graptolithusla가 정식으로 폐지된 것은 1954년의 국제 동물 명명 규약에서이며, 그 이유는 Graptolithusla가 원래 무생물을 지칭하기 위한 용어였기 때문이었다.[1]
1970년대에 전자 현미경이 발달하면서, 필석은 익새강(Pterobranchiala)에 가까운 생물이며, 반삭동물에 속하는 특수한 생물로 여겨지게 되었다.[1] 익새강에는 현존종도 있으며, 두개원반충속(Cephalodiscusla) 20종, 가시원반충속(Rhabdopleurala) 8종이 알려져 있다.[1] 두개원반충속은 1882년에 처음 발견되었다.[1]
2. 2. 연구사
"필석"이라는 이름은 1735년 린네가 실제 화석과 유사한 무기적 광물화 및 부착물을 지칭하기 위해 사용한 속인 ''Graptolithus''("바위에 쓴 글자")에서 유래되었다.[1] 1768년, ''자연의 체계'' 12권에서 그는 각각 식물 화석일 가능성이 있는 ''G. sagittarius''와 필석일 가능성이 있는 ''G. scalaris''를 포함시켰다.[1] 1751년 저서 ''Skånska Resa''에서 그는 현재 ''Climacograptus''(이중렬 필석 속)의 한 종류로 여겨지는 "이상한 종류의 화석 또는 필석"의 그림을 포함시켰다.[1]필석 화석은 이후 이끼벌레와 히드로충과 같은 다른 분지형 군체 동물들을 포함한 다양한 그룹으로 언급되었다.[1] Graptolithina이라는 용어는 1849년 Bronn에 의해 설립되었으며, 그는 이들을 정원뿔 두족류를 나타내는 것으로 간주했다.[1] 20세기 중반까지, 필석은 19세기 후반에 기술된 속인 ''Rhabdopleura''와 ''Cephalodiscus''에 속하는 살아있는 익새류와 밀접한 관련이 있는 독특한 그룹으로 인식되었다.[1] 속으로서의 ''Graptolithus''는 1954년 ICZN에 의해 공식적으로 폐기되었다.[1]
필석의 존재는 오래전부터 알려졌지만, 처음에는 생물이 아닌 돌의 한 형태라고 생각되었다.[2] 이 때문에 "그래프톨라이트"와 같이 광석과 같은 이름이 붙었다.[2] 1735년, 린네는 이를 무생물일 가능성이 높다고 하면서도, Graptolithusla 라는 학명을 붙여 속으로 취급했다.[2] 린네는 1768년, 저서 『자연의 체계』(''Systema Naturae|'')의 제12권에서 G. sagittariusla, G. scalarisla의 2종을 채록하고, "식물과 같기도 하고, 필석(무생물)과 같기도 하다"라고 해설했다.[2] 린네는 1751년에 출판한 『스카네 기행』(''Skånska Resasv'')에서도 "기묘한 화석, 혹은 필석"에 대해 언급하고 있다(린네가 발견한 것은 현재 biserial graptolites영어에 속하는 필석으로 여겨진다).[2] 이후의 연구자들은 이것과 유사한 생물 군을 부르는 데 Graptolithusla의 이름을 사용했다.[2] Graptolithusla가 정식으로 폐지된 것은 1954년의 국제 동물 명명 규약에서이며, 그 이유는 Graptolithusla가 원래 무생물을 지칭하기 위한 용어였기 때문이었다.[2]
1970년대에 전자 현미경이 발달하면서, 필석은 익새강(Pterobranchiala)에 가까운 생물이며, 반삭동물에 속하는 특수한 생물로 여겨지게 되었다.[2] 익새강에는 현존종도 있으며, 두개원반충속(Cephalodiscusla) 20종, 가시원반충속(Rhabdopleurala) 8종이 알려져 있다.[2] 두개원반충속은 1882년에 처음 발견되었다.[2]
3. 형태
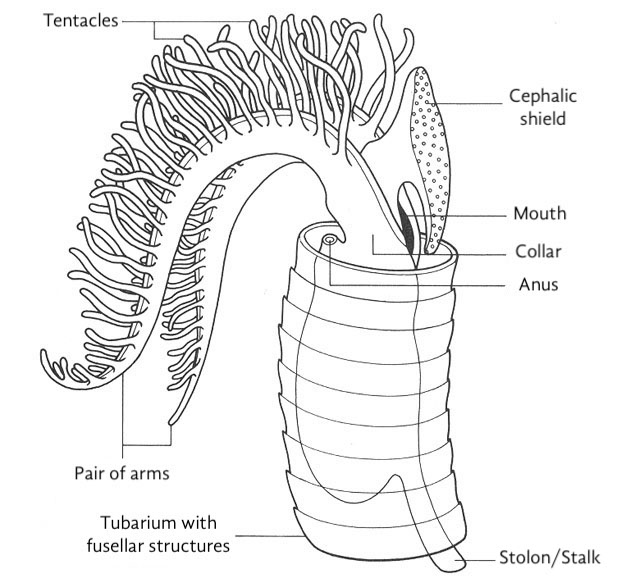
필석은 여러 개체가 모여 군체를 이루는 생물로, 껍데기로 덮여 있다는 점에서 산호나 이끼벌레와 유사하다. 필석 화석은 이 군집의 껍데기가 남은 것이다. 군집은 시큘라(sicula)라고 불리는 껍데기를 가진 하나의 개체에서 시작하여, 이 개체가 번식하면서 주위에 여러 개체들이 생겨나 군집을 형성한다. 각각의 개체가 자리 잡은 껍데기를 포벽(theca)이라 하며, 포벽들이 이어져 하나의 줄을 이루면 이를 스타이프(stipe)라고 한다.[22]
포벽 안에 사는 필석 개체를 개충(zooid)이라고 하는데, 보통 껍데기만 발견되기 때문에 개충에 대해 알려진 바는 많지 않다. 화석화된 개충이 보고된 적은 있으나, 신체 구조를 자세히 알 수 있을 만큼 보존 상태가 좋은 것은 드물다.[26] 따라서 필석 개충의 모델은 대부분 간벽충을 바탕으로 한다.

필석 군체는 rhabdosome영어이라고 불리며, sicula영어라고 불리는 초기 개체에서 시작하여 stipes영어(줄기)라고 불리는 다른 개체들이 이어진다. 각각의 개체는 theca영어라고 불리는 관 또는 작은 용기 형태를 하고 있으며, 이 군체의 분기나 배열 상태가 필석의 종류를 구별하는 중요한 단서가 된다.
수목형, 혹은 가지가 많은 화석은 대부분 목에 분류되는 덴드로이드 필석이다. 덴드로이드 필석은 캄브리아기에 등장하여 수저에 뿌리 같은 것으로 붙어 있었다. 가지가 적은 목은 오르도비스기 초기에 등장하여, 원양의 해면을 자유롭게 떠돌아다니거나 해초 등에 감겨 있었다. 목은 데본기 초기에 멸망할 때까지 동물성 플랑크톤으로서 번성하였고, 덴드로이드 필석은 석탄기 초까지 살아남았다.
3. 1. 군체 구조
필석은 마치 산호나 이끼벌레처럼 여러 개체가 껍데기로 덮힌 군집을 이루고 산다. 필석 화석은 이 군집 중에서도 껍데기가 남은 것이다.군집은 하나의 개체에서 시작하는데, 이 개체의 껍데기를 시큘라(sicula)라고 한다. 시큘라의 오목한 쪽으로 마치 꼬리와 같이 가느다란 구조물이 나기도 하는데, 이는 네마(nema)라고 한다.[22]
시큘라의 개체가 번식하면서 그 주위로 개체들이 하나둘씩 생겨나가고, 군집이 형성된다. 하나의 개체가 자리잡고 있는 껍데기를 포벽(theca, 복수형 thecae)이라고 하고, 포벽들이 나란히 이어져 하나의 줄을 이룰 때 그 줄을 스타이프(stipe)라고 한다.[22] 껍데기 속에 사는 각각의 개체는 일종의 줄기(stolon)로 이어져 있다고 생각된다.
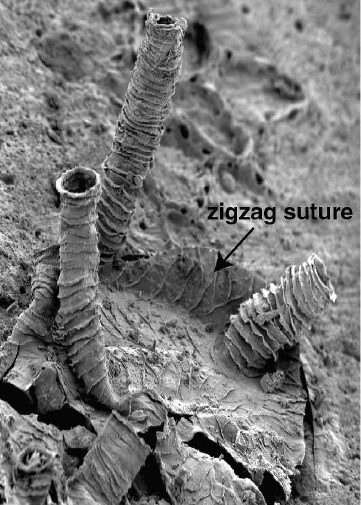
각 필석 군체는 초기 개체인 시큘라 조이드(sicular zooid)에서 시작되며, 그로부터 이후의 조이드가 발달한다. 이들은 모두 스톨론으로 연결되어 있는데, 이는 ''Rhabdopleura''(라브도플레우라)가 공유하지만 ''Cephalodiscus''(세팔로디스쿠스)는 공유하지 않는 진정한 군체 시스템이다. 이러한 조이드들은 두부 방패의 샘에서 분비된 일련의 관으로 구성된 유기 구조 내에 자리잡고 있다. 군체 구조는 공생체(coenecium, 현생 날개새류), 라브도솜(rhabdosome, 화석 필석류) 및 가장 흔하게 '''튜바륨'''(tubarium, 둘 다)을 포함하여 여러 다른 이름으로 알려져 있다. 각각 하나의 조이드가 차지하는 개별 관은 '''테카'''라고 알려져 있다. 튜바륨의 구성은 명확히 알려져 있지 않지만, 다양한 저자는 콜라겐 또는 키틴으로 만들어졌다고 제안한다. 일부 군체에는 더 큰 오토테카(autotheca)와 더 작은 비테카(bitheca)의 두 가지 크기의 테카가 있으며, 이러한 차이는 군체 내 조이드의 성적 이형성 때문일 수 있다고 제안되었다.
군체 발달 초기에 튜바륨은 가변적인 수의 가지('''스타이프'''로 알려짐)로 갈라지며 테카의 다양한 배열은 필석 화석을 식별하는 데 중요한 특징이다. 군체는 총 테카 열의 수(이열 군체는 두 열, 일열 군체는 한 열)와 군체당 초기 스타이프의 수(다지 군체는 많은 스타이프, 소지 군체는 두 개 또는 하나)로 분류할 수 있다. 각 테카 관은 대부분 '''푸젤리'''(fuselli, 단수: 푸젤룸, fusellum)라고 하는 일련의 겹쳐진 반원 반고리로 구성된다. 푸젤리는 화석으로 보존될 때 성장선과 유사하며, 두 개의 스택은 지그재그 패턴의 봉합선을 따라 만난다. 푸젤리는 튜바륨의 주요 보강 구성 요소이지만, 1개 이상의 느슨한 조직층인 피질의 도움을 받는다.
3. 2. 개체 (Zooid)
포벽 안에 살고 있는 하나의 필석 개체는 개충(zooid)이라고 불린다. 필석은 일반적으로 껍데기만이 발견되기 때문에 우리가 필석 개충에 대해 아는 것은 얼마 없다.화석화된 개충은 1978년 Bjerreskov에 의해 최초로 보고되었다. Bjerreskov는 필석 화석에 X선을 쬐어, 황철석이 포벽 주변으로 응집된 것을 발견했다. 그 학자는 이를 개충의 흔적으로 해석하였다.[23] 그 후로 프시그랍투스(''Psigraptus'')의 황철석으로 치환된 개충이 1984년에 보고되었다. 여기서는 개충과 개충 사이를 잇는 가는 실과 같은 것이 관찰되었는데, 이는 개체들을 연결하는 줄기로 해석되었다.[24] 줄기의 흔적은 이후 라트비아의 실루리아기 필석 화석에서도 보고되었다.[25]
이처럼 필석의 개충은 화석으로 남은 기록이 아예 없는 것은 아니다. 그러나 개충의 신체 구조를 일일이 파악할 수 있을 정도로 살아 생전의 모습이 온전하게 보존된 것은 없다.[26] 필석 개충의 모델이나 복원도는 대부분 간벽충을 바탕으로 한 것이 현실이다. 멸종 필석의 경우 포벽, 특히 포벽 입구의 형태가 다양하기 때문에 개충의 형태 역시 다양했으리라고 유추할 수 있다.[26]
4. 분류
필석아강에는 여러 하부 분류가 있는데, 그 중 대표적인 분류는 수지목(Dendroidea)과 필석목(Graptoloidea)이다. 수지목은 캄브리아기에서 석탄기까지, 필석목은 오르도비스기에서 데본기까지 화석이 발견된다. 필석목은 수지목 조상으로부터 오르도비스기 초기에 갈라져 나왔다.[22]
"필석"이라는 이름은 1735년 린네가 화석과 유사한 무기적 광물화 및 부착물을 지칭하기 위해 사용한 ''Graptolithus''("바위에 쓴 글자")에서 유래되었다. 이후 필석 화석은 이끼벌레와 히드로충과 같은 다른 분지형 군체 동물들을 포함한 다양한 그룹으로 언급되었다. 1970년대 이후 전자 현미경의 발달로 인해 필석류는 익새강과 가장 밀접한 관련이 있는 것으로 여겨지는데, 이는 반삭동물에 속하는 희귀한 현대 해양 동물 그룹이다.[7]
필석류는 몇몇 하위 과와 멸종된 두 개의 주요 목, 수지상필석목(저서 필석류)과 필석목(부유성 필석류)을 포함한다.
4. 1. 수지상필석목 (Dendroidea)
필석아강의 하부 분류 중 하나인 수지목(Dendroidea)은 부착성 생물로, 캄브리아기에서 석탄기까지 화석이 발견된다. 다만, ''Pseudocallograptus cf. salteri''와 같은 일부 종은 부유성 생활을 한 것으로 추정된다.[21] 수목처럼 가지가 많은 형태의 화석 대부분은 수지목에 분류되는 덴드로이드 필석이다. 이들은 캄브리아기에 등장하여 해저에 뿌리와 같은 것으로 붙어 있었다.필석 군체는 rhabdosome영어이라고 불리며, sicula영어라고 불리는 개체에서 시작하여 stipes영어(줄기)라고 불리는 다른 개체가 이어진다. 하나하나의 개체는 theca영어라고 불리는 관 또는 작은 용기 같은 형태를 하고 있다. 이 군체의 분기나 배열 상태는 필석의 종류를 구별하는 중요한 단서가 된다.
4. 2. 필석목 (Graptoloidea)
필석목(Graptoloidea)은 물 속을 떠다니는 부유성 생활을 했다. 이들은 오르도비스기에서 데본기까지 화석이 발견된다. 필석목은 수지목 조상으로부터 오르도비스기 초기에 갈라져 나왔는데, 가장 원시적인 필석목인 아니소그랍투스과(Anisograptidae)의 경우 부유성이라고 생각되나 수지목과 유사하게 생겼다. 대체로 나중에 분화되고 번성한 종류일수록 포벽의 형태가 복잡하고, 스타이프의 개수가 적다.[22] 특히 실루리아기와 데본기에 번성한 모노그랍투스과(Monograptidae)는 하나의 스타이프로만 이루어져 있다.필석은 군생하는 경우가 많으며, rhabdosome|랍도솜영어이라고 불린다. 그 군체는 sicula|시쿨라영어라고 불리는 개체에서 시작하며, 그곳에 stipes|스타이프영어(줄기)라고 불리는 다른 개체가 이어져 있다. 하나하나의 개체는 theca|테카영어라고 불리는 관 또는 작은 용기 같은 형태를 하고 있다. 이 군체의 분기나 배열 상태가 필석의 종류를 구별하는 중요한 단서가 된다.
가지가 많은 타입의 화석 대부분은 수지목(Dendroidea)에 분류되는 덴드로이드 필석이다. 덴드로이드 필석은 캄브리아기에 등장하여, 수저에 뿌리와 같은 것으로 붙어있는 타입이었다. 가지가 적은 필석목(Graptoloidea)은 오르도비스기 초기에 등장하여, 원양의 해면을 자유롭게 떠돌아다녔거나, 해초 등에 감겨 있었다. 필석목은 시대에 매우 적합하여, 데본기 초기에 멸망할 때까지 동물성 플랑크톤으로서 번성하였다.
4. 3. 하위 분류 (Maletz, 2014)
Maletz (2014)에 의한 필석류 분류는 다음과 같다.[2]
5. 생태
필석은 초기 고생대 생태계의 주요 구성 요소였으며, 특히 가장 풍부하고 다양한 종이 동물성 플랑크톤이었기 때문에 중요했다. 필석은 아마도 현탁물을 먹는 생물이었고 플랑크톤과 같은 먹이를 걸러 물을 섭취했다.[8]
필석의 존재는 오래전부터 알려졌지만, 처음에는 생물이 아닌 돌의 한 형태라고 생각되었다. 이 때문에 "그래프톨라이트"와 같이 광석과 같은 이름이 붙었다. 1735년 린네는 이를 무생물일 가능성이 높다고 하면서도, 라는 학명을 붙여 속으로 취급했다. 1768년 저서 『자연의 체계』('''') 제12권에서 , 2종을 채록하고, "식물과 같기도 하고, 필석(무생물)과 같기도 하다"라고 해설했다. 린네는 1751년 출판한 『스카네 기행』(''Skånska Resasv'')에서도 "기묘한 화석, 혹은 필석"에 대해 언급하고 있다(린네가 발견한 것은 현재 biserial graptolites영어에 속하는 필석으로 여겨진다). 이후 연구자들은 이와 유사한 생물군을 부르는 데 의 이름을 사용했다. 가 정식으로 폐지된 것은 1954년 국제 동물 명명 규약에서이며, 그 이유는 가 원래 무생물을 지칭하기 위한 용어였기 때문이었다.
1970년대 전자 현미경이 발달하면서, 필석은 익새강 에 가까운 생물이며, 반삭동물에 속하는 특수한 생물로 여겨지게 되었다. 익새강에는 현존 종도 있으며, 두개원반충속 20종, 가시원반충속 8종이 알려져 있다. 두개원반충속은 1882년에 처음 발견되었다.
5. 1. 서식 환경
필석은 초기 고생대 생태계, 특히 동물성 플랑크톤에서 중요한 구성 요소였다. 이들은 현탁물을 먹는 생물로, 플랑크톤과 같은 먹이를 걸러 물을 섭취했다.[8] 현대 익새류와 비슷하게, 먹이 효율을 높이고 포식자를 피하기 위해 수직으로 이동했을 것으로 추정된다. 연구에 따르면, 일부 종은 연안에서 공해에 이르기까지 표해수층 및 중심해수층에 주로 서식했다.[9]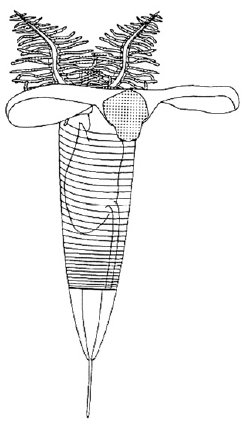
필석의 이동은 물 덩어리와 관련이 있었지만, 정확한 메커니즘(난류, 부력, 능동 수영 등)은 아직 불분명하다. Melchin과 DeMont(1995)는 필석이 "바다 나비"(나각류)처럼 머리 덮개나 먹이 촉수에서 발달한 짝을 이룬 근육 부속지를 이용해 노젓기 또는 수영을 통해 이동했다고 제안했다. 그러나 일부 종에서는 컵 구멍이 제한적이어서 이 가설이 실현 가능하지 않을 수 있다. 부력은 추가적인 컵 조직이나 가스 축적 제어 메커니즘에 의해 뒷받침되지 않으며, 능동 수영은 많은 에너지를 소모하여 튜바륨 건설에 사용될 수 있다.
필석 이동에 대한 의문은 여전히 남아있지만, 이러한 메커니즘은 종과 서식지에 따라 다를 수 있다. 퇴적물이나 다른 유기체에 부착되어 사는 저서성 종에게는 이동이 문제가 되지 않았다. 동물 개체는 튜바륨 내에서 제한적으로 이동할 수 있었고, 스톨론에 의해 움직임이 제한되었지만, 입 가까이에 있는 팔과 촉수를 사용하여 물을 걸러 음식 입자를 잡을 수 있었다.
필석은 군생하는 경우가 많으며, rhabdosome영어이라고 불린다. 군체는 sicula영어라는 개체에서 시작하며, stipes영어(줄기)라고 불리는 다른 개체가 이어진다. 각 개체는 theca영어라고 불리는 관 또는 작은 용기 형태를 하고 있다. 이 군체의 분기나 배열 상태는 필석의 종류를 구별하는 중요한 단서가 된다.
수목처럼 가지가 많은 타입의 화석은 대부분 Dendroideala목에 분류되는 덴드로이드 필석이다. 캄브리아기에 등장하여, 수저에 뿌리와 같은 것으로 붙어있는 타입이었다. 가지가 적은 Graptoloideala목은 오르도비스기 초기에 등장하여, 원양의 해면을 자유롭게 떠돌아다녔거나, 해초 등에 감겨 있었다. Graptoloideala목은 데본기 초기에 멸망할 때까지 동물성 플랑크톤으로서 번성했다. 덴드로이드 필석은 석탄기 초까지 살아남았다.
필석의 화석은 셰일이나 이암과 같은 퇴적암 내, 다른 해양 생물이 적은 지층에서 발견되는 경우가 많다. 이는 심해의 산소가 적은 곳에 침전되어, 다른 침전물이 겹쳐져서 만들어진 경우, 좋은 보존 상태로 발견되기 때문이다.
필석은 석회암이나 처트 안에서 발견되기도 하지만, 이러한 암석이 만들어지는 조건의 장소에는 다른 생물이 많이 살고, 침전된 필석의 사체도 먹혀버리기 때문에, 발견된 예는 적다.
필석의 화석은 바위 사이에 끼여 찌그러진 상태로 발견되기도 한다. 특히 황철석에 끼어 있는 경우가 많다. 이 경우, 원래의 입체 상태에서의 모습을 추정하는 것이 가능하다. 그 분기의 모습은 톱날 모양, 또는 튜닝 포크 모양인 경우가 많다. 이러한 화석은 식물로 오인되는 경우도 많다. 극히 드물게, 바위 표면에 탄소의 막으로서 발견되는 경우가 있으며, 여기에 비스듬히 빛을 비추면 빛나는 무늬로 관찰할 수 있다.
영국에서는, 아베레이디 해안(Abereiddy) , 디페드(Dyfed) 등 웨일스 지방의 오르도비스기 지층에서 자주 발견된다.

5. 2. 이동 방식
필석은 초기 고생대 생태계, 특히 동물성 플랑크톤에서 중요한 구성 요소였다. 이들은 현탁물을 먹는 생물로, 플랑크톤과 같은 먹이를 걸러 물을 섭취했다.[8]현대 익새류와의 유추를 통해, 필석은 먹이 효율을 높이고 포식자를 피하기 위해 수직으로 이동할 수 있었다고 추론된다. 상에 대한 생태 모델 및 연구에 따르면, 적어도 오르도비스기 종의 경우 일부 종 그룹은 연안에서 공해에 이르기까지 표해수층 및 중심해수층에 주로 국한되었다.[9]
필석의 이동 방식은 물 덩어리와 관련이 있었지만, 정확한 메커니즘(난류, 부력, 능동 수영 등)은 아직 명확하지 않다. Melchin과 DeMont(1995)는 필석의 이동이 무거운 주거 구조를 가진 현대의 자유 유영 동물과 유사하다고 제안했다. 특히 그들은 필석을 "바다 나비"(나각류), 즉 작은 유영 익족류 달팽이와 비교했다. 이 제안에 따르면, 필석은 머리 덮개 또는 먹이 촉수에서 발달한 짝을 이룬 근육 부속지의 물결 모양 운동을 통해 노젓기 또는 수영을 통해 이동했다.
일부 종에서는 컵 구멍이 매우 제한적이어서 부속지 가설이 실현 가능하지 않다. 부력은 어떠한 추가적인 컵 조직이나 가스 축적 제어 메커니즘에 의해서도 뒷받침되지 않으며, 능동 수영은 많은 에너지 낭비를 필요로 하는데, 이는 튜바륨 건설에 사용될 것이다.
필석 이동에 관해서는 아직 많은 의문이 있지만, 이러한 모든 메커니즘은 종과 서식지에 따라 가능한 대안이다. 퇴적물이나 다른 유기체에 부착되어 사는 저서성 종의 경우, 이것은 문제가 아니었다. 동물 개체는 이동할 수 있었지만 튜바륨 내에서 제한되었다. 비록 이러한 동물 개체의 움직임이 부유성 및 저서성 종 모두에서 가능하지만, 이는 스톨론에 의해 제한되며, 특히 먹이 섭취에 유용하다. 입 가까이에 있는 팔과 촉수를 사용하여 물을 걸러 음식 입자를 잡는다.
6. 생명 주기
필석류의 발생생물학 연구는 얕은 수역에서 발견된 종인 ''R. compacta''와 ''R. normani''의 발견으로 가능해졌으며, 필석 화석은 현존하는 대표자들과 유사한 발달을 보였을 것으로 추정된다. 생명 주기는 개체 발생과 아스토제니, 두 가지 사건을 포함하며, 주요 차이점은 발달이 개별 유기체에서 발생하는지 아니면 모듈형 콜로니 성장으로 발생하는지에 있다.
생명 주기는 유성 생식에 의해 생성된 플랑크톤성 플라눌라 유사 유충으로 시작되며, 이는 나중에 콜로니를 시작하는 ''sicular zooid''가 된다. ''Rhabdopleura''에서 콜로니는 수컷과 암컷 동물체를 가지지만 수정된 난자는 암컷 관에 보관되며, 수영할 수 있는 유충이 될 때까지 (4~7일 후) 새로운 콜로니를 시작하기 위해 정착할 때까지 그 안에 머문다. 각 유충은 동물체로의 변태가 일어나는 보호용 고치로 자신을 둘러싸고 (7~10일), 몸의 후면 부분에 부착되며, 거기에서 줄기가 결국 발달할 것이다.[10]
발달은 간접적이고 난황 영양이며, 유충은 섬모가 있고 색소가 있으며 배쪽에 깊은 우울증이 있다.[11] 아스토제니는 콜로니가 영구적인 말단 동물체의 끝에서 무성생식을 통해 성장할 때 발생하며, 그 뒤에서 새로운 동물체가 줄기에서 싹을 틔운다. 이를 단각형이라고 한다. 필석 화석에서는 새로운 동물체가 마지막 동물체의 끝에서 형성되었기 때문에 말단 동물체가 영구적이지 않았을 가능성이 있으며, 즉 합엽형 새싹이라고 할 수 있다. 이 새로운 유기체는 관 벽에 구멍을 내고 자체 튜브를 분비하기 시작한다.
7. 진화 발달 생물학에서의 중요성
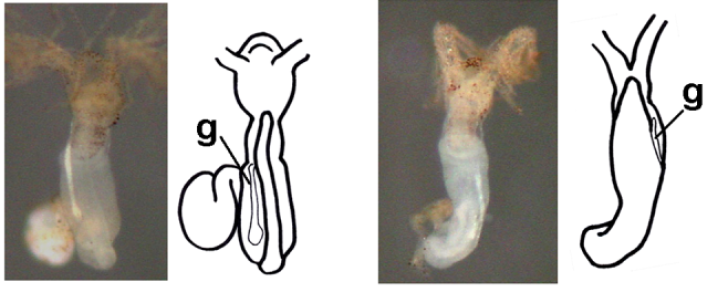
최근 몇 년 동안 살아있는 필석은 도토리벌레와 마찬가지로 진화 발달 생물학 연구의 반삭동물 모델로 사용되어 왔다. 예를 들어, 필석은 반삭동물의 비대칭성, 특히 생식선이 한쪽으로 무작위로 위치하는 경향이 있기 때문에 이를 연구하는 데 사용된다. ''Rhabdopleura normani''에서 고환은 비대칭적으로 위치하며, 구강판과 생식공과 같은 다른 구조물도 그럴 수 있다.[12] 이러한 발견의 중요성은 척삭동물이 반삭동물의 자매군이기 때문에 초기 척추동물의 좌우 비대칭성을 이해하는 것이다. 따라서 비대칭성은 후구동물에서 초기에 발달한 특징일 수 있다. 구조물의 위치가 일부 장새류에서도 엄격하게 확립되지 않았기 때문에, 반삭동물의 비대칭 상태는 강력한 발달적 또는 진화적 제약 하에 있지 않을 가능성이 높다. 최소한 생식선의 경우, 이러한 비대칭성의 기원은 튜바리움 기저부의 나선 방향, 익상새류의 일부 고유한 생물학적 메커니즘 또는 단순히 환경적 요인의 영향을 받을 수 있다.
신경 발달 패턴 형성에 관여하는 매우 보존된 유전자인 ''고슴도치''(hh)는 익상새류 대표로 ''Rhabdopleura''를 사용하여 반삭동물에서 분석되었다. 익상새류의 고슴도치 유전자는 다른 반삭동물인 장새류 ''Saccoglossus kowalevskii''와 비교하여 다른 패턴으로 발현되는 것으로 밝혀졌다. hh 유전자에서 자기 촉매 분해 부위에 있는 중요한 보존된 글리신-시스테인-페닐알라닌(GCF) 모티프는 N-말단에서 아미노산 트레오닌(T)의 삽입으로 인해 ''R. compacta''에서 변경되었으며, ''S. kowalesvskii''에서는 세린(S)이 글리신(G)으로 대체되었다. 이러한 돌연변이는 자가 단백질 분해 분열의 효율성을 감소시키므로 단백질의 신호 전달 기능을 감소시킨다. 이러한 독특한 메커니즘이 진화 과정에서 어떻게 발생했고 그룹에 어떤 영향을 미쳤는지는 분명하지 않지만, 수백만 년 동안 지속되었다면 기능적이고 유전적인 이점을 의미한다.[13]
8. 지질학적 중요성
필석은 고생대의 오르도비스기와 실루리아기 암석에서 주로 발견되는 화석으로, 전 세계적으로 분포한다. 특히 해저 화석이 드문 셰일과 이암에서 가장 흔하게 발견되는데, 이는 바닥 순환이 좋지 않고 산소가 부족하며 청소 동물이 없는 비교적 깊은 물에 퇴적된 환경에서 잘 보존되기 때문이다.
필석은 석회암과 처트에서도 발견되지만, 이러한 암석은 일반적으로 청소 동물을 포함하여 바닥에 사는 생물에게 더 유리한 환경에서 퇴적되었기 때문에 필석 유해가 다른 동물에게 먹혔을 가능성이 높다.
필석 화석은 종종 암석의 층리면을 따라 평평하게 발견되지만, 황철석 등으로 채워지면 3차원으로 발견될 수도 있다. 모양은 수지상 또는 분지형(예: ''Dictyonema''), 톱날 모양 또는 "튜닝 포크" 모양(예: ''Didymograptus murchisoni'') 등 다양하며, 화석 식물로 오인되기도 한다.
필석 화석은 일반적으로 암석 표면에 검은색 탄소 피막으로 보존되거나, 구조적으로 변형된 암석에서 밝은 회색 점토 피막으로 보존된다. 때로는 늘어나거나 왜곡되어 나타나기도 하지만, 표본을 빛에 기울여 보면 반짝이는 흔적으로 확인할 수 있다.
영국에서는 웨일스 디베드의 아베리디 베이, 스코틀랜드 남부 고원, 호수 지구, 웨일스 변경 지역 등에서 필석 화석이 많이 발견된다. 스코틀랜드의 돕스 린은 오르도비스기-실루리아기 경계에서 발견되는 종으로 유명하다. 필석은 넓은 분포를 가지기 때문에 미국, 캐나다, 호주, 독일, 중국 등 여러 지역에서도 풍부하게 발견된다.
8. 1. 표준 화석
필석은 고생대 암석의 연대를 측정하는 데 중요한 표준 화석이다. 지질학자들은 필석 화석을 이용하여 오르도비스기와 실루리아기의 암석을 백만 년 미만의 단위로 세분화할 수 있다. 오르도비스기 말에 발생한 전 세계적인 빙하기로 인해 네오그랍티네를 제외한 대부분의 필석이 사라졌다. 오르도비스기 빙하에서 살아남은 네오그랍티네로부터의 분화는 약 2백만 년 후에 시작되었다.[14]오르도비스기 대폭발(GOBE)은 군체의 형태와 시스에 영향을 미쳐 플랑크톤인 Graptoloidea와 같은 새로운 그룹을 탄생시켰다. 이후 이 그룹에 영향을 미친 가장 큰 대멸종 중 일부는 오르도비스기의 Hirnantian과 실루리아기의 Lundgreni였으며, 이때 필석 개체수가 극적으로 감소했다 (릴리풋 효과도 참조).[15]
필석의 다양성은 Aeronian의 Sedgwickii 사건 동안 크게 감소했다.[16] 이 사건은 현재의 캐나다, 리비아는 물론 아르헨티나의 라 칠카 층 (당시 곤드와나의 일부)과 같은 지역에서 증명되었다.[16]
필석 화석은 예측 가능한 보존, 광범위한 분포, 그리고 지질 시대에 걸친 점진적인 변화를 보이기 때문에 전 세계 암석 지층의 연대를 측정하는 데 사용될 수 있다.[7] 필석은 진화 속도가 빠르고 종류도 많았기 때문에 지질 시대를 파악하는 데 유용하게 사용된다.[17] 또한, 필석 화석을 통해 해당 지층이 생성된 수심과 수온을 추정할 수도 있다.
8. 2. 고환경 복원
필석 화석은 세계 각지에서 발견된다. 진화 속도가 빠르고 종류도 많았기 때문에 지질 시대를 아는 데 많이 사용된다.[17] 따라서 고생대의 표준 화석으로서 중요하며, 예를 들어 영국의 오르도비스기부터 실루리아기까지의 지층을 백만 년 단위로 식별할 수 있다. 오르도비스기 말의 빙하기에 필석의 대부분이 멸종했고, 실루리아기의 필석은 그 생존한 1, 2종에서 진화한 것이다. 또한, 필석 화석으로부터 해당 지층이 생성된 수심과 수온을 추정할 수도 있다.9. 한국의 필석
대한민국에서는 고생대 초 캄브리아기-오르도비스기에 형성된 퇴적암 지층 조선 누층군에서 필석의 화석이 발견된다.[27] 한국에서 발견된 필석 화석의 목록은 다음과 같다.
10. 연구자
참조
[1]
논문
Hemichordata (Pterobranchia, Enteropneusta) and the fossil record
[2]
논문
Phylogenetic analysis reveals that ''Rhabdopleura'' is an extant graptolite
[3]
논문
The classification of the Pterobranchia (Cephalodiscida and Graptolithina)
[4]
서적
Graptolite Paleobiology
https://www.wiley.co[...]
Wiley-Blackwell
[5]
간행물
Treatise on Invertebrate Paleontology. Part V. Graptolithina, with sections on Enteropneusta and Pterobranchia
Geological Society of America and University of Kansas Press
[6]
서적
Fundamentals of Invertebrate Palaeontology: Macrofossils
https://books.google[...]
[7]
서적
Life: A Natural History of the First Four Billion Years of Life on Earth
Alfred A. Knopf
[8]
웹사이트
Graptolites
http://samnoblemuseu[...]
2018-12-28
[9]
논문
Palaeoecology of the Graptoloidea
[10]
논문
Evolutionary crossroads in developmental biology: hemichordates
[11]
논문
Developmental Biology of Pterobranch Hemichordates: History and Perspectives
[12]
논문
Asymmetry in a Pterobranch Hemichordate and the Evolution of Left-Right Patterning
[13]
논문
Degenerate evolution of the hedgehog gene in a hemichordate lineage
[14]
논문
Graptoloid diversity and disparity became decoupled during the Ordovician mass extinction
[15]
논문
Biotic Crises in the History of Upper Silurian Graptoloids: A Palaeobiological Model
1993
[16]
논문
New insights on the Silurian graptolite biostratigraphy of the La Chilca Formation, Poblete Norte section, Central Precordillera of San Juan, Argentina: faunal replacement and paleoenvironmental implications
https://www.scielo.c[...]
[17]
서적
Life: a natural history of the first four billion years of life on earth
Alfred A. Knopf
[18]
웹사이트
真核生物の系譜
http://epp.eps.nagoy[...]
[19]
서적
Life: a natural history of the first four billion years of life on earth
Alfred A. Knopf
[20]
논문
Phylogenetic analysis reveals that Rhabdopleura is an extant graptolite
https://onlinelibrar[...]
[21]
논문
The classification of the Pterobranchia (Cephalodiscida and Graptolithina)
http://www.geology.c[...]
[22]
웹인용
Welcome to Stephen Hui Geological Museum
https://www.earthsci[...]
2021-11-12
[23]
논문
Discoveries on graptolites by X-Ray studies
http://yadda.icm.edu[...]
[24]
논문
Psigraptus, its classification, evolution and zooid
https://doi.org/10.1[...]
[25]
논문
Preservation of soft tissues in Silurian graptolites from Latvia
https://onlinelibrar[...]
[26]
논문
Graptolite reconstructions and interpretations
https://doi.org/10.1[...]
[27]
서적
한국화석도감
2003
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com