완족동물
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
완족동물은 좌우대칭의 체강동물로, 2개의 껍데기를 가지고 있으며, 촉수관을 이용해 먹이를 섭취한다. 전 세계적으로 300종 이상이 알려져 있으며, 화석은 캄브리아기부터 발견된다. 이들은 껍데기의 구조와 부착 방식, 촉수관의 형태 등에 따라 다양한 특징을 보인다. 완족동물은 전통적으로 무관절강과 유관절강으로 분류되었으나, 현재는 설각아문, 두각아문, 부리각아문의 세 아문으로 분류된다. 완족동물은 켄터키주의 주 화석으로 지정되었으며, 일부 종은 식용으로 이용되기도 한다.
더 읽어볼만한 페이지
- 완족동물 - 개맛
개맛은 녹색 직사각형 껍데기와 말굽 모양 육경을 가진 연체동물로, 육경을 이용해 호흡하고 먹이를 포획하며 점액성 관 속에서 플랑크톤을 섭취하며 해저에 서식한다. - 완족동물 - 무관절강
- 앙드레 마리 콘스탕 듀메릴이 명명한 분류군 - 원구류
원구류는 멸종된 갑주어류를 포함하는 무악류에 속하며, 칠성장어류와 먹장어류를 포함하고 턱이 없으며, 흉선, 비장, 수초, 교감 신경절이 없고, 턱이 있는 척추동물과는 독립적으로 산소 운반 헤모글로빈을 진화시켰다. - 앙드레 마리 콘스탕 듀메릴이 명명한 분류군 - 베일드카멜레온
베일드카멜레온은 아라비아반도 남서부 원산의 수목 생활을 하는 카멜레온 종으로, 머리의 투구와 뛰어난 색 변화 능력, 유연한 꼬리, 긴 혀 등의 특징을 가지며 애완동물로 거래되거나 침입종으로 간주되기도 한다. - 생물 분류에 관한 - 다람쥐
다람쥐는 등 쪽에 줄무늬가 있는 다람쥐속 설치류로, 홀로 생활하며 겨울잠을 자고 씨앗, 견과류, 곤충 등을 먹으며 맹금류 등의 먹이가 되고, 특히 한반도 서식 다람쥐는 새로운 종으로 분류될 가능성이 있으며, 유럽에서는 침입 외래종으로 지정되기도 하고 라임병을 옮길 수 있어 주의가 필요한 동물이다. - 생물 분류에 관한 - 황금랑구르
황금랑구르는 인도 아삼 주와 부탄에 분포하며 크림색에서 황금색 털을 가진 멸종위기종 영장류로, 서식지 파괴로 개체수가 감소하여 보호받고 있다.
| 완족동물 - [생물]에 관한 문서 | |
|---|---|
| 지도 정보 | |
| 기본 정보 | |
| 학명 | Brachiopoda |
| 명명자 | Duméril, 1806 |
| 영어 이름 | lamp shell |
| 다양성 | 약 100개의 현존하는 속, 약 5,000개의 화석 속 |
| 분류 | |
| 계 | 진핵생물 |
| 역 | 동물계 |
| 아계 | 진정후생동물아계 |
| 상문 | 관윤동물상문 |
| 문 | 완족동물문 |
| 하위 분류 | 무관절강 유관절강 또는 두개강 설형강 능형강 |
| 형태 및 특징 | |
| 형태 | 이매패류와 유사한 껍데기를 가짐 |
| 특징 | 껍데기는 등쪽과 배쪽의 두 개로 구성 껍데기 내부에 로포포어라는 촉수관 존재 대부분 해저에 부착 생활 |
| 생태 및 분포 | |
| 서식지 | 해양 (주로 얕은 바다) |
| 분포 | 전 세계 해양 |
| 먹이 | 플랑크톤 |
| 생활 방식 | 대부분 고착 생활 |
| 역사 | |
| 화석 기록 | 캄브리아기 초기부터 현재까지 |
| 경제적 가치 | |
| 경제적 가치 | 일부 종은 식용으로 이용되기도 함 |
| 사회경제적 가치 | 태국 망그로브 생태계에서 살아있는 화석, 링굴라 종의 사회경제적 가치 연구 |
| 기타 | |
| 참고 | 갯지렁이라고도 불림 |
2. 특징
완족동물은 선구동물과 후구동물의 특징을 모두 가진 좌우대칭의 체강동물이다. 2장의 각(shell)과 외투를 가지는 점에서 연체동물의 부족류와 유사하지만, 2각의 크기가 다르고 2각이 배복으로 붙어 있다는 점에서 부족류와 구별된다.
내부에는 내장낭(內臟囊, visceral sac), 2개의 외투(外套, mantle) 및 2개의 완(腕, arm)이라 불리는 촉수관이 있고 병부(자루)는 각의 바깥으로 길게 나와 있다. 외투에는 강모가 나 있으며, 이들은 외투 표피에서 자극을 전달하는 감각기로 작용한다. 근육은 체벽, 완, 각의 기부 및 병부에 발달한다.
소화관은 체강 내에 있으며 입, 식도, 소화샘으로 둘러싸인 위, 장, 직장, 항문으로 구성된다. 유관절류에는 항문이 없고 소화관이 막혀 있다. 혈관계는 개방혈관계이고 심장은 위의 위쪽에 있다. 배설기는 1쌍 또는 2쌍의 후신관으로, 내장낭 아랫부분에 있는 누두와 체강에 열려 있다. 유생은 자유유영을 한다.
완족동물은 촉수관을 이용하여 먹이를 섭취한다. 촉수관은 입을 둘러싼 촉수의 고리이며, 1쌍의 팔에 많은 가는 촉수가 나와서 형성된다. 촉수의 섬모 운동으로 물의 흐름을 만들어 먹이 입자를 걸러낸다. 무관절류의 소화관은 굽어져 항문으로 연결되지만, 관절류는 항문이 없다.
순환계는 개방순환계이지만 불완전하며, 장간막 위에 심장이 있다. 혈액과 체강액은 별개로 여겨진다. 가스 교환은 체표에서 이루어진다. 1쌍 또는 2쌍의 신관은 생식관 역할도 한다. 신경계는 등쪽과 배쪽에 신경절이 있고, 신경환으로 연결되어 있다.
2. 1. 껍데기 구조와 기능
완족류는 두 개의 껍데기(각)를 가지는데, 이는 동물의 등쪽(윗면)과 배쪽(아랫면)을 덮는다. 이는 껍데기가 측면(옆면)을 덮는 이매패류 연체동물과 다르다. 두 껍데기의 크기와 구조는 서로 다르며, 각각 독자적인 대칭적인 형태를 가지고 있고 서로 거울상이 아니다. 완족류 껍데기의 발생 과정은 호메오박스 유전자를 포함한 보존된 유전자 세트에 기반하는데, 이 유전자들은 연체동물의 껍데기 형성에도 사용된다.[5]등쪽 껍데기는 일반적으로 더 작으며, 안쪽 표면에 brachia("팔")를 가지고 있다. 이 brachia는 문의 이름의 유래이며, 먹이 섭취와 호흡에 사용되는 촉수관을 지지한다. 배쪽 껍데기는 일반적으로 더 크며, 경첩 근처에 자루를 위한 구멍이 있다. 대부분의 완족류는 이 자루를 통해 기질에 부착된다. 등쪽 및 배쪽 껍데기는 각각 배측 및 복측 껍데기라고도 불리지만, 일부 고생물학자들은 "배측"과 "복측"이라는 용어가 관련이 없다고 생각한다. 왜냐하면 그들은 "복측" 껍데기가 몸 아래쪽의 상면이 접힘으로써 형성되었다고 믿기 때문이다. 대부분의 완족류는 생활 위치에서 배쪽("아래쪽") 껍데기가 실제로 배측("위쪽") 껍데기 위에 위치한다. 많은 현생 관절완족류 종에서 두 껍데기는 모두 볼록하며, 표면에는 종종 성장선이나 기타 장식이 있다. 그러나 해저에 서식하는 무관절완족류인 설선류는 껍데기가 더 매끄럽고 평평하며 크기와 모양이 비슷하다.

관절완족류("관절")는 이빨과 소켓 구조를 가지고 있어 배쪽 및 등쪽 껍데기가 경첩으로 연결되고 측면 이동이 막힌다. 무관절완족류는 일치하는 이빨과 소켓이 없으며, 껍데기는 근육에 의해서만 연결된다.
모든 완족류는 배쪽 껍데기 안쪽에 위치한 내전근을 가지고 있으며, 이 근육은 경첩 앞쪽의 등쪽 껍데기 부분을 당겨 껍데기를 닫는다. 이 근육에는 비상시에 껍데기를 빠르게 닫는 "빠른" 섬유와 속도는 느리지만 껍데기를 오랫동안 닫아둘 수 있는 "고정" 섬유가 모두 있다. 관절완족류는 더 뒤쪽에 위치하고 경첩 뒤쪽의 등쪽 껍데기 부분을 당기는 외전근(또는 벌림근)을 사용하여 껍데기를 연다. 무관절완족류는 다른 개폐 메커니즘을 사용하는데, 근육이 체강(주요 체강)의 길이를 줄이고 바깥쪽으로 팽창시켜 껍데기를 벌린다. 두 강 모두 껍데기를 약 10도 각도로 연다. 무관절완족류가 사용하는 더 복잡한 근육 세트는 껍데기를 가위처럼 작동시킬 수도 있는데, 이는 설선류가 굴을 파는 데 사용하는 메커니즘이다.
각 껍데기는 세 층으로 구성되어 있으며, 바깥쪽에는 유기 화합물로 이루어진 각피층, 그 아래에는 두 개의 생체광물화 층이 있다. 관절완족류는 단백질로 이루어진 가장 바깥쪽 각피층, 그 아래에 방해석(탄산칼슘의 한 형태)으로 이루어진 "주층", 그리고 가장 안쪽에 단백질과 방해석의 혼합물이 있다. 무관절완족류 껍데기는 유사한 층 구조를 가지고 있지만, 그 구성은 관절완족류와 다르며, 무관절완족류의 강에 따라서도 다르다. 테레브라툴리다는 구멍이 있는 껍데기 구조를 가진 완족류의 한 예이다. 광물화된 층은 생체 조직의 작은 열린 관인 맹낭에 의해 구멍이 뚫려 있으며, 맹낭은 맨틀의 연장으로 주층의 바깥쪽에 거의 도달한다. 이러한 껍데기는 동물 생체 조직의 절반을 포함할 수 있다. 구멍이 없는 껍데기는 내부에 조직이 없는 단단한 껍데기이다. 가짜 구멍이 있는 껍데기는 방해석 막대를 따라 펼쳐지는 변형으로 형성된 결절을 가지고 있다. 이는 화석 형태에서만 알려져 있으며, 원래는 석회화된 구멍이 있는 구조로 잘못 해석되었다.[7][8]
자루를 가진 설선류와 원반류는 다른 물질이 끼워진 기질의 글리코사미노글리칸(길고 가지가 없는 다당류)을 가지고 있다. 각피층에는 키틴이 있고, 주 생체광물화층에는 인회석을 포함하는 인산칼슘이 있으며,[9] 가장 안쪽 층에는 콜라겐 및 기타 단백질, 키틴인산염 및 인회석을 포함한 복합 혼합물이 포함되어 있다. 자루가 없고 단단한 표면에 직접 고착되는 크라니이다는 키틴으로 이루어진 각피층과 방해석으로 이루어진 광물화 층을 가지고 있다. 껍데기 성장은 전주변성, 혼합주변성 또는 반주변성으로 설명할 수 있다. 크라니이다의 특징인 전주변성 성장에서는 새로운 물질이 가장자리를 따라 고르게 추가된다. 많은 현생 및 멸종 관절완족류에서 발견되는 혼합주변성 성장에서는 새로운 물질이 껍데기의 후방 영역에 추가되고 전방 경향을 보이며 다른 껍데기쪽으로 자란다. 설선류에서 발견되는 반주변성 성장은 혼합주변성 성장과 유사하지만, 대부분 평평한 판에서 일어나며 껍데기가 앞쪽과 바깥쪽으로 자란다.[10]
이매패류처럼 두 개의 껍데기를 가지지만, 이매패류의 껍데기는 몸의 좌우에 각각 하나씩 있는 반면, 완족동물의 껍데기는 배-등쪽에 있다고 여겨진다. 껍데기의 성분은 분류군에 따라 다르며, 관절류와 일부 무관절류는 탄산칼슘을, 다른 종류는 키틴질의 인산칼슘을 주성분으로 한다. 각각의 껍데기는 좌우대칭이지만, 등쪽 껍데기와 배쪽 껍데기의 모양은 다르다. 두 개의 껍데기는 관절류에서는 경첩으로 연결되지만, 무관절류는 경첩이 없고, 껍데기는 근육으로 연결된다.
2. 2. 부착 및 이동
대부분의 현생 완족동물 종들은 원통형의 병족(줄기)이라 불리는 신체 벽의 돌출부를 통해 단단한 표면에 부착한다. 병족은 키틴질의 표피(비세포성 "피부")를 가지고 있으며, 경첩 부분의 구멍을 통해 돌출된다.[12] 그러나, 무관절류인 ''크라니아(Crania)''와 관절류인 ''라카젤라(Lacazella)''처럼 병족이 없는 속도 있다. 이들은 "병족"(복측) 밸브의 후면을 표면에 고정시켜 전면이 표면에서 약간 위쪽으로 기울어지도록 한다.[12] 이러한 완족류에서는 복측 밸브에 병족 개구부가 없다.[12] ''네오티리스(Neothyris)''와 ''아나키네티카(Anakinetica)''와 같은 일부 관절류 속에서는 성체가 자람에 따라 병족이 퇴화하여 결국 표면에 느슨하게 놓이게 된다.[12]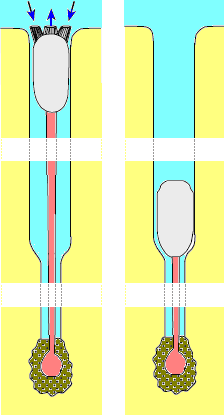
무관절류 완족류의 병족은 내부 기관을 수용하는 주 체강의 연장이다. 세로로 배열된 근육층이 병족의 표피를 둘러싸고 있다.[12] 목 혀모양완족류(Lingulida)에 속하는 종들은 긴 병족을 가지고 있으며, 이를 이용하여 부드러운 기질 속으로 파고들어가거나, 먹이를 먹기 위해 껍데기를 굴의 개구부까지 들어올리거나, 방해를 받으면 껍데기를 안으로 끌어들인다. 혀모양완족류는 굴의 위쪽 3분의 2 지점에서 몸을 위아래로 움직인다.[12] 그러나, 원반완족류(Discinida) 목의 병족은 짧고 단단한 표면에 부착한다.[12]
관절류 완족류의 병족에는 체강이 없으며, 그 상동은 불분명하다. 이는 유생의 신체의 다른 부분으로부터 형성되며, 결합조직으로 구성된 치밀한 중심부를 가지고 있다. 신체 후면의 근육은 병족을 곧게 펴거나 구부리거나 심지어 회전시킬 수 있다. 병족의 먼 쪽 끝은 일반적으로 뿌리 모양의 돌기 또는 짧은 유두("혹")를 가지고 있으며, 이를 통해 단단한 표면에 부착한다. 그러나, ''클리도노포라(Chlidonophora)'' 속의 관절류 완족류는 가지가 있는 병족을 이용하여 퇴적물에 고정된다. 일부 종은 전단이 위쪽으로 향하고 서 있고, 다른 종들은 병족 밸브가 위쪽으로 향한 채 수평으로 누워 있다.[12]
일부 초기 완족류—예를 들어 스트로포메나타(Strophomenata), 쿠토르기나타(Kutorginata) 및 오보렐라타(Obolellata)—는 병족을 사용하여 부착하지 않고, 병족과는 아무런 관련이 없는 "병족 초피"라고 알려진 완전히 다른 구조를 사용한다.[13][14]
2. 3. 촉수관
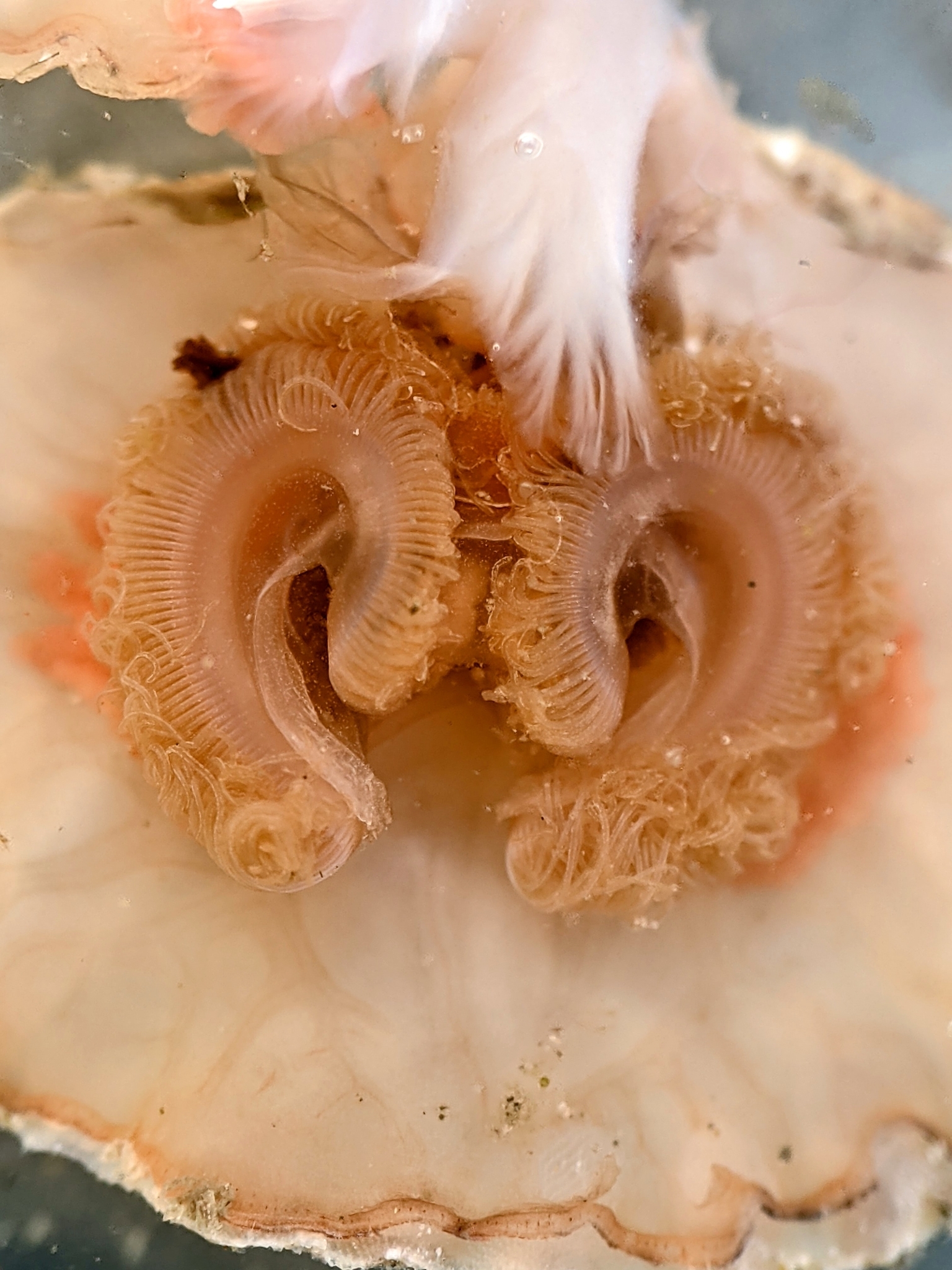
이끼벌레류와 포로니드류처럼, 완족류도 촉수관(lophophore)을 가지고 있다. 촉수관은 섬모(미세한 털)를 가진 촉수의 관으로, 물의 흐름을 만들어 물에서 먹이 입자를 걸러 먹는 것을 가능하게 한다. 이끼벌레류나 포로니드류의 촉수관은 단일하고 안으로 끌어들일 수 있는 줄기에 달린 촉수의 고리 모양인 반면, 완족류 촉수관의 기본 형태는 U자 모양이며, 문(phylum)의 이름의 유래가 된 팔(brachia, "arms")을 형성한다. 완족류의 촉수관은 안으로 끌어들일 수 없으며, 껍데기가 열릴 때 벌어지는 가장 앞쪽 부분의 내부 공간의 최대 3분의 2를 차지한다. 이 제한된 공간에서 충분한 여과 능력을 제공하기 위해, 큰 완족류의 촉수관은 중간 정도에서 매우 복잡한 모양으로 접혀 있다. 고리와 나선 모양이 흔하며, 어떤 종의 촉수관은 손가락을 펼친 손과 비슷한 모양으로 뒤틀린다. 모든 종에서 촉수관은 연골과 수정체골격(즉, 내부 체액의 압력)에 의해 지지되며, 체액은 촉수로 뻗어 있다. 일부 관절완족류는 팔껍데기(brachial valve) 안쪽에 부착된 촉수관을 지지하는 석회질 지지대인 브라키디움(brachidium)을 가지고 있는데, 이는 촉수관 근육의 극심한 감소와 일부 팔 신경의 감소로 이어졌다.[11]껍데기는 외투막에서 분비되어 만들어진다. 외투막은 껍데기의 안쪽을 덮고 있으며, 껍데기 안의 외투막으로 덮인 공간, 즉 외투강을 형성한다. 외투강은 물로 채워져 있으며, 촉수관이 있다. 촉수관은 입을 둘러싼 촉수의 고리로, 완족동물에서는 1쌍의 팔(arm영어)에 많은 가는 촉수가 나와서 이루어져 있다. 관절류에서는 이 팔은 팔골에 의해 지지되지만, 무관절류는 팔골을 갖지 않고, 촉수관은 체강액의 압력으로 지지된다.
촉수는 가장자리와 중앙을 따라 섬모(미세하게 움직이는 털)를 가지고 있다. 바깥쪽 섬모의 운동은 촉수 끝에서 기저부로 물의 흐름을 유도하고, 그곳에서 물이 빠져나간다. 촉수와 충돌하는 먹이 입자는 점액에 의해 포획되며, 중앙의 섬모는 이 혼합물을 촉수 기저부로 이동시킨다. 촉수 기저부를 따라 팔홈(brachial groove)이 이어지며, 이 홈의 섬모는 먹이를 입 쪽으로 이동시킨다. 완족류가 사용하는 방법은 "상류 수집(upstream collecting)"으로 알려져 있는데, 먹이 입자가 먹이 흐름을 만드는 섬모의 영역으로 들어올 때 포획된다. 이 방법은 관련된 포로니드류와 이끼벌레류, 그리고 익족류도 사용한다. 내장형동물류는 비슷하게 생긴 촉수관을 사용하지만, 이것은 단단하고 흐름은 기저부에서 끝으로 향하며, "하류 수집(downstream collecting)" 시스템을 형성하여 먹이 입자가 빠져나가려고 할 때 포획한다.
2. 4. 내부 구조
완족동물의 몸은 껍데기 내부 공간의 약 1/3만 차지하며, 경첩에 가장 가깝다. 나머지 공간은 맨틀 엽으로 둘러싸여 있으며, 이는 촉수관이 위치한 물로 채워진 공간을 둘러싼 돌출부이다.[1] 체강(체강)은 각 엽으로 관의 그물망으로 뻗어 있으며, 이는 맨틀 가장자리로 영양분을 운반한다.[2]맨틀 가장자리의 홈에 있는 비교적 새로운 세포는 외피층을 확장하는 물질을 분비한다. 이러한 세포는 홈에 있는 더 최근의 세포에 의해 맨틀 아랫면으로 점차 이동하고, 껍데기 판의 광물화된 물질을 분비하기 시작한다. 다시 말해, 판의 가장자리에서 외피층이 먼저 확장된 다음 외피층 아래 광물화된 층의 확장에 의해 강화된다.[2] 대부분의 종에서 맨틀 가장자리는 흔히 강모 또는 센털이라고 불리는 움직일 수 있는 강모를 가지고 있으며, 이는 동물을 방어하는 데 도움이 되고 감각기 역할을 할 수 있다. 일부 완족류에서는 강모 그룹이 맨틀강 안팎으로 물의 흐름을 제어하는 데 도움이 된다.[1]
대부분의 완족류에서, 맨틀의 맹낭(속이 빈 돌기)은 판의 광물화된 층을 관통하여 외피층으로 침투한다. 이러한 맹낭의 기능은 불확실하며, 글리코겐과 같은 화학 물질의 저장실일 수도 있고, 껍데기에 달라붙는 유기체를 막는 방어 물질을 분비할 수도 있고, 호흡에 도움이 될 수도 있다는 제안이 있다.[1] 실험에 따르면, 껍데기에 바셀린을 바르면 맹낭이 막혀 완족류의 산소 소비량이 감소한다.[2]
촉수관과 외투막만이 산소를 흡수하고 이산화탄소를 배출하는 표면이다. 산소는 체강의 체액에 의해 분포되는 것으로 보이며, 체강의 안감 수축이나 섬모의 박동에 의해 외투막을 통해 순환된다. 일부 종에서는 산소가 부분적으로 호흡 색소인 헤메리트린에 의해 운반되는데, 이는 체강 세포 내에서 운반된다.[1] 완족류의 최대 산소 소비량은 낮고, 최소 요구량은 측정할 수 없다.
완족류는 또한 위쪽에 있는 몸의 등쪽 부분에 위치한 근육질 심장에 의해 순환되는 무색의 혈액을 가지고 있다.[1] 혈액은 몸의 앞뒤로 뻗어 있는 혈관을 통과하며, 앞쪽의 촉수관과 뒤쪽의 장, 근육, 생식선 및 신관을 포함한 기관으로 분지된다. 혈액 순환은 완전히 폐쇄되지 않은 것으로 보이며, 체강액과 혈액은 어느 정도 섞여야 한다.[2] 혈액의 주요 기능은 영양분을 전달하는 것일 수 있다.[1]
성체 관절완족류의 "뇌"는 식도 위와 아래에 각각 하나씩, 두 개의 신경절로 이루어져 있다. 성체 무관절완족류는 아래쪽 신경절만 가지고 있다.[3] 신경절과 이들이 연결되는 교련에서 신경은 촉수관, 외투엽, 그리고 각 밸브를 작동시키는 근육으로 뻗어나간다. 외투의 가장자리에는 아마도 가장 많은 감각 수용체가 밀집해 있을 것이다. 감각 신경 세포에 직접 연결되어 있지는 않지만, 외투의 강모는 아마도 외투의 표피에 있는 수용체에 촉각 신호를 보낼 것이다. 많은 완족류는 위에 그림자가 나타나면 각 밸브를 닫지만, 이에 책임이 있는 세포는 알려져 있지 않다. 어떤 완족류는 동물의 위치 변화를 감지하는 평형낭을 가지고 있다.[1]
완족동물은 진체강을 가진 좌우대칭동물이다. 이매패류처럼 두 개의 껍데기를 가지지만, 이매패류의 껍데기는 몸의 좌우에 각각 하나씩 있는 반면, 완족동물의 껍데기는 배-등쪽에 있다고 여겨진다. 껍데기의 성분은 분류군에 따라 다르며, 관절류와 일부 무관절류는 탄산칼슘을, 다른 종류는 키틴질의 인산칼슘을 주성분으로 한다. 각각의 껍데기는 좌우대칭이지만, 등쪽 껍데기와 배쪽 껍데기의 모양은 다르다. 두 개의 껍데기는 관절류에서는 경첩으로 연결되지만, 무관절류는 경첩이 없고, 껍데기는 근육으로 연결된다. 껍데기 길이는 5센티미터 전후인 것이 많다.
배각의 후단에서 자루가 뻗어 나온다. 자루는 체벽이 뻗어 나와서 만들어진 것으로, 무관절류에서는 체강과 근육을 포함하며, 신축운동을 하지만, 관절류의 자루는 그것들을 갖추지 않고, 운동에 도움이 되지 않는다. 종에 따라 자루의 끝에 돌기가 있어서, 해저에 부착하는 데 사용된다. 자루가 없는 종도 있다.
껍데기는 외투막에서 분비되어 만들어진다. 외투막은 껍데기의 안쪽을 덮고 있으며, 껍데기 안의 외투막으로 덮인 공간, 즉 외투강을 형성한다. 외투강은 물로 채워져 있으며, 촉수관이 있다. 촉수관은 입을 둘러싼 촉수의 고리로, 완족동물에서는 1쌍의 팔에 많은 가는 촉수가 나와서 이루어져 있다. 관절류에서는 이 팔은 팔골에 의해 지지되지만, 무관절류는 팔골을 갖지 않고, 촉수관은 체강액의 압력으로 지지된다.
소화관은 U자형이다. 촉수관의 운동에 의해 입으로 들어온 먹이는 식도를 통해 위, 장으로 운반된다. 무관절류에서는 소화관이 굽어져 직장에 연결되고, 외투강의 안쪽이나 오른쪽에 열리는 항문으로 끝나지만, 관절류는 항문이 없고, 소화관은 막다른 곳(맹낭)이 된다.
순환계는 개방순환계이지만 불완전하다. 장간막 위에 심장을 가진다. 진정한 혈관은 없고, 복막으로 둘러싸인 관이 있다. 혈액과 체강액은 별개라고 여겨진다. 가스 교환은 체표에서 이루어진다. 1쌍 또는 2쌍의 신관을 가지며, 이것은 생식관의 역할도 한다.
신경계는 그다지 발달하지 않았다. 등쪽과 배쪽에 신경절이 있으며, 2개의 신경절은 신경환으로 연결되어 있다. 이러한 신경절과 신경환에서 전신으로 신경이 뻗어나간다.
3. 생태
완족동물은 모두 바다에 살며, 민물에 사는 종은 알려져 있지 않다. 대부분의 종은 강한 조류나 파도가 있는 곳을 피하며, 암석의 돌출부, 갈라진 틈, 동굴, 대륙붕의 가파른 경사면 및 심해저와 같은 곳에 서식한다. 그러나 일부 관절완족류는 켈프에 부착하거나, 예외적으로 조용한 조간대에서 서식하기도 한다. 가장 작은 완족동물인 ''Gwynia''는 길이가 약 에 불과하며, 자갈 알갱이 사이에서 산다.
알에서 부화한 후 노른자만 먹고 빠르게 정착하여 발달하는 측완류(Rhynchonelliforms)는 특정 지역에 고유종으로 존재하는 경우가 많으며, 제곱미터당 수천 마리에 달하는 밀집된 개체군을 형성한다. 어린 개체는 종종 더 성숙한 개체의 껍데기에 부착한다. 반면에, 유생이 정착하기 전에 최대 한 달 동안 헤엄치는 무관절완족류는 넓은 분포역을 가진다. 원반형(discinoid) 속인 ''Pelagodiscus''는 전 세계적으로 분포한다.
완족동물은 대사율이 낮아, 이매패류의 1/3에서 1/10 수준이다. 백악기 시대에 따뜻하고 얕은 바다에서 풍부했던 완족동물의 서식지는 이제 대부분 이매패류가 차지하고 있으며, 현재 대부분의 완족동물은 차갑고 빛이 적은 환경에서 서식한다.
완족동물의 껍데기에는 때때로 포식자에 의한 손상 흔적과 그 이후의 복구 흔적이 나타난다. 어류와 갑각류는 완족동물의 살을 맛없어하는 것으로 보인다. 화석 기록에 따르면, 복족류와 같은 굴착성 포식자는 이매패류나 성게류를 완족동물보다 10~20배 더 자주 공격했는데, 이는 이러한 포식자들이 실수로 완족동물을 공격하거나 다른 먹이가 부족할 때 공격했음을 시사한다. 먹이가 부족한 지역에서는 달팽이 ''Capulus ungaricus''가 이매패류, 달팽이, 관벌레, 완족동물로부터 먹이를 훔친다.
두흉류는 소규모로 상업적으로 어획된다. 주로 먹는 부분은 다육질의 자루이다.[21][22][23][24] 완족동물은 인공 구조물에 거의 정착하지 않는데, 아마도 오염에 취약하기 때문일 것이다. 이러한 특성 때문에 러시아의 일본해 연안에 건설 중인 석유 터미널 주변 환경 조건 측정에 ''Coptothyrus adamsi'' 개체군이 유용하게 사용될 수 있다.
완족동물은 미국의 켄터키주의 주 화석이다.[25]
모든 종이 바다의 저서생물이다. 많은 종은 자루의 끝을 저질에 부착시켜 몸을 고정하거나, 모래에 부착시켜 몸을 지탱하는 지점으로 삼는다. 자루가 없는 종은 단단한 저질에 몸을 직접 고정한다. 몸을 저질에 부착시키지 않는 종도 있다.
먹이를 얻기 위해 껍데기를 약간 열고, 촉수관의 섬모 운동에 의해 외투강 내에 물의 흐름을 만든다. 물속에 포함된 먹이 입자는 촉수 표면의 섬모에 의해 촉수 밑바닥에 있는 홈으로 들어가 입으로 운반된다. 주된 먹이는 식물플랑크톤이지만, 작은 유기물이라면 무엇이든 먹는다.
3. 1. 섭식
이끼벌레류와 포로니드류처럼, 완족동물도 촉수관(lophophore)을 가지고 있다. 촉수관은 섬모(미세한 털)를 가진 촉수의 관으로, 물의 흐름을 만들어 물에서 먹이 입자를 걸러 먹는 것을 가능하게 한다. 이끼벌레류나 포로니드류의 촉수관은 단일한, 안으로 끌어들일 수 있는 줄기에 달린 촉수의 고리 모양인 반면,[1] 완족류 촉수관의 기본 형태는 U자 모양이다.[2] 이는 문(phylum)의 이름의 유래가 된 팔(brachia, "arms")을 형성한다.[3] 완족류의 촉수관은 안으로 끌어들일 수 없으며, 껍데기가 열릴 때 벌어지는 가장 앞쪽 부분의 내부 공간의 최대 3분의 2를 차지한다. 이 제한된 공간에서 충분한 여과 능력을 제공하기 위해, 큰 완족류의 촉수관은 중간 정도에서 매우 복잡한 모양으로 접혀 있다. 고리와 나선 모양이 흔하며, 어떤 종의 촉수관은 손가락을 펼친 손과 비슷한 모양으로 뒤틀린다.[4] 모든 종에서 촉수관은 연골과 수정체골격(즉, 내부 체액의 압력)에 의해 지지되며,[5] 체액은 촉수로 뻗어 있다.[6] 일부 관절완족류는 팔껍데기(brachial valve) 안쪽에 부착된 촉수관을 지지하는 석회질 지지대인 브라키디움(brachidium)을 가지고 있다.[7] 이는 촉수관 근육의 극심한 감소와 일부 팔 신경의 감소로 이어졌다.[8]
촉수는 가장자리와 중앙을 따라 섬모(미세하게 움직이는 털)를 가지고 있다. 바깥쪽 섬모의 운동은 촉수 끝에서 기저부로 물의 흐름을 유도하고, 그곳에서 물이 빠져나간다. 촉수와 충돌하는 먹이 입자는 점액에 의해 포획되며, 중앙의 섬모는 이 혼합물을 촉수 기저부로 이동시킨다.[9] 촉수 기저부를 따라 팔홈(brachial groove)이 이어지며, 이 홈의 섬모는 먹이를 입 쪽으로 이동시킨다.[10] 완족류가 사용하는 방법은 "상류 수집(upstream collecting)"으로 알려져 있는데, 먹이 입자가 먹이 흐름을 만드는 섬모의 영역으로 들어올 때 포획된다. 이 방법은 관련된 포로니드류와 이끼벌레류, 그리고 익족류도 사용한다. 내장형동물류는 비슷하게 생긴 촉수관을 사용하지만, 이것은 단단하고 흐름은 기저부에서 끝으로 향하며, "하류 수집(downstream collecting)" 시스템을 형성하여 먹이 입자가 빠져나가려고 할 때 포획한다.[11]
물의 흐름은 열린 껍데기의 측면에서 팔(lophophore)로 들어가고 동물의 앞쪽에서 빠져나간다. 혀완족류(lingulid)에서는 입구와 출구 통로가 깔때기 역할을 하는 강모(chaetae) 무리에 의해 형성된다.[12] 다른 완족류에서는 입구와 출구 통로가 팔의 형태에 의해 구성된다.[13] 팔은 먹이 입자, 특히 식물 플랑크톤(작은 광합성 생물)을 포획하여 촉수 기저부를 따라 있는 팔홈(brachial groove)을 통해 입으로 전달한다.[14] 입은 팔 기저부에 있는 작은 구멍이다.[15] 먹이는 입, 근육질의 인두("throat"), 식도("gullet")를 통과한다.[16] 이들 모두는 섬모와 점액과 소화 효소를 분비하는 세포로 둘러싸여 있다.[17] 위 벽에는 먹이가 소화되는 가지가 있는 맹낭("pouches", 주머니)이 있다. 주로 세포 내에서 소화된다.[18]
영양분은 섬모에 의해 체강 전체, 맨틀엽을 포함하여 운반된다.[19] 대사에 의해 생성된 노폐물은 암모니아로 분해되며, 맨틀과 팔을 통해 확산에 의해 배출된다.[20] 완족류는 후신관을 가지고 있으며, 많은 문에서 암모니아 및 기타 용존 폐기물을 배출하는 데 사용된다. 그러나 완족류는 이 과정에서 배설의 첫 번째 단계를 수행하는 족세포의 징후가 없으며, 완족류의 후신관은 정자와 난자를 방출하는 데만 사용되는 것으로 보인다.[21][22]
완족류가 섭취하는 먹이의 대부분은 소화 가능하며, 고형 폐기물은 거의 생성되지 않는다.[23] 팔의 섬모는 소화되지 않은 물질의 개별 입자를 배출하기 위해 방향을 바꿀 수 있다. 동물이 원치 않는 더 큰 덩어리를 만나면, 입구 통로를 따라 있는 섬모는 일시적으로 멈추고, 덩어리와 접촉하는 촉수는 넓은 틈을 형성하기 위해 벌어지고, 그런 다음 섬모를 사용하여 덩어리를 맨틀 안쪽으로 천천히 버린다. 맨틀에는 자체 섬모가 있어서 껍데기 사이의 열림을 통해 덩어리를 씻어낸다. 팔이 막히면, 폐각근이 껍데기를 급격히 닫아 막힘을 제거하는 "재채기"를 일으킨다.[24] 일부 무관절 완족류에서는 소화관이 U자형이며, 몸벽 앞쪽에서 고형물을 배출하는 항문으로 끝난다.[25] 다른 무관절 완족류와 모든 관절 완족류는 막힌 끝을 가진 굽은 창자를 가지고 있으며 항문이 없다.[26] 이러한 동물들은 고형 폐기물을 점액과 함께 묶어서 주기적으로 창자 근육의 급격한 수축을 사용하여 "재채기"로 배출한다.[27]
완족동물은 껍데기를 약간 열고 촉수관의 섬모 운동으로 외투강 내에 물의 흐름을 만든다. 물속 먹이 입자는 촉수 표면의 섬모에 의해 촉수 밑바닥 홈으로 들어가 입으로 운반된다. 주된 먹이는 식물플랑크톤이지만, 작은 유기물이면 무엇이든 먹는다.
3. 2. 번식
대부분 종의 성체는 일생 동안 단일 성을 유지한다. 생식소는 발달 중인 생식세포(난자 또는 정자)의 덩어리이며, 대부분의 종은 각 껍데기에 두 개씩 총 네 개의 생식소를 가지고 있다.[1] 관절완족류의 생식소는 외투막 엽의 관에 위치하고, 무관절완족류의 생식소는 내장 근처에 위치한다.[2] 성숙한 생식세포는 주 체강으로 이동한 다음 후신관을 통해 외투강으로 배출되는데, 후신관은 입의 양쪽에 열려 있다. 대부분의 종은 난자와 정자를 모두 물속에 방출하지만, 일부 종의 암컷은 유생이 부화할 때까지 배를 육아낭에 보관한다.[1]세포분열은 방사형(세포는 서로 바로 위에 고리 모양으로 쌓여 형성됨), 전할구형(세포는 분리되어 있지만 인접해 있음), 조절형(세포가 발달하는 조직의 유형은 각 세포 내부가 아닌 인접 세포 간의 상호 작용에 의해 제어됨)이다.[1][3] 완족류의 원구는 닫히고 입과 항문은 새로운 개구부에서 발달하는데, 이는 일부 동물이 초기 배의 표면에 있는 "오목한 부분"인 원구를 깊게 하여 입과 항문을 발달시키는 것과 다르다.[1]
두판완족류(Lingulida와 Discinida)의 유생은 플랑크톤영양성(섭식성)이며, 몇 달 동안 플랑크톤으로 헤엄친다.[4] 이들은 소형 성체와 비슷하며, 껍데기, 외투막 엽, 외투강에서 나선형으로 감긴 자루, 그리고 먹이 섭취와 헤엄치는 데 사용되는 작은 촉수관을 가지고 있다.[1] 반면, 두건완족류 유생은 자루나 껍데기가 없다.[2] 껍데기가 무거워지면 어린 개체는 바닥으로 가라앉아 고착성 성체가 된다.[1] 관절완족류(Craniiformea와 Rhynchonelliformea) 유생은 난황영양성(비섭식성)이며, 난황만을 먹고 살며, 며칠 동안만 플랑크톤으로 남아있다. Rhynchonelliformea 유생은 Craniiformea가 두 개의 유생 엽만 가지고 있는 것과 달리 세 개의 유생 엽을 가지고 있다.[15] 이 유형의 유생은 몸과 촉수관이 되는 섬모가 있는 가장 앞쪽 엽, 자루가 되는 뒤쪽 엽, 그리고 뒤쪽 가장자리가 있는 치마와 같은 외투막을 가지고 있다. 변태 과정에서 자루가 표면에 부착되고, 앞쪽 엽은 촉수관과 다른 기관을 발달시키며, 외투막은 앞쪽 엽 위로 말려 올라가 껍데기를 분비하기 시작한다.[1] 차가운 바다에서는 완족류의 성장이 계절적이며, 겨울에는 종종 체중이 줄어든다. 이러한 성장 변화는 종종 껍데기에 성장선을 형성한다. 일부 속의 구성원은 수족관에서 먹이 없이 1년 동안 생존했다.[4]
완족동물은 유성생식만으로 번식하며, 무성생식은 전혀 알려져 있지 않다. 극소수의 자웅동체가 알려져 있지만, 대부분의 종은 자웅이체이다. 자웅이체인 경우에도, 성적이형은 거의 없다. 체외수정으로, 난자와 정자는 신관을 통해 해수 중에 방출되어 수정되는 것이 일반적이다. 일부 종에서는 난자는 암컷의 신관이나 외투강, 껍질의 오목한 곳 등에 머물러, 그곳에서 수정이 일어난다. 그 경우에는 수정란은 유생이 될 때까지 수정한 장소에서 보호된다.
수정란은 방사형의 전할을 거쳐 발생한다. 원구는 발생 과정에서 닫히고, 입은 이차적으로 생긴다. 모든 종이 부유하는 유생 시기를 가지지만, 유생의 특징은 관절완족류와 무관절완족류에서 크게 다르다. 무관절완족류의 유생은 촉수관과 두 개의 껍질, 육경을 갖추고, 성체와 그다지 다르지 않은 구조를 가진다. 성체와 다른 점은 육경이 구부러져 외투강 내에 있는 것과, 외투막에 비해 몸과 촉수관이 불균형적으로 크다는 것이다. 큰 촉수관은 이동과 섭식에 사용된다. 이 유생은 큰 변태를 거치지 않고 성체가 된다. 한편, 관절완족류의 유생에는 껍질이 없고, 몸은 두엽, 외투엽, 경엽의 세 부분으로 나뉜다. 스스로 먹이를 취할 수 없고, 난황의 영양에만 의존하며, 부유 기간은 짧다. 경엽을 사용하여 부착하면, 변태하여 성체가 된다.
4. 분류
완족동물은 크게 무관절강과 유관절강의 두 강(綱)으로 분류되었으나, 현재는 설각아문, 두각아문, 부리각아문의 세 아문(亞門)으로 분류하는 체계가 널리 쓰인다. 이 체계에서 부리각아문은 전통적인 분류의 유관절강에 해당하며, 무관절강은 키틴질 인산칼슘 껍데기를 가진 설각아문과 탄산칼슘 껍데기를 가진 두각아문으로 나뉜다.
완족동물 화석은 패각과 촉수관 형태가 매우 다양하지만, 현생 속들은 다양성이 적고 연체 특징을 제공한다. 이러한 형태적 특징과 더불어 수렴 진화와 역전 현상 때문에, 일부 분류학자들은 목(目)과 같은 상위 수준 분류를 정의하기에는 이르다고 보며, 속(屬)을 먼저 식별하고 이들을 중간 그룹으로 묶는 상향식 접근 방식을 선호한다.
반면, 다른 분류학자들은 일부 특징 패턴이 상위 수준 분류를 할 만큼 안정적이라고 믿지만, 상위 수준 분류 방식에 대해서는 의견이 다르다. 1869년에 정의된 "전통적인" 분류 외에, 1990년대에 패각 구성 물질을 기반으로 한 분류와 3부 구성 방식이 확립되었다.
- 전통적인 분류: 유관절강(밸브 사이에 이빨 있는 경첩)과 무관절강(근육으로만 연결된 경첩)으로 나뉜다.
- 패각 구성 물질 기반 분류: 방해석 패각을 가진 두개류와 유관절류를 석회질류로, 키틴과 인산칼슘 패각을 가진 설체류와 원반류를 설체류로 통합한다.
- 3부 구성 방식: 두개류를 두개형류로, 설체류와 원반류를 설체형류로, 린코넬리다와 테레브라툴리다를 린코넬리형류로 그룹화한다.
| "전통적인" 분류 | 무극류 | 유극류 | |||
|---|---|---|---|---|---|
| "석회질류" 접근 방식 | 설체류 | 석회질류 | |||
| 3부 구성 접근 방식 | 설체형류 | 두개형류 | 린코넬리형류 | ||
| 목 | 설체류 | 원반류 | 두개류 | 테레브라툴리다 | 린코넬리다 |
| 경첩 | 없음 | 이빨과 홈 | |||
| 항문 | 몸 앞쪽, U자형 창자 끝에 위치 | 없음 | 없음 | ||
| 자루 | 체강 포함, 근육이 관통 | 자루 없음 | 체강 없음, 몸과 연결되는 부위에 근육 존재 | ||
| 길고, 굴을 팜 | 짧고, 단단한 표면에 부착 | 없음, 표면에 고착 | 짧고, 단단한 표면에 부착 | ||
| 각피 | 글리코사미노글리칸과 키틴 | 키틴 | 단백질 | ||
| 패각의 주요(중간) 광물질 층 | 글리코사미노글리칸과 아파타이트 (인산칼슘) | 방해석 (탄산칼슘의 한 형태) | |||
| 패각의 내부 광물질 층 | 콜라겐 및 기타 단백질, 키틴인산염 및 아파타이트 (인산칼슘) | 방해석 | 단백질과 방해석 | ||
| 패각 개구부 주변의 강모 | 있음 | 있음 | 있음 | ||
| 체강 완전히 분할됨 | 있음 | 없음 | 있음 | ||
| 유생 | 플랑크토영양성(섭식) | 난황영양성(비섭식) | |||

4. 1. 전통적 분류
1869년에 정의된 "전통적인" 분류에서는 완족동물을 유극류(Articulata)와 무극류(Inarticulata)로 나눈다. 유극류는 밸브 사이에 이빨이 있는 경첩을 가지는 반면, 무극류는 근육만으로 연결된 경첩을 가진다.[16]| "전통적인" 분류 | 무극류 | 유극류 | |||
|---|---|---|---|---|---|
| "석회질류" 접근 방식 | 설체류 | 석회질류 | |||
| 3부 구성 접근 방식 | 설체형류 | 두개형류 | 린코넬리형류 | ||
| 목 | 설체류 | 원반류 | 두개류 | 테레브라툴리다 | 린코넬리다 |
| 경첩 | 없음 | 이빨과 홈 | |||
| 항문 | 몸 앞쪽, U자형 창자 끝에 위치 | 없음 | 없음 | ||
| 자루 | 체강 포함, 근육이 관통 | 자루 없음 | 체강 없음, 몸과 연결되는 부위에 근육 존재 | ||
| 길고, 굴을 팜 | 짧고, 단단한 표면에 부착 | 없음, 표면에 고착 | 짧고, 단단한 표면에 부착 | ||
| 각피 | 글리코사미노글리칸과 키틴 | 키틴 | 단백질 | ||
| 패각의 주요(중간) 광물질 층 | 글리코사미노글리칸과 아파타이트 (인산칼슘) | 방해석 (탄산칼슘의 한 형태) | |||
| 패각의 내부 광물질 층 | 콜라겐 및 기타 단백질, 키틴인산염 및 아파타이트 (인산칼슘) | 방해석 | 단백질과 방해석 | ||
| 패각 개구부 주변의 강모 | 있음 | 있음[17] | 있음 | ||
| 체강 완전히 분할됨 | 있음 | 없음 | 있음 | ||
| 유생 | 플랑크토영양성(섭식) | 난황영양성(비섭식) | |||
전통적으로 완족동물은 무관절강과 유관절강의 두 강(綱)으로 크게 나뉘어 왔지만, 설각아문, 두각아문, 부리각아문의 세 아문(亞門)으로 분류하는 체계가 널리 사용된다. 이 체계에서는 부리각아문이 전통적인 분류의 관절이 있는 강에 해당한다. 관절이 없는 강은 키틴질의 인산칼슘 껍데기를 가진 설각아문과 탄산칼슘 껍데기를 가진 두각아문으로 나뉜다.
다음은 전통적인 체계의 한 예이다. 멸종군에는 †를 붙였다.
- 무관절강
- * 삿갓조개목(린굴라목)
- * 이카리초우친목(정각류)
- * †오볼렐라목
- * †파테리나목
- 관절강
- * †오르티스목
- * †스트로포메나목
- * †펜타멜루스목
- * 부리조개목(린코넬라목)
4. 2. 현대적 분류
완족류 화석은 패각과 촉수관의 형태학적 다양성이 매우 크지만, 현생 속들은 다양성이 덜하고 연체(soft-body) 특징을 보인다. 화석과 현존하는 종 모두 형태학에 기반한 완족류의 포괄적인 분류를 어렵게 만드는 한계점을 가지고 있다. 또한 이 문은 상당한 수렴 진화와 역전(최근 그룹이 중간 그룹에서 보이는 특징을 잃고, 더 오래된 그룹에서 마지막으로 보였던 특징으로 되돌아가는 것)을 경험했다. 따라서 일부 완족류 분류학자들은 목과 같은 상위 수준의 분류를 정의하기에는 시기상조라고 믿고, 대신 속을 식별한 다음 이들을 중간 그룹으로 묶는 상향식 접근 방식을 권장한다.그러나 다른 분류학자들은 일부 특징 패턴이 상위 수준의 분류를 할 만큼 충분히 안정적이라고 믿고 있지만, 상위 수준의 분류가 무엇이어야 하는지에 대한 견해는 다르다. "전통적인" 분류는 1869년에 정의되었고, 두 가지 추가적인 접근 방식이 1990년대에 확립되었다.
- "전통적인" 분류에서 완족류는 유극류(Articulata)와 무극류(Inarticulata)로 나뉜다. 유극류는 밸브 사이에 이빨이 있는 경첩이 있는 반면, 무극류의 경첩은 근육에 의해서만 연결되어 있다.
- 1990년대에 패각의 구성 물질을 기반으로 고안된 분류는 두개류와 "유극류" 완족류를 석회질류에 통합했는데, 이들은 방해석 패각을 가지고 있다. 설체류와 원반류는 설체류에 통합되었는데, 이들은 키틴과 인산칼슘으로 만들어진 패각을 가지고 있다.
- 1990년대의 3부 구성 방식은 두개류를 자체 별도 그룹인 두개형류에 배치한다.[16] 설체류와 원반류는 설체형류로,[16] 린코넬리다와 테레브라툴리다는 린코넬리형류로 그룹화된다.
| "전통적인" 분류 | 무극류 | 유극류 | |||
|---|---|---|---|---|---|
| "석회질류" 접근 방식 | 설체류 | 석회질류 | |||
| 3부 구성 접근 방식 | 설체형류 | 두개형류 | 린코넬리형류 | ||
| 목 | 설체류 | 원반류 | 두개류 | 테레브라툴리다 | 린코넬리다 |
| 경첩 | 없음 | 이빨과 홈 | |||
| 항문 | 몸 앞쪽, U자형 창자 끝에 위치 | 없음 | 없음 | ||
| 자루 | 체강 포함, 근육이 관통 | 자루 없음 | 체강 없음, 몸과 연결되는 부위에 근육 존재 | ||
| 길고, 굴을 팜 | 짧고, 단단한 표면에 부착 | 없음, 표면에 고착 | 짧고, 단단한 표면에 부착 | ||
| 각피 | 글리코사미노글리칸과 키틴 | 키틴 | 단백질 | ||
| 패각의 주요(중간) 광물질 층 | 글리코사미노글리칸과 아파타이트 (인산칼슘) | 방해석 (탄산칼슘의 한 형태) | |||
| 패각의 내부 광물질 층 | 콜라겐 및 기타 단백질, 키틴인산염 및 아파타이트 (인산칼슘) | 방해석 | 단백질과 방해석 | ||
| 패각 개구부 주변의 강모 | 있음 | 있음[17] | 있음 | ||
| 체강 완전히 분할됨 | 있음 | 없음 | 있음 | ||
| 유생 | 플랑크토영양성(섭식) | 난황영양성(비섭식) | |||
약 330종의 현존하는 종이 인식되며, 100개 이상의 속으로 그룹화된다. 현대 완족류의 대부분은 린코넬리형류(유극류)이다.
1990년대 이후 수행된 유전자 분석은 서로 다른 생물 간의 관계에 대한 이해를 넓혔다. 이제 완족류는 이전에 가설적으로 제기되었던 것처럼 후구동물(예: 극피동물과 척삭동물)에 속하지 않고, 광범위한 그룹 원구동물의 하위 그룹인 좌우대칭동물에 포함되어야 한다는 것이 명확해졌다. 성체 형태는 다소 다르게 보이지만, 18S rRNA의 뉴클레오타이드 서열은 포로니드류(말굽벌레)가 관절이 없는 완족류와 관절 있는 완족류보다 더 가까운 근연종임을 나타낸다. 현재로서는 관절 없는 완족류 내부의 정확한 관계에 대한 증거의 무게는 불확실하다. 따라서 말굽벌레를 완족류에 포로나타(Phoronata)라는 강으로 포함시키는 것이 제안되었는데, 이는 Linguliformea 아문 내에 있는 Craniata와 Lingulata에 추가되는 것이다. 다른 아문인 Rhynchonelliformea는 현존하는 강 하나만 포함하며, 현존하는 목인 Rhynchonellida, Terebratulida, Thecideida로 세분된다.[18][19]
이는 멸종 그룹을 포함한 완족동물의 분류 체계를 목 수준까지 보여준다. 멸종 그룹은 (†) 기호로 표시된다.
- '''아문 린굴리포르메아'''
- * '''강 린굴라타'''
- ** 목 린굴리다
- ** 목 †아크로트레티다
- ** 목 †사이포노트레티다
- * '''강 †파테리나타'''
- ** 목 †파테리니다
- '''아문 크라니이포르메아'''
- * '''강 크라니아타'''
- ** 목 크라니이다
- ** 목 †크라니옵시다
- ** 목 †트리메렐리다
- '''아문 린코넬리포르메아'''
- * '''강 †칠레아타'''
- ** 목 †칠레이다
- ** 목 †딕티오넬리다
- * '''강 †오볼레라타'''
- ** 목 †오볼레리다
- ** 목 †나우카티다
- * '''강 †쿠토르기나타'''
- ** 목 †쿠토르기니다
- * '''강 †스트로포메나타'''
- ** 목 †빌링셀리다
- ** 목 †스트로포메니다
- ** 목 †프로덕티다
- ** 목 †오르토테티다
- * '''강 린코넬라타'''
- ** 목 린코넬리다
- ** 목 테레브라툴리다
- ** 목 테시데이다
- ** 목 †프로토르티다
- ** 목 †오르티다
- ** 목 †펜타메리다
- ** 목 †아트리피다
- ** 목 †아시리디다
- ** 목 †스피리페리다
- ** 목 †스피리페리니다
전통적으로는 관절이 있는 강과 관절이 없는 강의 두 강으로 크게 나뉘어 왔지만, 세 개의 아문(설각아문, 두각아문, 부리각아문)으로 분류하는 체계가 널리 사용되고 있다. 이 체계에서는 부리각아문이 전통적인 분류의 관절이 있는 강에 해당한다. 관절이 없는 강은 키틴질의 인산칼슘 껍데기를 가진 설각아문과 탄산칼슘 껍데기를 가진 두각아문으로 나뉜다.
그 외에, 껍데기의 성분을 중시하여, 탄산칼슘 껍데기를 가진 두각류와 부리각류를 묶고, 인산칼슘 껍데기를 가진 설각류와 나란히 배열하는 체계도 제안되고 있다. 아래에는 경첩의 유무에 따른 전통적인 두 강의 체계와 세 아문의 체계를 제시한다.
다음은 전통적인 체계의 한 예이다. 같은 두 강으로 나누는 체계라도 하위 분류에 대해서는 다른 것도 있다. 멸종군에는 †를 붙였다.
- 무관절강
- * 삿갓조개목(설각류, 린굴라목)
- * 이카리초우친목(정각류)
- * †오볼렐라목
- * †파테리나목
- 관절강
- * †오르티스목(오르시스목)
- * †스트로포메나목
- * †펜타멜루스목
- * 부리조개목(부리각류, 린코넬라목)
다음은 세 아문으로 분류하는 체계이다.
- 두각아문
- * 두각강
- ** 두각목(크라니아목, 정각목, 이카리초우친목)
- ** †목
- ** †목
- 설각아문
- * 설각강
- ** †아크로트레타목
- ** 삿갓조개목(린굴라목)
- ** †목
- * †파테리나강
- ** †파테리나목
- 부리각아문
- * †강
- ** †목
- ** †목
- * †강
- ** †목
- * †오볼렐라강
- ** †
- ** †오볼렐라목
- * 부리각강
- ** †목
- ** †아트리파목
- ** †오르티스목(오르시스목)
- ** †펜타멜루스목
- ** †목
- ** 부리조개목(부리각목, 린코넬라목)
- ** †스피리퍼목
- ** †목
- ** 호주키조개목(천각목)
- ** 테키데라목
- * †스트로포메나강
- ** †목
- ** †목
- ** †프로덕투스목
- ** †스트로포메나목
5. 진화


완족동물의 화석은 현생대의 모든 시대 지층에서 발견된다. 가장 오래된 것은 고생대 캄브리아기 초기 지층에서 발견되었으며, 처음에는 무절류만 발견되었지만 캄브리아기 중기가 되면 유절류도 등장한다. 유절류가 등장한 이후 무절류의 수는 감소하여 데본기 이후로는 거의 발견되지 않는다. 오르도비스기에는 유절류가 다양화되고, 데본기에 그 다양성이 최대가 된다. 그러나 고생대 말(P-T 경계)의 대량 멸종으로 완족동물의 다양성은 대부분 상실되었고, 그 후 완족동물은 과거처럼 번성하지 못하고 쇠퇴하는 경향에 있다. 완족동물의 화석종은 1만 3000종이 기재되어 있는 반면, 현생종은 350종 정도에 그친다.
현생 미역삿갓조개를 포함한 삿갓조개속은 살아있는 화석(오래전부터 형태를 크게 바꾸지 않고 살아온 생물)의 대표적인 예로 잘 알려져 있다. 현생종과 유사한 화석은 캄브리아기에서도 발견된다.
1990년대 이후 유전자 분석을 통해 생물 간의 관계에 대한 이해가 넓어졌다. 완족류는 후구동물(극피동물, 척삭동물 등)이 아닌, 원구동물의 하위 그룹인 좌우대칭동물에 포함된다는 것이 명확해졌다. 18S rRNA의 뉴클레오타이드 서열은 포로니드류(말굽벌레)가 관절이 없는 완족류와 관절 있는 완족류보다 더 가까운 근연종임을 나타낸다.[18][19]
현재 관절 없는 완족류 내부의 정확한 관계는 불확실하다. 포로나타(Phoronata)라는 강으로 말굽벌레를 완족류에 포함시키자는 제안이 있는데(B.L.Cohen & Weydmann), 이는 Linguliformea 아문 내에 있는 Craniata와 Lingulata에 추가된다. 다른 아문인 Rhynchonelliformea는 현존하는 강 하나만 포함하며, Rhynchonellida, Terebratulida, Thecideida 목으로 세분된다.[18][19]
완족동물은 포로니드(箒虫動物)문, 태형동물(外肛動物)문과 촉수관 등 여러 특징을 공유하기 때문에, 이 두 분류군과 함께 촉수동물(觸手動物) 또는 촉수관동물로 묶여 왔다. 촉수관동물은 구구동물적인 특징도 있지만, 방사형의 난할, 성체의 입이 원구에서 유래하지 않는 점, 몸의 3체절 구조 등 신구동물의 특징을 가지므로, 전통적으로는 신구동물의 한 분류군으로 여겨져 왔다. 그러나 분자계통학 결과는 촉수관동물을 구구동물에 포함하는 것을 지지하는 경우가 많아, 이것이 유력해지고 있다.
구구동물 중에서, 촉수관동물은 윤형동물(연체동물 등, 트로코포어 유생을 가진 분류군)과 함께 관륜동물로 묶는 견해가 지지되고 있다. 그러나 관륜동물 내에서 완족동물의 위치는 명확하지 않으며, 촉수관동물이 단계통군을 이룬다는 전통적인 견해도 의심받고 있다.
5. 1. 화석 기록
{| class="wikitable"|-
! 주요 화석 완족류 그룹의 연대표
|-
|
| 대 | 고생대 | 중생대 | 신생대 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 기 | 캄 | 오 | 실 | 데 | 석 | 페 | 트 | 쥐 | 백 | 팔 | 신생대 |
| 린굴라타 | colspan="11" | | ||||||||||
| 오보렐리다 | colspan="1"| | ||||||||||
| 스트로포메니다 | colspan="4" | | ||||||||||
| 오르티다 | colspan="5" | | ||||||||||
| 펜타메리다 | colspan="3" | | ||||||||||
| 링코넬리다 | colspan="9" | | ||||||||||
| 스피리페리다 | colspan="4" | | ||||||||||
| 테레브라툴리다 | colspan="8" | | ||||||||||
|}
12,000종이 넘는 화석 종들이 확인되었으며, 5,000개가 넘는 속으로 분류된다. 현존하는 완족동물 중 가장 큰 것은 길이가 100mm 정도이지만, 일부 화석은 최대 200mm의 너비를 가진다. 가장 초기의 완족동물은 초기 캄브리아기에서 발견되었는데, 무관절형이 먼저 나타난 후 관절형이 이어서 나타났다. 캄브리아기에서 세 종의 무광물화 종도 발견되었는데, 이들은 광물화된 조상으로부터 진화한 두 개의 뚜렷한 그룹을 나타내는 것으로 보인다. 무관절형인 ''Lingula''는 종종 "살아있는 화석"이라고 불리는데, 매우 유사한 속이 오르도비스기까지 거슬러 올라가 발견되기 때문이다. 반면에, 관절형 완족동물은 주요한 다양화를 일으켰고, 심각한 대량 멸종을 겪었지만, 가장 다양한 현생 그룹인 관절형 Rhynchonellida와 Terebratulida는 각각 오르도비스기와 석탄기 초기에 나타났다.
1991년 이후 클라우스 니엘센(Claus Nielsen)은 완족동물의 발달에 대한 가설을 제안했고, 2003년 코헨(Cohen)과 동료들은 이를 완족동물의 초기 진화에 대한 가설로 적용했다. 이 "완족동물 접힘" 가설은 완족동물이 ''Halkieria''와 유사한 조상으로부터 진화했다고 제안하는데, ''Halkieria''는 등에 "사슬 갑옷"과 앞뒤 끝에 껍데기를 가진 민달팽이와 같은 동물이다. 이 가설은 최초의 완족동물이 몸의 뒤쪽 부분을 앞쪽 아래로 접어서 껍데기를 두 개의 밸브로 변형시켰다고 제안한다.
그러나 2007년 이후의 화석들은 초기 캄브리아기 톰모티아류에 대한 새로운 해석과 완족동물이 톰모티아류로부터 진화했다는 새로운 가설을 뒷받침한다. 톰모티아류의 "갑옷"은 잘 알려져 있었지만 조립된 형태는 아니었고, 일반적으로 톰모티아류는 ''Halkieria''와 유사한 민달팽이 모양의 동물이라고 가정했는데, 다만 톰모티아류의 갑옷은 유기인산염 화합물로 만들어진 반면 ''Halkieria''의 갑옷은 방해석으로 만들어졌다는 점이 다르다. 그러나 새로운 톰모티아류인 ''Eccentrotheca''의 화석은 조립된 갑옷이 관을 형성하고 있음을 보여주는데, 이는 기어 다니는 민달팽이 모양의 동물이 아니라 고착성 동물임을 나타낸다. ''Eccentrotheca''의 유기인산염 관은 포로니드의 관과 유사하며, 포로니드는 촉수관으로 먹이를 먹는 고착성 동물로 완족동물과 매우 가까운 친척 또는 완족동물의 아그룹으로 간주된다.[26] 2008년에 발견되어 2009년에 설명된 또 다른 대부분 조립된 화석인 ''Paterimitra''는 완족동물의 밸브와 같지만 동물의 몸을 완전히 둘러싸고 있지는 않은 바닥에 두 개의 대칭적인 판을 가지고 있다.[27]
고생대에서 최고점에 달했을 때,[28] 완족동물은 가장 풍부한 여과 섭식자이자 암초 건설자 중 하나였으며, 조개의 제트 추진 방식으로 헤엄치는 것을 포함한 다른 생태적 지위를 차지했다. 그러나 "대멸종"으로 알려진 페름기-트라이아스기 멸종 사건 이후, 완족동물은 이전 다양성의 3분의 1만 회복했다. 완족동물의 다양성이 실제로 감소하고 있고 어떤 방식으로든 이매패류가 완족동물을 능가했다고 종종 생각되었다. 그러나 1980년에 굴드(Stephen Jay Gould)와 캘러웨이(Calloway)는 완족동물과 이매패류 모두 고생대부터 현대까지 증가했지만 이매패류가 더 빠르게 증가했다는 결론을 내린 통계 분석을 제시했다. 페름기-트라이아스기 멸종은 이매패류에게는 중간 정도로 심각했지만 완족동물에게는 파괴적이어서 완족동물이 처음으로 이매패류보다 다양성이 낮아졌고 페름기 이후 완족동물의 다양성은 매우 낮은 기준에서 증가했다. 이매패류가 완족동물을 능가했다는 증거는 없으며, 두 그룹 모두 단기간의 증가 또는 감소가 동시에 나타났다. 2007년 놀(Knoll)과 밤바흐(Bambach)는 완족동물이 페름기-트라이아스기 멸종에 가장 취약한 여러 그룹 중 하나였으며, 모두 석회질 경질부(탄산칼슘으로 구성됨)를 가지고 있으며 대사율이 낮고 호흡계가 약하기 때문이라고 결론지었다.
완족동물 화석은 고생대의 기후 변화를 나타내는 유용한 지표가 되었다. 오르도비스기의 대부분과 같이 지구 온도가 낮았을 때 적도와 극 사이의 큰 온도 차이로 인해 서로 다른 위도에서 다른 화석 집단이 생성되었다. 반면에 실루리아기의 대부분과 같이 따뜻한 시기에는 온도 차이가 작아져 저위도에서 중위도까지 모든 바다가 같은 소수의 완족동물 종에 의해 서식되었다.
완족동물의 화석은 현생대의 모든 시대 지층에서 발견된다. 가장 오래된 것은 고생대 캄브리아기 초기 지층에서 발견되었다. 처음에는 무절류만 발견되었지만, 캄브리아기 중기가 되면 유절류도 등장한다. 유절류가 등장한 이후 무절류의 수는 감소하여 데본기 이후로는 거의 발견되지 않는다. 오르도비스기에는 유절류가 다양화되고, 데본기에 그 다양성이 최대가 된다. 그러나 고생대 말(P-T 경계)의 대량 멸종으로 완족동물의 다양성은 대부분 상실되었다. 그 후 완족동물은 과거처럼 번성하지 못하고 쇠퇴하는 경향에 있다. 완족동물의 화석종은 1만 3000종이 기재되어 있는 반면, 현생종은 350종 정도에 그친다. 완족동물의 다양성 감소는 생태가 유사한 이매패류와의 경쟁에서 패배했기 때문으로 생각된다.
현생 미역삿갓조개를 포함한 삿갓조개속은 살아있는 화석(오래전부터 형태를 크게 바꾸지 않고 살아온 생물)의 대표적인 예로 잘 알려져 있다. 이와 같은 속의 화석(이견이 있지만, 어느 쪽이든 매우 유사한 형태를 가진 화석)이 오르도비스기・실루리아기부터 발견되기 때문이다. 같은 속으로 분류되지는 않지만, 현생종과 유사한 화석은 더 거슬러 올라가 캄브리아기에서도 발견된다.
5. 2. 진화 가설
1991년 이후 클라우스 니엘센(Claus Nielsen)은 완족동물의 발달에 대한 가설을 제안했고, 2003년 코헨(Cohen)과 동료들은 이를 완족동물의 초기 진화에 대한 가설로 적용했다. "완족동물 접힘" 가설은 완족동물이 Halkieria[25]와 유사한 조상으로부터 진화했다고 제안하는데, ''Halkieria''는 등에 "사슬 갑옷"과 앞뒤 끝에 껍데기를 가진 민달팽이와 같은 동물이다.[29] 이 가설은 최초의 완족동물이 몸의 뒤쪽 부분을 앞쪽 아래로 접어서 껍데기를 두 개의 밸브로 변형시켰다고 제안한다.[25]그러나 2007년 이후의 화석들은 초기 캄브리아기 톰모티아류에 대한 새로운 해석과 완족동물이 톰모티아류로부터 진화했다는 새로운 가설을 뒷받침한다. 톰모티아류의 "갑옷"은 잘 알려져 있었지만 조립된 형태는 아니었고, 일반적으로 톰모티아류는 ''Halkieria''와 유사한 민달팽이 모양의 동물이라고 가정했는데, 다만 톰모티아류의 갑옷은 유기인산염 화합물로 만들어진 반면 ''Halkieria''의 갑옷은 방해석으로 만들어졌다는 점이 다르다. 그러나 새로운 톰모티아류인 Eccentrotheca의 화석은 조립된 갑옷이 관을 형성하고 있음을 보여주는데, 이는 기어 다니는 민달팽이 모양의 동물이 아니라 고착성 동물임을 나타낸다. ''Eccentrotheca''의 유기인산염 관은 포로니드의 관과 유사하며, 포로니드는 촉수관으로 먹이를 먹는 고착성 동물로 완족동물과 매우 가까운 친척 또는 완족동물의 아그룹으로 간주된다.[26] 2008년에 발견되어 2009년에 설명된 또 다른 대부분 조립된 화석인 Paterimitra는 완족동물의 밸브와 같지만 동물의 몸을 완전히 둘러싸고 있지는 않은 바닥에 두 개의 대칭적인 판을 가지고 있다.[27]
그린란드의 시리우스 파셋(Sirius Passet)에서 발견된 할키에리아(Halkieria) 화석은 몸의 양쪽 끝 등쪽에 껍데기를 가지고 있었다. 사이먼 콘웨이 모리스(Simon Conway Morris)는 이 껍데기가 완족동물의 것과 유사하다는 점을 지적하며, 완족동물이 할키에리아와 같은 동물에서 진화했다고 추측했다. 그의 생각에 따르면, 양쪽 끝에 껍데기를 가진 할키에리아의 몸이 접히는 듯이 변화하여 두 개의 껍데기가 마주하게 됨으로써 완족동물의 체제가 기원했다는 것이다. 그는 또한 완족동물이 가진 강모가 할키에리아가 가진 경피에서 유래한다고 생각할 수 있다는 점과, 일부 현생 완족동물에서는 유생이 변태할 때 몸이 접히게 된다는 점을 근거로 제시하고 있다. 할키에리아의 경피는 환형동물의 강모에서 유래한다고 생각할 수도 있으므로, 이 가설은 완족동물과 환형동물이 근연이라는 가설과 일치한다. 한편, 할키에리아를 연체동물에 가깝다고 생각하는 의견도 있다.
그런데, 콘웨이 모리스의 주장이 옳다면, 완족동물의 껍데기는 모두 등쪽에 있다(배쪽은 안쪽으로 접힌다)는 것이 되며, 두 개의 껍데기가 배등에 있다는 전통적인 생각은 옳지 않게 된다.
5. 3. 계통
1990년대 이후 유전자 분석을 통해 생물 간의 관계에 대한 이해가 넓어졌다. 완족류는 후구동물(극피동물, 척삭동물 등)이 아닌, 원구동물의 하위 그룹인 좌우대칭동물에 포함된다는 것이 명확해졌다. 18S rRNA의 뉴클레오타이드 서열은 포로니드류(말굽벌레)가 관절이 없는 완족류와 관절 있는 완족류보다 더 가까운 근연종임을 나타낸다.[18][19]현재 관절 없는 완족류 내부의 정확한 관계는 불확실하다. 포로나타(Phoronata)라는 강으로 말굽벌레를 완족류에 포함시키자는 제안이 있는데(B.L.Cohen & Weydmann), 이는 Linguliformea 아문 내에 있는 Craniata와 Lingulata에 추가된다. 다른 아문인 Rhynchonelliformea는 현존하는 강 하나만 포함하며, Rhynchonellida, Terebratulida, Thecideida 목으로 세분된다.[18][19]
1940년대부터 1990년대까지는 배아 발생 및 형태학적 특징을 기반으로 한 계통수가 완족류를 후구동물 또는 그 자매군으로 분류했다.[29] 여기에는 척삭동물과 극피동물이 포함된다. 그러나 완족류를 후구동물과 연관 짓는 근거에 대한 정밀한 조사에서 문제점이 발견되었다.[33]
- 난자의 초기 세포 분열에서 방사형 분열은 초기 후생동물, 초기 탈피동물, 그리고 아마도 후구동물의 주요 하위 그룹인 초기 촉수동물의 원래 상태로 보인다. 따라서 방사형 분열은 완족류가 후구동물과 관련이 있음을 의미하지 않는다.[33]
- 후구동물과 전구동물의 체강이 각각 장체강 형성과 균열체강 형성이라는 서로 다른 과정을 통해 형성된다는 것이 전통적인 견해였다.[33] 그러나 1990년대 초 이후의 연구는 상당한 예외를 발견했다. 두 가지 유형의 체강 형성 모두 완족류에서 나타나므로, 완족류가 후구동물임을 의미하지 않는다.[33]
- "후구동물"과 "전구동물"이라는 용어는 원래 배아 초기 단계에 나타나는 함입인 원구에서 입이 형성되는 방식을 구분하는 데 사용되었다. 그러나 일부 "전구동물"은 전형적인 후구동물과 유사한 과정을 통해 입을 형성한다. 따라서 후구동물과 유사한 과정을 통해 입이 형성된다고 해서 완족류가 후구동물과 관련되어 있음을 의미하지 않는다.[33]
닐슨은 완족류와 밀접한 관련이 있는 엽형동물을 후구동물인 날개깃동물과 연관 짓는 견해를 가지고 있다. 그 이유는 이들의 촉수관이 세포당 하나의 섬모에 의해 움직이는 반면, 그가 전구동물로 간주하는 태형동물의 촉수관은 세포당 여러 개의 섬모를 가지고 있기 때문이다. 그러나 날개깃동물은 반삭동물이며 아마도 극피동물과 밀접한 관련이 있고, 날개깃동물과 다른 반삭동물의 최근 공통 조상이나 반삭동물과 극피동물의 최근 공통 조상이 고착성이고 촉수를 이용하여 먹이를 섭취했다는 증거는 없다.[33]
1988년 이후, DNA 유사성과 같은 생화학적 특징을 비교하는 분자 계통 발생학에 기반한 분석은 완족류를 연체동물, 환형동물, 편형동물을 포함하지만 절지동물을 포함하지 않는 전구동물 상문인 촉수동물에 속하는 것으로 분류했다. 이 결론은 리보솜 DNA, Hox 유전자, 미토콘드리아 단백질 유전자, 단일 핵 단백질 유전자, 그리고 핵 단백질 유전자 집합체 등 광범위한 유전자를 사용하는 분자 계통 발생학 연구에서 일치한다.
2000년과 2001년의 일부 통합 연구는 완족류를 촉수동물로 지지하는 반면, 1998년과 2004년의 다른 연구는 완족류를 후구동물로 결론지었다.
포로니다류는 촉수관으로 먹이를 먹고, 표면에 굴을 파거나 부착하여, 다당류(아마도 키틴일 가능성이 있음)와 해저 물질 입자를 섞어 만든 3겹의 관을 만든다. 전통적으로는 별개의 문으로 간주되었지만, 1997년부터 2000년까지 점점 더 자세해진 분자 계통 발생 연구 결과, 포로니다류는 완족류의 아종임을 결론지었다. 그러나 2005년 분석에서는 포로니다류가 태형동물의 아종이라는 결론을 내렸다.
2008년까지 모든 분자 계통 발생 연구와 절반의 통합 연구 결과는 완족류가 후구동물임을 결론지었지만, 완족류와 가장 가까운 후구동물 문이 무엇인지 확인할 수 없었다. 포로니다류(완족류의 아종)를 제외하고는 말이다. 그러나 2008년 두 가지 분석 결과, 완족류와 가장 가까운 후구동물 친척은 유형동물이었다. 저자들은 유형동물은 초기 세포 분열 단계에서 나선형 분열을 하고 트로코포라 유생을 형성하지만, 완족류는 방사형 분열을 하고 트로코포라 유생에서 진화한 흔적이 없는 유생을 가지고 있기 때문에 이를 놀라운 결과로 여겼다. 2008년 또 다른 연구에서도 완족류가 유형동물과 밀접한 관련이 있다는 결론을 내렸으며, 이는 후구동물 내 촉수관으로 먹이를 먹는 동물들의 분지군 촉수동물에 완족류가 속한다는 생각에 의문을 제기했다.
완족동물은 촉수관 등 여러 특징을 포로니드(箒虫動物)문, 태형동물(外肛動物)문과 공유하기 때문에, 이 두 분류군과 함께 촉수동물(觸手動物) 또는 촉수관동물로 묶여 왔다. 촉수관동물은 구구동물적인 특징도 있지만, 방사형의 난할, 성체의 입이 원구에서 유래하지 않는 점, 몸의 3체절 구조 등 신구동물의 특징을 가지므로, 전통적으로는 신구동물의 한 분류군으로 여겨져 왔다. 그러나 분자계통학 결과는 촉수관동물을 구구동물에 포함하는 것을 지지하는 경우가 많아, 이것이 유력해지고 있다.
구구동물 중에서, 촉수관동물은 윤형동물(연체동물 등, 트로코포어 유생을 가진 분류군)과 함께 관륜동물로 묶는 견해가 지지되고 있다. 그러나 관륜동물 내에서 완족동물의 위치는 명확하지 않다. 촉수관동물이 단계통군을 이룬다는 전통적인 견해도 의심받고 있다.
캐벌리어-스미스는 촉수관동물 3분류군 중에서 태형동물만은 다른 계통적 위치에 있지만, 완족동물과 포로니드는 단계통군을 이룬다고 생각하여, 이 두 분류군을 아문으로 포함하는 문을 제창했다. 헬름캄프 등의 연구는 완동물의 단계통성을 지지하며, 더 나아가 유형동물(紐形動物)이 그 자매군이라고 주장했다. 또한 코헨은 포로니드는 완족동물문에 포함된다고 말하고 있다. 한편, 던 등은 완족동물, 포로니드, 유형동물 3분류군이 단계통군이 된다는 점에서는 같은 결론에 도달했지만, 완족동물과 가장 가까운 근연종은 포로니드가 아니라 유형동물이라고 추정했다. 원도는 미토콘드리아 게놈상의 유전자 순서에 기반하여, 완족동물은 오히려 환형동물이나 유형동물에 가깝다고 추정하고, 포로니드와의 근연성에 의문을 제기하고 있다.
6. 인간과의 관계
인간과의 관계는 드문 편이다. 일본이나 동남아시아에서는 녹색바다민들레(ミドリシャミセンガイ) 등의 바다민들레속(シャミセンガイ属)이 식용으로 이용된다.
7. 한국 분포
참조
[1]
논문
Braquiópodos discínidos (Lingulida, Discinoidea) de la Formación Ixtaltepec, Carbonífero del área de Santiago Ixtaltepec, Oaxaca.
[2]
논문
Socioeconomic study and economic value of living fossil, Lingula sp. in mangrove ecosystem in Trat Province, Thailand
[3]
논문
Rediscovery of Terebratulina austroamericana Zezina, 1981 (Brachiopoda: Cancellothyrididae) from off northern Chile
https://pubmed.ncbi.[...]
[4]
서적
Brachiopoda
Geological Society of America/University of Kansas Press
[5]
논문
Brachiopod and mollusc biomineralisation is a conserved process that was lost in the phoronid–bryozoan stem lineage
2022-09-19
[6]
웹사이트
http://www.encyclope[...]
[6]
웹사이트
http://www.mcz.harva[...]
[7]
웹사이트
Marine Species Identification Portal : Brachiopoda of the North Sea
http://species-ident[...]
[8]
서적
Brachiopods
https://books.google[...]
Routledge
[9]
웹사이트
Apatite Ca5(PO4, CO3)3(F, Cl, OH) Hexagonal
http://www.uwex.edu/[...]
University of Wisconsin
2009-10-23
[10]
서적
Glossary of Geology
https://books.google[...]
Springer
[11]
논문
The nervous system of the most complex lophophore provides new insights into the evolution of Brachiopoda
[12]
웹사이트
Brachiopodia Morphology and Ecology
http://paleo.cortlan[...]
State University of New York College at Courtland
2010-03-06
[13]
논문
The attachment strategies of Cambrian kutorginate brachiopods: the curious case of two pedicle openings and their phylogenetic significance
[14]
논문
Earliest ontogeny of the Silurian orthotetide brachiopod ''Coolinia'' and its significance for interpreting strophomenate phylogeny
2017
[15]
논문
Gene Expression Patterns in Brachiopod Larvae Refute the "Brachiopod-Fold" Hypothesis
[16]
논문
Braquiópodos discínidos (Lingulida, Discinoidea) de la Formación Ixtaltepec, Carbonífero del área de Santiago Ixtaltepec, Oaxaca
[17]
서적
Part H, Brachiopoda (Revised)
Geological Society of America; University of Kansas
[18]
논문
Monophyly of brachiopods and phoronids
2000-02-07
[19]
논문
Molecular evidence that phoronids are a subtaxon of brachiopods (Brachiopoda: Phoronata) and that genetic divergence of metazoan phyla began long before the early Cambrian
http://eprints.gla.a[...]
2005-12-01
[20]
논문
Finding a home for hyoliths
https://academic.oup[...]
2019
[21]
웹사이트
The world’s oldest food delicacy revealed
https://web.archive.[...]
[22]
웹사이트
The Potency and Food Safety of Lamp Shells (Brachiopoda: Lingula sp.) as Food Resources
https://iopscience.i[...]
[23]
서적
Applied Palaeontology
https://books.google[...]
[24]
서적
Fishing in Many Waters
https://books.google[...]
[25]
웹사이트
Kentucky's State Fossil: Brachiopods
https://www.uky.edu/[...]
[26]
논문
The scleritome of ''Eccentrotheca'' from the Lower Cambrian of South Australia: Lophophorate affinities and implications for tommotiid phylogeny
[27]
논문
The scleritome of Paterimitra: an Early Cambrian stem group brachiopod from South Australia
2009-05
[28]
서적
Convergent Evolution on Earth. Lessons for the Search for Extraterrestrial Life
https://books.google[...]
MIT Press
2019
[29]
서적
The invertebrates
https://archive.org/[...]
McGraw-Hill
[29]
논문
Annelida and Arthropoda are not sister taxa: A phylogenetic analysis of spiralian metazoan morphology
[29]
서적
Animal evolution: Interrelationships of the living phyla
https://archive.org/[...]
Oxford University Press
[29]
논문
The phylogenetic position of Brachiopoda—a comparison of morphological and molecular data
1997-07
[30]
서적
Microscopic anatomy of invertebrates, volume 4: Aschelminthes
Wiley-Liss
[30]
논문
A critical reappraisal of the fossil record of the bilaterian phyla
2000-05
[31]
서적
Embryology and phylogeny in annelids and arthropods
Pergamon Press Ltd
[32]
논문
Dorsal or ventral: Similarities in fate maps and gastrulation patterns in annelids, arthropods and chordates
[33]
논문
Molecular Data Indicate the Protostome Affinity of Brachiopods
[34]
간행물
日本周辺海域産シャミセンガイ属の再検討
[35]
문서
이전의 학명은 Craniforma이다.
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com