석신산 탈수소효소
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
석신산 탈수소효소(SDH)는 미토콘드리아와 세균에 존재하는 효소로, 4개의 단백질 소단위체(SDHA, SDHB, SDHC, SDHD)로 구성된다. SDHA는 숙신산 산화 부위를, SDHB는 철-황 클러스터를 포함하며, SDHC와 SDHD는 유비퀴논 결합 부위를 형성한다. SDH는 석신산의 산화와 유비퀴논의 환원을 통해 전자 전달을 수행하며, 보결분자단으로 FAD를 사용한다. SDH는 5가지 타입으로 분류되며, 억제제로는 숙신산 포켓과 유비퀴논 포켓에 결합하는 물질이 있다. SDH의 기능 상실은 부신경절종, 신장 세포 암종, 미토콘드리아 복합체 II 결핍 등 다양한 질병을 유발할 수 있다.
더 읽어볼만한 페이지
- EC 1.3 - CH-CH 산화환원효소

2. 구조
미토콘드리아와 많은 세균에 존재하는 석신산 탈수소효소(Succinate Dehydrogenase, SDH)는 4개의 단백질 소단위체로 구성된다. 이들은 크게 두 개의 친수성 소단위체와 두 개의 소수성 소단위체로 나뉜다.
- 친수성 소단위체: 플라보 단백질(SDHA)과 철-황 단백질(SDHB)은 효소 활성이 일어나는 부분이다.
- 소수성 소단위체: SDHC와 SDHD는 세포막에 효소를 고정하는 역할을 한다.
2. 1. 소단위체 구성
석신산 탈수소효소는 4개의 단백질 소단위체로 구성되어 있는데, 2개는 친수성, 2개는 소수성이다. 친수성 소단위체는 플라보 단백질(SDHA)과 철-황 단백질(SDHB)이다. SDHA는 공유 결합된 플라빈 아데닌 디뉴클레오타이드(FAD) 보조 인자와 숙신산 결합 부위를 포함하고, SDHB는 3가지 철-황 클러스터([2Fe-2S], [4Fe-4S], [3Fe-4S])를 포함한다.[3] 소수성 막 앵커 소단위체는 SDHC와 SDHD이다. SDHC와 SDHD는 헴 b 그룹과 유비퀴논 결합 부위를 포함하는 막 결합 사이토크롬 b 복합체를 형성하며, 6개의 막횡단 나선을 가진다. 또한, 카르디올리핀과 포스파티딜에탄올아민이라는 두 개의 인지질 분자가 SDHC 및 SDHD 소단위체에서 발견된다.[4]각 소단위체의 특징은 다음과 같다.[29]
- SDHA: 플라보단백질 소단위체로, FAD 보조 인자가 공유 결합되어 있으며 숙신산 결합 부위가 존재한다.
- SDHB: 철-황 단백질 소단위체로, 3종류의 철-황 클러스터([2Fe-2S], [4Fe-4S], [3Fe-4S])를 포함한다.
- SDHC, SDHD: 소수성 소단위체로, 6개의 막관통 나선과 헴 b를 포함하는 사이토크롬 b를 구성한다. 인지질인 카르디올리핀과 포스파티딜에탄올아민이 결합하고 있다.
인간의 경우, SDHA 소단위체에는 두 가지 아이소타입(FpI, FpII)이 존재한다. 돼지 회충 및 예쁜꼬마선충에서도 SDHA 소단위체의 아이소타입이 발견되었다.[28]
2. 2. 기질 결합 부위
포유류 석신산 탈수소효소(SDH)에는 두 가지 뚜렷한 유비퀴논 결합 부위가 있는데, 기질에 가까운 QP와 기질에서 먼 QD가 있다. 유비퀴논에 대한 친화도가 더 높은 유비퀴논 결합 부위 QP는 SDHB, SDHC, SDHD로 구성된 틈새에 위치한다. 유비퀴논은 B 소단위체의 His207, C 소단위체의 Ser27 및 Arg31, D 소단위체의 Tyr83의 측쇄에 의해 안정화된다. 퀴논 고리는 C 소단위체의 Ile28과 B 소단위체의 Pro160에 의해 둘러싸여 있다. 이러한 잔기는 B 소단위체의 Il209, Trp163, Trp164, C 소단위체의 Ser27 (C 원자)과 함께 소수성 환경의 퀴논 결합 포켓 Qp를 형성한다.[6] 반대로 막간 공간에 더 가까운 유비퀴논 결합 부위 QD는 SDHD만으로 구성되어 있으며 유비퀴논에 대한 친화도가 더 낮다.[7]SDHA는 석신산의 산화를 위한 결합 부위를 제공한다. 서브유닛 A의 측쇄 Thr254, His354, Arg399는 분자를 안정화시키고, FAD는 산화되어 전자를 첫 번째 철-황 클러스터인 [2Fe-2S]로 전달한다.[8]
숙신산 결합 부위는 서브유닛 A의 Thr254, His354, 및 Arg399의 측쇄로 구성되어 있으며, 여기서 FAD에 의한 산화와 첫 번째 철-황 클러스터[2Fe-2S]로의 전자 전달이 일어난다.[30]
유비퀴논 결합 부위는 SdhB, SdhC, 및 SdhD로 구성된 간극에 위치해 있다. 유비퀴논은 서브유닛 B의 His207, 서브유닛 C의 Ser27과 Arg31, 그리고 서브유닛 D의 Tyr83의 각 측쇄에 의해 안정화된다. 퀴논 환은 서브유닛 C의 Ile28과 서브유닛 B의 Pro160에 둘러싸여 있다. 이러한 잔기는 서브유닛 B의 Ile209, Trp163 및 Trp164와, 서브유닛 C의 Ser27(탄소 원자)과 함께 퀴논 결합 포켓의 소수성 환경을 형성한다.[31]
2. 3. 산화 환원 중심
석신산 결합 부위와 유비퀴논 결합 부위는 FAD 및 철-황 클러스터를 포함하는 일련의 산화 환원 중심에 의해 연결된다.[1] 이 사슬은 효소 단량체를 통해 40 Å 이상 뻗어 있으며, 중심 간의 모든 가장자리 대 가장자리 거리는 생리학적 전자 전달에 대해 제안된 14 Å 제한 미만이다.[1] 석신산 결합 부위와 유비퀴논 결합 부위 사이의 FAD와 철-황 클러스터로 구성된 산화 환원 중심은 복합체를 거의 종단하여 총 거리가 40 Å에 달하지만, 각 산화 환원 중심 간의 거리는 생리적 전자 전달의 한계로 제안된 14 Å보다 짧다.[29]3. 조립 및 성숙
사람 미토콘드리아 석신산 탈수소효소의 모든 소단위체는 핵에서 암호화된다. 번역 후, SDHA 소단위체는 아포효소로 미토콘드리아 기질로 이동한다. 그 후, 첫 번째 단계 중 하나는 FAD 조효소의 공유 결합(공유 플라빈화)이다. 이 과정은 석신산 탈수소효소 조립 인자 2(SDHAF2;[9] 효모에서는 SDH5, 박테리아에서는 SDHE라고도 함)와 일부 크렙스 회로 중간체에 의해 촉진된다. 푸마르산은 SDHA의 공유 플라빈화를 가장 강력하게 자극한다.[10] 세균 시스템 연구를 통해 FAD 부착 메커니즘은 퀴논:메티드 중간체를 포함하는 것으로 나타났다.[11] 미토콘드리아 조립 과정에서 SDHA는 최종 복합체에 삽입되기 전에 석신산 탈수소효소 조립 인자 4(SDHAF4; 효모에서는 SDH8이라고 함)라고 하는 두 번째 조립 인자와 상호 작용한다.[7]
소단위체 SDHB의 Fe-S 보조 그룹은 단백질 복합체 ISU에 의해 미토콘드리아 기질에서 미리 형성된다. 이 복합체는 또한 성숙 과정에서 SDHB에 철-황 클러스터를 삽입할 수 있는 것으로 생각된다. 연구에 따르면 Fe-S 클러스터 삽입은 SDHA-SDHB 이량체 형성에 앞선다. 이러한 통합에는 SDHB 활성 부위 내 시스테인 잔기의 환원이 필요하다. 환원된 시스테인 잔기와 이미 통합된 Fe-S 클러스터는 모두 ROS 손상에 매우 취약하다. 두 개의 SDH 조립 인자, SDHAF1(SDH6)과 SDHAF3 (효모의 SDH7)은 ROS로 인한 Fe-S 클러스터 손상으로부터 소단위체 또는 이량체 SDHA-SDHB를 보호하는 방식으로 SDHB 성숙에 관여하는 것으로 보인다.[7]
SDHC 및 SDHD 소단위체로 구성된 소수성 앵커의 조립은 불분명하다. 특히 헴 b 삽입과 그 기능의 경우. 헴 b 보조 그룹은 복합체 II 내 전자 수송 경로의 일부가 아닌 것으로 보인다.[5] 조효소는 오히려 앵커의 안정성을 유지한다.
4. 반응 메커니즘
석신산 탈수소효소에 의해 촉매되는 화학 반응은 다음과 같다.
: 호박산 + 퀴논 푸마르산 + 퀴놀
이 반응은 가역적이지만 호기적 조건에서는 일반적으로 오른쪽 방향으로 진행된다. 혐기적 조건에서는 역반응인 푸마르산 환원 효소로 작용할 수도 있지만, 보통은 역반응을 담당하는 전문 복합체가 존재하거나(대장균 등) 혹은 돼지 회충처럼 일부 서브유닛을 교체하는 것이 알려져 있다.[28]
'''전자 터널링 효과''': 전자는 FAD를 경유하여 석신산으로부터 파생된 후, 터널 효과에 의해 [Fe-S]에서 [3Fe-4S] 클러스터로 중계된다. 이 전자는 그 후 활성 부위의 유비퀴논 분자까지 이동한다.
4. 1. 석신산 산화
석신산 탈수소효소는 보결분자단으로 FAD를 사용하며, 석신산의 산화 반응은 SDHA 소단위체에서 일어난다.[37] FAD는 산화되어 전자를 첫 번째 철-황 클러스터인 [2Fe-2S]로 전달한다.[8]석신산의 산화 반응 메커니즘은 아직 완전히 밝혀지지 않았지만, E2 또는 E1cb 메커니즘을 통해 푸마르산이 생성되는 것으로 제안된다. E2 메커니즘에서는 염기성 잔기 또는 보조 인자가 알파 탄소를 탈양성자화하고, FAD가 베타 탄소로부터 수소화물을 받아 석신산을 푸마르산으로 산화시킨다. E1cb 메커니즘에서는 FAD가 수소화물을 받기 전에 에놀레이트 중간체가 형성된다.
촉매하는 화학 반응은 다음과 같다.
: 호박산 + 퀴논 푸마르산 + 퀴놀
이 반응은 가역적이지만, 호기적 조건에서는 일반적으로 오른쪽 방향으로 진행된다.[28]
4. 2. 전자 전달
석신산 결합 부위와 유비퀴논 결합 부위는 FAD 및 철-황 클러스터를 포함하는 일련의 산화 환원 중심에 의해 연결된다.[1] 석신산의 산화를 통해 FAD에서 유도된 전자는 [Fe-S] 릴레이를 따라 [3Fe-4S] 클러스터에 도달할 때까지 터널링된 후, 활성 부위 내에서 대기 중인 유비퀴논 분자로 전달된다.[1]4. 3. 유비퀴논 환원
유비퀴논의 O1 카보닐 산소는 D 소단위체의 Tyr83과 수소 결합 상호작용을 통해 활성 부위에 위치한다.[6] [3Fe-4S] 철-황 클러스터에 전자가 존재하면 유비퀴논이 두 번째 방향으로 이동하여, 유비퀴논의 O4 카보닐기와 C 소단위체의 Ser27 사이에 두 번째 수소 결합 상호작용이 일어난다. 첫 번째 단일 전자 산화 환원 단계를 거치면 세미퀴논 라디칼 종이 형성된다. [3Fe-4S] 클러스터에서 두 번째 전자가 도착하면 유비퀴논은 유비퀴놀로 완전히 환원된다.[6]유비퀴논 결합 부위는 SdhB, SdhC, SdhD 소단위체로 구성된 틈새에 위치한다. 유비퀴논은 B 소단위체의 His207, C 소단위체의 Ser27과 Arg31, D 소단위체의 Tyr83의 측쇄에 의해 안정화된다. 퀴논 고리는 C 소단위체의 Ile28과 B 소단위체의 Pro160에 의해 둘러싸여 있다. 이러한 잔기들은 B 소단위체의 Ile209, Trp163, Trp164, C 소단위체의 Ser27 (C 원자)과 함께 퀴논 결합 주머니의 소수성 환경을 형성한다.[31]
SQR에서 퀴논을 완전히 환원하려면 2개의 전자와 2개의 프로톤이 필요하다. 물 분자(HOH39)가 활성 부위에 부착되어 B 소단위체의 His207, C 소단위체의 Arg31, D 소단위체의 Asp82에 배위된다고 알려져있다. 세미퀴논은 HOH39에서 유도된 프로톤에 의해 프로톤화되어 유비퀴놀로 환원된다. His207과 Asp82가 이 과정을 돕는 것으로 추정된다. 다른 연구에서는 D 소단위체의 Tyr83이 인접한 히스티딘과 유비퀴논의 O1 카보닐 산소에 배위되어, 히스티딘 잔기가 티로신의 pKa를 감소시켜 유비퀴논 중간체에 프로톤을 제공한다고 제안한다.[33]
4. 4. 헴의 기능
석신산 탈수소효소에서 헴의 기능은 아직 연구 중이다. 몇몇 연구에서는 [3Fe-4S]를 통해 유비퀴논으로 전달되는 전자가 헴과 유비퀴논 중간체 사이를 오갈 수 있다고 주장한다. 이러한 방식으로 헴 보조 인자는 전자 싱크 역할을 한다. 헴의 역할은 중간체가 분자 산소와 상호 작용하여 활성 산소(ROS)를 생성하는 것을 방지하는 것이다.[12]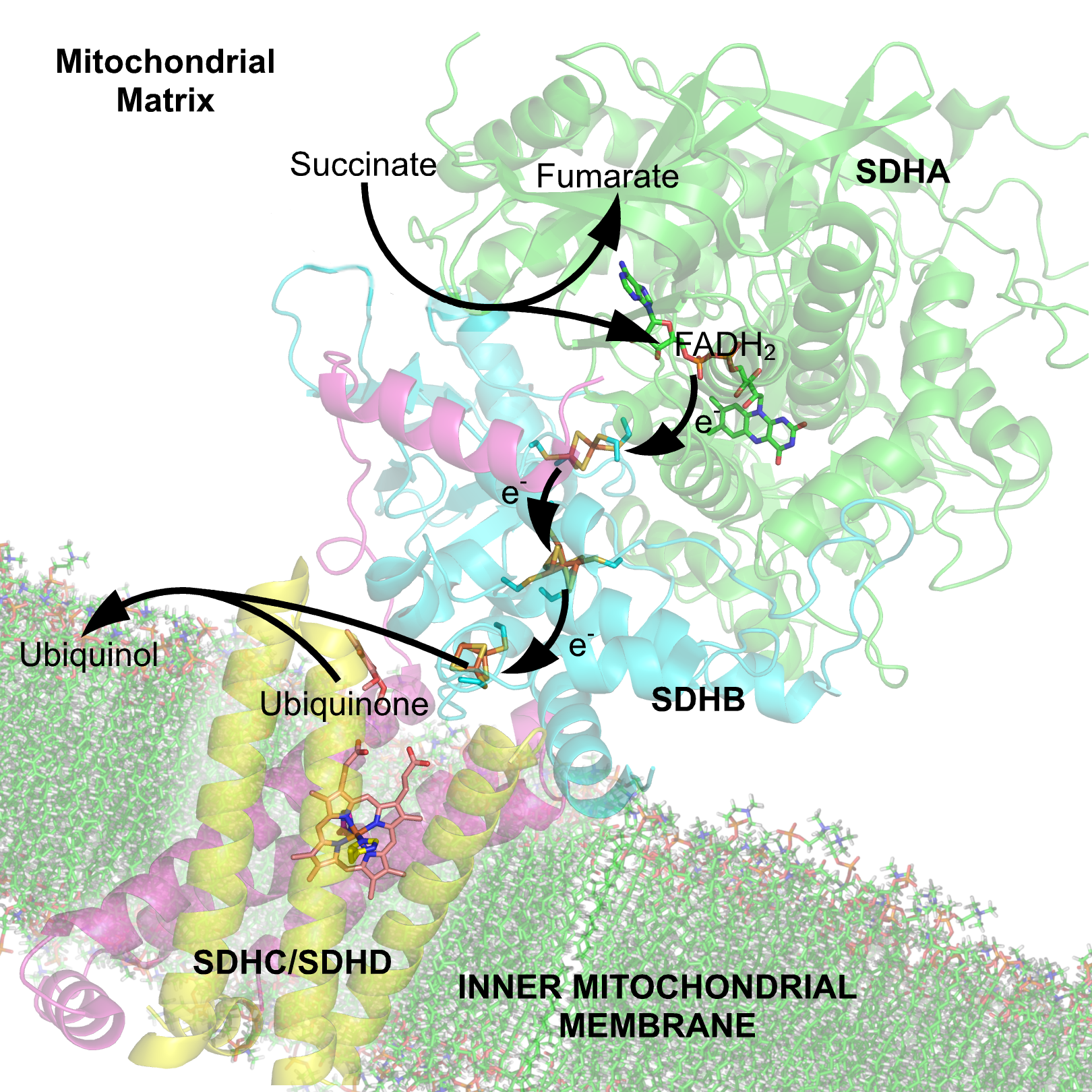
또한 전자가 [3Fe-4S] 클러스터에서 헴으로 직접 이동하는 것을 막는 게이팅 반응 메커니즘이 있을 수 있다는 가설도 있다. 유력한 후보는 [3Fe-4S] 클러스터와 헴 사이에 위치한 잔기 His207이다. B 서브유닛의 His207은 [3Fe-4S] 클러스터, 결합된 유비퀴논 및 헴과 직접 인접해 있으며, 이들 산화 환원 중심 사이의 전자 흐름을 조절할 수 있다.[33]
4. 5. 양성자 이동
SQR에서 퀴논을 완전히 환원하려면 두 개의 전자와 두 개의 프로톤이 필요하다. 물 분자(HOH39)가 활성 부위에 부착되어, 서브유닛 B의 His207, 서브유닛 C의 Arg31, 그리고 서브유닛 D의 Asp82에 배위된다고 주장된다. 세미퀴논은 HOH39에서 유도된 프로톤에 의해 프로톤화되고, 유비퀴논이 유비퀴놀로 환원되는 과정이 완료된다. His207과 Asp82가 이 기구를 용이하게 하는 것으로 생각된다. 다른 연구에서는 서브유닛 D의 Tyr83이 인접한 히스티딘과 유비퀴논의 O1 카르보닐 산소에 배위하고 있다고 제안한다. 이는 히스티딘 잔기가 티로신의 pKa를 감소시켜, 그 프로톤을 환원된 유비퀴논 중간체에 제공하기 쉽게 만들기 때문이다.[33]5. 분류
석신산 탈수소효소는 막 결합 서브유닛에 주목하여 다음과 같이 5가지 종류로 분류할 수 있다.[34]
이 외에도, 종자식물의 복합체 II는 7~8개의 서브유닛으로,[35] 트리파노소마에서는 12개의 서브유닛으로 구성되어 있다.[36]
6. 억제제
석신산 탈수소효소 억제제(SDHI)는 숙신산 포켓에 결합하는 억제제와 유비퀴논 포켓에 결합하는 억제제, 이렇게 두 가지 종류로 나뉜다. 유비퀴논 유형 억제제에는 카복신과 테노일트리플루오로아세톤이 있다. 숙신산 유사체 억제제에는 합성 화합물 말론산뿐만 아니라 말산 및 옥살아세트산과 같은 TCA 회로 중간체가 포함된다. 실제로 옥살아세트산은 복합체 II의 가장 강력한 억제제 중 하나이다. 일반적인 TCA 회로 중간체가 복합체 II를 억제하는 이유는 완전히 이해되지 않지만, 복합체 I에 의해 매개되는 역전자 전달로 인한 과산화물 생성을 최소화하는 데 보호 역할을 할 수 있다.[13] 아트페닌 5a는 유비퀴논 결합을 모방하는 매우 강력한 복합체 II 억제제이다.
유비퀴논 유형 억제제는 1960년대부터 농업에서 살균제로 사용되어 왔다. 카복신은 주로 녹병과 ''Rhizoctonia'' 질병과 같은 담자균류에 의해 발생하는 질병을 제어하는 데 사용되었다. 1980년대에는 단순한 벤즈아닐리드가 카복신과 유사한 활성을 갖는 것으로 밝혀졌으며, 베노다닐, 플루토라닐 및 메프로닐을 포함하여 다수가 시판되었다.[14] 최근에는 보스칼리드, 플루오피람, 플룩사피록사드, 피디플루메토펜 및 세닥산을 포함하여 다양한 식물 병원균에 대한 광범위한 스펙트럼을 가진 다른 화합물들이 개발되었다.[15][14] 일부 농업적으로 중요한 곰팡이는 새로운 세대의 유비퀴논 유형 억제제에 대해 민감하지 않다.[16] FRAC는 SDHI에 대한 실무 그룹을 운영하며[17] 저항성 관리 관행을 권장한다.[18]
복합체 II 억제제는 살충제 및 살비제로도 사용되며, IRAC 그룹 25에 속한다.[19]
7. 질병과의 연관성
석신산 탈수소효소는 산화적 인산화와 시트르산 회로 모두에서 기본적인 역할을 하기 때문에 모든 진핵생물에게 필수적이다. 돌연변이나 독소에 의해 석신산 탈수소효소의 기능이 상실되면 다양한 질병이 유발될 수 있다.
석신산 탈수소효소가 시트르산 회로에서 기능 부전을 겪으면, 종양 대사산물인 석신산이 축적되어 종양 발생을 유발할 수 있다. 이는 크롬친화성 세포에서 잘 알려져 있으며, 부신경절종, 신장 세포 암종, 위장관 기질 종양(GISTs)과 같은 신경내분비 종양을 유발한다.[20] 종양 발생을 유발하는 석신산 탈수소효소 돌연변이에 대한 침투율 데이터는 부족하며, 국제 지침은 모든 보인자에 대한 철저한 검사를 권장한다.[21] 석신산 탈수소효소 기능 상실 돌연변이에서 부신경절종의 침투율은 불완전하며 아단위체에 따라 다르다. SDHB 돌연변이의 침투율은 8%에서 37% 사이이며, SDHD 돌연변이의 침투율은 38%에서 64% 사이이며 일부 모계 각인 효과가 있다. SDHA 및 SDHC 돌연변이의 침투율은 연구가 부족하지만 1%에서 30% 사이일 가능성이 높다.[22][23] 포유류 석신산 탈수소효소는 에너지 생성뿐만 아니라 산소 감지 역할도 한다. 결함이 있는 석신산 탈수소효소로 인해 석신산이 축적되면 가성 저산소증과 혈관 신생이 유발될 수 있으며, 이 두 가지 모두 영상 검사에서 부신경절종의 뚜렷한 혈관성 및 특징적인 "소금과 후추" 외관에 기여한다.[24]
SDHA, SDHB, SDHD, SDHAF1의 이중 대립 유전자 기능 상실 돌연변이나 SDHA의 단일 대립 유전자 기능 상실 돌연변이는 미토콘드리아 복합체 II 결핍을 유발할 수 있다. 산화적 인산화의 이러한 파괴는 Leigh 증후군, 미토콘드리아 뇌병증, 시신경 위축, 근병증 및 다양한 질병을 유발할 수 있다. 이러한 증상은 출생 후 1년 이내 또는 자궁 내 사망에서 성인기에 시작되는 경미한 증상까지 다양할 수 있다.[25]
헌팅턴병 환자의 사후 뇌에서 석신산 탈수소효소 수치가 감소하는 것이 관찰되었으며, 증상 전 및 증상 헌팅턴병 환자 모두에서 에너지 대사 결함이 확인되었다.[26]
참조
[1]
논문
The quaternary structure of the Saccharomyces cerevisiae succinate dehydrogenase. Homology modeling, cofactor docking, and molecular dynamics simulation studies
2004-03
[2]
웹사이트
Using Histochemistry to Determine Muscle Properties
http://muscle.ucsd.e[...]
University of California, San Diego
2009-03-04
[3]
논문
Direct evidence for two distinct forms of the flavoprotein subunit of human mitochondrial complex II (succinate-ubiquinone reductase)
2003-08
[4]
논문
Architecture of succinate dehydrogenase and reactive oxygen species generation
2003-01
[5]
논문
Crystal structure of mitochondrial respiratory membrane protein complex II
2005-07
[6]
논문
Structural and computational analysis of the quinone-binding site of complex II (succinate-ubiquinone oxidoreductase): a mechanism of electron transfer and proton conduction during ubiquinone reduction
2006-03
[7]
논문
Protein-mediated assembly of succinate dehydrogenase and its cofactors
2014-12
[8]
논문
The reaction of N-ethylmaleimide at the active site of succinate dehydrogenase
1975-04
[9]
논문
The roles of SDHAF2 and dicarboxylate in covalent flavinylation of SDHA, the human complex II flavoprotein
2020-09
[10]
논문
How an assembly factor enhances covalent FAD attachment to the flavoprotein subunit of complex II
2022-10
[11]
논문
Crystal structure of an assembly intermediate of respiratory Complex II
2018-01
[12]
논문
The quinone binding site in Escherichia coli succinate dehydrogenase is required for electron transfer to the heme b
2006-10
[13]
논문
High rates of superoxide production in skeletal-muscle mitochondria respiring on both complex I- and complex II-linked substrates
2008-01
[14]
서적
Bioactive Carboxylic Compound Classes: Pharmaceuticals and Agrochemicals
2016
[15]
논문
Progress in understanding molecular mechanisms and evolution of resistance to succinate dehydrogenase inhibiting (SDHI) fungicides in phytopathogenic fungi
[16]
간행물
Differences between the succinate dehydrogenase sequences of isopyrazam sensitive Zymoseptoria tritici and insensitive Fusarium graminearum strains
2013-01
[17]
웹사이트
SDHI Fungicides Working Group
http://www.frac.info[...]
2020-01-31
[18]
웹사이트
Recommendations for SDHI
http://www.frac.info[...]
2020-03
[19]
서적
Modern Crop Protection Compounds
https://onlinelibrar[...]
Wiley‐VCH
2019-01-25
[20]
논문
Succinate dehydrogenase-deficient tumors: diagnostic advances and clinical implications
2012-07
[21]
논문
International consensus on initial screening and follow-up of asymptomatic SDHx mutation carriers
https://doi.org/10.1[...]
2021-07
[22]
논문
The penetrance of paraganglioma and pheochromocytoma in SDHB germline mutation carriers
2018-01
[23]
논문
Mitochondrial complex II and genomic imprinting in inheritance of paraganglioma tumors
2013-05
[24]
웹사이트
salt and pepper sign - paraganglioma
https://radiopaedia.[...]
[25]
논문
The genetic basis of isolated mitochondrial complex II deficiency
2020-09
[26]
논문
Delayed Onset and Reduced Cognitive Deficits through Pre-Conditioning with 3-Nitropropionic Acid is Dependent on Sex and CAG Repeat Length in the R6/2 Mouse Model of Huntington's Disease
https://www.reposito[...]
[27]
논문
The quaternary structure of the Saccharomyces cerevisiae succinate dehydrogenase. Homology modeling, cofactor docking, and molecular dynamics simulation studies
http://www.jbc.org/c[...]
2004-03
[28]
논문
Direct evidence for two distinct forms of the flavoprotein subunit of human mitochondrial complex II (succinate-ubiquinone reductase)
2003
[29]
논문
Architecture of succinate dehydrogenase and reactive oxygen species generation
http://www.sciencema[...]
2003-01
[30]
논문
The reaction of N-ethylmaleimide at the active site of succinate dehydrogenase
http://www.jbc.org/c[...]
1975-04
[31]
논문
Structural and computational analysis of the quinone-binding site of complex II (succinate-ubiquinone oxidoreductase): a mechanism of electron transfer and proton conduction during ubiquinone reduction
http://www.jbc.org/c[...]
2006-03
[32]
논문
UCSF Chimera--a visualization system for exploratory research and analysis
2004-10
[33]
논문
The quinone binding site in Escherichia coli succinate dehydrogenase is required for electron transfer to the heme b
http://www.jbc.org/c[...]
2006-10
[34]
논문
Quinol:fumarate oxidoreductases and succinate:quinone oxidoreductases: phylogenetic relationships, metal centres and membrane attachment
2002
[35]
논문
Functional and composition differences between mitochondrial complex II in Arabidopsis and rice are correlated with the complex genetic history of the enzyme
2010
[36]
논문
Novel mitochondrial complex II isolated from Trypanosoma cruzi is composed of 12 peptides including a heterodimeric Ip subunit
[37]
서적
Biochemistry
W.H. Freeman
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com