어룡
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
어룡은 중생대 바다에서 번성했던 멸종된 해양 파충류이다. 1699년 웨일스에서 발견된 화석 조각을 통해 처음 기술되었으며, 1811년 메리 애닝에 의해 최초의 완전한 어룡 화석이 발견되었다. 어룡은 뱀장어와 유사한 형태에서 시작하여 돌고래와 유사한 유선형 몸체로 진화했으며, 지느러미로 변형된 앞다리와 뒷다리, 꼬리지느러미를 가졌다.
어룡은 육식성이며, 다양한 크기와 생존 기간에 걸쳐 다양한 먹이를 섭취했다. 이들은 폐호흡을 하고 태생으로 새끼를 낳았으며, 고속 유영에 적합한 체형과 깊은 잠수 능력을 갖추었다. 어룡은 트라이아스기, 쥐라기, 백악기에 걸쳐 번성했지만, 백악기 후기에 멸종되었다.
더 읽어볼만한 페이지
- 어룡 - 운도로사우루스
운도로사우루스는 후기 쥐라기 시대에 살았던 해양 파충류로, 불완전하게 융합된 좌골치골과 강한 치열을 가지고 있으며, 삼각형 이빨로 해양 생물을 잡아먹는 육식성 포식자로 추정되지만, 계통 발생적 위치는 아직 논쟁 중이다. - 어룡 - 겐가사우루스
쥐라기에 이탈리아 겐가 지역에서 서식한 겐가사우루스는 약 2.4m의 몸길이를 가진 오프탈모사우루스과의 어룡으로, 비교적 완전한 화석 골격 특징으로 오프탈모사우루스와 구별된다. - 글로벌세계대백과를 인용한 문서/{{{분류 - 공 (악기)
공은 금속으로 제작된 타악기로, 다양한 문화권에서 의식, 신호, 음악 연주 등에 사용되며, 형태와 용도에 따라 여러 종류로 나뉜다. - 글로벌세계대백과를 인용한 문서/{{{분류 - 국무회의
국무회의는 대한민국 대통령을 의장으로, 예산, 법률안, 외교, 군사 등 국정 현안을 심의하는 중요한 기관이며, 대통령, 국무총리, 국무위원으로 구성되고, 정례회의는 매주 1회, 임시회의는 필요에 따라 소집된다. - 글로벌세계대백과를 인용한 문서/{{{분류2 - 공 (악기)
공은 금속으로 제작된 타악기로, 다양한 문화권에서 의식, 신호, 음악 연주 등에 사용되며, 형태와 용도에 따라 여러 종류로 나뉜다. - 글로벌세계대백과를 인용한 문서/{{{분류2 - 국무회의
국무회의는 대한민국 대통령을 의장으로, 예산, 법률안, 외교, 군사 등 국정 현안을 심의하는 중요한 기관이며, 대통령, 국무총리, 국무위원으로 구성되고, 정례회의는 매주 1회, 임시회의는 필요에 따라 소집된다.
| 어룡 - [생물]에 관한 문서 | |
|---|---|
| 기본 정보 | |
![[[익티오사우루스 소머세텐시스]] 골격 (NHMUK OR 2013)](https://cdn.onul.works/wiki/source/194c72d6134_b2687354.jpg) | |
![[[오프탈모사우루스]] 복원도](https://cdn.onul.works/wiki/noimage.png) | |
| 학명 | Ichthyosauria |
| 명명자 | Blainville, 1835 |
| 어원 | 고대 그리스어로 "물고기 도마뱀"을 의미 (ἰχθύς (ichthys, 물고기) + σαῦρος (sauros, 도마뱀)) |
| 생물학적 분류 | |
| 계 | 동물계 |
| 문 | 척삭동물문 |
| 강 | 파충강 |
| 아강 | 이궁아강 |
| 상목 계급 없음 | 쌍궁류 (Diapsida) |
| 목 | 어룡목 (Ichthyosauria) |
| 시간 척도 | |
| 생존 시기 | 트라이아스기 초기 – 백악기 후기 (인두절 부터 투로니아절까지), 페름기 후기에 기원했을 가능성이 있음. |
| 하위 분류군 | |
| 하위 그룹 | 텍스트 참조 |
| 기타 명칭 | |
| 후에네오사우리아 | Hueneosauria |
2. 역사
어룡은 현생 돌고래와 수렴 진화하여 유사한 생태적 지위를 차지했으며, 중생대의 대부분에 걸쳐 생존했다. 약 2억 5천만 년 전 트라이아스기 초기에 출현했으며, 이는 공룡(약 2억 3천만 년 전 출현)보다 약간 이른 시기이다. 어룡은 약 9천만 년 전 백악기 후반에 멸종했는데, 이는 공룡 멸종보다 약 2,500만 년 앞선다.
트라이아스기 초기에 육상 파충류 중 한 그룹이 바다로 돌아가 진화한 것으로 여겨지나, 정확한 조상 그룹은 아직 밝혀지지 않았다. 어룡은 이궁류에 속하는 것은 분명하지만, 현생 파충류와 플레시오사우루스 등을 포함하는 인룡형류나, 악어와 공룡 등을 포함하는 주룡형류보다는 더 원시적인 그룹에서 갈라져 나온 것으로 추정된다.
어룡은 특히 쥐라기에 크게 번성했으나 백악기 후반에 멸종하였고, 이후 바다의 최상위 포식자 자리는 플레시오사우루스나 모사사우루스류와 같은 다른 해양 파충류들이 차지하게 되었다.
어룡이 속한 분류군은 '''어룡목'''(Ichthyosauriala)이다. 1840년 영국의 생물학자 리처드 오웬은 '물고기 지느러미'라는 뜻의 Ichthyopterygiala라는 이름을 제안하기도 했으나, 현재 이 이름은 어룡목을 포함하는 더 상위 분류군인 어기류를 가리키는 데 사용된다. 어룡 화석의 발견과 연구에 대한 자세한 역사는 하위 문단에서 다룬다.
2. 1. 초기 발견
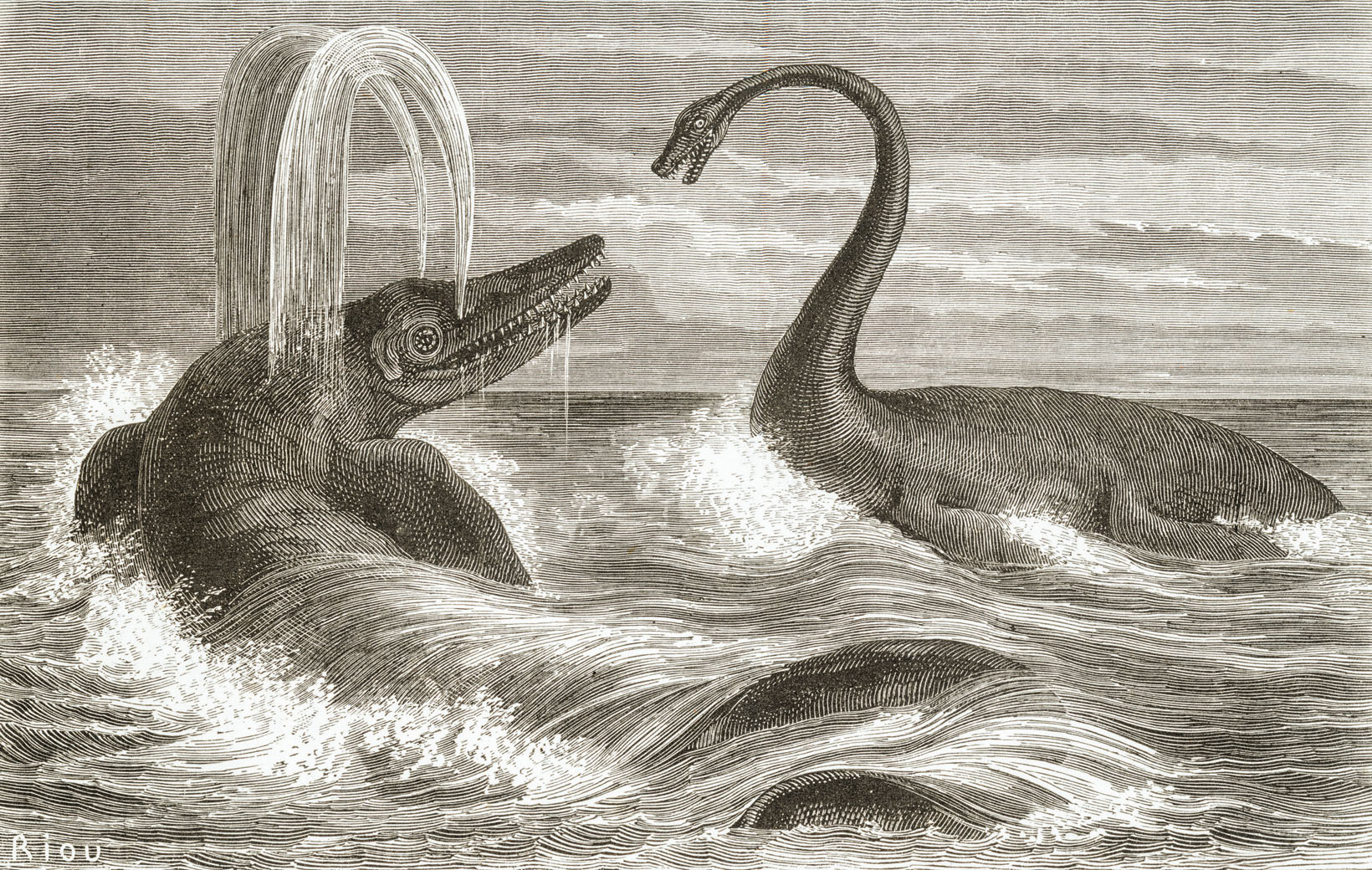
어룡에 대한 초기 연구는 단편적인 화석 조각에서 시작되었다. 1699년 웨일스 출신의 에드워드 르위드는 그의 저서 Lithophylacii Brittannici Ichnographia|리토필라키 브리타니키 이크노그라피아la에서 어룡의 뼈, 척추, 사지 골격 일부를 묘사한 삽화를 최초로 발표했다. 하지만 르위드는 이것을 물고기의 잔해로 생각했다.[5] 1708년에는 스위스 박물학자 요한 야코프 슈호이처가 두 개의 어룡 척추뼈 화석을 발견하고, 이를 대홍수 때 익사한 사람의 뼈라고 기술하며 세계적인 홍수의 명백한 증거로 여겼다.[6]
1766년에는 이빨이 있는 어룡의 턱뼈가 잉글랜드 바스 근처 웨스턴에서 발견되었고, 1783년 이 화석은 자연사 진흥 학회에 의해 악어류의 것으로 전시되었다. 1779년에는 존 월콧의 저서 ''화석의 설명과 그림''에도 어룡 뼈 삽화가 실렸다.[7] 18세기 말 영국에서는 화석 수집이 유행처럼 번졌고, 애쉬턴 레버나 존 헌터와 같은 수집가들의 소장품이 박물관으로 옮겨지면서 수십 점의 어룡 뼈와 이빨이 확인되었다. 그러나 당시에는 이 뼈들을 물고기, 돌고래, 악어의 것으로, 이빨은 바다사자의 것으로 잘못 분류하는 경우가 많았다.[8]
수집가들의 수요가 늘어나면서 상업적인 화석 발굴 활동도 활발해졌다. 19세기 초에는 더 완전한 형태의 골격들이 발견되기 시작했다. 1804년 세인트 도나츠의 에드워드 도노반은 턱, 척추, 갈비뼈, 어깨뼈 등을 포함하는 약 4m 길이의 어룡 표본을 발굴했지만, 이를 거대한 도마뱀으로 여겼다. 1805년 10월에는 제이콥 윌킨슨과 피터 호커 목사가 웨스턴에서 각각 어룡 골격을 발견했다는 신문 기사가 나왔고, 1807년 피터 호커의 사촌인 조셉 호커가 이 표본들을 묘사했다.[9] 이 표본들은 '호커의 악어'로 불리며 지질학자들 사이에서 유명해졌다. 1810년 스트랫퍼드어폰에이번 근처에서 발견된 어룡 턱뼈는 더 완전한 표본을 만들기 위해 플레시오사우루스의 뼈와 결합되기도 했는데, 이는 당시까지 어룡의 독특한 특징이 제대로 이해되지 못했음을 보여준다.
최초의 완전한 어룡 화석은 1811년 메리 애닝에 의해 영국 남부 라임 레지스(현재 쥐라기 해안으로 불리는 지역)에서 발견되었다. 이는 어룡 연구의 중요한 전환점이 되었다.
2. 2. 완전한 골격 발견과 학명
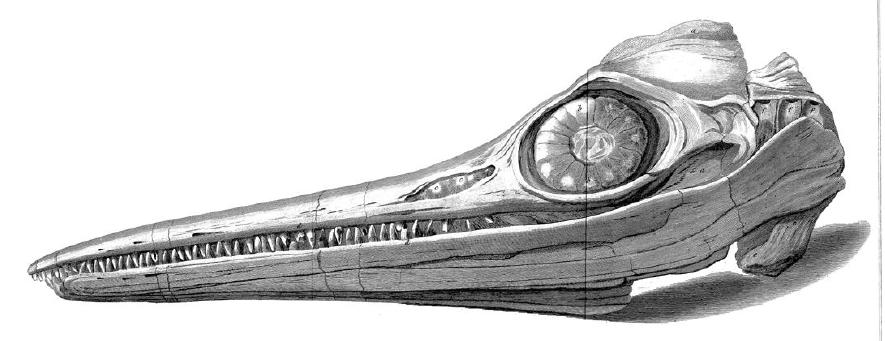
어룡 화석은 웨일스에서 나온 화석 조각을 바탕으로 1699년에 처음 기술되었다. 최초의 화석화된 척추는 1708년 두 차례 발표되었으며, 당시에는 세계적인 홍수의 명백한 증거로 여겨졌다.
최초의 완전한 어룡 골격은 1811년 영국 남부 라임 레지스에서 발견되었다. 이곳은 현재 쥐라기 해안으로 알려진 지역이다. 메리 애닝의 오빠인 조셉 애닝이 먼저 두개골을 발견했고, 1년 뒤인 1812년 어린 메리 애닝이 같은 개체의 몸통 부분을 찾아냈다. 그들의 어머니 몰리 애닝은 이 완전한 골격을 헨리 헨리 경에게 23파운드에 판매했다. 헨리 경은 이 화석을 윌리엄 불록의 런던 자연사 박물관에 빌려주었다. 이후 박물관이 문을 닫자 대영 박물관이 47파운드 5실링에 구입했으며, 현재는 독립된 런던 자연사 박물관 소장품(재고 번호 NHMUK PV R1158)으로 남아있다. 이 표본은 나중에 ''템노돈토사우루스 플라티오돈''으로 동정되었다.
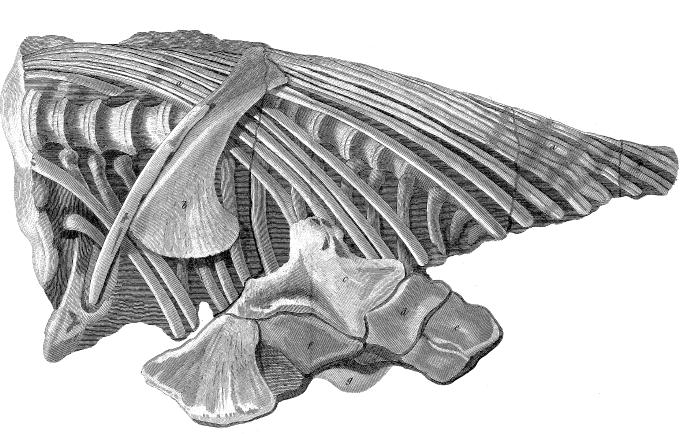
1814년, 애닝 남매가 발견한 표본은 에버라드 홈 교수에 의해 어룡에 관한 최초의 과학 논문에서 기술되었다.[10] 홈은 이 특이한 동물에 큰 관심을 보이며 기존 소장품들 중에서 추가적인 표본을 찾기 시작했다. 1816년에는 윌리엄 버클랜드와 제임스 존슨이 소유한 어룡 화석들을 기술했고,[11] 1818년에는 영국 전역의 자연학자들과 서신을 교환하며 얻은 정보를 발표했다.[12] 1819년에는 헨리 토마스 드 라 베체와 토마스 제임스 비치가 발견한 표본에 대한 두 편의 논문을 썼으며, 1820년에는 비치가 라임 레지스에서 발견한 표본에 대한 마지막 논문을 발표했다.[13] 홈의 연구는 어룡의 전반적인 해부학적 구조를 다루었지만, 세부적인 특징만을 강조했을 뿐 체계적인 설명은 부족했다.
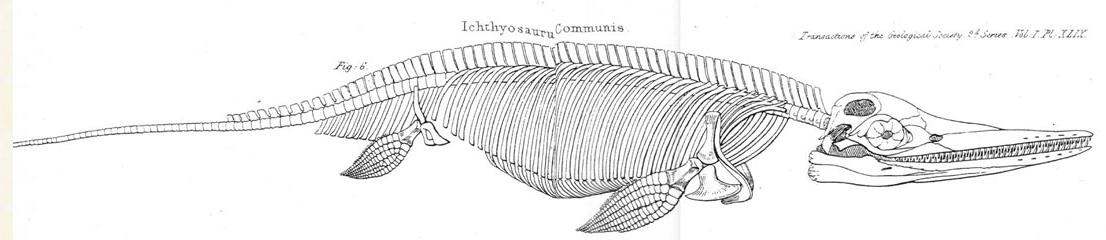
홈은 이 동물을 분류하는 데 어려움을 겪었다. 골격 요소 대부분은 파충류와 비슷했지만, 전체적인 모습은 물고기와 유사했기 때문에 처음에는 척추뼈의 납작한 형태를 근거로 물고기로 분류했다. 동시에 그는 이 생물을 물고기와 악어 사이의 과도기적 형태로 보았는데, 이는 진화적인 의미보다는 모든 생명체를 계층적으로 연결하는 "존재의 대사슬"(scala naturaela)에서의 위치를 의미했다. 1818년, 홈은 어룡의 부리뼈와 오리너구리의 흉골 사이의 피상적인 유사점을 발견하고, 이를 근거로 어룡이 오리너구리처럼 여러 주요 그룹의 특징을 결합한 과도기적 형태임을 강조하려 했다. 1819년에는 올름과 같은 영원과 도마뱀 사이의 형태로 간주하고, 마침내 ''Proteo-Saurus''라는 속명을 제안했다.[14][15]
그러나 1817년, 카를 디트리히 에버하르트 쾨니히는 이미 이 동물을 "물고기 도마뱀"이라는 뜻의 그리스어 ἰχθύς|ichthys|이크튀스grc(물고기)에서 유래한 ''Ichthyosaurus''라고 언급했다. 이 이름은 당시에는 정식으로 발표되지 않아 효력이 없는 nomen nudumla 상태였고, 쾨니히 자신은 1825년에야 이 이름을 출판했다.[16] 하지만 1819년 드 라 베체가 세 종의 ''Ichthyosaurus''를 명명하는 강연에서 이 이름을 사용했다. 이 강연 내용은 1822년에야 출판되었는데, 이때 드 라 베체의 친구인 윌리엄 코니베어가 이 세 종에 대한 설명과 함께 네 번째 종을 추가하여 공동으로 발표했다.[17] 이때 모식종으로 지정된 것은 현재는 유실된 골격을 바탕으로 한 ''Ichthyosaurus communis''였다. 코니베어는 ''Ichthyosaurus''가 ''Proteosaurus''보다 우선한다고 주장했는데, 현대적인 명명 규약 기준으로는 부정확하지만 결과적으로 ''Proteosaurus''라는 이름은 잊힌 이름(nomen oblitumla)이 되었다. 1821년, 드 라 베체와 코니베어는 어룡에 대한 최초의 체계적인 설명을 제공하면서, 새롭게 밝혀진 또 다른 해양 파충류 그룹인 플레시오사우루스와 비교했다.[18] 이 설명에는 그들의 친구이자 해부학자인 조셉 펜틀랜드의 기여가 컸다.
1835년, 앙리 마리 뒤크로테 드 블렌빌은 어룡을 포함하는 분류군으로 어룡목(Ichthyosauria)을 명명했다.[19] 1840년에는 리처드 오웬이 대안적인 개념으로 어룡상목(Ichthyopterygia)을 명명했다.[20]
이후에도 중요한 어룡 화석 발견이 이어졌다. 1905년 애니 알렉산더의 후원으로 캘리포니아 대학교의 존 C. 메리엄이 이끄는 탐사대가 네바다주 중부의 트라이아스기 얕은 바다 지층에서 25개의 표본을 발견했다. 이 표본 중 일부는 현재 캘리포니아 대학교 고생물학 박물관에 소장되어 있으며, 다른 일부는 암석에 묻힌 상태 그대로 네바다주 나이군의 베를린-어룡 주립공원에 전시되어 있다.
1977년에는 트라이아스기 어룡인 쇼니사우루스가 네바다주의 주(州) 화석으로 지정되었다. 네바다주는 약 17m에 달하는 쇼니사우루스의 완전한 골격을 소유한 유일한 곳이다. 1992년에는 캐나다의 고생물학자 엘리자베스 니콜스 박사(로열 티럴 고생물학 박물관 소속)가 역사상 가장 큰 어룡 화석 표본(길이 약 23m)을 발견하기도 했다.
2. 3. 19세기 연구와 대중화

이전에 상상조차 못 했던 거대한 해양 파충류 그룹이 멸종했다는 사실이 발견되자 과학계와 대중 모두 큰 충격을 받았고 상상력을 자극했다. 사람들은 어룡의 기이한 모습, 특히 눈구멍의 큰 공막환에 매료되었으며,[21] 때로는 살아있는 동물에게도 이런 특징이 보였을 것이라고 잘못 추측하기도 했다. 그 기괴한 형태는 사람들에게 소외감을 느끼게 했고, 어룡이 바다를 헤엄치던 시대 이후 엄청난 시간이 흘렀음을 깨닫게 해주었다.[22]
모든 사람이 어룡이 멸종했다고 생각한 것은 아니었다. 조지 영 목사는 1819년 위트비에서 골격을 발견했고, 1821년 자신의 저술에서 살아있는 표본을 여전히 발견할 수 있기를 희망했다.[23] 반대로, 지질학자 찰스 라이엘은 지구가 매우 오래되었으며 순환한다고 가정하여, 시간이 지나면 어룡이 다시 나타날 수도 있다고 생각했다. 이는 헨리 드 라 베체가 1830년에 그린 유명한 캐리커처에서 풍자되었다.

괴짜 수집가 토마스 호킨스의 활동 덕분에 대중의 인식이 높아졌다. 그는 어룡을 악마의 흉측한 창조물이라고 믿는 아담 이전설 지지자였다. 호킨스는 1834년에 ''어룡과 플레시오사우루스의 회고록''[24]과 1840년에 ''위대한 바다 용의 책''[25]을 출판했다. 첫 번째 책에는 존 사무엘슨 템플턴의 메조틴트 기법으로 제작된 삽화와 함께 과학적 설명이 포함되어 있어, 해당 주제에 대한 최초의 교과서로 여겨진다. 1834년 여름, 호킨스는 윌리엄 버클랜드와 기드온 만텔의 세금 납부 후, 당시 세계 최대 규모였던 자신의 광범위한 컬렉션을 대영 박물관에 팔았다. 그러나 큐레이터 코니히는 화석들이 루카 출신의 이탈리아 장인이 만든 석고로 심하게 복원되었다는 사실을 금방 발견했다. 가장 눈에 띄는 ''이크티오사우루스'' 표본의 경우, 꼬리 전체가 가짜였다. 버클랜드 교수가 이 사실을 미리 알고 있었다는 것이 밝혀지자, 박물관은 호킨스와 합의하여 가짜 부분에 진짜 골격 요소와 구별하기 위해 더 밝은 색상을 칠해야 했다.[26]

어룡은 1851년 세계 박람회를 위해 건립되었던 크리스탈 팰리스가 시드넘 힐에 1854년 재건되면서 더욱 대중적인 인기를 얻었다. 크리스탈 팰리스 공원의 주변 공원에는 리처드 오언의 지시에 따라 벤자민 워터하우스 호킨스가 디자인한 멸종된 동물의 실물 크기, 페인트칠된 콘크리트 조각상이 설치되었는데, 그중에는 어룡의 세 가지 모델이 포함되었다. 어룡이 먼 바다에 사는 동물이라는 것은 알려져 있었지만, 조각상들은 해안에서 햇볕을 쬐는 모습으로 묘사되었다. 이는 코니베어가 한때 설명했듯이, 그들의 모습을 더 잘 드러내기 위해 많은 19세기 삽화에서 따르던 관례였다. 이로 인해 어룡이 실제로 수륙 양생 생활을 했다는 오해가 생기기도 했다. 공원의 연못은 당시 조수 변화의 영향을 받아 물 높이가 변동했는데, 이로 인해 어룡 조각상이 간헐적으로 물에 잠기며 약간의 현실감을 더했다. 놀랍게도, 공막환이나 지느러미의 많은 마디와 같은 내부 골격 구조가 조각상 외부에 표현되기도 했다.

19세기 동안 명명된 어룡 속의 수는 점차 증가했으며, 새로운 발견을 통해 그들의 해부학적 구조에 대한 이해도 깊어졌다. 오언은 많은 화석에서 꼬리 뒤쪽이 아래로 굽어 있다는 점에 주목했다. 처음에는 이를 사후 효과, 즉 사체가 부패하면서 힘줄이 꼬리 끝을 아래로 잡아당기는 것으로 설명했다. 그러나 필립 그레이 이절턴 경의 이 주제에 대한 논문 이후,[27] 오언은 이 굽은 부분이 꼬리지느러미의 아래쪽 엽(lobe)을 지지했을 가능성을 고려했다.[28]
이 가설은 독일에서의 새로운 발견으로 확인되었다. 쥐라기 전기에 해당하는 홀츠마덴의 포시도니아 셰일에서는 이미 19세기 초부터 첫 번째 어룡 골격이 발견되었다.[29][30][31] 세기 후반 동안 발견 속도는 급증하여 매년 수백 개에 달했고, 궁극적으로 4,000개 이상이 발굴되어 어룡 표본의 대부분을 차지하게 되었다. 이 유적지는 또한 Konservat-Lagerstätte|콘서버트-라거슈테테deu였는데, 이는 발굴된 화석의 양뿐만 아니라 보존 상태 또한 매우 뛰어나다는 것을 의미했다. 골격은 매우 완전했고, 꼬리지느러미 및 등지느러미를 포함한 연조직 흔적이 종종 보존되었다. 또한, 뱃속에 배아를 가진 암컷 개체들도 발견되었다.[32]
2. 4. 20세기와 21세기 연구

20세기 초 어룡 연구는 독일의 고생물학자 프리드리히 폰 후네가 주도했다. 그는 독일에서 발견된 많은 표본을 바탕으로 방대한 연구 논문을 발표하며 어룡의 해부학적 지식을 크게 넓혔다.[33] 폰 후네는 해외 여러 지역을 방문하며 유럽 외 지역에서 발견된 화석들도 기술했다. 이 시기 북아메리카는 새로운 어룡 화석의 중요한 공급처가 되었다. 1905년 존 캠벨 메리엄이 이끌고 애니 몬태규 알렉산더가 자금을 지원한 1905년 도마뱀 원정대는 트라이아스기 당시 얕은 바다였던 네바다 중부에서 25개의 어룡 표본을 발견했다. 이 표본 중 일부는 현재 캘리포니아 대학교 고생물학 박물관에 소장되어 있으며, 다른 표본들은 암석에 묻힌 상태로 네바다주 나이 카운티의 베를린-어룡 주립공원에서 볼 수 있다.
1930년대부터 1970년대까지는 새로운 속이 명명되지 않는 등 연구가 다소 주춤했으나, 20세기 후반에 다시 활발해졌다. 1977년에는 17m 길이에 달하는 트라이아스기 어룡 ''쇼니사우루스''가 네바다주의 주 화석으로 지정되었다. 네바다주는 쇼니사우루스의 완전한 골격을 소유한 유일한 곳이다. 유효하다고 여겨지는 어룡 속의 약 절반 가량이 1990년 이후에 기술되었다. 1992년 캐나다 고생물학자 엘리자베스 니콜스는 23m 길이에 달하는 가장 큰 어룡 표본인 ''샤스타사우루스''를 발굴했다. 이러한 새로운 발견들은 어룡의 해부학과 생리에 대한 이해를 점진적으로 향상시켰다. 크리스토퍼 맥고언은 다수의 논문을 발표하며 어룡에 대한 대중의 관심을 높이는 데 기여했다.[34] 또한, 분지학이라는 새로운 연구 방법이 도입되어 동물 그룹 간의 관계를 정밀하게 계산할 수 있게 되었고, 1999년 모타니 료스케는 어룡의 계통 발생에 대한 최초의 포괄적인 연구를 발표했다.[35]
2003년, 맥고언과 모타니는 어룡과 그 근연종에 관한 현대적인 내용을 담은 첫 교과서를 출판했다.[36] 2016년[37]과 2020년[38][39]에는 각각 영국 릴스톡과 서머싯에서 거대한 어룡의 턱뼈 두 개가 발견되었다. 이 뼈들의 크기를 바탕으로 추정한 결과, 해당 어룡의 전체 길이는 최대 26m에 달할 것으로 보여, 현재까지 알려진 가장 큰 해양 파충류로 여겨진다. 이 화석은 약 2억 2백만 년 전의 것으로 추정된다.
3. 진화
어룡은 현생 돌고래와 수렴 진화하여 비슷한 생태적 지위를 차지했으며, 중생대 대부분의 기간 동안 생존했다. 약 2억 5천만 년 전 트라이아스기 초기에 육상 파충류 중 하나로부터 진화하여 수중 생활에 적응했지만, 정확한 조상은 아직 밝혀지지 않았다. 이는 공룡(약 2억 3천만 년 전 출현)보다 약간 이른 시기이다. 어룡은 이궁류에 속하지만, 인룡형류(도마뱀, 뱀, 플레시오사우루스 포함)나 주룡형류(거북, 악어, 공룡 포함)의 주요 계통보다는 더 오래된 계통에서 갈라져 나온 것으로 보인다.
가장 초기의 어룡 형태는 캐나다, 중국, 일본, 노르웨이 스피츠베르겐 섬의 초기 트라이아스기 지층에서 발견된다. 이들은 ''차오후사우루스'', ''그리피아'', ''우타츠사우루스'' 등을 포함하며, 이 시기 해양 생태계가 이미 대형 포식자를 지탱할 수 있었음을 보여준다.[157] 일부 초기 어룡은 타라트알콘처럼 최상위 포식자의 지위를 차지하기도 했다.
트라이아스기 동안 어룡은 바다뱀과 유사한 ''킴보스폰딜루스''부터 작은 ''믹소사우루스''까지 다양한 형태로 진화했다. 후기 트라이아스기에는 고전적인 ''샤스타사우루스''와 돌고래를 닮은 어룡들이 등장했으며, ''쇼니사우루스''와 같은 거대 종도 출현했다. 이 거대 종들은 노리안기 말기에 사라진 것으로 보인다.
트라이아스기-쥐라기 대멸종을 살아남은 어룡은 쥐라기 초기에 다시 번성했다. 이 시기에는 ''유리노사우루스'', ''익티오사우루스'', ''스테노프테리지우스'' 등 유선형의 돌고래와 유사한 형태가 주를 이루었다. 쥐라기 중기 이후에는 거대한 눈을 가진 오프탈모사우루스과가 번성했으며, 이들은 깊은 바다에서 먹이를 찾았을 것으로 추정된다.[158]
백악기에도 ''플라티프테리지우스''를 포함한 여러 속이 생존했지만[160], 후기 백악기 초반(약 9천만 년 전)에 모두 멸종했다[159][160]. 이는 공룡 멸종보다 약 2,500만 년 빠른 시점이다. 멸종은 두 단계로 진행된 것으로 여겨지며, 마지막 단계는 대규모 해양 무산소 사건과 관련이 있는 것으로 보인다.[161][162] 어룡이 사라진 후, 바다의 최상위 포식자 자리는 플레시오사우루스와 모사사우루스류가 차지하게 되었다.
어룡이 속한 목의 이름은 어룡목(Ichthyosauriala)이다. 1840년 리처드 오웬은 Ichthyopterygiala라는 이름을 제안했지만, 현재 이 이름은 어룡목의 상위 분류군인 어기류를 지칭하는 데 사용된다.
3. 1. 기원과 초기 어룡류


어룡의 기원은 오랫동안 논란의 대상이었다. 육상 척추동물 그룹과의 명확한 중간 형태 화석이 발견되지 않았으며, 알려진 가장 초기의 어룡 계통 종들은 이미 완전히 물속 생활에 적응한 상태였다. 2014년 중국에서 발견된 초기 트라이아스기 후기의 작은 기저 어룡형 화석은 양서류와 유사한 생활 방식을 가졌을 가능성을 시사하는 특징을 보여주었다.[40] 과거 1937년 프리드리히 폰 휘네는 어룡이 파충류가 아닌 양서류에서 독자적으로 진화한 계통이라는 가설을 제시하기도 했으나,[41] 오늘날 이 가설은 받아들여지지 않는다. 현재 학계에서는 어룡이 후기 페름기 또는 초기 트라이아스기에 육상에서 알을 낳던 양막류 사지동물로부터 유래했다는 점에 동의한다. 하지만 어룡 특유의 고도로 진화된 파생형질 때문에 정확한 조상을 파악하고 양막류 진화 계통수 내에서의 위치를 확립하는 데 어려움이 있다.[67]
어룡의 계통학적 위치에 대해서는 여러 가설이 제기되었다. 20세기 후반에는 어룡을 "원시" 파충류의 초기 분기점으로 여겨지는 무궁류에 속한다고 보기도 했다.[42] 이는 화석 기록상 어룡이 비교적 일찍 출현하고 다른 파충류 그룹과의 뚜렷한 연관성이 부족하다는 점을 설명하려는 시도였으나,[67] 무궁류 자체가 자연 분류군이 아닌 측계통군으로 밝혀지면서 이 가설은 힘을 잃었다. 현대의 분지학적 분석 결과, 어룡은 쌍궁류에 속한다는 것이 일관되게 나타난다. 일부 연구에서는 쌍궁류 계통수의 기저부에 위치시키기도 했지만,[43] 더 많은 분석은 어룡이 더 진화된 쌍궁류 하위 그룹인 신쌍궁류에 속함을 시사한다.[44]
1980년대 이후로는 어룡류와 또 다른 주요 해양 파충류 그룹인 사우롭테리지아(플레시오사우루스류 포함) 사이의 밀접한 관계가 유궁류 내에서 제기되었다. 1997년 존 메르크는 이들이 단계통군을 이루는 고궁류형 유궁류임을 주장했다.[45] 초기에는 유궁류 자체가 수생 적응 형질을 공유하는 다계통군으로 여겨져 논란이 있었으나, 최근 연구들은 어룡형류, 사우롭테리지아, 탈라토사우루스류가 하나의 큰 해양 고궁류형 단계통군을 형성하며, 이 그룹이 후기 페름기에 기원하여 초기 트라이아스기에 다양하게 분화했다는 가설을 지지하고 있다.[46][47][48]
가장 초기의 어룡 화석은 캐나다, 중국, 일본, 그리고 노르웨이의 스피츠베르겐 섬에서 발견된 초기 및 중기 트라이아스기(올레네키안과 아니시안) 지층에서 나왔으며, 연대는 최대 2억 4,600만 년 전까지 거슬러 올라간다. 이 초기 형태들에는 ''차오후사우루스''(Chaohusaurus), ''그리피아''(Grippia), ''우타추사우루스''(Utatsusaurus) 등이 포함된다. 이보다 더 오래된 화석의 존재 가능성도 제기되는데, 이는 어룡이 페름기 대멸종 이후 불과 200만 년 만인 약 2억 5천만 년 전에 이미 존재했음을 시사하며, 따라서 그 기원은 후기 페름기까지 거슬러 올라갈 수 있음을 암시한다.[50]
이 초기 어룡들은 후대의 돌고래와 유사한 형태보다는 지느러미가 달린 도마뱀에 더 가까운 모습이었다. 몸은 길쭉했고, 아마도 몸 전체를 물결치듯 움직이는 뱀장어형 유영을 했을 것으로 추정된다. 육상 동물의 특징인 튼튼한 가슴 골격과 골반을 가지고 있었고, 척추뼈에는 중력에 대항하여 몸을 지탱하기 위한 맞물림 구조가 남아 있었다. 그럼에도 불구하고 팔다리는 이미 완전히 지느러미 형태로 변형되어 수생 생활에 상당히 적응한 상태였다. 또한, 이들이 온혈동물이었고 알 대신 새끼를 낳는 태생이었을 가능성도 제기된다.
이러한 초기 어룡들은 후대의 "진짜 어룡"들과는 구조적으로 상당히 달랐기 때문에, 일부 연구자(모타니 등)는 이들을 어룡류(Ichthyosauria)에서 제외하고 더 큰 분류군인 어룡형류(Ichthyopterygia)의 기저 그룹으로 분류하기도 한다.[44] 그러나 이러한 분류 방식이 모든 연구자에게 받아들여지는 것은 아니다. 초기 어룡형류의 출현은 트라이아스기 초기의 해양 생태계가 이미 타라트알콘(Thalattoarchon)과 같은 대형 포식자를 지탱할 수 있을 만큼 복잡하게 발달했음을 보여주는 증거이기도 하다.[157]
3. 2. 후페수쿠스와의 관계

1959년 이후, 어룡 외에 또 다른 수수께끼의 고대 해양 파충류 그룹인 후페수키아가 알려졌다. 어룡류와 마찬가지로, 후페수키아는 뾰족한 주둥이를 가지고 있으며, 다섯 개 이상의 손가락이나 발가락을 갖는 다지증을 보인다. 그들의 팔다리는 육상 동물의 팔다리와 더 유사하여, 이들이 어룡류와의 진화 과정 중간 단계에 있는 과도기적 형태일 수 있다는 가능성이 제기되었다.
처음에 이러한 가능성은 후페수키아가 매우 뻣뻣한 몸통을 가지고 있어 헤엄치는 방식이 근본적으로 다르다는 이유로 거의 무시되었다. 두 그룹의 유사성은 서로 다른 환경에서 비슷하게 진화한 수렴 진화의 사례로 설명되었다. 게다가, 후페수키아의 기원 또한 불분명하여, 어룡류와의 가까운 관계가 어룡류의 일반적인 진화적 위치를 명확하게 해주는 데 큰 도움이 되지 못했다.
2014년에는 짧은 주둥이, 큰 지느러미, 뻣뻣한 몸통을 가진 작은 종인 ''카르토린쿠스''가 발표되었다. 이들은 물과 육지를 오가는 생활을 했을 가능성이 있다. 연구자 모타니는 카르토린쿠스가 어룡류보다 더 원시적인 형태라고 판단하여, 카르토린쿠스와 어룡류를 포함하는 포괄적인 분류군인 어룡형류를 새로 명명했다. 더 나아가, 후페수키아와 어룡형류를 묶어 어룡형류라는 더 큰 상위 그룹으로 분류했다. 어룡형류는 이궁류 파충류의 일부로 밝혀졌다.[49] 이 분류에 따르면 후페수키아는 어룡형류의 자매 그룹이며, 어룡형류 내에서는 카르토린쿠스가 어룡류의 자매 그룹이 된다.
3. 3. 트라이아스기 어룡의 다양성
가장 초기의 어룡 화석은 캐나다, 중국, 일본, 그리고 노르웨이의 스피츠베르겐에 있는 초기 및 중기 트라이아스기 (올레네키안기와 아니시안기) 지층에서 발견되었으며, 연대는 최대 2억 4,600만 년 전까지 거슬러 올라간다. 이러한 초기 형태에는 ''차오후사우루스'', ''그리피아'', ''우타추사우루스'' 등이 포함된다. 이보다 더 오래된 화석은 이들이 페름기 대멸종 이후 불과 200만 년 만인 약 2억 5,000만 년 전에 이미 존재했음을 시사하며, 이는 후기 페름기까지 거슬러 올라가는 더 이른 기원을 암시할 수 있다.[50] 이 초기 어룡들은 후대의 어룡들이 보여주는 물고기나 돌고래 같은 모습보다는 지느러미가 달린 도마뱀에 더 가까웠다. 몸은 길쭉했고, 아마도 몸 전체를 물결치듯 움직이는 뱀장어형 유영 방식을 사용했을 것이다. 육상 조상처럼 가슴 골격과 골반이 튼튼했고, 척추뼈에는 중력에 대항해 몸을 지탱하기 위한 맞물림 구조가 남아 있었다. 하지만 팔다리는 이미 완전히 지느러미 형태로 변형되어 있었다. 또한 이들은 온혈 동물이었고 태생이었을 가능성이 있다.이러한 초기 "원시 어룡"들은 후대의 "진정한 어룡"과 구조적으로 매우 달랐기 때문에, 일부 연구자들은 이들을 어룡목(Ichthyosauria)에서 제외하고 더 큰 분류군인 어룡형류(Ichthyopterygia)의 기저 그룹으로 분류하기도 한다.[44] 그러나 모든 연구자들이 이 분류 방식에 동의하는 것은 아니다. 초기 어룡형류의 출현은 트라이아스기 초기의 바다가 이미 대형 포식자를 지탱할 만큼 생태계가 회복되었음을 보여준다.[157] 이러한 환경 속에서 일부 어룡형류는 타라트알콘처럼 최상위 포식자의 위치를 차지하기도 했다.

기저 형태의 어룡들은 초기 트라이아스기와 중기 트라이아스기 경계 무렵에 진정한 어룡으로 빠르게 진화했다. 약 2억 4,500만 년 전에 등장한 이들은 이후 다양한 형태로 분화했는데, 여기에는 길이가 10m에 달하는 바다뱀과 유사한 형태의 킴보스폰딜루스(Cymbospondylus)와, 더 작고 전형적인 형태인 믹소사우루스(Mixosaurus) 등이 포함된다. 믹소사우루스류(Mixosauria)는 이미 뾰족한 두개골, 짧은 몸통, 더 수직적인 꼬리지느러미, 등지느러미, 그리고 많은 수의 작은 뼈로 이루어진 짧은 지느러미를 갖추어 물고기와 매우 유사한 모습을 보였다. 믹소사우루스류의 자매군은 더 진화한 메리아모사우루스류(Merriamosauria)였다.
후기 트라이아스기에 이르러 메리아모사우루스류는 크고 고전적인 형태의 샤스타사우루스류(Shastasauria)와, 더 진화하여 돌고래와 유사한 형태를 띤 유이치오사우루스류(Euichthyosauria)를 모두 포함하게 되었다. 전문가들은 샤스타사우루스류가 유이치오사우루스류로 진화하는 과정의 중간 단계(측계통군)인지,[51] 아니면 두 그룹이 더 이른 시기에 공통 조상으로부터 갈라져 나온 별개의 분류군인지에 대해 의견이 분분하다.[52] 유이치오사우루스류는 더 좁은 앞 지느러미를 가졌고 손가락(발가락) 수가 줄어드는 경향을 보였다. 기저 유이치오사우루스류에는 ''Californosaurus''와 ''Toretocnemus''가 속한다. 이보다 더 파생된 그룹인 파르비펠비아(Parvipelvia)는 골반 크기가 줄어든 특징을 보이며, 기저 형태로는 ''Hudsonelpidia''와 ''Macgowania''가 있다.

카르니아기와 노리아기 동안 샤스타사우루스류는 거대한 크기로 진화했다. 미국 네바다의 카르니아기 지층에서 여러 표본이 발견된 ''Shonisaurus popularis''는 길이가 15m에 달했다. 노리아기의 쇼니사우루스과(Shonisauridae)는 태평양 양쪽에서 발견된다. ''Himalayasaurus tibetensis''와 ''Tibetosaurus'' (아마도 동의어일 수 있음)는 티베트에서 발견되었는데, 이 거대한 어룡들(길이 10m~15m)은 일부 학자들에 의해 ''Shonisaurus'' 속에 포함되기도 한다.[53] 캐나다 브리티시컬럼비아의 파도넷 지층(Pardonet Formation)에서 발견된 거대한 ''Shonisaurus sikanniensis'' (2011년부터 2013년까지는 ''Shastasaurus'' 속으로 분류됨)는 길이가 21m에 달했을 것으로 추정된다.[68] 영국 서머싯에서 발견된 익티오타이탄(Ichthyotitan)은 길이가 26m에 달했을 것으로 추정되며, 만약 이 추정이 사실이라면 지금까지 알려진 가장 큰 해양 파충류가 된다.[37] 동시대의 다른 거대 어룡으로는 샤스타사우루스와 베사노사우루스(Besanosaurus) 등 샤스타사우루스류가 알려져 있다.[159]

후기 트라이아스기에 어룡은 크기와 다양성 면에서 정점을 이루었다. 이들은 다양한 생태적 지위를 차지했는데, 일부는 최상위 포식자였고 다른 일부는 작은 먹이를 사냥했다. 여러 종은 먹이를 빨아들이는 방식(흡입 섭취)이나 돌진하여 먹는 방식(ram feeding)에 특화되었을 가능성이 있으며, 단단한 먹이를 부수는 데 적합한 이빨을 가진 형태(경골어류 포식)도 알려져 있다. 그러나 후기 트라이아스기 말로 가면서 어룡의 다양성은 감소한 것으로 보인다. 거대 종들은 노리아기 말에 사라진 것으로 추정된다. 레티아기(최후기 트라이아스기)의 어룡 화석은 영국에서 발견되는데, 이들은 초기 쥐라기의 어룡과 매우 유사한 형태를 보인다. 이러한 다양성 감소의 원인으로는 상어, 경골어류, 그리고 최초의 플레시오사우루스와의 경쟁 심화가 거론된다. 공룡과 마찬가지로 어룡과 그들의 동시대 경쟁자였던 플레시오사우루스는 트라이아스기-쥐라기 멸종 사건에서 살아남았고, 이후 초기 쥐라기의 비어있는 생태적 지위를 채우며 빠르게 다시 다양화했다.
3. 4. 쥐라기와 백악기 어룡

초기 쥐라기 동안 어룡은 여전히 다양한 종을 유지했으며, 몸길이는 1m에서 10m에 달했다. 이 시기 영국과 독일에서는 잘 보존된 표본들이 많이 발견되었는데, 대표적인 속으로는 ''Eurhinosaurus'', ''Ichthyosaurus'', ''Leptonectes'', ''Stenopterygius'', 그리고 대형 포식자인 ''Temnodontosaurus''가 있다. ''Suevoleviathan''과 같이 좀 더 원시적인 parvipelvians도 존재했다. 하지만 전체적인 형태학적 다양성은 트라이아스기에 비해 크게 줄어들었다.[54] 거대한 몸집을 가진 종류나, 흡입 방식으로 먹이를 먹거나 단단한 먹이를 부숴 먹던 종류는 사라졌다.[55] 이 시기 어룡 대부분은 유선형의 돌고래와 비슷한 thunniform 몸체를 가졌지만, ''Leptonectes''와 ''Eurhinosaurus''를 포함하는 Eurhinosauria와 같은 일부 그룹은 더 길쭉한 몸과 긴 주둥이를 가지고 있었다.
중기 쥐라기의 어룡 화석은 상대적으로 드물게 발견된다. 이는 해당 시기의 전반적인 화석 기록이 부족하기 때문일 수 있다. 후기 쥐라기에는 Ophthalmosauridae과가 출현하며 번성했다. 몸길이 약 4m 정도인 ''Ophthalmosaurus''와 그 근연종들이 대표적인데, 이들은 전반적인 모습이 ''Ichthyosaurus''와 매우 유사했다. 특히 ''Ophthalmosaurus''는 매우 큰 눈을 가지고 있어, 아마도 빛이 적은 깊은 바다에서 사냥했을 것으로 추정된다.[56][158] 전통적으로 후기 쥐라기에는 어룡의 다양성이 감소한 것으로 여겨졌으나, 백악기의 새로운 발견들은 이 시기의 다양성이 기존에 생각했던 것보다 훨씬 높았을 가능성을 시사한다.

전통적으로 어룡은 백악기에 접어들면서 다양성이 더욱 줄어든 것으로 여겨졌다. 이 시기의 모든 화석은 한때 단일 속인 ''Platypterygius''에 속하는 것으로 생각되기도 했다.[160] 이 마지막 어룡 속은 백악기 후기 초반인 세노마니아기, 약 9,500만 년 전에 멸종한 것으로 보이며[159][160], 이는 중생대의 다른 대형 파충류 그룹들이 백악기 말까지 생존했던 것과는 대조적이다. 과거에는 이러한 멸종의 원인으로 플레시오사우루스와 같은 다른 해양 포식자와의 경쟁이나, 어룡 스스로가 특정 환경에 지나치게 특수화되어 당시 빠르게 진화하던 경골어류를 따라잡지 못했기 때문이라는 설명이 제시되었다.[57] 이 관점에서는 어룡이 진화적으로 정체되었으며, ''Platypterygius''가 보여준 유일한 혁신은 열 개의 손가락뿐이라고 보았다.[58]
하지만 최근 연구들은 백악기 어룡이 이전에 생각했던 것보다 훨씬 다양했음을 보여준다. 과거 "Platypterygius"로 분류되었던 많은 화석들이 실제로는 여러 다른 속들에 속한다는 것이 밝혀졌다. 2012년 기준으로, ''Acamptonectes'', ''Sveltonectes'', ''Caypullisaurus'', ''Maiaspondylus''를 포함하여 최소 8개의 어룡 계통이 쥐라기와 백악기의 경계를 넘어 생존했던 것으로 알려졌다.[59] 2013년에는 백악기 초기의 원시적인 툰노사우루스류인 ''Malawania''가 발견되기도 했다.[60] 실제로 대륙이 분열되면서 해안선이 늘어난 초기 백악기 동안 어룡의 새로운 방산진화가 일어났을 가능성도 제기된다.[61]
어룡의 멸종은 단번에 일어난 것이 아니라 두 단계에 걸쳐 진행된 것으로 보인다.[62][162] 첫 번째 멸종 사건은 세노마니아기 초기에 발생하여, 당시 존재했던 어룡의 세 가지 주요 먹이 방식 중 '부드러운 먹이 전문 포식자'와 '일반 포식자' 그룹이 사라지고, 특화되지 않은 최상위 포식자 그룹만 남게 되었다.[62][162] 두 번째 멸종 사건은 세노마니아-투로니아 경계 사건이라 불리는 대규모 해양 무산소 사건 동안 발생했다.[161] 이 사건으로 해양 생태계가 큰 타격을 입으면서 마지막 남은 최상위 포식자 그룹마저 멸종했고[162], 가장 마지막까지 살아남았던 계통인 ''Platypterygius hercynicus''는 약 9,300만 년 전에 자취를 감추었다.[63]
따라서 어룡의 멸종은 오랜 기간에 걸친 점진적인 쇠퇴가 아니라, 세노마니아기와 투로니아기에 발생한 급격한 환경 변화와 기후 변동에 따른 두 차례의 사건 때문이었을 가능성이 높다.[62][64] 해수면 상승과 해양 무산소 상태의 확산[161] 등 당시의 생태적 변화와 불안정성이 어룡의 이동 경로, 먹이 확보, 번식지에 영향을 미쳐 멸종을 초래한 것으로 추정된다. 이 시기에는 어룡 외에도 일부 미세 플랑크톤, 암모나이트, 벨렘나이트, 암초를 만들던 이매패류 등 다른 많은 해양 생물들도 함께 멸종했다.[62] 초기 모사사우루스류와의 경쟁이 멸종의 주된 요인이었을 가능성은 낮다. 왜냐하면 어룡의 생태적 지위를 대체할 만한 대형 모사사우루스류는 어룡이 멸종하고 약 300만 년이 지난 후에야 등장했기 때문이다.[62] 또한 플레시오사우루스류 폴리코틸리드 역시 어룡이 차지했던 일부 생태적 지위를 채웠을 수 있지만, 이들은 어룡과 약 1,900만 년 동안 공존했다.[62]
4. 해부학적 특징
어룡은 물고기나 돌고래와 유사한 유선형 몸체를 가진 해양 파충류이다. 평균 몸길이는 2m에서 4m 정도였지만, 종에 따라 크기는 매우 다양했다. 머리는 상괭이와 비슷했으며, 일반적으로 길고 이빨이 있는 주둥이를 가졌으나 파라오프탈모사우루스처럼 이빨이 없는 종도 있었다. 앞다리와 뒷다리는 노와 같은 지느러미로 변형되었고, 등에는 살로 된 등지느러미가, 꼬리에는 상어와 유사한 초승달 모양의 수직 꼬리지느러미가 발달한 경우가 많았다.[54]
어룡은 폐로 호흡했기 때문에 주기적으로 수면 위로 올라와야 했으며, 새끼를 낳는 태생 방식으로 번식했다. 이는 몸 안에서 알이 부화하여 새끼가 태어나는 방식으로, 태아를 품고 있거나 출산 도중 사망한 화석을 통해 확인되었다. 참치처럼 빠른 유영에 적합한 몸 형태를 가졌으며, 최적 효율 유영 속도는 초당 1.5m (시속 5~6km 전후)로 추정된다.[156] 몸길이 2.4m의 스테노프테리지우스는 약 163kg에서 168kg, 4m의 오프탈모사우루스는 약 930kg에서 950kg 정도의 체중이었을 것으로 추정된다.
어룡의 외형적 특징은 육상 파충류였던 조상으로부터 진화하여 수중 생활에 적응한 결과로, 물고기나 돌고래와의 유사성은 수렴 진화의 대표적인 예로 꼽힌다.[71] 특히 등지느러미와 꼬리지느러미는 뼈 구조 없이 연조직으로 이루어져 있어 초기 복원에서는 생략되기도 했으나, 이후 보존 상태가 좋은 화석들을 통해 그 존재가 확인되었다.
눈은 매우 크고 공막륜으로 보호되어 있어 빛이 적은 환경에서도 먹이를 찾는데 유리했을 것으로 보인다. 주요 먹이는 벨렘나이트와 같은 두족류였으나, 일부 종은 물고기, 갑각류, 또는 작은 파충류를 먹기도 했다.
4. 1. 전반적인 형태


어룡은 전반적으로 현생 물고기나 돌고래와 유사한 방추형 몸체를 가졌다. 특히 후기 어룡들은 이러한 경향이 두드러졌으나, 가장 초기의 어룡 계통 구성원은 뱀장어와 비슷한 길쭉한 모습을 보였다.[71] 어룡은 짧은 목과 일반적으로 길고 이빨이 있는 주둥이를 가진 머리를 가졌다. 다만 파라오프탈모사우루스처럼 이빨이 없는 어룡도 존재했다. 앞다리와 뒷다리는 노와 같은 지느러미로 완전히 변형되었다. 많은 종이 등쪽에 살로 이루어진 등지느러미를 가졌으며, 꼬리 끝에는 상어와 유사한 초승달 모양의 수직 꼬리지느러미가 있었다.[54] 꼬리지느러미는 위아래로 넓게 퍼져 있으며, 추진력을 제어하는 데 사용되었다.
어룡의 평균 길이는 약 2m에서 4m였지만, 크기는 매우 다양했다.[68] 어떤 개체는 길이가 0.3m에 불과했고, 일부 종은 훨씬 거대했다. 트라이아스기의 ''Shonisaurus popularis''는 길이가 약 15m에 달했으며,[68] ''Ichthyotitan severnensis''와 같은 종은 20m에서 25m까지 자랐을 것으로 추정된다.[37][38] 모타니 료스케의 추정에 따르면, 몸길이 2.4m 크기의 ''Stenopterygius''는 약 163kg에서 168kg의 무게였고, 4m 크기의 ''Ophthalmosaurus icenicus''는 약 930kg에서 950kg의 무게였다.[70]
어룡은 파충류였지만 물고기 및 돌고래와의 형태적 유사성은 수렴 진화의 대표적인 예로 꼽힌다. 진화 생물학자 스티븐 제이 굴드는 육상 파충류였던 조상에게는 없던 등지느러미와 꼬리지느러미가 물속 생활에 적응하며 물고기와 거의 동일한 위치와 형태로 발달한 점을 지적하며 이를 강조했다.[71] 이러한 구조의 유사성은 공통 조상으로부터 물려받은 상동성이 아니라, 동일한 환경 압력에 대한 적응의 결과인 유사성이다.[71]
초기 쥐라기 동안 어룡은 여전히 다양한 종이 존재했으며, 길이는 1m에서 10m에 달했다. 이 시기에는 ''Ichthyosaurus'', ''Stenopterygius''와 같이 유선형의 돌고래 같은 몸체를 가진 thunniform 형태가 많았지만, ''Leptonectes''나 ''Eurhinosaurus''처럼 몸과 주둥이가 더 긴 형태도 있었다.[54] 중기 쥐라기 이후, 특히 후기 쥐라기와 백악기에는 Ophthalmosauridae에 속하는 어룡들이 주를 이루었다. 이들은 ''Ophthalmosaurus''로 대표되며, ''Ichthyosaurus''와 유사한 전반적인 구조를 가졌으나, 특히 매우 큰 눈을 가진 것이 특징이다. 이 큰 눈은 공막륜으로 보호받았으며, 빛이 적은 심해나 야간에 사냥하는 데 유리했을 것으로 추정된다.[56]
어룡은 폐로 호흡했기 때문에 주기적으로 수면 위로 올라와야 했다. 또한 태생으로 번식했는데, 이는 몸 안에서 알이 부화하여 새끼가 태어나는 방식이다. 실제로 태아를 품고 있거나 출산 도중 사망한 것으로 보이는 화석이 발견되면서 태생임이 확인되었다. 폐호흡을 하는 해양 파충류가 알을 낳으려면 바다거북처럼 육지로 올라와야 하므로, 완전 수중 생활을 했던 어룡에게 태생은 필수적인 적응이었다.
어룡은 참치처럼 고속 유영에 적합한 몸 형태를 가졌으며, 현생 고래처럼 깊은 바다에도 잠수했을 것으로 추정된다. 모타니 료스케는 어룡의 최적 효율 유영 속도를 초당 1.5m (시속 5~6km 전후)로 추정했는데, 이는 현생 참치류와 거의 비슷한 수준이다.[156]
어룡의 등지느러미와 꼬리지느러미는 연골이나 살로 이루어져 있어 화석으로 잘 남지 않기 때문에, 이크티오사우루스의 초기 복원에서는 등지느러미가 없는 모습으로 그려지기도 했다. 그러나 1890년대 독일 홀츠마덴의 라거슈테테에서 발견된 보존 상태가 매우 좋은 표본들을 통해 등지느러미와 꼬리지느러미의 흔적이 명확히 밝혀졌다.
4. 2. 두개골과 턱

기저 어룡류는 이미 길고 삼각형 모양의 두개골을 가지고 있었으며, 협의의 어룡류로 진화하면서 주둥이는 매우 뾰족해졌다. 주둥이는 주로 전상악골로 이루어져 있고, 그 뒤의 상악골은 보통 더 짧다. 때로는 상악골이 전상악골의 뒤쪽 가지에 의해 외부 콧구멍과 분리되기도 한다. 이 때문에 전상악골에는 이빨이 많지만, 상악골에는 이빨이 적거나 아예 없을 수도 있다. 주둥이 뒤쪽 윗부분은 비골이 형성한다. 더 진화한 종들은 비골 뒤쪽을 나누는 정중선의 구멍인 내비공(foramen internasalela)을 가진다. 비골은 보통 뼈로 된 콧구멍의 위쪽과 앞쪽 가장자리를 형성하며, 콧구멍은 종종 눈구멍 바로 앞에 위치한다. 그러나 일부 트라이아스기 종에서는 전상악골이 뒤로 길게 뻗어 비골을 콧구멍에서 제외시키기도 한다.[72]
두개골 뒤쪽은 거대한 눈구멍이 차지하며, 때로는 뒤쪽 측면의 대부분을 덮는다. 눈구멍 안에는 물의 압력으로부터 눈을 보호하는 작고 겹치는 뼈 조각들로 이루어진 원형 구조인 큰 공막륜이 있다. 어룡류는 상대적으로나 절대적으로나 알려진 모든 척추동물 중에서 가장 큰 눈구멍을 가졌다. 다른 뒤쪽 두개골 요소들은 대개 압축되고 융합되어 구별하기 어렵다. 두개골 뒤쪽 윗부분의 요소는 전통적으로 두정골로 여겨졌고, 인두골과 사각관골은 때때로 융합된 것으로 보였다. 그러나 1968년 알프레드 셔우드 로머는 이 뼈가 실제로는 인두골이라고 주장했고,[73] 1973년 맥고완이 이를 지지했다.[74] 하지만 1990년 존 스티브 마사르는 대부분의 연구자들에게 원래의 식별, 즉 두정골이라는 해석이 옳다고 설득했다.[75] 두정골은 두정골 개구부의 뒤쪽 가장자리를 형성한다. 측면의 하부 측두 개구부는 없다. 두정골 개구부의 앞쪽 가장자리는 보통 후전두골이 형성한다. 매우 원시적인 ''우타추사우루스''에서만 후안와골과 인두골이 여전히 개구부 가장자리에 닿는다. 한 쌍의 두정골 개구부 사이에서 두개골 지붕은 좁으며, 일부 종은 턱 근육 부착을 위한 세로 방향의 볏을 가진다. 기초 어룡류는 한 쌍의 두정골 사이에 두정안 개구부를 가지고 있다. 고유 어룡류에서는 이 개구부가 앞쪽으로 이동하여 처음에는 두정골과 전두골 사이 경계에 위치하다가, 더 진화된 종에서는 전두골 사이로 완전히 이동한다. 후두정골과 탁상골은 없다. 두개골 뒤쪽과 구개골의 뼈는 종종 불완전하게 골화되어 부분적으로 연골 상태로 남아 있었던 것으로 보인다. 후두과는 일반적으로 매우 볼록하다. 고막에서 중이로 소리를 전달하는 뼈인 등골은 길쭉하고 구멍이 없다. 익상돌기 이빨은 일반적으로 없다.[72]
아랫턱 역시 주둥이처럼 길쭉하다. 하지만 유리노사우루스나 엑스칼리보사우루스 같은 일부 종에서는 주둥이 앞부분이 아랫턱보다 훨씬 앞으로 튀어나와 있다. 아랫턱 앞부분은 보통 낮지만, 뒤쪽의 깊이는 매우 다양하다. 아랫턱의 대부분은 이빨을 가진 뼈인 앞쪽의 치골로 이루어져 있다. 안쪽 면에서는 치골이 분리골로 덮여 있으며, 분리골은 두 아랫턱이 만나는 지점인 결합부까지 앞으로 뻗어 있다.[72] 턱 관절은 수평적인 씹기 운동을 허용하지 않고, 턱을 수직으로 열고 닫는 단순한 경첩 역할을 한다.[76]
4. 3. 치아
어룡의 치아는 일반적으로 원뿔 모양이었다.[72] 식성에 따라 다양한 형태를 보였는데, 물고기를 먹는 종은 길고 가느다란 치관을 가지며 약간 뒤로 굽어 있었다. 더 큰 먹이를 잡는 데 특화된 형태는 더 짧고 넓으며 곧은 치아를 가지고 있었고, 때로는 절삭 날이 존재하기도 했다. 최상위 포식자인 탈라토아르콘은 납작한 칼날 모양의 더 큰 치아를 가졌다. 조개류를 먹는 경골류 종은 낮고 볼록한 치아를 빽빽하게 가지고 있었으며, 이들의 치아 법랑질에는 깊은 수직 홈과 주름이 있었다.[72]| 식성/종류 | 치아 특징 |
|---|---|
| 물고기 섭취 종 | 길고 가는 치관, 뒤로 약간 굽음 |
| 큰 먹이 섭취 종 | 짧고 넓고 곧은 치아, 때로는 절삭 날 존재 |
| 최상위 포식자 (탈라토아르콘) | 납작한 칼날 모양의 큰 치아 |
| 경골류 (조개류 섭취) | 낮고 볼록한 치아, 빽빽하게 배열, 치아 법랑질에 깊은 수직 홈과 주름 |
많은 어룡의 치열은 여러 치아 모양을 결합한 이형치를 가졌는데, 예를 들어 앞쪽에는 작은 치아, 뒤쪽에는 더 큰 치아가 있는 식이었다. 치아는 보통 치아 소켓에 위치했지만, 파생된 종에서는 일반적인 치아 홈을 가졌으며 이는 홈니상치아로 알려진 상태이다. 홈니상치아의 경우, 성체가 되면 때때로 이가 없어지기도 했다. 치아 소켓에 있는 치아는 때때로 턱뼈와 융합되기도 했다. 어룡 치아의 상아질에서는 눈에 띄는 수직 주름이 관찰된다.[72]
4. 4. 골격과 연조직
가장 초기의 어룡은 뱀장어와 비슷한 모습이었으나, 후기로 갈수록 전형적인 물고기나 돌고래와 유사한 형태로 진화했다. 짧은 목과 긴 주둥이를 가진 머리는 돌고래와 비슷했으며, 앞다리와 뒷다리는 완전히 지느러미 형태로 변형되었다. 일부 종은 등지느러미와 수직에 가까운 꼬리지느러미를 가졌다. 외형상 물고기와 매우 흡사하지만, 어룡은 파충류이지 물고기는 아니다.
진화 생물학자 스티븐 제이 굴드는 어룡을 수렴 진화의 대표적인 예로 꼽았다. 이는 어룡과 물고기의 유사한 신체 구조가 공통 조상으로부터 물려받은 상동적인 특징이 아니라, 비슷한 수중 환경에 적응하면서 독립적으로 발달한 유사적인 특징이기 때문이다. 굴드는 다음과 같이 설명했다:
육상 조상을 가진 이 바다 파충류는 물고기와 매우 강력하게 수렴하여, 실제로 등지느러미와 꼬리를 정확한 위치와 정확한 유압 설계를 갖도록 진화시켰다. 이러한 구조는 조상이 되는 육상 파충류가 등이나 꼬리에 전조 역할을 하는 혹이나 날이 없었기 때문에 더욱 놀랍다.[71]
모타니 료스케는 1999년, 보다 진화한 형태의 어룡류(Ichthyosauria)를 가까운 초기 어룡 그룹과 구별되는 몇 가지 해부학적 특징을 기준으로 정의했다. 그 특징들은 다음과 같다.
| 부위 | 특징 |
|---|---|
| 콧구멍 | 두개골 측면에 위치하며 위에서는 거의 보이지 않음. |
| 안와 상단 | 전두골과 후두정골이 이루는 뼈 막대로 구성됨. |
| 후안와골 | 측면에서 볼 때 상측두창(supratemporal fenestra) 형성에 참여하지 않음. |
| 정수리 눈 구멍 | 두정골과 전두골의 경계에 위치함. |
| 익상골 측면 날개 | 불완전하거나 가변적으로 골화됨. |
| 척골 | 원래의 축 뒤쪽 부분이 없음. |
| 뒤쪽 등쪽 척추뼈 | 원반 모양임. |

초기 어룡 복원도에는 등지느러미와 꼬리지느러미의 넓은 부분이 그려지지 않았다. 이는 이 부분들이 단단한 뼈로 지지되지 않아 화석으로 잘 보존되지 않았기 때문이다. 꼬리 아랫부분만이 척추뼈로 지지된다. 1880년대 들어 어룡의 몸 전체 윤곽이 발견되기 시작했고, 1881년 리처드 오언은 잉글랜드 배로우어폰소어의 초기 쥐라기 암석에서 꼬리지느러미 윤곽이 보존된 어룡 화석을 보고했다.[78] 이후 잘 보존된 다른 표본들을 통해, ''차오후사우루스''와 같은 일부 원시적인 어룡들은 꼬리지느러미가 약하게 발달하여 등쪽 부분만 존재했고, 이로 인해 꼬리가 노와 같은 모양이었음이 밝혀졌다.[79] 시간이 지나면서 해당 표본의 꼬리 부분 윤곽은 희미해졌다.[82]
등지느러미의 존재 여부는 오랫동안 논란거리였다. 19세기 후반 독일 홀츠마덴의 라거슈테테(보존 상태가 매우 좋은 화석 산지)에서 발견된 표본들은 몸 전체의 윤곽, 특히 등지느러미의 존재를 시사하는 검은색 흔적을 보여주었다. 이러한 윤곽은 특수한 환경 조건 덕분에 보존되었는데, 실제 조직 자체보다는 미생물 매트의 흔적일 가능성이 제기되었다.[80] 1987년, 데이비드 마틸은 박테리아 활동에 의한 간접적인 보존 방식 때문에 이 윤곽들이 실제 형태를 정확하게 반영한다고 보기 어렵다고 주장했다. 그는 발견된 흔적들이 실제 등지느러미가 아니라 몸에서 떨어져 나온 피부 조각이거나, 심지어 화석 준비 과정에서 등지느러미가 있을 것이라는 기대 하에 인위적으로 만들어졌을 가능성도 제기하며 진짜 등지느러미는 발견되지 않았다고 결론지었다. 그는 어룡이 돌고래와 달리 뒷지느러미를 유지한 이유가 등지느러미가 없어 안정성을 확보하기 위해서였을 수 있다고 설명했다.[81] 그러나 다른 연구자들은 윤곽이 다듬어졌을 가능성은 인정하면서도, 보존된 등지느러미 중 상당수는 실제 몸 윤곽을 어느 정도 반영하는 진품일 가능성이 높다고 보았다. 웁살라 대학교 고생물학 박물관의 R158 표본처럼, 박테리아 매트 특유의 희미한 가장자리를 보여 인위적인 수정이 가해지지 않았음에도 참치와 유사한 몸 윤곽과 등지느러미를 뚜렷하게 보존하고 있는 사례도 있다.[82] 1993년 마틸 역시 일부 등지느러미 표본은 진짜일 수 있음을 인정했다.[80]
등지느러미가 보존된 화석들은 지느러미가 뼈 구조가 시사하는 것보다 훨씬 넓고 뾰족한 형태였으며, 단단한 섬유 조직으로 지지되었음을 보여준다. 일부 표본에서는 4개의 콜라겐 층이 관찰되었고, 각 층의 섬유는 서로 교차하는 구조를 이루었다.[83]
2017년에는 독일 포시도니아 셰일에서 발견된 약 1억 8270만 년 전의 ''스테노프테리지우스'' 척추 화석(탄산염 결절 내 보존)에서 콜라겐 섬유, 콜레스테롤, 혈소판, 적혈구 및 백혈구와 같은 연조직의 분자 수준 증거가 발견되었다. 이는 화석화되지 않은 원래의 유기 조직으로, 결절 내부의 보호 환경 덕분에 이례적으로 잘 보존된 것으로 보인다. 발견된 적혈구는 현대 포유류의 1/4에서 1/5 크기였는데, 이는 당시(토아르시움절)의 낮은 산소 농도 환경에서 산소 흡수 효율을 높이기 위한 적응이었을 수 있다. 콜레스테롤의 높은 탄소-13 동위원소 비율은 어룡이 식품 체인에서 상위에 위치하며 주로 물고기와 두족류를 먹었음을 시사한다.[84] 2018년에는 ''스테노프테리지우스''에서 블러버(지방층)의 증거도 발견되었다.[132]
일반적으로 어룡의 피부는 화석에 남은 윤곽으로 미루어 볼 때, 비늘 없이 매끄럽고 탄력적이었던 것으로 보인다.[85] 그러나 이러한 잔해는 실제 피부 자체가 아니라 세균 성장에 의해 형성된 윤곽일 가능성이 높다. 예외적으로 솔호펜 석회암에서 발견된 ''에기로사우루스'' 표본에서는 미세한 부분까지 보존될 수 있는 암석의 특성 덕분에 실제 피부의 흔적이 보고되었는데, 여기서는 미세한 비늘이 보이는 듯했다.[86]
어룡의 색깔을 정확히 알기는 어렵다. 1956년 메리 화이트아는 영국 화석 R 509의 피부 표본에서 적갈색 색소 과립을 함유한 색소 세포(멜라닌 세포)를 발견했다고 보고했다.[87] 전통적으로 어룡은 상어, 펭귄 등 현대 해양 동물처럼 사냥 시 위장을 위해 위쪽은 어둡고 아래쪽은 밝은 대비음영을 가졌을 것으로 추정되었다.[76] 그러나 2014년 룬드 대학교의 요한 린드그렌은 어룡 표본 YORYM 1993.338의 피부에서 검은색 멜라닌을 함유한 구조인 멜라노솜을 발견하고, 어룡이 체온 조절이나 심해 위장을 위해 균일하게 어두운 색을 띠었을 가능성이 높다고 결론지었다. 이는 대비음영을 가진 것으로 밝혀진 모사사우루스과나 선사 시대 장수거북과는 대조적이다.[88][89] 하지만 2015년 다른 연구에서는 피부의 멜라노솜 기저층이 파충류에게 흔하지만 반드시 어두운 외모를 의미하지는 않으며, 다른 색소포(이리도포어, 잔토포어, 에리트로포어 등)가 색깔에 영향을 미치지만 화석으로 잘 보존되지 않는다는 점을 지적하며 린드그렌의 해석에 의문을 제기했다. 따라서 YORYM 1993.338이 대비음영, 녹색 등 다른 색이나 무늬를 가졌을 가능성도 배제할 수 없다.[90] 2018년 린드그렌 연구팀은 ''스테노프테리지우스'' 표본에서 발견된 유멜라닌 함유 멜라노포어의 분포 변화를 근거로 어룡이 대비음영을 가졌을 것이라고 다시 주장했다.[132]
어룡의 평균 몸길이는 2m에서 4m 정도였지만, 이보다 훨씬 크거나 작은 종들도 존재했다. 머리는 상괭이와 비슷하게 생겼고, 길고 이빨이 있는 주둥이를 가졌으나, 파라오프탈모사우루스처럼 이빨이 없는 종도 있었다. 꼬리지느러미는 크고 위아래로 넓게 퍼져 있었으며, 추진력을 제어하는 데 사용되었다.
어룡은 폐로 호흡했기 때문에 주기적으로 수면 위로 올라와야 했다. 태생이었는데, 이는 태아를 품고 있거나 출산 도중 죽은 화석이 발견되면서 확인되었으며, 화석 발견 이전부터 예상되었던 사실이다. 폐호흡 해양 생물이 알을 낳으려면 바다거북이나 바다뱀처럼 육지로 올라가야 하기 때문이다. 몸 형태는 참치처럼 빠른 유영에 적합했으며, 현생 고래처럼 깊은 바다까지 잠수했을 것으로 추정된다(모타니 료스케, 2000). 어룡의 최적 효율 유영 속도는 초당 1.5m(시속 5~6km 정도)로 추정되는데, 이는 현생 참치류와 비슷한 수준이다.[156]
체중은 몸길이 2.4m인 스테노프테리지우스가 약 163kg에서 168kg, 몸길이 4m인 ''오프탈모사우루스''가 약 930kg에서 950kg 정도였을 것으로 추정된다(모타니 료스케).
먹이의 경우, 많은 어룡이 벨렘나이트라는 두족류에 크게 의존했다. 일부 초기 어룡은 갑각류의 껍데기를 부수는 데 적합한 이빨을 가지고 있었으며, 물고기도 주식으로 삼았을 가능성이 높다. 소수의 대형 종들은 튼튼한 턱과 이빨을 이용해 자신보다 작은 파충류를 잡아먹었을 것으로 보인다. 어룡은 크기가 매우 다양하고 오랜 기간 존속했기 때문에 먹이 종류도 폭넓었을 것으로 생각된다. 전형적인 어룡은 눈이 매우 크고 공막륜으로 보호되어 있었는데, 이 때문에 빛이 적은 밤에도 먹이를 찾을 수 있었을 것으로 여겨진다.
5. 생태
어룡은 외형상 물고기와 유사할 뿐만 아니라, 상어 중 람나상어류, 참치, 돌고래와 유사한 발달 특징을 공유하는데, 이는 수렴 진화의 좋은 예로 꼽힌다.[92] 고생물학자 스티븐 제이 굴드는 이러한 구조적 유사성이 상동성이 아닌 유사 관계임을 지적하며 어룡을 수렴 진화의 대표적 사례로 들었다. 이러한 유사성은 어룡이 이들과 비슷한 활동 수준(체온 조절 포함)과 생태적 지위를 가졌을 가능성을 시사한다.[92] 어룡은 중생대의 바다에 널리 서식했던 해양 파충류이다.[92]
현대의 고래류처럼 어룡도 공기를 호흡했으며, 온혈동물이었을 가능성이 제기된다. 뼈의 조직학적 연구 결과나 이빨의 산소 동위원소 비율 분석 등은 이들이 높은 체온을 유지했음을 시사한다.[126][127][128][129][130][131][132] 높은 신진대사는 빠른 유영 속도를 가능하게 했을 것이다.[133]
어룡의 몸길이는 평균 2m에서 4m 정도였으나 크기는 다양했다. 대부분 길고 이빨이 있는 주둥이를 가졌지만, 파라오프탈모사우루스처럼 이빨이 없는 종도 있었다. 매우 큰 눈은 공막륜이라는 뼈 고리로 보호되었고, 꼬리지느러미는 유영에 중요한 역할을 했다. 초기 복원에서는 등지느러미가 누락되기도 했으나, 1890년대 독일 홀츠마덴에서 발견된 보존 상태가 매우 좋은 화석들을 통해 등지느러미의 존재가 확인되었다.
어룡은 태생으로 번식했으며, 이는 육지에 오르기 어려운 신체 구조 때문으로 추정된다. 먹이는 주로 벨렘나이트와 같은 두족류나 물고기였으나, 종에 따라 다양했다. 참치와 유사한 유선형 몸체는 빠른 유영에 적합했다.
5. 1. 서식지와 먹이


어룡은 주로 연안보다는 넓은 바다에 서식했으며, 중생대의 모든 바다에서 발견되었다. 이는 가장 초기의 어룡류에게도 해당되어 특정 지역을 그들의 기원으로 단정하기 어렵다.[92]
어룡은 육식성이었으며, 크기가 매우 다양하고 오랫동안 생존했기 때문에 다양한 먹이를 섭취했을 가능성이 높다. 뾰족한 주둥이를 가진 종은 작은 동물을 잡는 데 적합했다. 일부 학자들은 위턱이 돌출된 유린호사우리아(Eurhinosauria)가 황새치처럼 주둥이를 이용해 먹이를 찔렀을 것이라고 추측하기도 한다. 어룡의 위 내용물 화석에서 가장 흔하게 발견되는 것은 두족류(특히 벨렘나이트)의 잔해이다.[93][94] 그 외에도 물고기[95], 다른 척추동물, 심지어 더 작은 어룡을 먹기도 했다.[96][97] 거대한 트라이아스기 어룡인 ''Thalattoarchon''은 크고 날카로운 이빨을 가져 자신과 비슷한 크기의 먹이를 사냥하는 대형 포식자였을 것으로 보이며,[98] ''Himalayasaurus''와 여러 ''Temnodontosaurus'' 종 역시 매우 큰 먹이를 사냥하는 데 적응했다.[99] 이러한 식성은 물고기와 두족류의 잔해가 포함된 분뇨 화석(코프로라이트)을 통해 확인된다. 1835년 버클랜드(Buckland)는 비늘로 식별 가능한, 부분적으로 소화된 물고기 덩어리가 든 표본을 보고했으며,[100] 1968년 연구에서는 이 물고기가 ''Pholidophorus'' 속이며, 두족류의 부리와 빨판 갈고리도 함께 발견되었다고 밝혔다. 이러한 단단한 먹이 입자는 위장에 보관되었다가 주기적으로 토해냈을 것으로 추정된다.[101] 때로는 익사한 동물의 사체를 먹기도 했다. 2003년 보고된 ''Platypterygius longmani'' 표본의 위장에서는 물고기와 거북이 외에 육상 조류의 뼈도 발견되었다.[102]
일부 초기 어룡은 경식성으로, 조개를 부수는 데 적합한 평평하고 볼록한 이빨을 가지고 얕은 바다의 저서생물을 먹었다. 다른 종들은 비교적 짧은 턱을 빠르게 열어 먹이를 입으로 빨아들이는 흡입 섭식을 했을 가능성이 제기되었다. 이는 ''Shonisaurus''나 짧은 주둥이를 가진 ''Guanlingsaurus liangae'' 같은 종에게서 추정되었다.[103] 그러나 2013년 연구에서는 어룡의 설골(혀의 기저부 뼈)이 흡입 섭식을 지원할 만큼 충분히 발달하지 않았다고 지적하며, 대신 입을 크게 벌리고 헤엄치며 먹이를 모으는 돌진 섭식을 했을 가능성을 제시했다.[104]
전형적인 어룡은 매우 큰 눈을 가졌으며, 공막륜이라는 뼈 고리로 보호받았다. 이는 어룡이 밤이나 깊은 바다에서 사냥했을 가능성을 시사한다. 현존하는 동물 중 이 정도로 큰 눈을 가진 것은 대왕오징어와 콜로살오징어뿐이다.[105] 따라서 시각은 사냥에 중요한 감각이었을 것으로 보인다. 청력은 등자뼈가 매우 튼튼한 구조인 점으로 미루어 좋지 않았을 수 있다. 그러나 입천장의 홈은 후각이 발달했거나 전기 수용 기관이 존재했을 가능성을 암시한다.[106]
어룡 역시 다른 동물들의 먹이가 되었다. 트라이아스기에는 주로 상어와 다른 어룡이 천적이었으며,[107] 쥐라기에는 대형 사경룡류와 탈라토수키아가 추가되었다. 이는 포식자의 위 내용물 화석을 통해 확인되는데, 예를 들어 2009년에는 사경룡류 표본의 위장에서 어룡 배아가 발견되기도 했다.[108]
위석(위 속의 돌)이 어룡 골격과 함께 발견된 사례가 몇 차례 있는데, 이는 소화를 돕거나 부력을 조절하는 데 사용되었을 수 있다. ''난노프테리지우스''와 ''판지앙고사우루스'' 화석에서 위석이 발견되었다.[91]
5. 2. 유영과 잠수
어룡의 주요 추진력은 몸통의 좌우 운동에서 나왔다. 초기 형태는 뱀장어형 운동 방식으로 몸통 전체와 꼬리를 물결치듯 움직였다.[79] 이는 일반적으로 비효율적인 방식으로 여겨진다. 이후 등장한 Parvipelvia와 같은 후기 형태는 몸통과 꼬리가 짧아지고, 몸의 마지막 3분의 1 또는 꼬리 끝부분만 움직이는 더 효율적인 카랑기폼 또는 튜니폼 운동 방식을 사용했을 가능성이 높다. 이들 종의 몸통은 상당히 뻣뻣했다.꼬리는 두 개의 엽으로 나뉘었으며, 아래쪽 엽은 꼬리뼈에 의해 지지되었다. 꼬리뼈는 아래쪽 엽의 윤곽을 따라 배 쪽으로 휘어져 있었다. 초기 어룡은 다소 비대칭적인, 즉 아래쪽 엽이 위쪽 엽보다 더 큰 '이형'(heterocercal) 꼬리지느러미를 가졌다. 이는 상어와 달리 아래쪽 엽이 가장 큰 형태였다. 더 진화한 형태는 거의 수직에 가까운 대칭적인 꼬리지느러미를 가졌다. 상어는 물보다 무거운 몸(비중성 부력)을 보상하기 위해 비대칭 꼬리지느러미를 사용하여 꼬리가 아래로 누르는 힘으로 몸 전체를 위로 향하게 한다. 이렇게 앞으로 헤엄치면 무게로 인한 가라앉는 힘을 상쇄할 만큼의 양력이 발생한다. 1973년 McGowan은 어룡의 꼬리지느러미 비대칭성이 상어와 반대이므로, 어룡은 물보다 가벼운 양성 부력을 가졌을 것이며, 이는 위석과 골다공증 또는 밀도 높은 뼈가 없는 것으로 확인될 것이라고 주장했다. 그는 꼬리가 몸을 아래로 향하게 하고, 앞지느러미가 몸 앞쪽을 더 아래로 누르며 피치(pitch)를 제어하는 데 사용되었을 것이라고 보았다.[109] 하지만 1987년 Michael A. Taylor는 다른 가설을 제시했다. 어룡은 (부레가 없는) 상어와 달리 폐 속 공기 양을 조절하여 부력을 조절할 수 있었다는 것이다. 따라서 꼬리는 주로 중립적인 추진력을 제공하고, 작은 부력 변화는 지느러미 각도를 미세하게 조절하여 안정화시켰을 것이라고 보았다.[110] 1992년 McGowan은 이 견해를 받아들여, 상어의 꼬리는 좁은 엽을 가지고 있어 더 수직적이고 대칭적인 후기 어룡의 꼬리와 좋은 비교 대상이 아니라고 지적했다. 후기 어룡의 꼬리지느러미는 참치와 더 유사하며, 이는 높은 순항 속도를 유지하는 비슷한 능력을 시사한다.[111] 2002년 모타니 료스케의 비교 연구는 현존 동물에서 작은 꼬리지느러미 엽이 높은 박동 빈도와 양의 상관관계를 보인다고 결론지었다.[112] 현대 연구자들은 일반적으로 어룡이 음성 부력, 즉 물보다 무거웠다는 데 동의한다.[113]
1994년 Judy Massare는 어룡이 가장 빠른 해양 파충류였다고 결론지었다. 길이 대비 깊이 비율이 3~5 사이로, 물 저항을 최소화하는 최적의 수치였다. 매끄러운 피부와 유선형 몸체는 난류 발생을 억제했다. 유체역학적 효율(에너지가 전방 운동으로 전환되는 정도)은 돌고래에 근접하여 약 0.8로 추정되었다. Massare는 어룡이 플레시오사우루스보다 5분의 1 더 빨랐으며, 이 차이의 절반은 어룡의 신진대사가 30% 더 높다는 가정으로 설명될 수 있다고 보았다. 그의 모델에 따르면 순항 속도는 시속 5km 약간 안 되었다.[114] 하지만 2002년 모타니 료스케는 Massare의 계산 오류를 수정하여 예상 순항 속도를 시속 2km 미만으로 낮췄다. 이는 현대 고래류보다는 약간 느린 속도이다.[115] 그러나 플레시오사우루스와 모사사우루스류의 추정 속도도 함께 하향 조정되었기 때문에, 어룡은 여전히 상대적으로 빠른 해양 파충류의 위치를 유지했다. 최적 효율 속도는 초당 1.5m(시속 5~6km 전후)로 추정되며, 이는 현생 참치류와 거의 동등하다.[156]

어룡은 길이가 다양한 지느러미 모양의 사지를 가졌다. 일반적인 해석은 이 지느러미들이 등지느러미, 꼬리지느러미와 함께 방향 안정성을 위한 조종면으로 사용되어 요(yaw)를 제어하고 피치(pitch)와 롤(roll)을 안정시키는 역할을 했으며, 추진에는 사용되지 않았다는 것이다. 그러나 1980년대 독일 고생물학자 Jürgen Riess는 다른 모델을 제시했다.[116] 그는 플레시오사우루스의 앞다리 비행 운동을 연구한 후, 긴 지느러미를 가진 어룡들이 적어도 앞지느러미를 이용해 강력한 추진 스트로크를 했으며 위아래로 움직였다고 주장했다. 이는 퇴화하지 않은 어깨뼈 구조와, 기능적으로 사용되지 않으면 불필요했을 완벽한 날개형 단면을 가진 손뼈의 진화를 설명할 수 있다고 보았다. 그는 퀸즐랜드 폐어와 아마존강 돌고래에서 유사점을 찾았다고 생각했으며, 이들도 긴 지느러미를 추진에 사용한다고 추정했다. Riess는 여러 논문을 통해 이 가설을 상세히 설명했다.[117][118][119] 이 대체 해석은 다른 연구자들에게 일반적으로 받아들여지지 않았다. 1998년 Darren Naish는 폐어와 강 돌고래가 실제로는 지느러미를 그런 식으로 사용하지 않으며, 예를 들어 현대의 혹등고래는 뼈 모자이크로 지지되는 매우 긴 앞지느러미를 가졌지만 주로 방향타 역할을 한다고 지적했다.[120] 2013년 연구는 Platypterygius와 같은 넓은 어룡 지느러미가 추진이 아닌 조종면으로 사용되었음을 시사했다.[121]

많은 현존하는 폐호흡 해양 척추동물은 깊이 잠수할 수 있다. 어룡의 잠수 능력에 대한 몇 가지 증거가 있다. 깊은 곳에서 빠르게 상승하면 감압병을 유발할 수 있는데, 이로 인한 뼈 괴사는 쥐라기와 백악기 어룡 화석에서 잘 나타난다(각각 표본의 15%, 18%). 반면 트라이아스기 종에서는 드물게 나타난다. 이는 초기 어룡들이 깊이 잠수하지 않았다는 신호일 수도 있지만, 후기 시대에 포식 압력이 커져 더 자주 빠르게 수면으로 도피해야 했기 때문일 수도 있다.[122] 그러나 후자의 가능성은 현대 동물에서 감압병 손상이 제한된 수의 급격한 상승 사건이 아니라, 정상적인 잠수 행동 중 점진적인 퇴화 축적으로 발생한다는 사실에 의해 반박된다.[123] 어룡의 눈 크기 또한 잠수 능력의 증거를 제공한다. 어룡의 눈은 척추동물 중에서 상대적으로나 절대적으로 가장 크다. 현대 표범해표는 시야를 확보하며 최대 1km까지 잠수할 수 있다. 모타니 료스케는 어룡이 상대적으로 훨씬 큰 안와(눈구멍)를 가졌으므로 훨씬 더 깊이 잠수할 수 있었을 것이라고 제안했다.[124] 눈 지름이 25cm에 달하는 ''Temnodontosaurus''는 아마도 1600m 깊이에서도 볼 수 있었을 것이다.[125] 이러한 깊이에서 큰 눈은 거대한 물체를 보는 데 특히 유용했을 것이다.[105] ''Ophthalmosaurus''과 같은 후기 종들은 상대적으로 더 큰 눈을 가졌는데, 이는 쥐라기 후기와 백악기 어룡들의 잠수 능력이 더 뛰어났음을 시사한다.
현대의 고래류와 마찬가지로 어룡은 공기를 호흡했다. 고래와 돌고래는 포유류이며 온혈동물이다. 어룡은 전통적으로 파충류로 분류되어 냉혈동물로 여겨졌으나, 1970년대 이후 중생대의 주요 파충류인 수각류 공룡, 익룡, 플레시오사우루스 등이 온혈동물이었을 것이라는 주장이 제기되었다. 이는 이들의 생태적 지배력을 설명하는 데 도움이 된다. 어룡 역시 내온성(온혈성)을 가졌을 수 있다는 직접적인 증거가 있다. 1990년 비비안 드 부프레닐은 어룡의 뼈 조직학 연구를 통해, 빠른 성장과 강한 혈관 형성이 특징인 섬유판상 골 조직을 가지고 있음을 밝혔다. 이는 온혈동물의 특징과 같다.[126] 초기 트라이아스기 어룡들도 이미 이러한 특징을 보였다.[127][128] 2012년에는 매우 원시적인 형태인 Utatsusaurus조차 이러한 뼈 유형을 가지고 있어, 어룡의 조상이 이미 온혈동물이었음을 시사한다는 보고가 있었다.[129] 높은 신진대사에 대한 추가적인 직접 증거는 이빨의 산소 동위원소 비율 분석에서 나왔는데, 이는 주변 해수 온도보다 약 20°C 높은 35°C에서 39°C 사이의 체온을 나타낸다.[130][131] 단열 효과가 있는 블러버(지방층)의 존재 역시 동물이 스스로 열을 생산해야 하는 온혈성을 뒷받침한다.[132] 내온성에 대한 간접적인 증거는 후기 어룡의 몸 형태에서도 찾을 수 있다. 짧은 꼬리와 수직 꼬리지느러미는 높은 신진대사로만 유지될 수 있는 높은 순항 속도에 최적화된 것으로 보인다. 이런 방식으로 헤엄치는 현존 동물들은 모두 완전히 온혈동물이거나, 상어나 참치처럼 몸 중심부의 높은 온도를 유지한다.[133] 이 주장은 뱀장어처럼 몸을 움직여 헤엄쳤던 초기 형태에는 적용되지 않는다.
1996년 리처드 코웬은 어룡이 온혈동물이었음을 받아들이면서도, 1987년 데이비드 캐리어가 지적한 파충류 호흡의 제약, 즉 '캐리어의 제약'을 받았을 것이라고 추정했다. 몸을 물결치듯 움직이는 방식이 폐에서 공기를 밀어내어 이동 중 호흡을 방해한다는 것이다.[134] 코웬은 어룡이 돌고래처럼 물 위로 뛰어오르면서 이 문제를 극복했을 것이라고 가설을 세웠다. 즉, 계속 물 밖으로 뛰어오르며 점프할 때마다 신선한 공기를 들이마셨을 것이라는 추측이다.[135] 다른 연구자들은 적어도 후기 어룡의 경우, 뻣뻣한 몸통 구조 때문에 캐리어의 제약이 적용되지 않았을 것으로 본다. 이는 효과적인 호흡 및 산소 저장 시스템을 시사하는 뛰어난 잠수 능력으로도 뒷받침된다. 따라서 이들 종에게 돌고래처럼 뛰어오르는 유영 방식은 필수가 아니었을 것이다. 그럼에도 불구하고 어룡은 돌고래에 비해 콧구멍 위치가 낮아, 공기를 들이쉬기 위해 머리를 약간 기울인 채 자주 수면으로 올라왔을 것으로 추정된다.[136]
5. 3. 번식과 성장

어룡은 알을 낳는 난생 조상에서 유래한 파충류임에도 불구하고, 새끼를 낳는 태생(Viviparity) 방식으로 번식했다. 이는 어룡이 공기 호흡을 위해 물 밖으로 나와야 했지만, 유선형 몸체 때문에 거북이나 일부 바다뱀처럼 알을 낳기 위해 육지로 이동하기 어려웠기 때문으로 추정된다. 대신 고래나 돌고래처럼 물속에서 새끼를 낳았을 것이다. 어룡이 태생이라는 사실은 1845년 12월 9일, 박물학자 조셉 채닝 피어스가 ''Ichthyosaurus communis'' 화석에서 2.5m 길이의 어미 산도 안에 머리가 뒤쪽을 향한 채 있는 11cm 길이의 작은 배아를 발견하면서 처음 확인되었다.[137]

이후 독일 홀츠마덴 지역에서는 태아를 품은 성체 어룡 화석이 다수 발견되었다. 1880년, 해리 고비어 시리가 이끈 영국 고생물학 위원회는 어룡이 물속에서 출산했으며, 어미 몸 안에 태아가 있는 화석은 출산 중 새끼가 먼저 죽어 어미까지 사망한 경우일 수 있다고 보았다.[138] 이는 새끼가 익사하는 것을 막기 위해 꼬리부터 태어나는 고래나 돌고래와 비교되기도 했다. 만약 새끼가 머리부터 태어나면 질식할 위험이 있고, 죽은 새끼가 산도에 걸리면 어미도 위험해지기 때문이다.[139] 하지만 다른 설명으로는, 임신한 암컷이 다른 원인으로 죽은 뒤 사체가 분해되면서 발생한 가스가 태아를 머리부터 밀어냈을 가능성도 제기되었다.

2014년에는 새끼 세 마리를 낳다가 죽은 초기 어룡 ''Chaohusaurus'' 암컷 화석이 보고되었다. 이 화석에서는 이미 태어난 두 마리와 아직 산도에 있는 한 마리가 발견되었는데, 이는 초기 어룡이 후기 어룡과 달리 머리부터 새끼를 낳았음을 시사한다. ''Chaohusaurus''는 매우 원시적인 어룡으로, 이 발견은 어룡의 육상 조상 단계에서 이미 태생이 시작되었을 가능성을 보여준다.[140][141]
2023년 발표된 연구는 임신한 어룡 표본들을 종합적으로 분석하여 태아의 출산 방향 진화를 조사했다. 연구 결과, 원시적인 어룡인 ''Chaohusaurus''와 ''Cymbospondylus''는 머리부터 출산한 증거를 보였고, ''Mixosaurus''는 머리부터와 꼬리부터 출산한 증거가 모두 나타났다. 반면, ''Stenopterygius'', ''Besanosaurus'', ''Qianichthyosaurus'', ''Platypterygius'' 등 더 진화한 메리아모사우리아 어룡들은 꼬리부터 출산한 증거를 보였다. 연구자들은 후기 어룡이 꼬리부터 출산하는 것을 선호하게 된 이유로, 출산 시 머리보다 골반을 먼저 밀어내는 것이 어미에게 더 쉬웠거나, 어미의 자세 제어(트림 컨트롤)에 드는 에너지를 줄일 수 있었기 때문일 수 있다고 주장했다. 또한 ''Mixosaurus''에서 두 가지 방향이 모두 나타난 점을 들어, 단순히 질식 위험 때문에 꼬리부터 출산하게 되었다는 기존 가설에는 동의하지 않았다.[142]
어룡의 태아는 태반 포유류나 플레시오사우루스에 비해 매우 작았고, 한 배에 낳는 새끼의 수는 많은 편이었다. ''Stenopterygius''의 경우 한 암컷 화석에서 7마리, 다른 암컷에서는 11마리의 태아가 확인되기도 했다. 태아의 크기는 어미 길이의 최대 4분의 1 정도였다.[143] 새끼는 태어날 때부터 성체와 거의 비슷한 신체 비율을 가졌으며, 성장 과정(개체 발생)에서는 주로 뼈들이 서로 합쳐지고 더 단단해지는 변화가 일어났다.[144] 최소한 한 마리의 신생아 ''I. communis'' 표본의 위 내용물 분석 결과, 어린 개체도 두족류와 물고기를 먹었던 것으로 나타났다.[145] 이는 ''Stenopterygius''처럼 성장하면서 작은 물고기에서 큰 두족류로 먹이가 바뀌는 종들과는 다른 모습이다.[146]
악어, 대부분의 바다거북, 일부 도마뱀 등은 알이 부화하는 환경 온도에 따라 새끼의 성별이 결정되지만(온도 의존성 성 결정), 새끼를 낳는 파충류는 이러한 방식을 사용하지 않는다. 2009년의 한 연구는 현생 파충류, 조류, 포유류 94종을 분석하여, 태생을 하는 동물에게는 유전적으로 성별이 결정되는 방식(성 염색체 등)이 매우 중요하다고 결론지었다. 특히 해양 파충류의 경우, 태생이 진화하기 이전에 이미 안정적인 해양 환경에 적응하면서 유전적 성 결정 방식을 갖게 되었을 것으로 보았다.[147] 따라서 어룡을 포함한 모사사우루스, 플레시오사우르 등 중생대 해양 파충류들도 유전자에 의해 성별이 결정되었을 가능성이 높다.[148]
5. 4. 사회적 행동과 지능
어룡은 종종 무리 또는 사냥 집단으로 생활했을 것으로 추정된다.[72] 하지만 어룡의 구체적인 사회적 행동 특성에 대한 증거는 아직 부족하다. 일부 연구에서는 성적 이형성이 존재했을 가능성을 제시한다. 예를 들어, ''Eurhinosaurusla''와 ''Shastasaurusla''의 골격에서는 두 가지 형태가 나타나는데, 더 긴 주둥이, 더 큰 눈, 더 긴 몸통, 더 짧은 꼬리, 그리고 추가적인 발가락 뼈를 가져 더 긴 지느러미를 가진 개체는 암컷일 수 있다. 특히 더 긴 몸통은 배아를 위한 공간을 제공했을 가능성이 있다.[149]일반적으로 어룡의 뇌는 현대 냉혈 파충류의 뇌와 비슷하게 크기가 작고 길쭉한 모양을 하고 있었다. 그러나 1973년, 맥고완(McGowan)은 잘 보존된 표본의 자연 뇌 주형을 연구한 결과, 종뇌가 예상했던 것만큼 작지는 않다는 점을 지적했다. 큰 눈의 크기로 미루어 볼 때 시각엽은 예상대로 컸으며, 후각엽과 소뇌 역시 특별히 크지는 않았지만 잘 분화되어 있었다.[74]
6. 병리학
화석을 통해 어룡의 구체적인 행동 양상을 파악하기는 어렵지만, 일부 화석에 남은 흔적을 통해 병리학적 정보를 얻을 수 있다. 예를 들어, 한 어룡 화석에서는 주둥이 부위에 다른 개체에게 물린 자국이 발견되었다. 2011년 벤자민 키어(Benjamin Kear) 등이 호주에서 발견하여 분석한 이 상처는 측정 결과, 다른 어룡, 아마도 같은 종의 어룡에게 공격당한 흔적으로 밝혀졌다. 이는 동종 간의 싸움 중 얼굴을 물어뜯는 행위였을 가능성을 시사한다. 상처 부위에서는 뼈가 다시 자라난 흔적, 즉 치유의 징후가 보여 공격받은 개체가 살아남았음을 알 수 있다.[150][151]
또 다른 사례로는 스발바르 제도에서 발견된 길이 약 9m에 달하는 매우 큰 어룡 화석이 있다. 이 화석은 꼬리 부분을 제외하고는 거의 완전한 상태로 보존되었다. 정밀 조사 결과, 이 어룡은 암모나이트를 사냥하던 중(목 부위에서 암모나이트 껍질 발견) 같은 서식지에 살았던 플리오사우루스의 공격을 받아 꼬리가 잘린 것으로 추정된다. 공격을 받은 어룡은 이후 깊은 바다로 가라앉아 익사했고, 심해저에서 화석화된 것으로 보인다. 이 발견은 내셔널 지오그래픽의 특별 프로그램 "바다 괴물의 죽음"(Death of a Sea Monstereng)을 통해 대중에게 알려졌다.[152]
참조
[1]
논문
Earliest Triassic ichthyosaur fossils push back oceanic reptile origins
[2]
웹사이트
Fossil Saved from Mule Track Revolutionizes Understanding of Ancient Dolphin-Like Marine Reptile
https://www.scienced[...]
2013-05-15
[3]
웹사이트
Malawania from Iraq and the Cretaceous Ichthyosaur Revolution (part II)
http://blogs.scienti[...]
2013-05-15
[4]
웹사이트
What are dinosaurs?
https://www.nhm.ac.u[...]
Natural History Museum, London
2024-11-04
[5]
서적
Lithophylacii Brittannici Ichnographia, sive Lapidum aliorumque Fossilium Brittanicorum singulari figurà insignium
[6]
서적
Piscium Querelae et Vindiciae
Gessner
[7]
서적
Descriptions and Figures of Petrifications Found in the Quarries, Gravel-Pits etc. Near Bath. Collected and Drawn by John Walcott, Esq.
S. Hazard
[8]
간행물
"The roles played by museums, collections, and collectors in the early history of reptile palaeontology", pp. 5–31 in: Richard Moody, E. Buffetaut, D. Naish, D. M. Martill (eds). Dinosaurs and Other Extinct Saurians: A Historical Perspective
Geological Society of London
[9]
간행물
Gentleman's Magazine
[10]
논문
Some Account of the Fossil Remains of an Animal More Nearly Allied to Fishes Than Any of the Other Classes of Animals
[11]
논문
Some Farther Account of the Fossil Remains of an Animal, of Which a Description Was Given to the Society in 1814
[12]
논문
Additional Facts Respecting the Fossil Remains of an Animal, on the Subject of Which Two Papers Have Been Printed in the Philosophical Transactions, Showing That the Bones of the Sternum Resemble Those of the Ornithorhynchus Paradoxus
[13]
논문
On the mode of formation of the canal containing the spinal marrow and of the form of the fins (if they deserve that name) of the Proteosaurus
[14]
논문
An account of the fossil skeleton of the Proteo-Saurus
[15]
논문
An account of the fossil skeleton of the Proteo-Saurus
[16]
서적
Icones Fossilium Sectiles
[17]
논문
Additional notices on the fossil genera Ichthyosaurus and Plesiosaurus
https://zenodo.org/r[...]
[18]
논문
Notice of the discovery of a new animal, forming a link between the Ichthyosaurus and crocodile, together with general remarks on the osteology of Ichthyosaurus
https://zenodo.org/r[...]
[19]
논문
Description de quelques espèces de reptiles de la Californie, précédée de l'analyse d'une système générale d'Erpetologie et d'Amphibiologie
[20]
논문
Report on British fossils reptiles
[21]
서적
Science and Religion in Neo-Victorian Novels: Eye of the Ichthyosaur
Routledge
[22]
논문
'The World-Renowned Ichthyosaurus': A Nineteenth-Century Problematic and Its Representations
[23]
논문
Account of a singular fossil skeleton, discovered at Whitby in February 1819
[24]
서적
Memoirs on Ichthyosauri and Plesiosauri; Extinct monsters of the ancient Earth
Relfe and Fletcher
[25]
서적
The Book of the Great Sea-dragons, Ichthyosauri and Plesiosauri, Gedolim Taninum of Moses. Extinct Monsters of the Ancient Earth
W. Pickering
[26]
서적
The Dragon Seekers: How an Extraordinary Circle of Fossilists Discovered the Dinosaurs and Paved the Way for Darwin
Basic Books
[27]
논문
On certain peculiarities in the cervical vertebrae of ''Ichthyosaurus'' hitherto unnoticed
[28]
간행물
"XXXVI.—Note on the Dislocation of the Tail at a certain point observable in the Skeleton of many Ichthyosauri", Transactions of the Geological Society of London
[29]
서적
De ichthyosauri sive proteosauri fossilis speciminibus in agro bollensi in Wirttembergia repertis
Cotta
[30]
논문
Über einen kolossalen ''Ichthyosaurus trigonodon''
[31]
논문
Über Ichthyosauren in den Lias-Schiefern der Gegend von Boll in Württemberg
[32]
논문
Über die Fortpflanzungsweise des ''Ichthyosaurus''
[33]
서적
Die Ichthyosaurier des Lias und ihre Zusammenhänge
Gebrüder Borntraeger
[34]
서적
The successful dragons: a natural history of extinct reptiles
Samuel Stevens & Company
[35]
논문
Phylogeny of the Ichthyopterygia
[36]
서적
"Ichthyopterygia — Handbuch der Paläoherpetologie Part 8"
Verlag Dr. Friedrich Pfeil, München
2003
[37]
논문
A giant Late Triassic ichthyosaur from the UK and a reinterpretation of the Aust Cliff 'dinosaurian' bones
2018-04-09
[38]
논문
The last giants: New evidence for giant Late Triassic (Rhaetian) ichthyosaurs from the UK
2024-04-17
[39]
웹사이트
Ancient fossils lead to discovery of largest known marine reptile
https://www.cnn.com/[...]
2024-04-17
[40]
논문
A basal ichthyosauriform with a short snout from the Lower Triassic of China
2014
[41]
논문
Die Frage nach der Herkunft der Ichthyosaurier
[42]
논문
On the cranial morphology of ichthyosaurs
[43]
논문
Ichthyosauria: A preliminary phylogenetic analysis of diapsid affinities
[44]
논문
Ichthyosaur relationships illuminated by new primitive skeletons from Japan
[45]
논문
A phylogenetic analysis of the euryapsid reptiles
[46]
논문
European origin of placodont marine reptiles and the evolution of crushing dentition in Placodontia
[47]
논문
Successive climate crises in the deep past drove the early evolution and radiation of reptiles
2022-08
[48]
논문
Ancestral body plan and adaptive radiation of sauropterygian marine reptiles
2022-12
[49]
논문
A basal ichthyosauriform with a short snout from the Lower Triassic of China
[50]
논문
Earliest Triassic ichthyosaur fossils push back oceanic reptile origins
[51]
문서
Maisch and Matzke 2000
[52]
논문
A new genus of ichthyosaur from the Late Triassic Pardonet Formation of British Columbia: bridging the Triassic-Jurassic gap
[53]
논문
The status of ''Himalayasaurus tibetensis'' (Ichthyopterygia)
http://mygeologypage[...]
2017-11-17
[54]
웹사이트
The sea dragons bounce back
http://phys.org/news[...]
[55]
논문
Resetting the evolution of marine reptiles at the Triassic-Jurassic boundary
[56]
문서
Motani 2000
[57]
논문
A functional analysis of the skull of ''Goronyosaurus nigeriensis'' (Squamata: Mosasauridae) and Its Bearing on the Predatory Behavior and Evolution of the Enigmatic Taxon
[58]
논문
Extinction of ichthyosaurs: a catastrophic or evolutionary paradigm?
[59]
간행물
"A severe drop in Eurasian ichthyosaur diversity prior to their late Cenomanian extinction: local or global signal?",
2012
[60]
논문
A basal thunnosaurian from Iraq reveals disparate phylogenetic origins for Cretaceous ichthyosaurs
[61]
논문
Cretaceous Ichthyosaurs: Dwindling Diversity, or the Empire Strikes Back?
[62]
논문
Extinction of fish-shaped marine reptiles associated with reduced evolutionary rates and global environmental volatility
2016-03-08
[63]
논문
New Ophthalmosaurid Ichthyosaurs from the European Lower Cretaceous Demonstrate Extensive Ichthyosaur Survival across the Jurassic–Cretaceous Boundary
[64]
논문
"'Rigid Swimmer' and the Cretaceous Ichthyosaur Revolution (part I)"
http://blogs.scienti[...]
2011
[65]
논문
The Ichthyosauria
http://www.naturkund[...]
2017-10-08
[66]
논문
Observations on Triassic ichthyosaurs. Part XII. A new Lower Triassic ichthyosaur genus from Spitzbergen
[67]
논문
Phylogeny, systematics, and origin of the Ichthyosauria – the state of the art
http://www.palaeodiv[...]
[68]
논문
Giant Ichthyosaurs of the Triassic — A New Species of ''Shonisaurus'' from the Pardonet Formation (Norian: Late Triassic) of British Columbia
[69]
논문
Giant ichthyosaurs of the Early Jurassic
[70]
웹사이트
Size of Ichthyosaurs
http://www.ucmp.berk[...]
2000-11-15
[71]
서적
Bent out of Shape
W. W. Norton & Co
[72]
논문
Ichthyosauria: their diversity, distribution, and phylogeny
[73]
논문
An ichthyosaur skull from the Cretaceous of Wyoming
[74]
논문
The cranial morphology of the Lower Liassic latipinnate ichthyosaurs of England
[75]
논문
The affinities and ecology of Triassic ichthyosaurs
[76]
논문
Reptiles that took on the sea
[77]
논문
An evolutionary and developmental perspective on the loss of regionalization in the limbs of derived ichthyosaurs
http://doc.rero.ch/r[...]
[78]
간행물
A Monograph of the Fossil Reptilia of the Liassic Formations. Part III, Ichthyopterygia
Palaeontographical Society
[79]
논문
Eel-like swimming in the earliest ichthyosaurs
[80]
논문
Soupy Substrates: A Medium for the Exceptional Preservation of Ichthyosaurs of the Posidonia Shale (Lower Jurassic) of Germany
[81]
논문
Prokaryote mats replacing soft tissues in Mesozoic marine reptiles
[82]
논문
Evolution of Fish-Shaped Reptiles (Reptilia: Ichthyopterygia) in Their Physical Environments and Constraints
[83]
논문
Rare soft-tissue preservation showing fibrous structures in an ichthyosaur from the Lower Lias (Jurassic) of England
[84]
논문
Palaeobiology of red and white blood cell-like structures, collagen and cholesterol in an ichthyosaur bone
[85]
논문
The ichthyosaur integument: skin fibers, a means for a strong, flexible and smooth skin
[86]
논문
A new ichthyosaur from the upper Jurassic lithographic limestones of Bavaria
https://cyberleninka[...]
[87]
논문
XCVII. On the colour of an ichthyosaur
[88]
논문
Skin pigmentation provides evidence of convergent melanism in extinct marine reptiles
[89]
웹사이트
Fossil pigments reveal the colors of ancient sea monsters
https://www.scienced[...]
[90]
논문
A guide to the field of palaeo colour
2015-06-01
[91]
논문
Gastroliths in the Triassic ichthyosaur ''Panjiangsaurus'' from China
http://doc.rero.ch/r[...]
[92]
논문
Further evidence for the wide geographical distribution of ichthyosaur taxa (Reptilia, Ichthyosauria)
[93]
논문
On the skin and food of ichthyosauri and teleosauri.
1856
[94]
논문
The gastric contents of an ichthyosaur from the Lower Lias of Lyme Regis, Dorset.
1968
[95]
논문
Euthynotus cf. incognitos (Actinopterygii, Pachycormidae) als Mageninhalt eines Fischsauriers aus dem Posidonieschiefer Suddeutschlands (unterer Jura, Lias Epsilon).
2000
[96]
논문
Uber die Nahrung eines Leptopterygius (Ichthyosauria, Reptilia) aus dem suddeutschen Posidonienschiefer (Unterer Jura) mit Bemerkungen uber den Magen der Icthyosaurier.
1989
[97]
서적
A revision of the Longipinnate Ichthyosaurs of the Lower Jurassic of England, with descriptions of two new species (Reptilia: Ichthyosauria)
https://archive.org/[...]
Royal Ontario Museum
[98]
논문
Macropredatory ichthyosaur from the Middle Triassic and the origin of modern trophic networks
[99]
간행물
A revision of the longipinnate ichthyosaurs of the Lower Jurassic of England, with descriptions of two new species (Reptilia: Ichthyosauria). Life Sciences Contribution of the Royal Ontario Museum 97.
[100]
논문
On the discovery of coprolites, or fossil faeces, in the Lias at Lyme Regis, and in other formations
https://zenodo.org/r[...]
[101]
논문
The gastric contents of an ichthyosaur from the Lower Lias of Lyme Regis, Dorset
[102]
논문
Unusual gut contents in a Cretaceous ichthyosaur
[103]
논문
Short-Snouted toothless ichthyosaur from China suggests late Triassic diversification of suction feeding ichthyosaurs
[104]
논문
Absence of suction feeding ichthyosaurs and its implications for Triassic mesopelagic paleoecology
[105]
논문
A unique advantage for giant eyes in giant squid
[106]
간행물
"Reassessment of the English Cretaceous ichthyosaur ''Platypterygius campylodon''"
The University of Glasgow Press, Glasgow
2007
[107]
논문
Tooth morphology and prey preference of Mesozoic marine reptiles
1987-06-18
[108]
논문
A plesiosaur containing an ichthyosaur embryo as stomach contents from the Sundance Formation of the Bighorn Basin, Wyoming
https://mds.marshall[...]
2019-11-30
[109]
논문
Differential growth in three ichthyosaurs: ''Ichthyosaurus communis'', ''I. breviceps'', and ''Stenopterygius quadriscissus'' (Reptilia, Ichthyosauria)
https://www.archive.[...]
1973
[110]
논문
A reinterpretation of ichthyosaur swimming and buoyancy
1987
[111]
논문
The ichthyosaurian tail: sharks do not provide an appropriate analogue
1992
[112]
논문
Scaling effects in caudal fin propulsion and the speed of ichthyosaurs
2002
[113]
논문
Float, explode or sink: post-mortem fate of lung-breathing marine vertebrates
http://doc.rero.ch/r[...]
2012
[114]
서적
"Swimming capabilities of Mesozoic marine reptiles: a review"
Cambridge University Press
1994
[115]
논문
Swimming speed estimation of extinct marine reptiles: energetic approach revisited
2002
[116]
간행물
"How to reconstruct paleoecology? – Outlines of a holistic view and an introduction to ichthyosaur locomotion"
Attempto Verlag (Tübingen)
1984
[117]
간행물
"Biomechanics of ichthyosaurs"
1985
[118]
논문
Fortbewegungsweise, Schwimmbiophysik und Phylogenie der Ichthyosaurier
1986
[119]
논문
Locomotion and phylogeny of the ichthyosaurs
1989
[120]
논문
Did ichthyosaurs fly?
1998
[121]
논문
Locomotory capabilities in the Early Cretaceous ichthyosaur ''Platypterygius australis'' based on osteological comparisons with extant marine mammals
2013
[122]
논문
Adaptations for marine habitat and the effect of Triassic and Jurassic predator pressure on development of decompression syndrome in ichthyosaurs
2012
[123]
논문
Deep-diving dinosaurs
2012
[124]
논문
Large eyes in deep diving ichthyosaurs
1999
[125]
논문
What to do with a 10-inch eyeball? – Evolution of vision in ichthyosaurs
1999
[126]
논문
Bone histology of the ichthyosaurs: comparative data and functional interpretation
1990
[127]
논문
Caractères structuraux et mode de croissance du femur d'''Omphalosaurus nisseri'', ichthyosaurien du Trias moyen de Spitsberg
1987
[128]
논문
The palaeohistology of the basal ichthyosaur ''Mixosaurus'' Baur, 1887 (Ichthyopterygia, Mixosauridae) from the Middle Triassic: Palaeobiological implications
2011
[129]
논문
"Osteohistology of ''Utatsusaurus hataii'' (Reptilia: Ichthyopterygia): Implications for early ichthyosaur biology"
2012
[130]
논문
Regulation of body temperature by some Mesozoic marine reptiles
2010
[131]
웹사이트
Warm-Blooded Marine Reptiles at the Time of the Dinosaurs
https://www.scienced[...]
Science Daily
2010-06-14
[132]
논문
Soft-tissue evidence for homeothermy and crypsis in a Jurassic ichthyosaur
https://lup.lub.lu.s[...]
2018
[133]
논문
Evolution of fish–shaped reptiles (Reptilia: Ichthyopterygia) in their physical environments and constraints
2005
[134]
논문
The evolution of locomotor stamina in tetrapods: circumventing a mechanical constraint
1987
[135]
서적
"Locomotion and respiration in marine air-breathing vertebrates"
University of Chicago Press
1996
[136]
논문
Über einen möglichen Auftauchmodus bei den Ichthyosauriern
1993
[137]
논문
Notice of what appears to be the Embryo of an ''Ichthyosaurus'' in the Pelvic cavity of ''Ichthyosaurus'' (''communis''?)
https://zenodo.org/r[...]
1846
[138]
논문
Report on the mode of reproduction of certain species of ''Ichthyosaurus'' from the Lias of England and Würtemberg, by a Committee
1880
[139]
논문
An ichthyosaur embryo from the Lower Lias (Jurassic: Hettangian) of Somerset, England, with comments on the reproductive biology of ichthyosaurs
1993
[140]
웹사이트
Ancient reptile birth preserved in fossil: Ichthyosaur fossil may show oldest live reptilian birth
https://www.scienced[...]
[141]
논문
Terrestrial Origin of Viviparity in Mesozoic Marine Reptiles Indicated by Early Triassic Embryonic Fossils
[142]
논문
Heads or tails first? Evolution of fetal orientation in ichthyosaurs, with a scrutiny of the prevailing hypothesis
[143]
논문
Neue Erkenntnisse zur Fortpflanzungsbiologie der Ichthyosaurier (Reptilia)
[144]
논문
In utero foetal remains of the Cretaceous ichthyosaurian ''Platypterygius'': ontogenetic implications for character state efficacy
[145]
웹사이트
The first known neonate Ichthyosaurus communis skeleton from the Lower Jurassic, UK
http://dx.doi.org/10[...]
2024-11-26
[146]
논문
Trophic niche ontogeny and palaeoecology of early T oarcian S tenopterygius ( R eptilia: I chthyosauria)
https://onlinelibrar[...]
2016-05
[147]
논문
Genotypic sex determination enabled adaptive radiations of extinct marine reptiles
[148]
뉴스
For Extinct Monsters of the Deep, a Little Respect
https://www.nytimes.[...]
2010-03-22
[149]
논문
The sexual dimorphism of ''Shastasaurus tangae'' (Reptilia: Ichthyosauria) from the Triassic Guanling Biota, China
[150]
논문
Healed bite marks on a Cretaceous ichthyosaur
[151]
웹사이트
Battle scars found on an ancient sea monster
https://www.scienced[...]
[152]
간행물
Death of a Sea Monster
National Geographic Channel
2011
[153]
뉴스
Indian Ichthyosaur fossil proves this ancient sea monster roamed the world
http://www.news.com.[...]
News Corp Australia Network
2017-10-30
[154]
논문
Discovery of the first ichthyosaur from the Jurassic of India: Implications for Gondwanan palaeobiogeography
2017-10-25
[155]
웹사이트
Stunning Jurassic 'Sea Monster' Found in India
https://news.nationa[...]
National Geographic
2017-10-25
[156]
서적
宇宙137億年の謎が2時間でわかる本
KAWADE夢文庫
[157]
논문
The largest hupehsuchian (Reptilia, Ichthyosauromorpha) from the Lower Triassic of South China indicates early establishment of high predation pressure after the Permo-Triassic mass extinction
https://doi.org/10.1[...]
2030-03-19
[158]
논문
Large eyeballs in diving ichthyosaurs
http://mygeologypage[...]
2020-03-25
[159]
논문
レスキューとしての企画展 「復興、南三陸町・歌津魚竜館」─世界最古の魚竜のふるさと
http://www.museum.to[...]
東北大学総合学術博物館
2020-03-18
[160]
논문
New Ophthalmosaurid Ichthyosaurs from the European Lower Cretaceous Demonstrate Extensive Ichthyosaur Survival across the Jurassic–Cretaceous Boundary
[161]
논문
セノマニアン/チューロニアン期境界前後の軟体動物相 : 北海道大夕張地域と米国西部内陸地域の比較(<特集>白亜紀海洋無酸素事変の解明)
https://doi.org/10.1[...]
日本古生物学会
[162]
논문
Extinction of fish-shaped marine reptiles associated with reduced evolutionary rates and global environmental volatility
2016-03-08
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com