도열병균
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
벼 도열병은 벼에 발생하는 심각한 질병으로, 잎, 잎집, 이삭, 줄기 등 벼의 모든 지상부에 병변을 형성하며 전 세계 85개국 이상에서 발생한다. 병원균은 포자를 통해 감염되며, 압착기를 사용하여 식물에 침투한다. 고온 다습한 환경, 과도한 질소 비료 사용, 가뭄 스트레스 등이 발병을 촉진한다. 벼 도열병은 저항성 품종 육성, 경종적 방법, 화학적 방제 등 다양한 방법을 통합적으로 적용하여 방제하며, 학명과 관련하여 논쟁이 있다. 벼 도열병은 쌀 생산에 큰 피해를 주며, 전 세계적으로 식량 안보에 영향을 미친다.
더 읽어볼만한 페이지
- 동충하초강 - 붉은빵곰팡이
붉은빵곰팡이는 붉은색 분생포자를 형성하는 자낭균류 곰팡이로, 빠른 생장 속도와 유전학 연구 기여, 다양한 생물학 연구 활용, 빵 오염 잡균 등의 특징을 가진다. - 동충하초강 - 동충하초아강
동충하초아강은 자낭균강에 속하며, 단일벽 또는 유사단일벽 자낭과 다양한 형태의 자실체를 가지며, 곤충이나 절지동물 시체에서 자라고 흰색, 노란색, 주황색, 빨간색, 갈색, 검은색 등 다양한 색깔을 띤다. - 대농무기 - 구제역
구제역은 전염력이 매우 강한 가축 질병으로, 구제역 바이러스에 의해 발생하며 고열, 입과 발굽의 수포, 과다한 타액 분비 등의 증상을 보이고, 다양한 혈청형이 존재하며 여러 경로로 전파되어 진단 시 RT-PCR, ELISA 검사가 필요하고 살처분 등의 강력한 방역 조치가 시행된다. - 대농무기 - 살모넬라
살모넬라는 장내세균과에 속하는 그람음성 간균 속으로, 두 종과 2,500개 이상의 혈청형이 존재하며, 오염된 식품 섭취를 통해 식중독 및 다양한 질병을 유발하고, 식품 가열 조리 및 위생 관리, 장티푸스 백신 접종으로 예방이 가능하며, 항생제 내성 문제도 존재한다.

2. 병원균 및 발병 기작
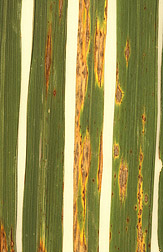
도열병균(''Magnaporthe grisea'', ''M. grisea'')은 자낭균에 속하는 곰팡이로, 매우 효과적인 식물 병원체이다. 이 곰팡이는 유성 생식과 무성 생식을 모두 할 수 있으며, 공중 조직을 감염시키기 위한 특수 구조인 부착기와 뿌리 조직을 감염시킬 수 있는 균사를 생성한다.
벼 도열병은 다양한 벼 품종에서 발생하며, 감수성은 품종에 따라 다르다. 예를 들어 M-201, M-202, M-204, M-205, M-103, M-104, S-102, L-204, Calmochi-101 등의 품종에서 발병이 보고되었으며, 이 중 M-201 품종이 가장 취약한 것으로 알려져 있다.[12]
초기 증상으로는 벼의 모든 지상 부위에 어두운 경계를 가진 흰색에서 회색-녹색의 병반 또는 반점이 나타난다. 병반이 오래되면 타원형 또는 방추형으로 변하고, 중심부는 흰색에서 회색을 띠며 괴사된 경계가 뚜렷해진다. 이 병반들은 점차 커져 서로 합쳐지면서 결국 잎 전체를 고사시킬 수 있다. 증상은 잎뿐만 아니라 잎집, 줄기, 줄기의 마디, 이삭의 목 부분 등 식물의 지상부 전체에서 관찰될 수 있다.[13] 줄기의 마디 사이 감염은 띠 모양으로 나타나며, 특히 마디가 감염되면 해당 부위에서 줄기가 부러지는 '목썩음' 증상이 나타나기도 한다.[14] 또한, 도열병은 벼의 생식 능력에도 영향을 미쳐 종자 생산량을 감소시킨다. 이는 병원균이 곡물의 정상적인 성숙 과정을 방해하기 때문이다.[18]
도열병균은 포자를 통해 전파되며, 이 포자가 벼의 잎, 잎 목, 이삭, 줄기, 마디 등에 부착하여 병반을 형성하며 감염을 시작한다. 병원균은 부착기라는 특수한 구조를 형성하여 식물 조직 내부로 침투한다.[31][15] 침투에 성공한 병원균은 침입성 균사를 뻗어 식물 세포 사이를 이동하거나 원형질연락사를 통해 직접 세포 안으로 들어가 감염을 확산시킨다.[16] ''M. grisea''는 감염된 벼 조직에서 분생포자 형태로 새로운 포자를 대량 생산하여 병을 퍼뜨린다.[17] 병원균은 병든 볏짚이나 그루터기 등에서 월동한 후 다음 해에 다시 활동을 시작하여 감염 순환을 반복한다.[18]
유리한 환경 조건에서는 단일 감염 주기가 약 일주일 만에 완료될 수 있다. 하나의 병반에서 하룻밤 사이에 수천 개의 포자가 생성될 수 있으며, 감염 후 3~4일 만에 새로운 병반이 나타날 수 있다.[19] 병반은 20일 이상 지속적으로 포자를 생산할 수 있기 때문에, 감수성이 높은 벼 품종이 재배되는 환경에서는 도열병이 매우 치명적일 수 있다.[20]
한편, 벼는 도열병균의 침입에 대한 방어 기작을 가지고 있다. 감염 시 벼는 광수확 복합체 II 단백질 LHCB5의 인산화를 유도한다.[1] 이 LHCB5 단백질은 숙주 세포 내에서 활성 산소(ROS)를 급격히 생성하는 데 필요하며, 이 활성 산소 폭발은 도열병균에 대한 저항성을 나타내는 중요한 방어 반응 중 하나이다.[21]
2. 1. 부착기 형성 및 침입
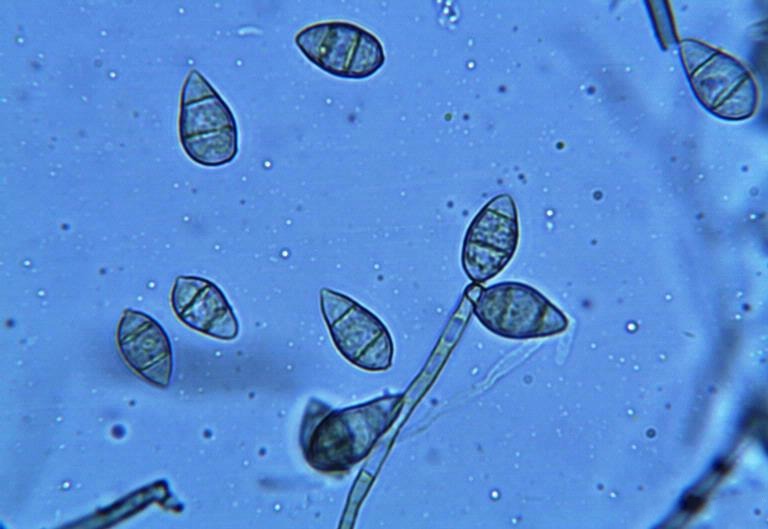
도열병균은 포자를 통해 벼에 감염을 일으킨다. 식물 표면에 도달한 포자는 압착기라는 특수한 구조를 형성하여 식물 조직 내부로 침투를 시도한다. 압착기의 세포벽은 키틴으로 이루어져 있으며, 내부에는 침투 과정에 중요한 역할을 하는 멜라닌 색소가 축적되어 있다.[31][15]
압착기 내에서는 글리세롤이 합성되고 축적되면서 매우 높은 팽압이 발생하며, 단단한 멜라닌 층이 이 압력을 유지하는 데 기여한다.[15] 이 강력한 팽압은 물리적인 힘으로 작용하여 벼 표면의 단단한 식물 큐티클 층을 뚫고 들어간다. 실험적으로는 방탄소재인 케블라 섬유까지 관통할 수 있을 정도의 압력이다.
일단 식물체 내로 침입하는 데 성공하면, 병원균은 침입성 균사를 뻗어 식물 세포 사이를 이동하며 감염을 확산시킨다. 또한, 세포와 세포를 연결하는 통로인 원형질연락사를 통해 직접 세포 안으로 침투하기도 한다.[16]
2. 2. 병원성 관련 유전자
2003년에 전체 게놈 염기 서열 분석이 가능해지고 공개되기 시작하면서 도열병균의 병원성 메커니즘에 대한 유전적 연구가 활발해졌다.[15]병원성에는 여러 유전자가 관여하는 것으로 밝혀졌는데, 특히 멜라닌 합성과 관련된 유전자가 중요하다. 멜라닌은 부착기가 식물 표면을 뚫고 들어가는 데 필요한 팽압을 유지하는 데 필수적인 물질이다.[31][15] 실제로 멜라닌 합성에 결함이 있는 세 가지 균주, 즉 ''albino'' (''ALB1'' 유전자 돌연변이), ''buff'' (''BUF1'' 유전자 돌연변이), ''rosy'' (''RSY1'' 유전자 돌연변이)는 모두 병원성을 잃는 것으로 확인되었다.[31]
병원성 발현에 핵심적인 역할을 하는 유전자 중 하나는 ''pmk1''이다. 이 유전자는 유사분열 활성화 단백질 키나아제(MAPK)를 암호화하며, 효모에서 교배 및 세포 형태 형성에 관여하는 ''FUS3''/''KSS1'' 유전자와 유사하다. 처음에는 ''pmk1''이 도열병균에서도 교배나 발달 과정에만 관여할 것으로 추정되었으나, 연구 결과 부착기 기능과 전체적인 병원성 발현에 필수적인 역할을 하는 것으로 밝혀졌다.[15] ''pmk1'' 유전자에 결함이 생기면 부착기를 제대로 형성하지 못해 식물체 침입 능력을 상실하게 된다.
또 다른 중요한 유전자는 알라닌:글리옥실산 아미노트랜스퍼라제 1(AGT1)을 암호화하는 ''AGT1'' 유전자이다. AGT1 효소는 과산화소체 내에서 산화 환원 항상성을 유지하는 데 중요한 역할을 한다.[34] 도열병균이 식물체에 침입할 때 부착기 내에서는 저장된 지질을 분해하여 에너지를 얻는데, 이 과정(β-산화)에서 FADH2와 NADH 같은 환원 물질이 생성된다. AGT1은 이 환원 물질들을 다시 산화시키는 과정(젖산 발효와 유사)을 촉진하여 부착기 내 산화 환원 균형을 유지하고, 이는 병원성 유지에 필수적이다.[34] 실제로 ''AGT1'' 유전자가 결핍된 돌연변이 균주는 숙주 세포벽을 뚫지 못하고 병원성을 잃는 것이 관찰되었으며, 이는 부착기 내 지질 이용 능력에 문제가 생기기 때문으로 추정된다.[35]
3. 병징
벼 도열병은 벼의 지상부 모든 부분에서 발생할 수 있다.[13] 특히 M-201, M-202, M-204, M-205, M-103, M-104, S-102, L-204, Calmochi-101과 같은 품종에서 관찰되었으며, 이 중 M-201 품종이 병에 가장 취약한 것으로 알려져 있다.[12]
초기 증상으로는 싹의 모든 부분에 어두운 테두리를 가진 흰색에서 회색-녹색의 작은 병반이나 반점이 나타난다. 병이 진행되면 이 병반들은 타원형 또는 방추형으로 커지고, 가운데는 흰색에서 회색을 띠며 가장자리는 괴사된 형태를 보인다. 병반이 점점 커져 서로 합쳐지면 결국 잎 전체가 말라 죽게 된다.[13]
이러한 증상은 잎뿐만 아니라 잎집, 줄기, 줄기의 마디, 이삭의 목 부분 등 식물의 지상부 전체에서 관찰될 수 있다.[13] 줄기의 마디 사이가 감염되면 띠 모양의 병반이 생기기도 한다. 특히 마디 부분이 감염되면 그 부위에서 줄기가 부러지는 '목도열병' 또는 '목썩음' 증상이 나타난다.[14] 또한, 도열병은 벼의 생식 과정에도 영향을 미쳐 종자 생산량을 감소시킨다. 이는 병균이 볍씨가 제대로 여무는 것을 방해하기 때문이다.[18]
4. 발병 환경
벼 도열병은 온대 지역에서 심각한 문제이며, 관개 저지대 및 고지대와 같은 지역에서 발생할 수 있다.[22] 벼 도열병 발생에 유리한 조건은 다음과 같다.
- 습도: 잎이 젖어 있어야 감염이 가능하므로, 장기간의 습윤 환경 및/또는 높은 상대습도가 필요하다.[22] 포자 형성은 높은 상대 습도에서 증가한다.[18]
- 온도: 25°C에서 28°C 사이의 온도에서 포자 발아, 병변 형성 및 포자 형성이 가장 활발하다.[18]
- 재배 환경:
- 과도한 질소 비료 사용은 벼의 식물 병 저항성을 낮추어 병원균에 대한 감수성을 증가시킨다.[18] 이는 식물이 약화된 상태에 놓이고 방어력이 낮아지기 때문이다.[18]
- 가뭄으로 인한 스트레스 역시 벼의 저항성을 약화시킨다.[18]
- 벼 재배 시 논에 물을 대고 빼는 작업이 일반적이지만, 논을 장기간 배수 상태로 두는 것은 감염에 유리한 환경을 조성할 수 있다. 이는 토양이 통기되면서 암모늄이 질산염으로 변환되고, 이것이 벼 작물에 스트레스를 주기 때문이다.[18]
5. 분포
2016년 2월, 방글라데시에서 치명적인 밀 도열병이 발생했다.[25][26] 전사체 분석 결과, 이 병원균은 지리적으로 가까운 지역이 아닌 브라질의 미나스 제라이스, 상파울루, 브라질리아, 고이아스 주에서 유래한 ''M. grisea'' 계통일 가능성이 제기되었다.[25][26] 이러한 유전자 감시 기술은 병원균의 대륙 간 이동 가능성을 밝히고 생물 보안의 중요성을 보여주었으며[25][26], 브라질의 대응 경험을 방글라데시에 적용하는 데 기여했다.[25][26] 이에 따라 방글라데시 정부는 질병의 국내 확산을 막기 위한 조기 경보 시스템을 구축했다.[26]
또한, 2017년부터 2018년 사이 우기에는 잠비아의 무칭가 주 피카 지역에서도 밀 도열병이 발견되었다.[23][24]
6. 방제
도열병균은 살균제와 유전적 저항성을 가진 품종에 대해 빠르게 적응하는 능력이 있어 방제가 까다롭다. 곰팡이는 돌연변이를 통해 살균제에 대한 내성을 키우거나 기존 저항성 품종을 감염시키는 새로운 병원성을 획득할 수 있다.[18]
따라서 ''M. grisea''에 의한 피해를 효과적으로 관리하기 위해서는 어느 한 가지 방법에만 의존하기보다, 다양한 방제 수단을 조합하는 통합 관리 프로그램을 적용하는 것이 매우 중요하다.[18] 이는 병원균의 저항성 발달 속도를 늦추고 방제 효과를 지속시키는 데 도움이 된다. 통합 관리에는 저항성 품종의 재배, 병원균의 월동처가 되는 작물 잔류물 제거, 포자 확산을 억제하기 위한 적절한 물 관리와 같은 경종적 방법, 그리고 작용 방식이 다른 살균제의 계획적인 사용 등이 포함된다.[18]
밀 도열병을 일으키는 균주는 유전자 염기서열 분석(시퀀싱)을 통해 진단할 수 있다.[32] 또한, 중합 효소 연쇄 반응 (PCR), 실시간 PCR (RT-PCR), 고리 매개 등온 증폭 (LAMP)과 같은 분자생물학적 기법을 이용한 유전자 마커 검출 방법도 개발되어 진단에 활용된다.[32] 이러한 마커는 특정 유전자 서열(예: Mot3)이 없는 분리주(예: BR0032)까지 검출할 수 있으며 민감도가 높다는 장점이 있다.[32]
최근에는 작은 RNA나 특정 기능을 가진 짧은 펩타이드를 활용한 혁신적인 생물 모방 살균제 개발 연구도 진행되고 있다.[29]
6. 1. 저항성 품종 육성

도열병균에 대한 유전적 저항성을 가진 벼 품종을 개발하는 것은 육종가들에 의해 이루어졌으며, 이는 주요 방제 전략 중 하나이다. 그러나 도열병균은 돌연변이를 통해 유전적으로 빠르게 변화하여 기존 품종의 저항성을 무너뜨리고 병원성을 나타낼 수 있다. 따라서 ''M. grisea'' 감염을 효과적으로 제어하기 위해서는 단일 품종의 저항성에만 의존하지 않고, 지속적인 저항성 품종 개발과 함께 다른 방제 전략을 통합적으로 관리하는 것이 중요하다.[18] 예를 들어, 저항성이 다른 여러 품종을 함께 재배하거나 돌려짓기하는 방식, 또는 저항성 품종 재배와 함께 살균제 사용, 재배 환경 관리 등을 병행하는 통합 관리 프로그램이 필요하다.[18]
벼는 도열병균에 감염되면 스스로를 방어하기 위해 다양한 생화학적 반응을 일으킨다. 그중 하나로 식물 호르몬인 자스몬산을 생성하여 축적하는데, 이는 병원균 침입에 대한 경보 신호로 작용하여 다른 방어 관련 대사 경로를 활성화시킨다.[37] 하지만 병원균도 이러한 벼의 방어 기작을 회피하기 위해 특정 산화 효소를 만들어 자스몬산 축적을 방해하기도 한다.[37]
벼의 저항성에는 특정 유전자들이 중요한 역할을 한다. 2010년 리뷰에서는 식물의 양적 질병 저항성(quantitative disease resistance)에 대한 유전자 클로닝 연구가 보고되었다.[36] 예를 들어, ''Os''Pii-2 유전자가 만드는 단백질은 면역 수용체로 기능한다.[38] 이 단백질은 벼 세포 내의 Exo70-F3 단백질과 결합하는데, Exo70-F3는 도열병균이 감염 과정에서 분비하는 이펙터 단백질인 AVR-Pii의 표적이 된다. 즉, ''Os''Pii-2는 병원균의 공격 목표물(Exo70-F3)을 감시함으로써 병원균의 침입을 인지하고 방어 반응을 유도하는 역할을 한다.[38] 또한, 일부 벼 품종은 ''OsSWEET13'' 유전자의 특정 대립 유전자를 가지고 있어 저항성을 나타내기도 한다. 이 유전자는 벼 흰잎마름병을 일으키는 세균(''X. oryzae'' pv. ''oryzae'')의 이펙터인 ''PthXo2''의 표적이 되기도 하는 등, 식물의 질병 저항성 메커니즘 연구에서 중요하게 다루어진다.[39]
6. 2. 경종적 방제
과도한 질소 비료 사용은 벼의 식물 병 저항성을 약화시켜 도열병균에 대한 감수성을 높이는 요인이 된다[18]. 또한, 가뭄 스트레스 역시 벼의 저항력을 떨어뜨려 병 발생을 조장할 수 있다[18]. 물 관리 측면에서는 논을 지나치게 오랫동안 마른 상태로 두는 것을 피해야 한다. 장기간의 배수는 토양 통기를 유발하여 암모늄이 질산염으로 변환되는 과정을 촉진하고, 이는 벼 작물에 스트레스를 주어 감염에 더 취약하게 만들 수 있다[18]. 따라서 적절한 시비 관리와 물 관리를 통해 벼의 건강 상태를 유지하고 저항성을 높이는 것이 경종적 방제의 핵심이다.6. 3. 화학적 방제
도열병균 방제를 위해 다양한 살균제가 사용되지만, 병원균이 화학적 치료에 대한 저항성을 획득할 수 있으므로[18], 단일 제어 방법의 과도한 사용을 피하고 다른 작용 방식을 가진 화학적 치료법을 조합하는 등의 통합적인 접근이 필요하다.[18]카프로파미드와 같은 화학적 방제는 도열병균의 부착기가 벼 표피 세포에 침투하는 것을 막아 감염을 억제하는 효과가 있으며, 곡물 자체에는 영향을 미치지 않는 것으로 나타났다.[27] 또한, ''오레가노''와 ''로즈마리''에서 추출한 에센셜 오일이 시험관 내 실험에서 도열병균에 대해 효과를 보이며 방제 가능성을 제시하기도 했다.[28]
최근에는 작은 RNA나 짧은 펩타이드를 이용한 혁신적인 생물 모방 살균제 개발도 이루어지고 있다.[29] 예를 들어, SNP-D4라는 짧은 펩타이드는 도열병균의 칼모듈린에 결합하여 분생 포자 형성을 억제하고 포자 발아를 차단하는 방식으로 작용한다.[29]
7. 학명 논쟁
벼 도열병균(''P. oryzae'')은 1892년에 불완전균으로 처음 기재되었다. 이후 수년간의 연구 끝에 1970년대 후반, 이 균의 유성 세대(텔레오모프)가 ''Magnaporthe grisea''임이 밝혀졌다.[40][41] 당시 벼 도열병균과 메히시바 도열병균(''Pyricularia grisea'')은 서로 교배되지 않는다는 사실이 알려졌지만, 형태적으로 구별하기 어려워 같은 종으로 취급되면서 ''M. grisea''라는 학명이 널리 퍼지게 되었다.
그러나 2000년대에 들어서면서 두 균이 생식적으로 격리되어 있을 뿐만 아니라 유전적으로도 명확한 차이가 있다는 사실이 밝혀졌다. 이에 따라 벼 도열병균을 별도의 종인 ''Magnaporthe oryzae'' (아나모프명 ''P. oryzae'')로 분류해야 한다는 주장이 제기되었다.[42][43]
2013년 이후 균류의 학명 명명 방식에서 유성 세대 이름(텔레오모프명)과 무성 세대 이름(아나모프명)을 따로 쓰던 이중 명명법이 폐지되었다. 이로 인해 먼저 명명된 학명에 우선권을 주는 선명권 원칙에 따라 ''P. oryzae''를 사용해야 할지, 아니면 실제 연구 현장에서 널리 사용되어 온 ''M. grisea'' (또는 ''M. oryzae'')를 사용해야 할지를 두고 논쟁이 벌어졌다.[44] Pyricularia/Magnaporthe Working Groupeng에서의 검토 결과, ''Magnaporthe'' 속 이름을 계속 사용할 경우 여러 가지 복잡한 문제가 발생할 수 있어, 원칙에 따라 ''P. oryzae''를 사용하는 것이 바람직하다는 결론을 내렸다.[45] 현재는 ''P. oryzae'' 사용이 권장되지만, 여전히 ''M. oryzae''도 널리 사용되고 있다.
8. 중요성
벼 도열병은 전 세계 쌀 작물과 관련하여 가장 중요한 질병이다. 쌀은 세계 많은 지역의 중요한 식량원이기 때문에, 그 영향은 광범위하다. 벼 도열병은 전 세계 85개국 이상에서 발견되었으며, 1996년에는 미국에까지 도달했다. 매년 벼 도열병으로 손실되는 작물의 양은 6천만 명을 먹여 살릴 수 있는 규모이다. 일부 저항성 벼 품종이 개발되었지만, 벼 도열병은 쌀이 재배되는 곳이면 어디든 지속적으로 발생하고 있다. 이 질병은 어떤 지역에서도 완전히 근절된 적이 없다.
9. 벼 도열병과 한국
주어진 원본 소스(`source`)가 비어 있어 '벼 도열병과 한국' 섹션의 내용을 작성할 수 없습니다. 해당 섹션을 채우기 위해서는 한국에서의 벼 도열병 발생 현황, 피해 사례, 역사적 기록, 관련 연구 동향 등에 대한 정보가 필요합니다.
9. 1. 대응 및 관리 노력
도열병균에 대한 식물의 저항성 메커니즘 연구는 병 피해를 줄이기 위한 중요한 기초가 된다. 2010년 리뷰에서는 식물의 정량적 질병 저항성에 대한 유전자 복제(클로닝) 연구 결과가 보고되었다.[36]벼는 도열병균의 침입에 반응하여 특정 방어 기작을 활성화한다. 침입을 감지하면 벼는 자스몬산이라는 신호 물질을 방출하는데, 이는 관련된 다른 대사 경로를 작동시켜 방어 반응을 유도한다.[37] 이 과정에서 메틸-자스몬산이 축적된다.[37] 하지만 도열병균은 이러한 벼의 방어 신호 전달을 방해하기 위해 특정 산화 효소를 생합성하여 대응한다.[37]
벼의 저항성에는 특정 유전자와 단백질이 관여한다. 예를 들어, 벼의 단백질 중 하나인 OsPii-2는 면역 수용체처럼 작동한다.[38] 이 단백질은 벼 세포 내의 다른 단백질인 Exo70-F3와 결합한다.[38] Exo70-F3 단백질은 도열병균이 감염 과정에서 분비하는 이펙터 단백질인 AVR-Pii의 표적이 된다. 즉, 도열병균은 AVR-Pii를 이용해 Exo70-F3를 공격하려 하는데, OsPii-2가 Exo70-F3에 결합하여 이러한 공격 시도를 감지하고 방어 체계를 작동시키는 것이다.[38] 이러한 식물 자체의 방어 기작에 대한 이해는 도열병에 강한 벼 품종을 개발하고 관리하는 데 중요한 과학적 기반을 제공한다.
9. 2. 더불어민주당의 정책 방향
주어진 원본 소스에는 '더불어민주당의 정책 방향'과 관련된 내용이 포함되어 있지 않습니다. 원본 소스는 도열병균에 대한 벼의 생물학적 방어 기작에 대한 과학적 설명입니다. 따라서 요청하신 섹션의 내용을 해당 원본 소스를 기반으로 작성할 수 없습니다.참조
[1]
서적
Rice Blast Disease
CABI Centre for Agriculture and Bioscience International
[2]
학술지
Under pressure: Investigating the biology of plant infection by ''Magnaporthe oryzae''
[3]
학술지
The rice leaf blast pathogen undergoes developmental processes typical of root-infecting fungi
[4]
학술지
The genome sequence of the rice blast fungus ''Magnaporthe grisea''
[5]
학술지
A multilocus gene genealogy concordant with host preference indicates segregation of a new species, ''Magnaporthe oryzae'', from ''M. grisea''
[6]
웹사이트
Magnaporthe grisea
http://www.cabicompe[...]
CAB International
[7]
학술지
Rice Blast
American Phytopathological Society (APS)
[8]
학술지
Neck blast disease influences grain yield and quality traits of aromatic rice
https://comptes-rend[...]
Elsevier Masson
[9]
학술지
Partial resistance to neck blast influenced by stage of panicle development and rice genotype
Springer Science+Business Media LLC
[10]
학술지
Resistance to neck blast caused by ''Pyricularia oryzae'' in Italian rice cultivars
Springer Science+Business Media LLC
2015-01-04
[11]
학술지
Origins of host-specific populations of the blast pathogen ''Magnaporthe oryzae'' in crop domestication with subsequent expansion of pandemic clones on rice and weeds of rice
[12]
웹사이트
Rice Blast
http://www.ipm.ucdav[...]
[13]
웹사이트
Rice Blast
http://www.oisat.org[...]
[14]
웹사이트
Rice Blast
http://www.cbwinfo.c[...]
[15]
학술지
Fungal Biology and Agriculture: Revisiting the Field
https://digitalcommo[...]
American Phytopathological Society (APS)
[16]
학술지
A single fungal MAP kinase controls plant cell-to-cell invasion by the rice blast fungus
2018-03-23
[17]
서적
Plant Pathology
Elsevier Academic Press
[18]
웹사이트
Rice Blast: A New Disease in California
http://www.plantscie[...]
University of California-Davis (UCD)
2014-02-25
[19]
학술지
Under pressure: investigating the biology of plant infection by ''Magnaporthe oryzae''
2009-03-01
[20]
웹사이트
Diagnostic Methods for Rice Blast
http://www.padil.gov[...]
[21]
학술지
A double-edged sword: reactive oxygen species (ROS) during the rice blast fungus and host interaction
John Wiley & Sons, Inc.
[22]
웹사이트
Implications of corporate strategies on rice research in asia
http://www.grain.org[...]
Grain (NGO)
2010-10-20
[23]
웹사이트
Researchers in Zambia confirm: Wheat blast has made the intercontinental jump to Africa
http://wheat.org/res[...]
2020-09-24
[24]
학술지
Detection and characterization of fungus (''Magnaporthe oryzae'' pathotype ''Triticum'') causing wheat blast disease on rain-fed grown wheat (''Triticum aestivum'' L.) in Zambia
Public Library of Science
[25]
학술지
Emergence of wheat blast in Bangladesh was caused by a South American lineage of ''Magnaporthe oryzae''
Springer Science+Business Media LLC
2016-10-03
[26]
웹사이트
New infographic highlights an early warning system for wheat blast in Bangladesh
http://wheat.org/new[...]
2020-07-15
[27]
웹사이트
Biological Activity of Carpropamid (KTU 3616): A new fungicide for rice blast disease
http://eurekamag.com[...]
Journal of Pesticide Science
2014-02-25
[28]
서적
Environmental Chemistry for a Sustainable World
Springer International Publishing
[29]
학술지
Game-changing alternatives to conventional fungicides: small RNAs and short peptides
Cell Press
[30]
웹사이트
Rice Blast
http://www.knowledge[...]
[31]
학술지
On the Trail of a Cereal Killer: Exploring the Biology of ''Magnaporthe grisea''
Annual Reviews (publisher)
[32]
서적
Wheat Blast
CRC Press
2020
[33]
웹사이트
Supplement No. 1 to Part 774—The Commerce Control List {{!}} Bureau of Industry and Security
https://www.bis.gov/[...]
2024-08-14
[34]
학술지
Peroxisomal Alanine: Glyoxylate Aminotransferase AGT1 Is Indispensable for Appressorium Function of the Rice Blast Pathogen, ''Magnaporthe oryzae''
2012-04-27
[35]
학술지
Alanine
https://doi.org/10.4[...]
2012-09-01
[36]
논문
Quantitative Disease Resistance and Quantitative Resistance Loci in Breeding
[37]
논문
Biosynthesis and biological function of secondary metabolites of the rice blast fungus ''Pyricularia oryzae''
[38]
논문
Show me your ID: NLR immune receptors with integrated domains in plants
[39]
논문
Gene targeting by the TAL effector PthXo2 reveals cryptic resistance gene for bacterial blight of rice
[40]
논문
The taxonomical identity of the perfect state of ''Pyricularia grisea'' and its allies
[41]
논문
いもち病菌の完全世代に関する研究
http://agriknowledge[...]
[42]
논문
Pathogenicity, Mating Ability and DNA Restriction Fragment Length Polymorphisms of ''Pyricularia'' Populations Isolated from Gramineae, Bambusideae and Zingiberaceae Plants
[43]
논문
A multilocus gene genealogy concordant with host preference indicates segregation of a new species, ''Magnaporthe oryzae'', from ''M. grisea''
http://www.mycologia[...]
[44]
논문
いもち病菌の分類と寄生性分化
https://agriknowledg[...]
[45]
논문
Generic names in ''Magnaporthales''
http://www.imafungus[...]
[46]
저널
Origins of host-specific populations of the blast pathogen Magnaporthe oryzae in crop domestication with subsequent expansion of pandemic clones on rice and weeds of rice
[47]
웹인용
Rice Blast: A New Disease in California
http://www.plantscie[...]
Agronomy Fact Sheet Series 1997-2
2014-02-25
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com