유즐동물
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
유즐동물은 8개의 섬모판열을 가지고 있으며, 섬모판열을 움직여 이동하는 투명하고 연약한 이분방사대칭 동물이다. 자포동물과 달리 자포 대신 접착세포를 가지고 먹이를 잡으며, 대부분 자웅동체로 체외 수정을 통해 번식한다. 전 세계 바다에 서식하며, 플랑크톤 생활을 하고 다른 유즐동물이나 해파리 등을 잡아먹는 포식자이다. 유즐동물은 유촉수강과 무촉수강으로 분류되며, 최근 연구에서는 다른 모든 동물의 자매군일 가능성이 제기되기도 한다.

더 읽어볼만한 페이지
- 유즐동물 - 유촉수강
유촉수강은 빗해파리류의 한 강으로, Cyclocoela와 맹장빗해파리아강으로 나뉘며 각 아강은 특정한 형태적, 생태적 특징을 지닌 여러 목으로 구성된다. - 유즐동물 - 넓적빗해파리목
넓적빗해파리목은 납작하고 타원형 몸체를 가진 해양 무척추동물로, 위장색을 띠며 해저나 바위에 서식하고, 유성생식과 무성생식을 하며 외부 수정을 통해 번식하고, 일부 종은 착생생물과 공생 관계를 맺기도 한다. - 생물 분류에 관한 - 다람쥐
다람쥐는 등 쪽에 줄무늬가 있는 다람쥐속 설치류로, 홀로 생활하며 겨울잠을 자고 씨앗, 견과류, 곤충 등을 먹으며 맹금류 등의 먹이가 되고, 특히 한반도 서식 다람쥐는 새로운 종으로 분류될 가능성이 있으며, 유럽에서는 침입 외래종으로 지정되기도 하고 라임병을 옮길 수 있어 주의가 필요한 동물이다. - 생물 분류에 관한 - 황금랑구르
황금랑구르는 인도 아삼 주와 부탄에 분포하며 크림색에서 황금색 털을 가진 멸종위기종 영장류로, 서식지 파괴로 개체수가 감소하여 보호받고 있다. - 표시 이름과 문서 제목이 같은 위키공용분류 - 라우토카
라우토카는 피지 비치레부섬 서부에 위치한 피지에서 두 번째로 큰 도시이자 서부 지방의 행정 중심지로, 사탕수수 산업이 발달하여 "설탕 도시"로 알려져 있으며, 인도에서 온 계약 노동자들의 거주와 미 해군 기지 건설의 역사를 가지고 있고, 피지 산업 생산의 상당 부분을 담당하는 주요 기관들이 위치해 있다. - 표시 이름과 문서 제목이 같은 위키공용분류 - 코코넛
코코넛은 코코넛 야자나무의 열매로 식용 및 유지로 사용되며, 조리되지 않은 과육은 100g당 354kcal의 열량을 내는 다양한 영양 성분으로 구성되어 있고, 코코넛 파우더의 식이섬유는 대부분 불용성 식이섬유인 셀룰로오스이며, 태국 일부 지역에서는 코코넛 수확에 훈련된 원숭이를 이용하는 동물 학대 문제가 있다.

2. 특징
유즐동물은 8개의 섬모판열을 가지고 있으며, 이 섬모판열은 섬모가 융합된 빗 모양의 구조로 되어 있다. 이 섬모를 물결치듯 움직여 이동한다.[134] 섬모판열은 빛의 반사에 의해 무지개색으로 빛나는 경우가 있는데, 이는 생물발광이 아닌 구조색이다.[135]
몸은 이분방사대칭이며, 투명하고 연약하다.[133] 대부분의 종은 무색투명하지만, 일부 심해 종은 뚜렷한 색을 띠기도 한다.
자포동물과 달리 자포가 없고, 대신 교포를 가지고 있어 먹이를 잡는다.[134] 일부 종은 먹이인 해파리의 자포를 흡수하여 사용하기도 한다(도자포).[134]
몸은 두 개의 주요 세포층 사이에 젤리 같은 중교질층이 있는 구조로, 자포동물과 유사하지만, 더 복잡한 근육과 신경계를 가지고 있다.[17][18]
소화계는 입, 인두, 위, 그리고 관 시스템으로 구성되며, 항문 구멍을 통해 노폐물을 배출한다.[26]
뇌나 중추 신경계는 없지만, 표피 아래에 신경망을 가지고 있으며,[26] 몸의 균형을 감지하는 평형포를 가지고 있다.[26]
대부분 자웅동체이며, 체외 수정을 통해 번식한다.[133][26] 손상된 조직을 재생할 수 있으며, 일부 종은 분열생식을 통해 번식하기도 한다.[59][26]
유즐동물은 신경계와 근육 세포 유형이 다른 동물들과 다르다는 점에서 독특하며, 최근 연구에서는 유즐동물이 다른 모든 동물의 자매군일 가능성이 제기되기도 한다.[36][47]
빗해파리는 대부분의 해양 환경에서 발견된다. 영하 2°C의 극지방 수역에서부터 30°C의 열대 지방까지, 연안 근처와 대양 중앙, 수면에서 수심 7000미터 이상의 심해까지 분포한다.[73] 가장 잘 알려진 속은 ''Pleurobrachia'', ''Beroe'', 그리고 ''Mnemiopsis''이며, 이러한 플랑크톤성 연안 종들은 해안 근처에서 채집될 가능성이 가장 높기 때문이다.[29][77] 담수에서는 빗해파리가 발견된 적이 없다.
유즐동물은 전통적으로 촉수의 유무에 따라 유촉수강 (Tentaculata)과 무촉수강 (Nuda)의 두 강으로 나뉜다.
3. 생태
대부분 종의 성체는 손상되거나 제거된 조직을 재생할 수 있다.[59]
빗해파리는 일부 연안 지역에서는 여름철에 풍부할 수 있지만, 다른 지역에서는 드물고 찾기 어렵다. 빗해파리가 매우 많은 수로 서식하는 만에서는, 빗해파리에 의한 포식이 요각류와 같은 작은 동물성 플랑크톤의 개체군을 조절할 수 있다. 그렇지 않으면 이러한 요각류가 해양 먹이 사슬의 중요한 부분인 식물 플랑크톤(플랑크톤성 식물)을 멸종시킬 수도 있다.
거의 모든 유즐동물은 포식자이다. 초식 유즐동물은 없으며, 일부 기생하는 속(genus)이 단 하나뿐이다.[77] 먹이가 풍부하면 자신의 체중의 10배를 하루에 먹을 수 있다.[76] ''Beroe''속은 주로 다른 유즐동물을 먹지만, 다른 표수종들은 현미경적인 크기의 동물플랑크톤(플랑크톤성 동물)부터, 연체동물과 어류의 유생을 포함하여, 요각류, 단각류, 심지어 크릴과 같은 작은 성체 갑각류에 이르기까지 다양한 크기의 먹이를 잡아먹는다. ''Haeckelia''속에 속하는 종들은 해파리를 잡아먹고, 자신의 먹이의 자포(쏘는 세포)를 접착세포 대신 자신의 촉수에 통합한다.[27]
유즐동물은 유기물과 염분 및 물의 비율이 낮아 다른 동물의 먹이로 적합하지 않다고 여겨졌기 때문에 과거 해양 먹이 사슬의 "막다른 길"로 여겨졌다. 또한, 가능한 포식자의 내장에서 유즐동물의 잔해를 확인하기 어려운 경우가 많은데, 빗 모양의 섬모는 때때로 충분히 오래 남아 단서를 제공한다. 연어(Oncorhynchus keta)에 대한 자세한 조사에 따르면, 이 물고기는 유즐동물을 같은 무게의 새우보다 20배 빠르게 소화하며, 유즐동물이 충분히 있다면 좋은 먹이가 될 수 있다는 것을 보여주었다. 베로이드는 주로 다른 유즐동물을 먹는다. 일부 해파리와 거북이는 다량의 유즐동물을 먹으며, 해파리는 유즐동물 개체군을 일시적으로 멸종시킬 수 있다. 유즐동물과 해파리는 종종 계절적인 개체군 변화가 크기 때문에, 이들을 먹는 대부분의 물고기는 잡식성이며, 전문적인 해파리 먹이보다 개체군에 더 큰 영향을 미칠 수 있다.
일부 말미잘의 유생은 유즐동물에 기생하며, 어른이 되면 물고기에 기생하는 일부 편형동물의 유생도 마찬가지이다.[80] 대부분의 유즐동물 종은 자웅동체이며, 적어도 일부 종의 유체는 성체의 크기와 형태에 도달하기 전에 번식이 가능하다. 이러한 자웅동체와 조기 번식의 결합으로 인해 소규모 개체군이 폭발적으로 증가할 수 있다.
유즐동물은 모든 식물플랑크톤(플랑크톤성 식물)을 먹는 동물플랑크톤의 과다 증식을 억제함으로써 해양 생태계의 균형을 유지하는 역할을 할 수 있다.[81] 식물플랑크톤은 무기물에서 유기물을 생산하는 주요 해양 생산자이다.[82]
1980년대 후반에는 서대서양 유즐동물인 ''Mnemiopsis leidyi''가 선박의 밸러스트탱크를 통해 흑해와 아조프해로 우연히 유입되었으며, 어류의 유생과 소형 갑각류를 섭취하여 성어의 먹이를 감소시켜 어획량 감소의 원인으로 지목되었다.[81] ''Mnemiopsis''는 매우 빠르게 번식하고 넓은 범위의 수온과 염분을 견딜 수 있기 때문에 새로운 지역으로 침입하는 데 적합하다. 해당 지역의 ''Mnemiopsis'' 개체군은 결국 ''Mnemiopsis''를 먹는 북미산 유즐동물인 ''Beroe ovata''의 우연한 유입[85]과 1991년부터 1993년까지의 지역 기후의 저온화[84]에 의해 통제되었다.
1990년대 후반에는 ''Mnemiopsis''가 카스피해에 나타났다. ''Beroe ovata''는 그 직후에 도착했으며, 그곳에서 ''Mnemiopsis''의 영향을 줄일 것으로 예상되지만 완전히 제거하지는 못할 것이다. ''Mnemiopsis''는 1990년대 후반에 동부 지중해에 도달했으며, 현재 북해와 발트해에서 번성하고 있는 것으로 보인다.[27]
4. 분류
하지만 분자 계통 분석은 이러한 전통적인 분류를 지지하지 않으며, 유즐동물 문의 내부 계통 관계는 아직 완전히 밝혀지지 않았다.[133] 과거에는 자포동물과 함께 강장동물로 분류되기도 했으나, 최근에는 독립된 동물문으로 분류하는 것이 일반적이다.
4. 1. 유촉수강 (Tentaculata)

유촉수강(Tentaculata) 빗해파리목(Cydippida) 동물들은 대체로 둥근 형태를 띠며, 구형, 원통형, 알 모양 등 다양하다. 흔히 해안가에서 볼 수 있는 "바다 구즈베리"인 빗해파리(Pleurobrachia)는 좁은 쪽에 입이 있는 알 모양의 몸체를 가지기도 하지만,[26] 일부 개체는 더 둥글다. 몸체 반대쪽에는 길고 가는 촉수 한 쌍이 있으며, 각각 촉수 주머니 안에 들어 있다.[17] 일부 빗해파리목 종들은 몸체가 납작해져서 촉수가 있는 면이 더 넓다.[26]
빗해파리목 유즐동물의 촉수는 보통 작은 촉수(tentilla)가 가장자리에 늘어서 있지만, 몇몇 속은 가지가 없는 단순한 촉수를 가진다. 촉수와 작은 촉수는 미세한 접착세포로 덮여 있어 먹이를 잡는다. 접착세포는 표피 바깥층에 있는 버섯 모양 세포로, 점착성 물질이 든 소포(vesicles)가 있는 돔형 머리, 세포를 고정하는 줄기, 줄기에 감겨 머리와 뿌리에 연결된 나선형 실의 세 부분으로 구성된다.[26] 지름 1.5mm인 ''Minictena luteola''는 다섯 가지 유형의 접착세포를 가진다.[48][49]
해파리를 주로 먹는 ''해켈리아''(Haeckelia)는 먹이의 자포(nematocyte)를 자신의 촉수에 통합한다. 일부 자포동물을 먹는 민달팽이도 방어를 위해 자포를 몸에 통합한다.[50] ''유플로카미스''(Euplokamis)의 작은 촉수는 다른 빗해파리목 종들과 달리 횡문근을 포함하며, 이완 시 나선형으로 감긴다.[51]
입 근처에서 반대쪽 끝까지 8줄의 빗 모양 섬모열이 몸 전체에 고르게 분포한다.[17] "빗"은 메타크로날 리듬으로 움직이며, 멕시코 웨이브와 비슷하다.[53]
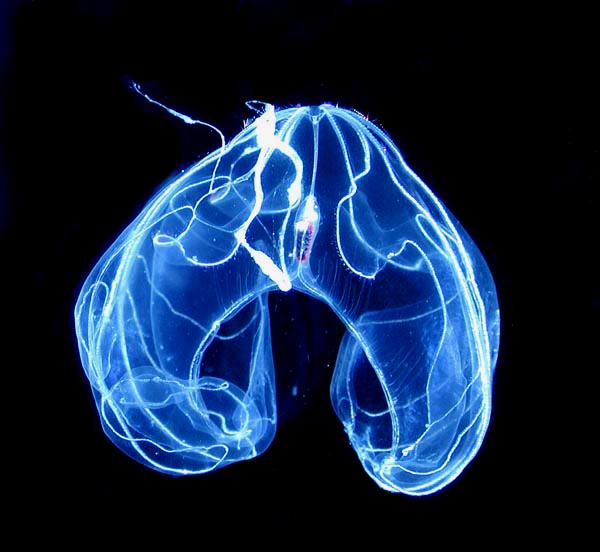
유즐동물(Lobata)은 입보다 바깥쪽으로 돌출된 한 쌍의 근육질 컵 모양 돌기인 엽(lobe)을 가진다. 눈에 잘 띄지 않는 촉수는 입가에서 시작하여 복잡하게 구불거리는 홈을 따라 엽 안쪽 표면으로 퍼진다. 많은 유즐동물 종은 입 양쪽 엽 사이에 네 개의 귀(auricle)를 가지는데, 이는 섬모가 있는 젤라틴질 돌기로, 미세한 먹이를 입 쪽으로 유도하는 물 흐름을 생성한다. 이러한 구조를 통해 유즐동물은 부유성 플랑크톤을 지속적으로 섭취한다.[26]
유즐동물은 8개의 빗판열(comb-rows)을 가지며, 입이 없는 극(aboral pole)에서 시작하여 보통 몸보다 엽까지 확장되지 않는다. 귀가 있는 종에서는 귀 가장자리의 섬모가 4개 빗판열 섬모의 연장이다. 대부분 유즐동물은 물속에서 이동할 때 빗판열의 섬모를 추진력으로 사용한다.[5]
가네시다(Ganeshida)는 작은 구엽(oral lobe) 한 쌍과 촉수 한 쌍을 가진다. 몸은 원형 단면이며, 인두(pharynx)는 구엽 안쪽 표면으로 뻗어 있다.[26]
탈라소칼리시다(Thalassocalycida)는 1978년에 발견되었으며 단 한 종만 알려져 있다.[57] 해파리와 비슷하며, 몸이 짧고 입에서 가장 먼 쪽 표면에 짧은 빗판(comb-row)이 있다. 종(bell)의 움직임과 짧은 촉수를 사용하여 먹이를 잡는다.[26]
세스티다(Cestida)("벨트 동물")는 리본 모양의 플랑크톤성 동물로, 입과 반대쪽 끝 기관이 리본 반대쪽 가장자리 중앙에 있다. 각 반대쪽 끝에는 빗판 한 쌍이 있고, 입 가장자리를 따라 홈에서 나오는 텐틸라(tentilla)가 몸 표면 대부분을 가로지른다. 세스티다는 몸을 흔들거나 빗판을 움직여 헤엄친다.[26][58]
대부분의 플라티크테니다(Platyctenida)는 납작한 타원형 몸체를 가지며, 반대쪽 끝 표면에 텐틸라를 가진 촉수 한 쌍이 있다. 인두를 뒤집어 근육질 "발"로 사용하여 표면에 붙어 기어 다닌다. 알려진 플라티크테니드 종 중 하나를 제외하고는 모두 빗판이 없다.[26]
4. 2. 무촉수강 (Nuda)
오이빗해파리목 (Beroida)은 촉수가 없고, 큰 입을 가진 유즐동물이다. 다른 유즐동물이나 해파리를 잡아먹는다.[55]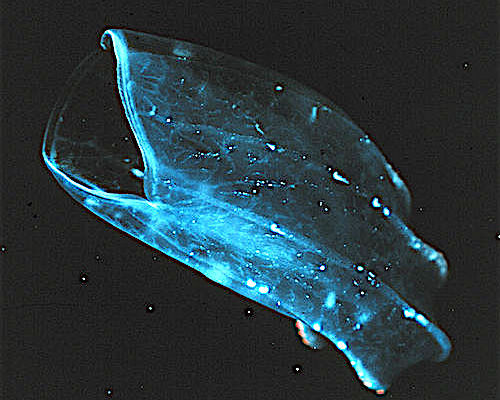
베로이다(Beroida)목, 또는 누다(Nuda)목으로도 알려진 이들은 먹이를 잡는 부속물이 없지만, 큰 입 안쪽에 위치하고 주머니 모양의 몸 대부분을 차지하는 큰 인두(pharynx)에는 구강 끝에 "대섬모(macrocilia)"가 있다. 이 수천 개의 큰 섬모가 융합된 다발은 너무 커서 통째로 삼킬 수 없는 먹이(거의 항상 다른 유즐동물)를 "물어뜯어"낼 수 있다.[55] 일부 ''Beroe'' 종의 입술 부분, 대섬모 밭 앞에는 위벽에 접착 상피세포로 이루어진 좁은 두 줄이 있는데, 이 세포들이 서로 연결되어 먹이를 먹지 않을 때 입을 닫는다. 이러한 밀착된 폐쇄는 먹이를 추격할 때 동물의 앞부분을 유선형으로 만들어준다.[56]
5. 진화
유즐동물은 몸이 연약하여 화석으로 남기 매우 어렵지만, 캄브리아기 중기 지층에서 촉수가 없고 섬모판열이 많은 유즐동물 화석이 발견되기도 했다.[3] 1990년대 중반까지는 데본기 지층에서 발견된 화석 2개가 전부였으나, 이후 버제스 셰일 등에서 캄브리아기 중기 화석 3개가 추가로 발견되었다. 이 화석들은 현생 유즐동물과 달리 촉수가 없고 섬모판열이 24~80개로 훨씬 많았으며, 현생종에는 없는 내부 기관도 가지고 있었다.[137] 중국에서 발견된 화석들은 캄브리아기에 유즐동물이 이미 다양하게 분화되어 있었지만, 섬모판열이 몸에서 돌출된 날개 위에 있는 등 현생종과는 여러 면에서 달랐음을 보여준다.[138]
유즐동물과 다른 동물과의 계통 관계는 오랫동안 논쟁의 대상이었다. 초기 학자들은 형태적 유사성을 근거로 유즐동물을 자포동물과 함께 강장동물문으로 분류하기도 했다. 그러나 유전자 연구 결과, 유즐동물의 뉴런은 다른 동물의 뉴런과 독립적으로 진화했을 가능성이 제기되었고,[87] 최근에는 유즐동물과 자포동물을 별개의 문으로 분류하는 추세이다. 분자 계통 발생학적 연구에 따르면, 자포동물과 후구동물이 유즐동물보다 서로 더 가깝게 관련되어 있다.
최근 연구에서는 유즐동물이 다른 모든 동물의 자매군일 가능성이 제기되기도 한다.[8][105] 이는 유즐동물의 신경계와 근육 세포가 다른 동물들과 매우 다르다는 점을 시사한다. 유즐동물의 신경 세포와 신경계는 다른 동물들과 다른 생화학적 특성을 가지고 있다. 예를 들어, 다른 동물에서 발견되는 세로토닌, 도파민 등의 신경 전달 물질을 만드는 데 필요한 유전자와 효소가 없으며,[43] 신경 전달 물질로 L-글루탐산을 사용한다.[45] 유즐동물의 신경계 유전자 함량은 알려진 동물 중 가장 작으며,[46] 신경계의 일부는 시냅스 없이 직접 융합된 뉴런으로 구성되어 있다.[36] 이러한 특징들은 유즐동물이 독자적인 진화 경로를 거쳤음을 보여준다.
하지만 유즐동물을 다른 모든 동물의 자매군으로 배치하는 것이 유즐동물 게놈의 높은 진화율 때문이라는 주장도 있으며,[104] 해면동물이 가장 먼저 갈라진 동물 분류군이라는 주장도 있다.[113] 또 다른 연구에서는 해면동물이 다른 모든 현존 동물의 자매군이라는 가설을 반박하고 유즐동물을 다른 모든 동물의 자매군으로 재배치하기도 했다.[12]
참조
[1]
논문
Raman spectra of a Lower Cambrian ctenophore embryo from southwestern Shaanxi, China
2007-04-00
[2]
논문
The first fossil ctenophore from the Lower Devonian of West Germany
1983-06-09
[3]
논문
Middle Cambrian Ctenophores from the Stephen Formation, British Columbia, Canada
1996-03-29
[4]
논문
A vanished history of skeletonization in Cambrian comb jellies
2015-07-00
[5]
백과사전
Ctenophora
[6]
보고서
World Register of Marine Species
http://www.marinespe[...]
VLIZ
2024-02-19
[7]
논문
A large and consistent phylogenomic dataset supports sponges as the sister group to all other animals
[8]
논문
Broad phylogenomic sampling improves resolution of the animal tree of life
[9]
논문
The genome of the Ctenophore ''Mnemiopsis leidyi'' and its implications for cell type evolution
2013-12-13
[10]
논문
Genomic data do not support comb jellies as the sister group to all other animals
[11]
서적
Spineless: The science of jellyfish and the art of growing a backbone
Riverhead Books (Penguin Random House)
2017-11-00
[12]
논문
Ctenophore relationships and their placement as the sister group to all other animals
2017-11-00
[13]
논문
Miscues misplace sponges
2016-02-09
[14]
논문
A sisterly dispute
2016-01-00
[15]
논문
Ancient gene linkages support ctenophores as sister to other animals
2023-05-17
[16]
논문
Meeting report of Ctenopalooza: The first international meeting of ctenophorologists
2016-12-00
[17]
서적
Invertebrate Zoology
Oxford University Press
[18]
서적
Invertebrate Zoology
https://archive.org/[...]
Brooks / Cole
[19]
논문
Evolution of striated muscle: Jellyfish and the origin of triploblasty
2005-06-00
[20]
bioRxiv
Atlas of Neuromuscular Organization in the Ctenophore, Pleurobrachia bachei {{small|(A. Agassiz, 1860)}}
2018-08-16
[21]
논문
The invasive ctenophore ''Mnemiopsis leidyi'' overwinters in high abundances in the subarctic Baltic Sea
[22]
백과사전
Ctenophore
Oxford University Press
[23]
서적
Invertebrate Zoology
https://archive.org/[...]
Brooks / Cole
[24]
서적
Invertebrate Zoology
Oxford University Press
[25]
논문
The ctenophore genome and the evolutionary origins of neural systems
2014-06-00
[26]
서적
Invertebrate Zoology
https://archive.org/[...]
Brooks / Cole
[27]
웹사이트
Ctenophores – some notes from an expert
http://faculty.washi[...]
University of Washington
2009-02-05
[28]
논문
Evolution of collagens
[29]
논문
A golden age of gelata: past and future research on planktonic ctenophores and cnidarians
2004-11-00
[30]
논문
Intracellular Fate Mapping in a Basal Metazoan, the Ctenophore ''Mnemiopsis leidyi'', Reveals the Origins of Mesoderm and the Existence of Indeterminate Cell Lineages
1999-10-00
[31]
논문
The fine structure of the cilia from ctenophore swimming-plates
[32]
논문
Escape of the ctenophore ''Mnemiopsis leidyi'' from the scyphomedusa predator ''Chrysaora quinquecirrha''
1997-06-00
[33]
논문
Density is Altered in Hydromedusae and Ctenophores in Response to Changes in Salinity
https://www.biodiver[...]
1984-02-00
[34]
웹사이트
Alien-like comb jellies have a nervous system like nothing ever seen before
https://www.livescie[...]
[35]
웹사이트
The jellyfish with a nervous system that is causing a shiver in the scientific community
https://web.archive.[...]
[36]
논문
Syncytial nerve net in a ctenophore adds insights on the evolution of nervous systems
https://doi.org/10.1[...]
2023-04-20
[37]
논문
Cambrian comb jellies from Utah illuminate the early evolution of nervous and sensory systems in ctenophores
2021
[38]
논문
The gluey tentacles of comb jellies may have revealed when nerve cells first evolved
2019-01-10
[39]
웹사이트
Did the ctenophore nervous system evolve independently?
http://ryanlab.whitn[...]
[40]
논문
Larval body patterning and apical organs are conserved in animal evolution
2014-01-29
[41]
논문
Larval nervous systems: true larval and precocious adult
2015-02-15
[42]
논문
Early animal evolution: a morphologist's view
2019-07
[43]
웹사이트
Aliens in our midst
https://aeon.co/essa[...]
2017-08-01
[44]
논문
Antioxidant enzymes that target hydrogen peroxide are conserved across the animal kingdom, from sponges to mammals
2023-02-13
[45]
논문
Neural system and receptor diversity in the ctenophore Beroe abyssicola
2019-08-15
[46]
서적
Structure and Evolution of Invertebrate Nervous Systems
Oxford University Press
[47]
논문
The phylogenetic position of ctenophores and the origin(s) of nervous systems
2015
[48]
논문
Five types of colloblast in a cydippid ctenophore, ''Minictena luteola'' Carré and Carré: An ultrastructural study and cytological interpretation
1993
[49]
웹사이트
Characterizing functional biodiversity across the phylum Ctenophora using physiological measurements
https://archives.eve[...]
[50]
논문
Ingestion of a medusa (''Aegina citrea'') by the nematocyst-containing ctenophore (''Haeckelia rubra'', formerly ''Euchlora rubra''): Phylogenetic implications
1984-02
[51]
논문
Structure and function of the prehensile tentilla of ''Euplokamis'' (Ctenophora, Cydippida)
1988-03
[52]
논문
Mechanisms of ciliary co-ordination in Ctenophores
1973
[53]
논문
Physical constraints on the evolution of ctenophore size and shape
1990-04
[54]
논문
An undescribed lobate ctenophore, ''Lobatolampea tetragona'' gen. nov. & spec. nov., representing a new family, from Japan
http://www.repositor[...]
2000-03
[55]
논문
Visualization of changes in ciliary tip configuration caused by sliding displacement of microtubules in macrocilia of the ctenophore ''Beroe''
1985
[56]
논문
Reversible epithelial adhesion closes the mouth of ''Beroe'', a carnivorous marine jelly
https://www.biodiver[...]
1991
[57]
논문
What determines the likelihood of species discovery in marine holozooplankton: is size, range or depth important?
2005-06
[58]
서적
Pacific Coast Pelagic Invertebrates: A Guide to the Common Gelatinous Animals
https://archive.org/[...]
Sea Challengers and Monterey Bay Aquarium
[59]
논문
The ontogeny and maintenance of adult symmetry properties in the ctenophore, ''Mnemiopsis mccradyi''
1986-12
[60]
웹사이트
Rapid physiological integration of fused ctenophores
https://www.cell.com[...]
[61]
논문
A reconstruction of sexual modes throughout animal evolution
2017-12
[62]
논문
Ctenophores are direct developers that reproduce continuously beginning very early after hatching
2022
[63]
논문
Not all ctenophores are hermaphrodites. Studies on the systematics, distribution, sexuality and development of two species of Ocyropsis
1986-02
[64]
논문
Developmental expression of 'germline'- and 'sex determination'-related genes in the ctenophore ''Mnemiopsis leidyi''
2016
[65]
논문
Ctenophore population recruits entirely through larval reproduction in the central Baltic Sea
2012
[66]
논문
Nitric oxide signaling in ctenophores
2023
[67]
웹사이트
This Benjamin Button-like Jellyfish Can Age in Reverse, From Adult to Juvenile
https://www.zmescien[...]
[68]
웹사이트
Phylum Ctenophora: list of all valid scientific names
http://faculty.washi[...]
2007-05
[69]
논문
Optical properties of the iridescent organ of the comb-jellyfish Beroë cucumis (Ctenophora)
2006-04-14
[70]
논문
Not all ctenophores are bioluminescent: Pleurobrachia
https://www.biodiver[...]
1995-12
[71]
논문
Bioluminescence spectra of shallow and deep-sea gelatinous zooplankton: Ctenophores, medusae and siphonophores
1999-04-08
[72]
논문
Genomic organization, evolution, and expression of photoprotein and opsin genes in Mnemiopsis leidyi: a new view of ctenophore photocytes
2012
[73]
논문
Depth- and temperature-specific fatty acid adaptations in ctenophores from extreme habitats
2021-11-01
[74]
논문
First record of a ctenophore in lakes: The comb-jelly ''Mnemiopsis leidyi'' ({{small|A. Agassiz, 1865}}) invades the Fayum, Egypt
2016
[75]
논문
Molecular and morphological confirmation of an invasive American isopod; ''Livoneca redmanii'' ({{small|Leach, 1818}}), from the Mediterranean region to Lake Qaroun, Egypt
https://journals.ekb[...]
2019
[76]
논문
Laboratory studies of ingestion and food utilization in lobate and tentaculate ctenophores 1: Ctenophore food utilization
1978-07
[77]
논문
Comparative feeding behavior of planktonic ctenophores
2007-12
[78]
논문
Planktonic feeding and evolutionary significance of the lobate body plan within the Ctenophora
1998-10
[79]
논문
Herbivorous fishes ''Siganus rivulatus'' (Siganidae) and ''Zebrasoma desjardinii'' (Acanthuridae) feed on Ctenophora and Scyphozoa in the Red Sea
2016
[80]
논문
Predation on pelagic coelenterates: A review
2005-06
[81]
논문
Estimating the predatory impact of gelatinous zooplankton
1995-07
[82]
논문
Primary production of the biosphere: Integrating terrestrial and oceanic components
http://www.escholars[...]
1998-07-10
[83]
논문
The ctenophore ''Mnemiopsis'' in native and exotic habitats: U.S. estuaries versus the Black Sea basin
2001
[84]
논문
Invasion dynamics of the alien ctenophore ''Mnemiopsis leidyi'' and its impact on anchovy collapse in the Black Sea
2008-12
[85]
서적
Preliminary investigation on the molecular systematics of the invasive ctenophore ''Beroe ovata''
Springer Netherlands
2004
[86]
서적
Effects of ''Beroe'' c.f. ''ovata'' on gelatinous and other zooplankton along the Bulgarian Black Sea Coast
Springer Netherlands
2006
[87]
뉴스
Comb jelly nseurons spark evolution debate
https://www.quantama[...]
2015-06-12
[88]
논문
The Cambrian "explosion" of metazoans and molecular biology: Would Darwin be satisfied?
http://www.ijdb.ehu.[...]
2009-02-14
[89]
논문
A late-surviving stem-ctenophore from the Late Devonian of Miguasha (Canada)
2021
[90]
논문
Eoandromeda and the origin of Ctenophora
2011-09-20
[91]
뉴스
Ancient sea jelly shakes evolutionary tree of animals
https://www.scientif[...]
2018-06-21
[92]
논문
Lower Cambrian vendobionts from China and early diploblast evolution
2006-05-05
[93]
논문
Cambrian sessile, suspension feeding stem-group Ctenophores and evolution of the comb jelly body plan
2019-04
[94]
논문
Tentacular nature of the 'column' of the Cambrian diploblastic ''Xianguangia sinica''
2023-01
[95]
웹사이트
Ancient jellies had spiny skeletons, no tentacles
https://www.livescie[...]
2015-07-10
[96]
서적
The Early Evolution of Metazoa and the Significance of Problematic Taxa
Cambridge University Press
1991
[97]
논문
Cladistic analyses of the animal kingdom
1996-04
[98]
서적
Ueber die Morphologie und die Verwandtschaftsverhältnisse der wirbellosen thiere. Ein Beitrag zur Charakteristik und Classification der thierischen Formen
1923
[99]
서적
Systematische Phylogenie der Wirbellosen Thiere
1896
[100]
서적
The Invertebrates
McGraw Hill
1940
[101]
논문
Phylogenomics Revives Traditional Views on Deep Animal Relationships
2009-04-28
[102]
논문
The phylogenetic position of the comb jellies (Ctenophora) and the importance of taxonomic sampling
2004-12
[103]
논문
Phylogeny of Medusozoa and the evolution of cnidarian life cycles
2002
[104]
논문
Improved phylogenomic taxon sampling noticeably affects non-bilaterian relationships
2010-09
[105]
논문
Assessing the root of bilaterian animals with scalable phylogenomic methods
2009-12-22
[106]
논문
The homeodomain complement of the ctenophore ''Mnemiopsis leidyi'' suggests that Ctenophora and Porifera diverged prior to the ParaHoxozoa
[107]
논문
Nuclear receptors from the ctenophore ''Mnemiopsis leidyi'' lack a zinc-finger DNA-binding domain: Lineage-specific loss or ancestral condition in the emergence of the nuclear receptor superfamily?
[108]
논문
Genomic insights into Wnt signaling in an early diverging metazoan, the ctenophore ''Mnemiopsis leidyi''
[109]
논문
Evolution of sodium channels predates the origin of nervous systems in animals
[110]
논문
The Genome of the Ctenophore ''Mnemiopsis leidyi'' and its Implications for Cell Type Evolution
2013-12-13
[111]
논문
Error, signal, and the placement of Ctenophora sister to all other animals
2015-05-05
[112]
논문
Extracting phylogenetic signal and accounting for bias in whole-genome data sets supports the Ctenophora as sister to remaining Metazoa
2015-12-01
[113]
논문
Phylogenomics revives traditional views on deep animal relationships
2009-04-01
[114]
논문
Deep metazoan phylogeny: When different genes tell different stories
2013-04-01
[115]
논문
Genomic data do not support comb jellies as the sister group to all other animals
2015-12-15
[116]
논문
Topology-dependent asymmetry in systematic errors affects phylogenetic placement of Ctenophora and Xenacoelomorpha
2020-12-11
[117]
논문
Hidden diversity of Ctenophora revealed by new mitochondrial COI primers and sequences
[118]
논문
Rapid evolution of the compact and unusual mitochondrial genome in the ctenophore, ''Pleurobrachia bachei''
[119]
논문
Evolutionary conservation of the antimicrobial function of mucus: A first defence against infection
2018-07-04
[120]
보도자료
Into the brain of comb jellies: Scientists explore the evolution of neurons
https://neuroscience[...]
Okinawa Institute of Science and Technology
2022-08-08
[120]
논문
Mass spectrometry of short peptides reveals common features of metazoan peptidergic neurons
2022-10-01
[121]
논문
The last common ancestor of animals lacked the HIF pathway and respired in low-oxygen environments
[122]
논문
Conserved biophysical features of the CaV2 presynaptic Ca2+ channel homologue from the early-diverging animal Trichoplax adhaerens
2020-12-25
[123]
presentation slides
Evolution and Development
http://emb.carnegies[...]
Carnagie Science
2024-10-08
[124]
논문
Hox genes pattern the anterior-posterior axis of the juvenile but not the larva in a maximally indirect developing invertebrate, Micrura alaskensis (Nemertea)
[125]
논문
Hox gene expression during the development of the phoronid Phoronopsis harmeri
[126]
웹사이트
Independent Innexin Radiation Shaped Signaling in Ctenophores
https://academic.oup[...]
[127]
서적
The Origins and Relationships of Lower Invertebrates
https://archive.org/[...]
Clarendon Press
[128]
논문
A molecular phylogenetic framework for the phylum Ctenophora using 18S rRNA genes
2001-11-01
[129]
논문
Raman spectra of a Lower Cambrian ctenophore embryo from southwestern Shaanxi, China
2007-04-01
[130]
논문
The first fossil ctenophore from the Lower Devonian of West Germany
1983-06-09
[131]
논문
Middle Cambrian Ctenophores from the Stephen Formation, British Columbia, Canada
1996-03-29
[132]
논문
Eoandromeda and the origin of Ctenophora
2011-09-20
[133]
문서
藤田10
[134]
문서
発生の時、2細胞期に割球を分離するとクシ板が4列のものが2匹になり、また、4細胞期に割球を分離するとクシ板が2列のものが4匹になる。
[135]
논문
The development of radial and biradial symmetry: The evolution of bilaterality
http://icb.oxfordjou[...]
[136]
웹사이트
Phylum Ctenophora: list of all valid species names.
https://faculty.wash[...]
[137]
논문
Middle Cambrian Ctenophores from the Stephen Formation, British Columbia, Canada
The Royal Society
1996-03-01
[138]
논문
The Cambrian "explosion" of metazoans and molecular biology: would Darwin be satisfied?
http://www.ijdb.ehu.[...]
[139]
논문
Lower Cambrian Vendobionts from China and Early Diploblast Evolution
http://kasprzyk.home[...]
[140]
서적
The Early Evolution of Metazoa and the Significance of Problematic Taxa
[141]
저널
Cladistic analyses of the animal kingdom
[142]
서적
Ueber die Morphologie und die Verwandtschaftsverhältnisse der wirbellosen thiere. Ein Beitrag zur Charakteristik und Classification der thierischen Formen
1923
[143]
서적
Systematische Phylogenie Der Wirbellosen Thiere, Invertebrata, Part 2: Des Entwurfs Einer Systematischen Stammesgeschichte
1896
[144]
서적
The Invertebrates: Volume I, Protozoa Through Ctenophra
McGraw Hill
1940
[145]
저널
Phylogenomics revives traditional views on deep animal relationships
[146]
저널
The phylogenetic position of the comb jellies (Ctenophora) and the importance of taxonomic sampling
[147]
저널
Phylogeny of Medusozoa and the evolution of cnidarian life cycles
[148]
저널
Improved phylogenomic taxon sampling noticeably affects nonbilaterian relationships.
[149]
저널
Broad phylogenomic sampling improves resolution of the animal tree of life.
[150]
저널
Assessing the root of bilaterian animals with scalable phylogenomic methods.
[151]
저널
The homeodomain complement of the ctenophore Mnemiopsis leidyi suggests that Ctenophora and Porifera diverged prior to the ParaHoxozoa.
[152]
서적
The Origins and Relationships of Lower Invertebrates
Clarendon Press
[153]
저널
A Molecular Phylogenetic Framework for the Phylum Ctenophora Using 18S rRNA Genes
http://www.lifesci.u[...]
Academic Press
2009-02-14
[154]
저널
The Genome of the Ctenophore Mnemiopsis leidyi and Its Implications for Cell Type Evolution
2013-12-13
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com