해면동물
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
해면동물은 고착성 다세포 동물로, 편모세포를 통해 물을 흡수하고 배출하는 독특한 구조를 가지고 있다. 몸은 중교질로 구성되며, 내부에는 다양한 세포들이 존재한다. 해면동물은 무성 생식과 유성 생식을 모두 하며, 생존 기간은 수 년에서 최대 200년 이상이다. 해면동물의 계통 발생은 논쟁의 대상이었으나, 현재는 빗해파리류가 가장 기저적인 동물일 가능성이 높으며, 해면동물은 다른 모든 동물의 자매군으로 여겨진다. 해면동물은 스펀지, 의약품 개발, 수질 정화 등 다양한 분야에서 활용되며, 문화적으로도 네모바지 스폰지밥과 같은 캐릭터의 모티브가 되기도 한다.
더 읽어볼만한 페이지
- 해면동물 - 보라해면
보라해면은 한국 바닷가에 흔하게 분포하는 보라색 해면으로, 조간대 바위 밑이나 옆에 서식하며 몸의 일부분으로 재생이 가능하다. - 해면동물 - 목욕해면
목욕해면(Spongia officinalis)은 지중해 등 열대 및 아열대 해역에 분포하며, 고대부터 목욕 및 청소 용도로 사용되었고 현재는 세척 용도와 오락 목적으로 활용되는 해면 동물이다. - 생물 분류에 관한 - 다람쥐
다람쥐는 등 쪽에 줄무늬가 있는 다람쥐속 설치류로, 홀로 생활하며 겨울잠을 자고 씨앗, 견과류, 곤충 등을 먹으며 맹금류 등의 먹이가 되고, 특히 한반도 서식 다람쥐는 새로운 종으로 분류될 가능성이 있으며, 유럽에서는 침입 외래종으로 지정되기도 하고 라임병을 옮길 수 있어 주의가 필요한 동물이다. - 생물 분류에 관한 - 황금랑구르
황금랑구르는 인도 아삼 주와 부탄에 분포하며 크림색에서 황금색 털을 가진 멸종위기종 영장류로, 서식지 파괴로 개체수가 감소하여 보호받고 있다. - 글로벌세계대백과를 인용한 문서/{{{분류 - 공 (악기)
공은 금속으로 제작된 타악기로, 다양한 문화권에서 의식, 신호, 음악 연주 등에 사용되며, 형태와 용도에 따라 여러 종류로 나뉜다. - 글로벌세계대백과를 인용한 문서/{{{분류 - 국무회의
국무회의는 대한민국 대통령을 의장으로, 예산, 법률안, 외교, 군사 등 국정 현안을 심의하는 중요한 기관이며, 대통령, 국무총리, 국무위원으로 구성되고, 정례회의는 매주 1회, 임시회의는 필요에 따라 소집된다.

2. 생물학적 특징
해면동물은 대부분 물속의 박테리아 및 기타 미세한 먹이를 섭취하지만, 일부는 광합성 미생물을 내생공생체로 기주하며, 이러한 공생 관계는 종종 소비량보다 더 많은 음식과 산소를 생산한다.[13] 먹이가 부족한 환경에 서식하는 소수의 해면 종은 주로 작은 갑각류를 포식하는 육식 동물로 진화했다.[13]
대부분의 해면동물은 입수공을 통해 들어온 물에서 박테리아와 같은 미세한 먹이 입자를 걸러 섭취한다. 50μm(마이크로미터)보다 큰 입자는 입수공에 들어갈 수 없으며 피나코사이트가 식세포작용을 통해 섭취한다. 0.5μm~50μm의 입자는 입수공에 포획되어 피나코사이트 또는 아르케오사이트에 의해 섭취된다. 0.5μm 미만의 박테리아 크기의 입자는 편모세포에 의해 포획되고 섭취되는데, 편모세포는 일반적으로 해면동물 먹이의 80%를 포획한다.[27] 아르케오사이트는 소포에 포장된 먹이를 소화하지 않는 세포로 운반하며, 일부 해면동물은 아르케오사이트가 영양분을 운반하는 통로 역할을 하는 내부 섬유를 가지고 있다.[17]
유리해면은 해수에 용해된 영양분을 이용하여 생존하며 실트에 매우 민감하다는 주장이 있었으나, 2007년 연구에서는 증거가 없음을 발견했다. 유리해면은 물에서 박테리아와 기타 미생물을 매우 효율적으로(약 79%) 추출하고 부유 퇴적물 입자를 처리하여 먹이를 추출한다.[29]
해면동물의 세포는 물이 몸을 통과함에 따라 확산에 의해 산소를 흡수하며, 이산화탄소 및 암모니아와 같은 수용성 노폐물도 확산된다. 아르케오사이트는 입수공을 막을 위험이 있는 미네랄 입자를 제거하여 중교질을 통해 운반하고 배출되는 물 흐름으로 배출하지만, 일부 종은 이를 골격에 통합한다.[17]
일부 해면 종은 육식성을 띠는데, 먹이 입자가 매우 부족한 수역에서 갑각류와 기타 작은 동물을 잡아먹는다. 지금까지 137종만 발견되었으며,[31] 대부분 과 Cladorhizidae에 속하지만, Guitarridae와 Esperiopsidae의 일부 구성원도 육식성이다.[34] 대부분의 육식성 해면동물은 심해에 서식하며,[33] 심해 탐사 기술의 발전으로 더 많은 종이 발견될 것으로 예상된다.[17][34] 한 종은 얕은 깊이의 지중해 동굴에서 일반적인 여과 섭식 해면과 함께 발견되었는데, 가는 실로 1mm 미만의 갑각류를 얽어서 며칠에 걸쳐 소화한 후 정상 모양으로 돌아간다.[33] 대부분의 알려진 육식성 해면동물은 물 흐름 시스템과 편모세포를 완전히 상실했다. 그러나 속 ''Chondrocladia''는 먹이를 포획하는 데 사용되는 풍선 모양의 구조를 부풀리기 위해 고도로 수정된 물 흐름 시스템을 사용한다.[34][35]
담수 해면동물은 종종 아케오사이트 및 다른 세포 내에 내생공생체로서 녹조류를 숙주로 삼고, 조류가 생산하는 영양분으로부터 이익을 얻는다. 많은 해양 종은 다른 광합성 생물을 숙주로 삼는데, 가장 일반적인 것은 시아노박테리아이지만, 어떤 경우에는 디노플라젤라류이기도 하다. 공생하는 시아노박테리아는 일부 해면의 전체 생체 조직 질량의 3분의 1을 차지할 수 있으며, 일부 해면은 이러한 미생물로부터 에너지 공급의 48%~80%를 얻는다.[17] 슈투트가르트 대학교의 2008년 연구에서는 실리카로 만들어진 침골이 광합성 내생공생체가 서식하는 중교질층으로 빛을 전달한다고 보고했다.[36] 광합성 생물을 숙주로 삼는 해면은 먹이 입자가 비교적 부족한 해역에서 가장 흔하며, 종종 햇빛을 최대한 많이 모을 수 있는 잎 모양을 하고 있다.[82]
열수분출공 근처에 서식하는 육식성 해면은 메탄영양 박테리아를 숙주로 삼고 일부를 소화하기도 한다.[82]
해면동물은 면역계가 없지만, 다른 종으로부터의 이식은 거부한다. 일부 해양 종에서는 회색 세포가 이물질 거부 반응에서 주요 역할을 하며, 침입을 받으면 화학 물질을 생성하여 침입자가 해면의 내부 수송 시스템을 이용하지 못하게 한다. 침입이 지속되면 회색 세포는 그 부위에 집중되어 독소를 방출한다.[82]
대부분의 해면동물은 해산이며, 조간대에서 심해까지 다양한 서식지를 가진다. 담수산 종도 적지만 존재한다. 암반, 해조류, 조가비 등 단단한 기반 위에 붙어서 성장하는 것이 많지만, 심해의 진흙 바닥에는 뿌리 모양의 구조로 고착하는 것도 있다.
해면은 여과섭식자이며, 체내를 통과하는 물에서 유기물 미립자나 미생물을 잡아 영양으로 삼는다.
해면의 체내에는 많은 미생물이 공생하고 있으며, 종에 따라서는 전체 부피의 40%를 미생물이 차지한다.[132] 그 대부분은 해면 체내에서만 발견되는 종이다.
2. 1. 구조
해면동물은 다세포이며, 종속영양이고, 세포벽이 없으며, 정자를 생성한다는 점에서 다른 동물과 유사하다.[10] 하지만 다른 동물과 달리 진정한 조직[11]과 기관[11]이 없다. 일부는 방사대칭이지만, 대부분은 비대칭이다. 해면동물의 몸 형태는 중앙 공동을 통한 물 흐름의 효율을 극대화하도록 적응되어 있는데, 이 공동에서 물은 영양분을 공급하고, 그런 다음 출수공이라고 하는 구멍을 통해 빠져나간다.
많은 해면동물은 규질골편(탄산칼슘 또는 이산화규소의 골격과 같은 조각) 및/또는 스폰진(변형된 콜라겐 단백질의 한 종류)[10]으로 된 내골격을 가지고 있다. 중교질이라는 내부의 젤라틴성 기질은 내골격 역할을 하며, 암석과 같은 단단한 표면에 부착하는 연한 해면에서 유일한 골격이다. 일반적으로 중교질은 광물질 골편, 스폰진 섬유 또는 둘 다에 의해 단단해진다.
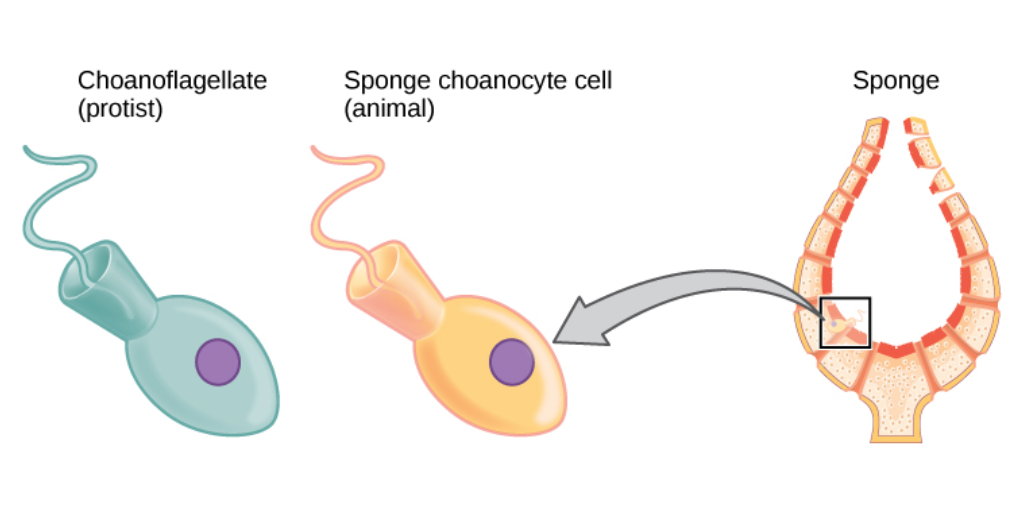
자포동물(해파리 등)과 빗해파리와 마찬가지로, 그리고 다른 모든 알려진 후생동물과 달리, 해면동물의 몸은 두 개의 주요 세포층 사이에 끼인 비생체 젤리 같은 물질인 중교질로 구성된다.[82][19] 해면동물은 척추동물과 같은 신경계는 없지만, 전혀 다른 신경계를 가질 수 있다.[7]
해면동물의 몸은 속이 비어 있으며, 주로 콜라겐으로 이루어진 젤리 같은 물질인 중교질과 콜라겐으로 만들어진 치밀한 섬유 네트워크에 의해 모양이 유지된다. 18가지의 서로 다른 세포 유형이 확인되었다.[22] 내부 표면은 편모가 하나씩 있는 원통형 또는 원추형의 칼라(collar)로 둘러싸인 편모세포로 덮여 있다. 채찍 모양의 편모의 파동 운동은 해면동물의 몸을 통해 물을 이동시킨다. 모든 해면동물은 중교질을 통해 내부로 이어지는 통로인 '''입수공(ostia)'''을 가지고 있으며, 대부분의 해면동물에서 이는 닫을 수 있는 입구 밸브를 형성하는 관 모양의 세관세포에 의해 조절된다. 판 모양의 세포인 피나코사이트는 편모세포로 덮여 있지 않은 중교질의 다른 모든 부분을 덮는 단층의 외부 피부를 형성하며, 피나코사이트는 입수공으로 들어가기에는 너무 큰 음식 입자도 소화한다.[17][82] 동물의 기저부에 있는 피나코사이트는 해면동물을 고정하는 역할을 한다.[82]
중교질 내에는 다음과 같은 다양한 유형의 세포들이 살고 이동한다.[17][82]
- 로포사이트는 중교질을 통해 천천히 이동하며 콜라겐 섬유를 분비하는 아메바와 같은 세포이다.
- 콜렌사이트는 또 다른 유형의 콜라겐 생성 세포이다.
- 섬유세포는 중교질의 일부를 형성하는 다당류를 분비한다.
- 난세포와 정자세포는 생식 세포이다.
- 골편세포는 많은 해면동물의 골격을 형성하고 일부 종에서는 포식자에 대한 방어를 제공하는 광물화된 골편("작은 가시")을 분비한다.
- 골편세포 대신 또는 골편세포와 함께, 데모스폰지는 콜라겐의 한 형태를 분비하는 해면세포를 가지고 있는데, 이것은 폴리머화되어 중교질을 굳게 하는 두꺼운 섬유질 물질인 스폰진을 형성한다.
- 근세포("근육 세포")는 신호를 전달하고 동물의 일부가 수축하게 한다.
- "회색 세포"는 해면동물의 면역계에 해당하는 역할을 한다.
- 원세포(또는 아메바세포)는 전능성을 가진 아메바와 같은 세포이며, 다시 말해 각 세포는 다른 어떤 유형의 세포로도 변형될 수 있다. 또한 먹이를 먹는 데 중요한 역할을 하며 입수공을 막는 잔해를 제거하는 역할도 한다.
유리해면은 이 기본적인 계획에 독특한 변형을 보여준다. 실리카로 만들어진 골편은 막대 사이에 생체 조직이 거미줄처럼 매달려 있는 발판과 같은 틀을 형성하며, 대부분의 세포 유형이 이 안에 들어 있다.[17] 이 조직은 어떤 면에서는 단일 외부 세포막을 공유하는 많은 세포처럼 행동하고, 다른 면에서는 여러 세포핵을 가진 단일 세포처럼 행동하는 합포체이다.
대부분의 해면은 굴뚝과 비슷하게 작동한다. 아래쪽에서 물을 흡입하여 위쪽의 출수공으로 배출한다. 주변의 물 흐름은 위쪽이 더 빠르기 때문에 베르누이 원리에 의해 발생하는 흡입 효과가 일부 작업을 무료로 수행한다. 해면은 출수공과 입수공(흡입공)을 완전히 또는 부분적으로 닫고 편모의 박동을 다양하게 조절함으로써 물 흐름을 조절할 수 있으며, 물에 모래나 실트가 많으면 물 흐름을 멈출 수 있다.[17]
피나코사이트와 편모세포의 층은 더 복잡한 동물의 상피와 유사하지만, 세포 간 연결이나 기저판(아래쪽의 얇은 섬유질 시트)으로 단단히 결합되어 있지 않다. 이러한 층의 유연성과 섬모세포에 의한 중교질층의 재구성을 통해 동물은 평생 동안 자신의 형태를 조절하여 지역의 물 흐름을 최대한 활용할 수 있다.[17]
해면에서 가장 단순한 몸 구조는 "아스콘형"으로 알려진 관 또는 꽃병 모양이지만, 이는 동물의 크기를 심각하게 제한한다. 몸 구조는 편모세포의 단일 층으로 둘러싸인 줄기 모양의 해면강으로 특징지어집니다. 단순히 크기를 키우면 부피 대 표면적의 비율이 증가하는데, 표면적은 길이 또는 너비의 제곱에 비례하여 증가하는 반면 부피는 세제곱에 비례하여 증가하기 때문이다. 음식과 산소가 필요한 조직의 양은 부피에 따라 결정되지만, 음식과 산소를 공급하는 펌핑 용량은 편모세포가 덮인 면적에 따라 달라진다. 아스콘형 해면은 직경이 1mm를 넘는 경우가 거의 없다.[17]

일부 해면은 몸벽이 주름진 "사이콘형" 구조를 채택하여 이러한 제한을 극복한다. 주름의 안쪽 주머니에는 편모세포가 늘어서 있으며, 이는 입수공을 통해 주름의 바깥쪽 주머니에 연결된다. 편모세포 수의 증가, 즉 펌핑 용량의 증가로 인해 사이콘형 해면은 직경이 수 센티미터까지 자랄 수 있다.
"류콘형" 패턴은 편모세포가 늘어선 방들의 네트워크를 포함하고 서로 연결되며, 물 흡입구와 배출구에 관을 통해 연결된 중교질층으로 내부를 거의 완전히 채움으로써 펌핑 용량을 더욱 향상시킨다. 류콘형 해면은 직경이 1m를 넘게 자라며, 어떤 방향으로든 성장하면 편모세포 방의 수가 증가하기 때문에 더 넓은 범위의 형태를 취할 수 있다. 예를 들어, "피복형" 해면은 부착된 표면의 형태를 따른다. 모든 담수 해면과 대부분의 천해 해양 해면은 류콘형 몸체를 가지고 있다. 유리해면의 물 통로 네트워크는 류콘형 구조와 유사하다.[17]
세 가지 유형의 구조 모두에서 편모세포가 늘어선 영역의 단면적은 흡입 및 배출 채널의 단면적보다 훨씬 크다. 이는 편모세포 근처의 흐름을 느리게 하여 음식 입자를 포획하기 쉽게 만듭니다.[17] 예를 들어, 높이 약 10cm이고 직경이 1cm인 작은 류콘형 해면인 ''류코니아''에서는 분당 6cm의 속도로 8만 개가 넘는 흡입관으로 물이 들어간다. 그러나 ''류코니아''는 지름의 합이 관의 지름보다 훨씬 큰 200만 개가 넘는 편모가 있는 방을 가지고 있기 때문에 방을 통과하는 물의 흐름은 시간당 3.6cm로 느려져 편모세포가 음식을 쉽게 포획할 수 있다. 모든 물은 초당 약 8.5cm의 속도로 단일 출수공을 통해 배출되어 폐기물을 어느 정도 멀리 운반할 수 있을 만큼 충분히 빠르다.[25]
동물학에서 골격(skeleton)이란 관절이 있든 없든, 생체광물화(biomineralization)되었든 아니든, 동물의 어느 정도 단단한 구조를 말한다. 대부분의 해면동물에서 중교질(mesohyl)은 내골격(endoskeleton) 역할을 하며, 암석과 같은 단단한 표면에 부착하는 연한 해면동물에서는 유일한 골격이다. 보다 일반적으로 중교질은 광물질인 해면골편(sponge spicule)이나 스폰진 섬유, 또는 둘 다에 의해 단단해진다. 대부분의 종에서 발견되지만 모든 종에 있는 것은 아닌 골편(spicule)[26]은 이산화규소(silica) 또는 탄산칼슘으로 만들어지며, 모양은 단순한 막대기에서 최대 6개의 광선을 가진 3차원 "별" 모양까지 다양하다. 골편은 골세포(sclerocyte)에 의해 생성[17]되며, 분리되어 있거나 관절로 연결되거나 융합될 수 있다.[16]
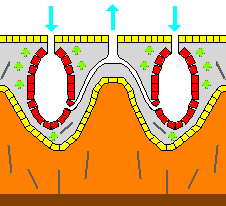
일부 해면동물은 유기 성분의 바깥쪽에 완전히 위치하는 외골격(exoskeleton)을 분비하기도 한다. 예를 들어, 각질해면(sclerosponge) ("딱딱한 해면")은 유기물이 광물질 내의 구멍에 편모세포(choanocyte)가 있는 얇은 층을 형성하는 거대한 탄산칼슘 외골격을 가지고 있다. 이러한 외골격은 동물의 피부를 형성하는 피복세포(pinacocyte)에 의해 분비된다.[17] 해면은 부착성 동물이다. 기본적으로 방사대칭 형태를 취하지만, 실제로는 환경에 따라 크게 변한다. 표면에는 소공이라고 하는 많은 구멍이 있으며, 여기에서 물과 먹이를 섭취한다. 또한, 대공이라고 하는 개구부가 상부에 있으며, 여기에서 물을 내뿜는다. 위강이라고 하는 내부의 공동부에는 편모를 가진 깃세포가 많이 있으며, 이 편모에 의해 소공에서 대공으로의 물 순환을 일으킨다. 이러한 체내에 물을 순환시키는 구조를 통틀어 수관계(canal system)라고 한다.
2. 2. 섭식
해면동물은 물을 통해 흐르는 물에서 먹이 입자를 걸러서 섭취한다. 50μm(마이크로미터)보다 큰 입자는 입수공에 들어갈 수 없으며 피나코사이트가 식세포작용을 통해 섭취한다. 0.5μm~50μm의 입자는 입수공에 포획되어 피나코사이트 또는 아르케오사이트에 의해 섭취된다. 0.5μm 미만의 박테리아 크기의 입자는 편모세포에 의해 포획되고 섭취되는데, 편모세포는 일반적으로 해면동물 먹이의 80%를 포획한다.[27] 아르케오사이트는 소포에 포장된 먹이를 소화하지 않는 세포로 운반하며, 일부 해면동물은 아르케오사이트가 영양분을 운반하는 통로 역할을 하는 내부 섬유를 가지고 있다.[17]유리해면은 해수에 용해된 영양분을 이용하여 생존하며 실트에 매우 민감하다는 주장이 있었으나, 2007년 연구에서는 증거가 없음을 발견했다. 유리해면은 물에서 박테리아와 기타 미생물을 매우 효율적으로(약 79%) 추출하고 부유 퇴적물 입자를 처리하여 먹이를 추출한다.[29] 칼라체는 먹이를 소화하고 소포에 싸서 다이네인 "모터" 분자에 의해 미세소관 다발을 따라 합포체 전체로 운반한다.[17]
해면동물의 세포는 물이 몸을 통과함에 따라 확산에 의해 산소를 흡수하며, 이산화탄소 및 암모니아와 같은 수용성 노폐물도 확산된다. 아르케오사이트는 입수공을 막을 위험이 있는 미네랄 입자를 제거하여 중교질을 통해 운반하고 배출되는 물 흐름으로 배출하지만, 일부 종은 이를 골격에 통합한다.[17]
먹이 입자가 매우 부족한 수역에서는 일부 종이 갑각류와 기타 작은 동물을 잡아먹는다. 지금까지 137종만 발견되었으며,[31] 대부분 과 Cladorhizidae에 속하지만, Guitarridae와 Esperiopsidae의 일부 구성원도 육식성이다.[34] 대부분의 육식성 해면동물은 심해에 서식하며,[33] 심해 탐사 기술의 발전으로 더 많은 종이 발견될 것으로 예상된다.[17][34] 한 종은 얕은 깊이의 지중해 동굴에서 일반적인 여과 섭식 해면과 함께 발견되었는데, 가는 실로 1mm 미만의 갑각류를 얽어서 며칠에 걸쳐 소화한 후 정상 모양으로 돌아간다.[33]
대부분의 알려진 육식성 해면동물은 물 흐름 시스템과 편모세포를 완전히 상실했다. 그러나 속 ''Chondrocladia''는 먹이를 포획하는 데 사용되는 풍선 모양의 구조를 부풀리기 위해 고도로 수정된 물 흐름 시스템을 사용한다.[34][35]
광합성을 하는 내생 공생체를 가진 해면동물은 소비하는 것보다 최대 세 배나 많은 산소를 생산하며, 소비하는 것보다 많은 유기물도 생산한다. 이러한 서식지 자원에 대한 기여는 오스트레일리아의 그레이트 배리어 리프에서는 상당하지만 카리브해에서는 비교적 미미하다.[27]
대부분의 해면동물은 퇴적물식동물로서 해수에서 유기 잔해 입자와 미생물을 걸러 먹는다. 특히 해면동물은 산호초 먹이 그물에서 퇴적물을 더 높은 영양 단계로 재활용함으로써 퇴적물식동물로서 중요한 역할을 한다.[72]
해면은 부착성 동물이며, 방사대칭 형태를 취하지만 환경에 따라 크게 변한다. 표면에는 소공이라고 하는 많은 구멍이 있으며, 여기에서 물과 먹이를 섭취한다. 또한, 대공이라고 하는 개구부가 상부에 있으며, 여기에서 물을 내뿜는다. 위강이라고 하는 내부의 공동부에는 편모를 가진 깃세포가 많이 있으며, 이 편모에 의해 소공에서 대공으로의 물 순환을 일으킨다. 이러한 체내에 물을 순환시키는 구조를 통틀어 수관계(canal system)라고 한다.
해면은 여과섭식자이며, 체내를 통과하는 물에서 유기물 미립자나 미생물을 잡아 영양으로 삼는다. 그것들은 깃세포나 내부의 식세포에 의해 확보된다. 그러나, 심해 등에는 육식성 해면이라고 불리는 것이 있으며, 그것들은 더 큰 소형 갑각류 등을 잡아 소화한다.
해면의 체내에는 많은 미생물이 공생하고 있으며, 종에 따라서는 전체 부피의 40%를 미생물이 차지한다.[132]
2. 3. 공생
해면동물은 녹조류와 같은 다양한 생물들과 공생 관계를 맺는다. 특히 담수 해면동물은 아케오사이트 및 다른 세포 내에 내생공생체로서 녹조류를 포함하고 있으며, 녹조류가 생산하는 영양분을 얻는다.[17] 많은 해양 종은 시아노박테리아나 디노플라젤라류와 같은 광합성 생물을 숙주로 삼는다. 공생하는 시아노박테리아는 일부 해면의 전체 생체 조직 질량의 3분의 1을 차지하며, 해면은 이 미생물로부터 에너지 공급의 48%~80%를 얻는다.[17] 2008년 슈투트가르트 대학교 연구에 따르면, 실리카로 만들어진 해면의 골편은 광합성 내생공생체가 서식하는 중교질층으로 빛을 전달한다.[36] 이러한 광합성 생물을 숙주로 삼는 해면은 먹이 입자가 비교적 부족한 해역에서 흔하며, 햇빛을 최대한 많이 모을 수 있는 잎 모양을 띠는 경우가 많다.[82]열수분출공 근처에 서식하는 육식성 해면은 메탄영양 박테리아를 숙주로 삼고 일부를 소화하기도 한다.[82] 해면동물은 다른 유기체와도 다양한 공생 관계를 맺는 것으로 알려져 있다. 예를 들어, 껍질 모양의 해면인 ''리소덴도릭스(Lissodendoryx) 콜롬비엔시스(colombiensis)''는 해초 해면에 둘러싸여 서식 범위를 넓히는데, 해초 해면은 불가사리에게 맛이 없어 ''리소덴도릭스''를 보호하고, 그 대가로 해초 해면은 해저 퇴적물에서 멀리 떨어진 높은 위치를 얻는다.[53]
''시날페우스(Synalpheus)'' 속 새우들은 해면 속에 군집을 이루며, 각 새우 종은 서로 다른 해면 종에 서식하여 ''시날페우스'' 속을 가장 다양한 갑각류 속 중 하나로 만든다. 특히, 시날페우스 레갈리스(Synalpheus regalis)는 해면을 먹이원으로 사용할 뿐만 아니라, 다른 새우와 포식자로부터 자신을 방어하는 데에도 사용한다.[54] 최대 16,000마리의 새우가 큰머리해면(loggerhead sponge)에 서식하며, 해면이 먹이를 얻기 위해 바닷물을 여과하는 과정에서 쌓이는 큰 입자를 먹고 산다.[55] 소라게와 같은 다른 갑각류들은 특정 해면 종인 ''슈도스폰고소라이테스(Pseudospongosorites)''를 가지고 공생하며, 게와 해면 모두 복족류 껍데기를 차지한다. 게와 해면이 껍데기를 벗어날 때까지, 게는 적절한 대체 껍데기를 찾을 때까지 해면의 몸을 껍데기 대신 보호 수단으로 사용한다.[56]
해면은 미생물 공동체와 공생하여 다양한 해면 미생물군을 형성할 수 있다. 해면은 매우 특이적인 미생물 군집을 숙주할 수 있으며, 이 미생물 군집은 숙주 생물량의 최대 35%를 차지할 수 있다.[75] 이러한 특정 공생 관계를 홀로바이오틱 관계라고 한다. 해면과 관련된 미생물 군집은 다양한 이차 대사산물을 생성하여 화학적 방어와 같은 메커니즘을 통해 포식자로부터 보호하는 데 도움을 준다.[76]
해면은 박테리아세포 내 공생체와 피나코더 세포층 아래에서 가장 많은 빛을 받아 광영양 작용에 이용하는 시아노박테리아 또는 미세조류를 포함하여 다양한 공생 관계를 가진다. 해면은 알파프로테오박테리아, 방선균문, 클로로플렉스문, 니트로스피로문, "시아노박테리아", 감마-분류군, 후보문 포리박테리아, 및 타우마르케아를 포함하여 50개 이상의 서로 다른 미생물 문과 후보 문을 숙주할 수 있다.[76]
해면의 체내에는 많은 미생물이 공생하고 있으며, 종에 따라서는 전체 부피의 40%를 미생물이 차지한다.[132] 그 대부분은 해면 체내에서만 발견되는 종이다. 현재, 세균과 고세균 모두에, Poribacteria(해면 Porifera에 따름)나 타움고세균(''Cenarchaeum symbiosum'' 등)과 같은 새로운 문이 제안되고 있다.[133]
3. 생식과 발생
해면동물은 무성생식과 유성생식으로 번식한다. 무성생식은 몸 일부가 떨어져 나가 새로운 개체가 되거나(출아법), '아포(芽胞)'라는 싹을 만들어 번식하는 방식이다. 유성생식의 경우, 대부분 자웅동체이지만, 생식 기관은 없다. 정자는 편모세포에서, 난자는 원세포나 편모세포가 변형되어 만들어진다. 정자는 물을 통해 다른 해면동물로 이동, 난자와 수정한다. 수정란은 물속에 방출되거나 어미 해면 안에서 유생으로 자란다.[17]
해면동물의 난자와 정자 형성 과정은 다른 동물들과 매우 유사하며, 감수분열에 필요한 유전자들도 가지고 있다. 이는 감수분열이 진핵생물 진화 초기에 나타났음을 시사한다.[42]
온대 지역 해면동물은 수명이 수년에 불과하지만, 열대나 심해 종은 200년 이상 살기도 한다. 일부 석회질 해면은 1년에 0.2mm만 자라므로, 1m 크기 해면은 5,000년 정도 된 것으로 추정된다.[17]
해면동물은 발생 과정에서 외배엽이나 내배엽 같은 배엽이 형성되지 않고, 뚜렷한 기관도 분화되지 않는다. 이 때문에 해면동물은 판형동물과 함께 측생동물로 분류되며, 기관계가 분화된 진정후생동물과는 구별된다. 과거에는 해면동물이 깃편모충이라는 단세포생물에서 유래하여 다른 다세포 동물과는 계통이 다르다는 주장도 있었지만, 해면동물에게도 다세포 생물의 발생에 관여하는 유전자들이 존재한다는 사실이 밝혀지면서 이러한 주장은 힘을 잃었다.
3. 1. 생식
해면동물은 무성생식과 유성생식을 모두 한다.무성생식 방법으로는 파편화, 출아법, 생아 생성이 있다. 해면의 파편은 해류나 파도에 의해 떨어져 나갈 수 있다. 해면은 피나코사이트와 편모세포의 이동성과 중교질층의 재구성을 이용하여 적절한 표면에 다시 부착한 후 며칠 안에 작지만 기능적인 해면으로 재생된다.[17] 고운 천으로 짜낸 해면도 같은 능력을 통해 재생될 수 있다.[17] 해면 파편은 결합세포가 중교질층을 생성하고 원세포가 다른 모든 세포 유형을 생성할 수 있을 때에만 재생될 수 있다.[27] 매우 적은 종만이 출아법으로 번식한다.[17]
생아는 일부 해양 해면과 많은 담수 해면 종이 죽을 때 수천 개씩 생성하는 "생존 캡슐"이며, 일부, 주로 담수 종은 가을에 정기적으로 생성한다. 해면세포는 스폰진으로 된 껍질을 만들고, 종종 침골로 강화하여 영양분으로 가득 찬 원세포 덩어리를 감싼다.[17] 담수 생아에는 광합성 공생체가 포함될 수도 있다.[37] 그런 다음 생아는 휴면 상태가 되어 추위, 건조, 산소 부족 및 염분의 극심한 변화를 견딜 수 있다.[17] 담수 생아는 종종 온도가 내려가고 몇 달 동안 차가운 상태를 유지한 후 거의 "정상" 수준에 도달해야 활성화된다.[37] 생아가 발아하면, 덩어리 바깥쪽의 원세포는 피나코사이트로 변형되고, 껍질의 구멍 위의 막이 터지고, 세포 덩어리가 서서히 나타나며, 남은 원세포의 대부분은 기능적인 해면을 만드는 데 필요한 다른 세포 유형으로 변형된다. 서로 다른 개체의 같은 종의 생아는 하나의 해면을 형성하기 위해 결합할 수 있다.[17] 일부 생아는 모 해면 내에 남아 있으며, 봄에는 오래된 해면이 부활했는지 아니면 자신의 생아에 의해 "재집락화"되었는지 구별하기 어려울 수 있다.[37] 체표에서 싹이 자라 번식하는 것 외에, 아포(芽球)라고 불리는 싹을 체외로 방출하여 번식하는 종도 있다.
유성생식의 경우, 대부분의 해면동물은 자웅동체(동시에 양성의 기능을 가짐)이지만, 생식 기관인 생식소는 없다. 정자는 편모세포 또는 중교질 내로 가라앉아 정자낭을 형성하는 전체 편모세포실에 의해 생성되는 반면, 난자는 일부 종에서는 원세포 또는 편모세포의 변형을 통해 형성된다. 각 난자는 일반적으로 "간호세포"를 섭취하여 난황을 얻는다. 산란 중에 정자는 낭에서 터져 나와 출수공을 통해 배출된다. 만약 동일 종의 다른 해면동물과 접촉하면, 물의 흐름에 의해 편모세포로 운반되어 둘러싸이지만, 소화하는 대신 아메바형태로 변형되어 중교질을 통해 난자로 정자를 운반한다. 대부분의 경우 난자는 운반체와 그 짐을 모두 둘러싼다.[17]
몇몇 종은 수정란을 물속에 방출하지만, 대부분은 부화할 때까지 난자를 보유한다. 난자를 보유함으로써 부모는 수직 전달을 통해 공생 미생물을 자손에게 직접 전달할 수 있는 반면, 난자를 물속에 방출하는 종은 수평적으로 공생체를 획득해야 한다(수직 전달된 공생체를 가진 유생이 수평적으로 다른 공생체를 획득하는 경우를 포함하여, 두 가지 방식의 조합이 가장 일반적일 것이다).[38][39] 네 가지 유형의 유생이 있지만, 모두 편모 또는 섬모의 바깥층 세포가 유생의 이동을 가능하게 하는 세포 덩어리인 난황영양성(비영양성) 유생이다. 며칠 동안 헤엄친 후, 유생은 가라앉아 정착할 장소를 찾을 때까지 기어 다닌다. 대부분의 세포는 원세포로 변형된 다음, 소형 성체 해면에서 위치에 적합한 유형으로 변형된다.[17][40]
유리해면 배아는 분리된 세포로 분열하여 시작하지만, 32개의 세포가 형성되면 빠르게 유생으로 변형된다. 외부적으로는 중앙을 따라 움직임에 사용하는 섬모띠가 있는 난형이지만, 내부적으로는 거미줄 모양의 주요 합포체가 그 주위와 사이에 드리워져 있고 중앙에 여러 개의 칼라체가 있는 편모합포체가 있는 골편의 전형적인 유리해면 구조를 가지고 있다. 그런 다음 유생은 부모의 몸을 떠난다.[41]
해면동물의 난자형성(난자형성)과 정자형성(정자형성)(생식세포형성(생식세포형성))의 세포학적 과정은 다른 후생동물과 매우 유사하다.[42] DNA 재조합 및 이중 가닥 절단 복구 유전자를 포함한 고전적인 감수분열(감수분열) 유전자 세트의 대부분 유전자는 진핵생물(진핵생물)에서 보존되어 있으며, 해면동물(예: ''Geodia hentscheli'' 및 ''Geodia phlegraei'')에서 발현된다.[42] 해면동물은 가장 초기 분기된 동물로 간주되기 때문에, 이러한 발견은 재조합 및 DNA 복구 능력을 포함한 감수분열의 기본 도구가 진핵생물 진화 초기에 존재했음을 시사한다.[42] 유성생식 또한 다양하며, 자웅동체와 자웅이체의 양쪽 종류가 있다. 많은 종에서는 수정 후, 유생이 될 때까지 부모의 몸속에서 자라는 태생이지만, 난생인 종도 존재한다.
3. 2. 발생
온대 지역의 해면동물은 수년 정도 살지만, 일부 열대 종이나 심해 종은 200년 이상 살 수 있다. 일부 석회질 해면(demosponge)은 연간 0.2mm 정도만 자라며, 그 속도가 일정하다면 1m 너비의 표본은 약 5,000년 된 것이다. 어떤 해면은 수 주 만에 유성 생식을 시작하지만, 다른 해면은 수년이 될 때까지 기다린다.[17]발생 과정에서 외배엽이나 내배엽과 같은 배엽 형성은 일어나지 않는다. 또한, 다세포생물이면서도 명확한 기관 분화가 보이지 않는다. 이 때문에 해면동물문과 판형동물문(마찬가지로 조직 분화가 보이지 않는다)은 측생동물(Parazoa)로 분류되며, 기관계가 분화한 다른 동물인 진정후생동물(Eumetazoa)과 구별된다. 깃세포가 깃으로 둘러싸인 편모를 가지고 있다는 점에서 단세포생물인 깃편모충과 해면동물의 유사성이 오래전부터 지적되어 왔다. 이러한 점으로 미루어 보아, 과거에는 해면동물이 깃편모충의 군체로부터 유래하여 다세포 동물과는 계통이 다르다는 주장도 있었다. 그러나 해면동물에는 호메오박스 유전자, TGB-β 유전자 등 다세포 생물로서의 분화, 발생에 관여하는 유전자군이 이미 존재한다는 것이 밝혀지고 있으며, 다른 다세포 동물과의 차이는 그렇게 크지 않다는 증거도 제시되고 있다.
4. 진화와 계통
해면동물은 분자시계와 생체지표 연구를 통해 캄브리아기 대폭발 이전부터 이미 존재했음이 시사된다. 하지만 규산 침골을 가진 데모스폰지는 캄브리아기까지 화석 기록에 나타나지 않는다.[88] 7억 5천만 년 전 암석에서 발견되었다는 보고[89]는 검증되지 않았다.
에디아카라기 더우샨투오층(약 5억 8천만 년 전)에서는 침골, 피나코사이트, 포로사이트, 아르케오사이트, 클레로사이트, 내부 공동을 포함한 데모스폰지 화석이 발견되었다.[90] 유리해면 화석은 오스트레일리아, 중국, 몽골(약 5억 4천만 년 전)에서,[91] 초기 캄브리아기 해면동물 속인 ''Kiwetinokia''는 멕시코에서 발견되었다.[92] 석회해면의 탄산칼슘 침골은 오스트레일리아의 초기 캄브리아기 암석(약 5억 3천만 년 전)에서 발견되었고, 다른 데모스폰지들은 초기 캄브리아기 청장 생물군(약 5억 2천 5백만 년 전)에서 발견되었다.[93] 캐나다 노스웨스트 준주에서는 8억 9천만 년 전 해면동물 화석이 발견되었는데, 이는 최초의 동물이 신원생대 산소 대사 사건 이전에 나타났음을 시사한다.[94]
담수 해면은 에오세 중기(약 4천 8백만 년 전) 지층에서 가장 오래된 화석이 발견되어 비교적 최근에 나타난 것으로 보인다.[91] 현대 해면의 약 90%가 데모스폰지이지만, 이들의 골격은 부드러운 스폰진으로 구성되어 화석화가 잘 되지 않아 다른 종류보다 화석 유해가 드물다.[95] 가장 오래된 해면 공생 생물은 실루리아기 초기 랜드로베리절 지층에서 알려져 있다.[96]
화학적 추적자인 24-이소프로필 콜레스탄은 데모스폰지에서 생성되지만 진정동물에서는 생성되지 않는다고 알려져 있다.[97] 이전에는 18억 년 전 암석에서 이 물질의 흔적이 보고되었지만,[98] 최근 연구에 따르면 약 6억 3천 5백만 년 전 마리노아 빙하기 말 이후에만 나타난다.[100][101][102]
아르케오시아타는 초기 캄브리아기 암석에서 흔하게 발견되지만 캄브리아기 말(약 4억 9천만 년 전)에 멸종된 것으로 보이는[93] 해면, 자포동물, 조류, 유공충 등 다양한 생물로 분류되어 왔으나, 1990년대 이후로는 독특한 해면 그룹으로 간주되고 있다.[103]
찬셀로리아는 해면과의 관련성이 제기되기도 했으나,[104] 피부 구조 차이 등으로 인해 해면과 더 복잡한 동물의 중간일 수 있다는 주장도 있다.[105] 2008년 연구에서는 할키에리아와의 유사성이 제기되었으나,[106] 딜레마가 존재한다.
해면동물은 발생 과정에서 외배엽이나 내배엽과 같은 배엽 형성이 일어나지 않고, 명확한 기관 분화도 보이지 않아 측생동물로 분류된다.[134] 깃편모충과의 유사성으로 인해 깃편모충 군체에서 유래했다는 주장도 있었으나, 호메오박스 유전자 등 다세포 생물로서의 분화, 발생에 관여하는 유전자군이 이미 존재하여 다른 다세포 동물과의 차이가 크지 않다는 증거도 제시되고 있다.
4. 1. 후생동물과의 계통 관계
해면동물의 계통 발생은 계통 발생학의 등장 이후로 많은 논쟁이 있어 왔다. 원래 가장 기저적인 동물 문으로 여겨졌으나, 현재로서는 빗해파리가 그 자리를 차지할 가능성이 높다는 상당한 증거가 있다.[84][85] 또한, 해면동물 문의 단계통성 또한 의문시되고 있다. 여러 연구들은 다른 모든 동물인 후생동물이 해면동물 내부에서 갈라져 나왔다는 결론을 내렸고, 일반적으로 석회해면류와 동질골격류가 유해면류보다 다른 동물들과 더 가깝다는 결과를 얻었다.[86][87] 해면동물 내부의 계통 관계는 불확실성이 적은 편이다. 해면동물의 단계통성을 지지하든 그렇지 않든 거의 모든 연구에서 동질골격류와 석회해면류의 가까운 관계가 밝혀졌다. 유리해면류의 위치 또한 상당히 확실하며, 대부분의 연구에서 유리해면류가 유해면류의 자매군으로 나타난다.1990년대에는 해면동물이 모두 단계통군(monophyletic group)으로 여겨졌다. 즉, 모든 해면동물은 스스로 해면동물인 공통 조상(last universal common ancestor)에서 유래했고, 다른 모든 후생동물(다세포 동물)의 "자매군"(sister-group)으로 여겨졌다. 후생동물 자체는 단계통군을 형성한다. 반면, 1990년대 일부 분석에서는 동물의 가장 가까운 진화적 친척이 깃편모충이라는 생각을 되살렸다. 깃편모충은 해면동물의 편모세포와 매우 유사한 단세포 유기체이다. 이는 대부분의 후생동물이 해면과 매우 유사한 조상에서 진화했고, 따라서 해면동물이 단계통군이 아닐 수도 있음을 시사한다. 왜냐하면 동일한 해면과 같은 조상이 현대 해면과 비해면 후생동물 모두에게 기원이 될 수 있기 때문이다.[107]
2001년 이후의 분석은 진후생동물(해면보다 복잡한 동물)이 특정 해면동물 그룹과 다른 해면동물 그룹보다 더 밀접하게 관련되어 있다는 결론을 내렸다. 이러한 결론은 모든 해면동물의 최종 공통 조상이 해면동물이 아닌 진후생동물의 직계 조상이기도 하기 때문에 해면동물이 단계통군이 아니라는 것을 의미한다. 리보솜 DNA 비교를 기반으로 한 2001년 연구는 해면동물 내에서 가장 기본적인 분류는 육방해면류와 나머지 사이이며, 진후생동물은 다른 유형의 해면보다 석회해면류(탄산칼슘 골편을 가진 해면)와 더 밀접하게 관련되어 있다는 결론을 내렸다.[107] 2007년에는 RNA 비교를 기반으로 한 분석과 주로 골편 비교를 기반으로 한 또 다른 분석에서 해면류와 육방해면류가 석회해면류보다 서로 더 밀접하게 관련되어 있으며, 석회해면류는 진후생동물과 더 밀접하게 관련되어 있다는 결론을 내렸다.[91][108]
다른 해부학적 및 생화학적 증거는 진후생동물을 해면류의 하위 그룹인 동질모르파와 연결한다. 2007년 육방해면류와 빗해파리류를 제외한 핵 DNA 비교는 다음과 같은 결론을 내렸다.
- 동질모르파는 진후생동물과 가장 밀접하게 관련되어 있다.
- 석회해면류가 그 다음으로 가깝다.
- 다른 해면류는 이러한 그룹의 진화적 "숙모"이다.
- 캄브리아기 암석에서 화석이 발견되는 주머니 모양의 동물인 샹셀로리드는 해면일 수 있다.[109]
동질모르파의 정자는 다른 해면에는 없는 진후생동물의 정자와 특징을 공유한다. 동질모르파와 진후생동물 모두에서 세포층은 다른 해면에서는 발견되지 않는 "IV형" 콜라겐으로 구성된 카펫과 같은 기저막에 부착되어 결합되어 있다. 하지만 모든 해면류의 중교질을 강화하는 스폰진 섬유는 "IV형" 콜라겐과 유사하다.[110]
위에서 설명한 분석은 해면동물이 모든 후생동물, 즉 해면동물과 더 복잡한 그룹을 포함한 모든 다세포 동물의 조상과 가장 가깝다는 결론을 내렸다. 그러나 2008년 균류부터 인간까지 21개 속의 각각 150개 유전자를 비교한 또 다른 분석(해면동물은 두 종만 포함)에서는 빗해파리류(빗해파리)가 표본에 포함된 후생동물 중 가장 기본적인 계통이라고 제안했다.[111][112][113][114] 이것이 사실이라면, 현대 빗해파리는 다른 후생동물과 독립적으로 복잡한 구조를 발달시켰거나, 해면동물의 조상은 더 복잡했고 알려진 모든 해면은 극적으로 단순화된 형태이다. 이 연구는 판형동물과 같은 더 넓은 범위의 해면과 다른 단순한 후생동물을 사용하여 추가 분석을 권장했다.[111]
하지만 데이터를 재분석한 결과, 분석에 사용된 컴퓨터 알고리즘이 다른 종의 유전자와 현저하게 다른 특정 빗해파리 유전자의 존재로 인해 오도되었다는 것을 보여주었다. 해면동물은 다른 모든 동물의 자매군이거나 조상인 준단계통군(paraphyletic grade)이다.[115][116] 사용 가능한 모든 데이터(형태학적, 발생학적 및 분자적)를 결합하여 구성된 '계통수'는 해면동물이 실제로 단계통군이며, 자포동물과 함께 후구동물의 자매군을 형성한다는 결론을 내렸다.[117][118]
2017년에 발표된 후생동물 규모에서 1,719개 단백질의 매우 크고 내부적으로 일관된 정렬은 (i) 동질모르파, 석회해면류, 육방해면류 및 해면류를 대표하는 해면동물이 단계통군이며, (ii) 해면동물이 다른 모든 다세포 동물의 자매군이며, (iii) 빗해파리류가 두 번째로 초기 분기 동물 계통으로 나타나며, (iv) 판형동물이 세 번째 동물 계통으로 나타나고, 그 뒤를 이어 플라눌라류가 후구동물의 자매군인 자포동물이 나타난다는 것을 보여주었다.[119]
2021년 3월 더블린의 과학자들은 해면동물이 다른 모든 동물의 자매군이라는 추가적인 증거를 발견했고,[120] 2023년 5월 Schultz 등은 게놈 동형성(synteny)에서 비가역적 변화 패턴을 발견하여 빗해파리류가 다른 모든 동물의 자매군이라는 강력한 증거를 제시했다.[121]
5. 분류
칼 폰 린네는 대부분의 부착성 동물을 동물류 강의 편형동물목에 속하는 것으로 분류했지만, ''해면'' 속을 해조류목의 식물로 잘못 분류했다.[77] 이후 오랫동안 해면은 나머지 동물계를 구성하는 후생동물과 분리된 아계 측생동물("동물 옆에")에 속하는 것으로 여겨졌다.[103]
해면동물문은 주로 골격의 구성에 따라 여러 강으로 나뉜다.[16][27]
- 육방해면강(유리해면)은 규산질 침골을 가지는데, 가장 큰 것은 여섯 개의 광선을 가지며 개별적으로 존재하거나 융합될 수 있다.[16] 그들의 몸의 주요 구성 요소는 많은 수의 세포가 단일 외피 세포막을 공유하는 합포체이다.[27]
- 석회해면강은 방해석(탄산칼슘의 한 형태)으로 만들어진 골격을 가지는데, 이는 별개의 침골이나 큰 덩어리를 형성할 수 있다. 모든 세포는 하나의 핵과 막을 가지고 있다.[27]
- 대부분의 해면류는 연조직 내에 규산질 침골이나 해면질 섬유, 또는 둘 다를 가지고 있다. 그러나 몇몇은 탄산칼슘의 또 다른 형태인 아라고나이트로 만들어진 거대한 외부 골격을 가지고 있다.[16][27] 모든 세포는 하나의 핵과 막을 가지고 있다.[27]
- 화석해면류는 캄브리아기의 화석으로만 알려져 있다.[103]
1970년대에는 거대한 탄산칼슘 골격을 가진 해면이 별도의 강인 각질해면류(혹은 "산호 해면")에 속하는 것으로 분류되었다.[78] 그러나 1980년대에 이것들이 모두 석회해면류 또는 해면류의 구성원이라는 것이 밝혀졌다.[79]
지금까지 과학 논문에서는 약 9,000종의 해면동물이 확인되었는데,[27] 그중 유리해면이 약 400종, 석회해면이 약 500종, 나머지는 해면류이다.[17] 그러나 얕은 바다에서조차 수직 암벽과 동굴 벽, 암석과 산호 바위의 갤러리 등 일부 서식지는 거의 조사되지 않았다.[27] 해면동물은 전통적으로 석회해면강(Calcarea), 육방해면강(Hexactinellida), 그리고 데모스폰지아강(Demospongiae)의 세 가지 강으로 분류되었다. 그러나 최근 연구에 따르면 데모스폰지아강에 속하는 것으로 여겨졌던 동질골격해면강(Homoscleromorpha)은 다른 해면동물 강과는 유전적으로 매우 다른 계통 관계를 가지고 있다는 사실이 밝혀졌다.[14] 따라서 동질골격해면강은 최근 네 번째 해면동물 강으로 인정받게 되었다.[80][81]
해면동물은 주로 골격의 구성 성분에 따라 다음과 같은 강으로 분류된다.[82] 아래 표는 진화 순서대로 위에서 아래로 나열되어 있다.
; 석회해면강
: 골격의 주성분은 탄산칼슘이며, 모두 해산인 해면동물이다.
:* 칼키네아아강 Calcinea
:** 로이케티다목 Leucettida
:** 클라토리니다목 Clathorinida - 클라토리나해면
:* 칼카로네아아강 Calcaronea
:** 아미해면목 Leucosolenida - 바구니해면
:** 츠보해면목 Syncettida - 컵해면
:* 파레트로니다아강 Pharetronida
:** 페트로비오나목 Inozoa
:** 소라시아목 Sophinctizoa
; 보통해면강
: 현생하는 해면의 95%가 이 강에 속한다. 골격은 상당히 유연성 있는 해면질 섬유인 스폰진으로 구성되어 있다. 스폰진의 주성분은 다른 모든 동물이 가지는 세포외기질인 콜라겐의 조상 물질이다.
:* 동골해면아강 Homosclromorpha
:** 동골해면목 Homoscleromorhida - 김해면
:* 사방해면아강 Tetrectinomorpha
:** 유성해면목 Choristida - 두꺼운피부해면
:** 나선해면목 Spirophorida - 젤리해면, 토우나스해면
:** 이시해면목 Lithistida - ''Theonella cylindrica''
:** 경해면목 Hadromerida - 빵해면·유자해면
:** 중축해면목 Axinelida - 뿔가지해면
:* 각질해면아강 Ceractinomorpha
:** 그물각질해면목 Dictyoceratida - 목욕해면
:** 수상각질해면목 Dendroceratida
:** 단골해면목 Haplosclerida - 보라해면、거친해면、솜털해면
:** 이와해면목 Petrosida - ''Petrosia ficiformis''
:** 다골해면목 Poecilosclerida
:** 갯벌해면목 Halichondrida - 주황갯벌해면、검정갯벌해면
; 육방해면강
: 유리해면이라고도 불리며, 육방성상의 규산질의 골편을 주로 하는 골격을 가진다. 심해의 모래밭 등에 서식하고 있다.
:* 양반아강 Amphidiscophora
:** 양반목 Amphidiscosa
:* 육방성아강 Hexasteriphora
:** 육방목 Dictynia
:** 해면동굴목 Lyssacina - 해면동굴
:** 아우록시스티스목 Lycniscosa
; 경골해면강
: 탄산칼슘 골격의 주위를 규산질 골편과 해면 조직이 둘러싸고 있다. 대부분이 화석 종이다.
:* 아스트로스크레라목 Astrosclerida
:* 케라토포렐라목 Ceratoporellida
:* 층판해면목 Tabulospongida
6. 인간과의 관계
해면은 다양한 방식으로 인간과 관계를 맺고 있다.
보통해면강에 속하는 6종의 해면은 해면질 섬유로만 이루어져 있어 화장용이나 목욕용 스펀지로 사용된다. 지중해산, 홍해산 해면이 부드럽고 품질이 좋다고 알려져 있다. 해저에서 채취한 해면의 조직을 부패시켜 씻어내고 남은 골격이 상점에서 볼 수 있는 스펀지이다. 일본에서는 유리해면의 일종인 카이로우도우케츠가 독특한 모양으로 장식용으로 사용되었다.
해면은 수중 미생물과 유기물을 제거하여 수질 정화에 기여한다. 또한, 해면에서 발견되는 복잡한 구조의 유기 화합물은 의약품 개발에 활용될 가능성이 있다. 항 HIV 약으로 사용되는 지도부딘은 해면 유래 천연물과 유사한 구조를 가지고 있으며,[135] 할리콘드린 B[136]나 환상 구아니딘류[137] 등은 항암제, 항 HIV 약, 항말라리아약으로 작용하는 것으로 확인되었다.
조선 시대부터 해면은 삽입형 생리대(탐폰)로 사용되기도 했다. 건조시킨 해면을 질에 삽입하여 경혈을 흡수시키는 방식으로, 씻어서 재사용할 수 있다.
6. 1. 이용



1997년 서호주 샤크 만의 큰돌고래들이 해면을 도구로 사용한다는 보고가 있었다. 돌고래는 해양 해면을 주둥이에 부착하는데, 이는 모래 바닥에서 먹이를 찾을 때 보호하기 위해 사용되는 것으로 추정된다.[122] '스폰징(sponging)'으로 알려진 이 행동은 이 만에서만 관찰되었으며, 거의 전적으로 암컷에게서 나타난다. 2005년 연구에 따르면 어미가 딸에게 이 행동을 가르치고 모든 해면 사용자가 밀접하게 관련되어 있어, 이것이 상당히 최근에 나타난 혁신임을 시사한다.[123]
대부분의 해면동물 속(genus)의 탄산칼슘 또는 실리카 해면침(Sponge spicule)은 대부분의 용도에 너무 거칠지만, ''히포스폰기아(Hippospongia)''와 ''스폰기아(Spongia)'' 두 속은 부드럽고 전적으로 섬유질 골격을 가지고 있다.[14] 초기 유럽인들은 투구 완충재, 휴대용 식수 용기, 도시 상수 필터 등 여러 용도로 부드러운 해면을 사용했다. 합성 스펀지가 발명될 때까지 세척 도구, 페인트와 세라믹 유약 도포, 은밀한 피임 도구로 사용되었다. 그러나 20세기 중반까지 과도한 어획으로 인해 해면동물과 관련 산업 모두 멸종 위기에 놓였다.[124]
현재 스펀지와 같은 질감을 가진 많은 물체는 해면동물에서 유래하지 않은 물질로 만들어진다. 합성 스펀지는 개인 및 가정용 세척 도구, 유방 임플란트,[125] 피임 스펀지[126] 등에 사용된다. 일반적으로 사용되는 재료는 셀룰로오스 폼, 폴리우레탄 폼, 그리고 덜 자주 사용되는 실리콘 폼이다.
주방이나 샤워실에서 사용하기 위해 일반적으로 판매되는 수세미 "스펀지"는 동물이 아닌 박과(Cucurbitaceae) 식물인 수세미외(Luffa aegyptiaca)의 섬유질 "골격"에서 유래한다.[127]
보통해면강에 속하는 6종의 해면은 해면질 섬유로만 이루어져 단단한 골편이 없기 때문에 화장용이나 목욕용 스펀지로 사용된다. 지중해산, 홍해산 해면이 부드럽고 품질이 높다고 여겨진다. 해저에서 채취한 해면의 조직을 부패시켜 씻어내고 남은 골격이 상점에서 볼 수 있는 스펀지가 된다. 일본에서는 유리해면의 일종인 카이로우도우케츠가 모양이 독특하여 장식 등으로 사용되었다.
해면은 수중에 떠다니는 먹이를 여과 섭식하기 때문에 수질 오염의 원인이 되는 수중 미생물과 유기물을 제거하는 역할을 한다.
해면에서는 복잡한 구조를 가진 유기 화합물이 다수 발견되었으며, 의약품 후보로 기대되고 있다. 항 HIV 약으로 사용되는 지도부딘은 해면 유래 천연물과 유사한 구조를 가지고 있으며,[135] 할리콘드린 B[136]나 환상 구아니딘류[137] 등은 항암제, 항 HIV 약, 항말라리아약으로 작용하는 것이 확인되었다.
삽입형 생리대(탐폰)로 사용되는 경우가 있다. 건조시킨 해면을 질에 삽입하여 경혈을 흡수시킨다. 꺼내서 씻고 말리면 재사용할 수 있으며, 조선 시대부터 사용되었던 것으로 알려져 있다.
6. 2. 환경 정화
광합성을 하는 내생 공생체를 가진 해면동물은 소비하는 것보다 최대 세 배나 많은 산소를 생산하며, 소비하는 것보다 많은 유기물도 생산한다. 이러한 서식지 자원에 대한 기여는 오스트레일리아의 그레이트 배리어 리프에서는 상당하지만 카리브해에서는 비교적 미미하다.[27]대부분의 해면동물은 퇴적물식동물로서 해수에서 유기 잔해 입자와 미생물을 걸러 먹는다. 특히 해면동물은 산호초 먹이 그물에서 퇴적물을 더 높은 영양 단계로 재활용하는 중요한 역할을 한다.[72]
해면은 수중에 떠다니는 먹이를 여과 섭식하기 때문에 수질 오염의 원인이 되는 수중 미생물과 유기물을 제거하는 역할을 한다.
6. 3. 의약품 개발
해면동물은 복잡한 면역계는 없지만, 다른 종의 이식은 거부하고 같은 종의 이식은 받아들인다. 일부 해양 종에서 회색 세포는 이물질 거부 반응에서 주된 역할을 하며, 침입 시 화학 물질을 생성하여 침입자가 해면 내부 수송 시스템을 이용하지 못하게 한다. 침입이 지속되면 독소를 방출하여 해당 부위 세포를 죽인다. 이 "면역" 체계는 최대 3주 동안 활성화될 수 있다.[82]해면동물은 바이러스, 세균, 종양, 곰팡이를 제어하는 화학 물질을 가지고 있어 의학적으로 잠재력이 있다.[128][129] 이러한 화학 물질은 해면동물 자체 또는 해면동물의 미생물 공생체에서 발견된다.

보호 수단이 없는 해면동물은 다양한 화합물을 합성하는데, 그중 하나는 옥실리핀이라는 산화된 지방산 유도체이다. 이들은 항암, 항균, 항진균 특성을 가진다. 오키나와산 ''Plakortis'' 해면동물에서 분리된 플라코리딘 A는 마우스 림프종 세포에 대한 세포독성 물질로서 잠재력을 보였다.[130][131]
해면에서 발견된 복잡한 구조의 유기 화합물은 의약품 후보로 기대된다. 항 HIV 약 지도부딘은 해면 유래 천연물과 유사하며,[135] 할리콘드린 B,[136] 환상 구아니딘류[137] 등은 항암제, 항 HIV 약, 항말라리아약으로 작용한다.
6. 4. 기타
보통해면강에 속하는 6종의 해면은 해면질 섬유로만 이루어져 단단한 골편이 없기 때문에 화장용이나 목욕용 스펀지로 사용된다. 지중해산, 홍해산 해면이 부드럽고 품질이 높다고 여겨진다. 해저에서 채취한 해면의 조직을 부패시켜 씻어내고 남은 골격이 상점에서 볼 수 있는 스펀지가 된다. 일본에서는 유리해면의 일종인 카이로우도우케츠가 모양이 독특하여 장식 등으로 사용되었다.해면은 수중에 떠다니는 먹이를 여과 섭식하기 때문에 수질 오염의 원인이 되는 수중 미생물과 유기물을 제거하는 역할을 한다.
해면에서는 복잡한 구조를 가진 유기 화합물이 다수 발견되었으며, 의약품 후보로 기대되고 있다. 항 HIV 약으로 사용되는 지도부딘은 해면 유래 천연물과 유사한 구조를 가지고 있으며[135], 그 외에도 할리콘드린 B[136]나 환상 구아니딘류[137] 등이 항암제나 항 HIV 약, 항말라리아약으로 작용하는 것이 확인되었다.
삽입형 생리대(탐폰)로 사용되는 경우가 있다. 건조시킨 해면을 질에 삽입하여 경혈을 흡수시킨다. 꺼내서 씻고 말리면 재사용할 수 있다. 조선 시대부터 사용되었던 것으로 알려져 있다.
참조
[1]
논문
Ediacarian sponge spicule clusters from southwestern Mongolia and the origins of the Cambrian fauna
https://www.research[...]
1997-04
[2]
논문
Sterol and genomic analyses validate the sponge biomarker hypothesis
https://www.pnas.org[...]
2016-02-22
[3]
논문
Animals die more shallowly: they aren't deceased, they're dead. Animals in the polish linguistic worldview and in contemporary life sciences
2018
[4]
웹사이트
Porifera
https://www.marinesp[...]
Flanders Marine Institute
2024-05-08
[5]
사전
porifera
Merriam-Webster
2024-05-12
[6]
논문
Improved Modeling of Compositional Heterogeneity Supports Sponges as Sister to All Other Animals
2017-12
[7]
논문
Alternative neural systems: What is a neuron? (Ctenophores, sponges and placozoans)
2022-12-23
[8]
사전
Spongiology
Merriam-Webster
2017-12-27
[9]
사전
σπόγγος
https://www.perseus.[...]
2021-09-05
[10]
웹사이트
Structure of Sponges
https://www.qm.qld.g[...]
2019-09-27
[11]
논문
The Porifera Ontology (PORO): enhancing sponge systematics with an anatomy ontology
2014-09-08
[12]
논문
Evaluating multiple alternative hypotheses for the origin of Bilateria: an analysis of 18S rRNA molecular evidence
1998-12
[13]
논문
Prey capture and digestion in the carnivorous sponge ''Asbestopluma hypogea'' (Porifera: Demospongiae)
[14]
서적
Sponges
Hutchinson
[15]
서적
Biology 2e
https://d3bxy9euw4e1[...]
OpenStax
2018
[16]
서적
Systema Porifera: A Guide to the Classification of Sponges
https://books.google[...]
Kluwer Academic/Plenum
[17]
서적
Invertebrate Zoology
https://archive.org/[...]
Brooks / COLE Publishing
[18]
논문
Evolutionary conservation of the antimicrobial function of mucus: a first defence against infection
2018-07-04
[19]
서적
Invertebrate Zoology
Oxford University Press
[20]
웹사이트
Studies in Life Sciences, University of Alberta: Glass Sponge Ecology
http://www.biology.u[...]
2008-03-16
[21]
서적
Invertebrate Zoology
Brooks / Cole
2004
[22]
논문
Profiling cellular diversity in sponges informs animal cell type and nervous system evolution
2021-11
[23]
논문
Blue-light-receptive cryptochrome is expressed in a sponge eye lacking neurons and opsin
2012-04
[24]
서적
Invertebrate Zoology
Brooks / Cole
2004
[25]
서적
Integrated Principles of Zoology
McGraw-Hill
[26]
웹사이트
Marine Species Identification Portal: Halisarca dujardini
http://species-ident[...]
2019-08-02
[27]
서적
Encyclopedia of Life Sciences
John Wiley & Sons
[28]
논문
Ecology of siliceous sponges: Application to the environmental interpretation of the Upper Jurassic sponge facies (Oxfordian) from Spain
http://www.ucm.es/BU[...]
[29]
논문
In situ feeding and metabolism of glass sponges (Hexactinellida, Porifera) studied in a deep temperate fjord with a remotely operated submersible
[30]
논문
Global diversity of sponges (Porifera)
[31]
뉴스
4 new species of 'killer' sponges discovered off Pacific coast
http://www.cbc.ca/ne[...]
2014-04-19
[32]
논문
Predation on copepods by an Alaskan cladorhizid sponge
[33]
논문
Carnivorous sponges
[34]
논문
A new genus of carnivorous sponges (Porifera: Poecilosclerida, Cladorhizidae) from the deep N-E Pacific, and remarks on the genus ''Neocladia''
http://www.mapress.c[...]
[35]
논문
New species from the deep Pacific suggest that carnivorous sponges date back to the Early Jurassic
[36]
논문
Light inside sponges
[36]
뉴스
Nature's 'fibre optics' experts
http://news.bbc.co.u[...]
2008-11-11
[37]
서적
Pennak's Freshwater Invertebrates of the United States: Porifera to Crustacea
https://books.google[...]
John Wiley and Sons
[38]
논문
Global patterns in symbiont selection and transmission strategies in sponges
2022-10-26
[39]
논문
Symbiont transmission in marine sponges: reproduction, development, and metamorphosis
2022-05-00
[40]
논문
Some Like It Fat: Comparative Ultrastructure of the Embryo in Two Demosponges of the Genus Mycale (Order Poecilosclerida) from Antarctica and the Caribbean
2015-03-18
[41]
논문
Embryogenesis in the glass sponge Oopsacas minuta: Formation of syncytia by fusion of blastomeres
2006-02-16
[42]
논문
The Molecular Machinery of Gametogenesis in Geodia Demosponges (Porifera): Evolutionary Origins of a Conserved Toolkit across Animals
2020-12-16
[43]
논문
Kinetics and rhythm of body contractions in the sponge Tethya wilhelma (Porifera: Demospongiae)
2004-12-00
[44]
논문
A Post-Synaptic Scaffold at the Origin of the Animal Kingdom
2007-06-06
[45]
논문
Hierarchical assembly of the siliceous skeletal lattice of the hexactinellid sponge Euplectella aspergillum
2007-00-00
[46]
논문
Latitudinal variation in spongivorous fishes and the effectiveness of sponge chemical defenses
https://web.archive.[...]
2008-01-00
[47]
서적
Deep-sea Biology: A Natural History of Organisms at the Deep-Sea Floor
https://books.google[...]
Cambridge University Press
[48]
논문
Video-monitored predation by Caribbean reef fishes on an array of mangrove and reef sponges
1996-00-00
[49]
논문
Chemical defenses and resource trade-offs structure sponge communities on Caribbean coral reefs
2014-03-00
[50]
서적
Extraordinary Animals: An Encyclopedia of Curious and Unusual Animals
https://archive.org/[...]
Greenwood Publishing Group
[51]
논문
Population Dynamics of a Sponge Disease on Caribbean Reefs
http://archive.rubic[...]
Proceedings of the American Academy of Underwater Sciences 31st Symposium
[52]
논문
Aplysina red band syndrome: a new threat to Caribbean sponges
2006-07-00
[52]
웹사이트
New disease threatens sponges
http://www.practical[...]
2006-10-17
[53]
논문
Collaboration among sponge species increases sponge diversity and abundance in a seagrass meadow
2008-06-00
[54]
논문
Species boundaries, specialization, and the radiation of sponge-dwelling alpheid shrimp
[55]
서적
Coral Reefs: Cities Under The Seas
The Darwin Press, Inc.
[56]
논문
Population dynamics and epibiont associations of hermit crabs (Crustacea: Decapoda: Paguroidea) on Dog Island, Florida
https://museumsvicto[...]
2022-01-24
[57]
논문
Utilizing sponge spicules in taxonomic, ecological and environmental reconstructions: a review
2020-12-18
[58]
논문
Foundation Species Abundance Influences Food Web Topology on Glass Sponge Reefs
Frontiers Media SA
2020-09-24
[59]
논문
Coral mucus fuels the sponge loop in warm- and coldwater coral reef ecosystems
[60]
논문
Differential recycling of coral and algal dissolved organic matter via the sponge loop
2017-03-00
[61]
논문
In situ release of mucus and DOC-lipid from the corals Acropora variabilis and Stylophora pistillata in different light regimes.
1987-07-00
[62]
논문
Coral mucus functions as an energy carrier and particle trap in the reef ecosystem
2004-00-00
[63]
논문
Net release of dissolved organic matter by the scleractinian coral Acropora pulchra
2009-00-00
[64]
논문
Organic matter release by dominant hermatypic corals of the Northern Red Sea.
2010-09-00
[65]
논문
Expulsion of zooxanthellae by symbiotic cnidarians from the Red Sea.
1987-04-00
[66]
논문
Preferential expulsion of dividing algal cells as a mechanism for regulating algal-cnidarian symbiosis
https://www.biodiver[...]
2000-00-00
[67]
논문
Diurnal lipid and mucus production in the staghorn coral Acropora acuminata
1980-00-00
[68]
논문
Autotrophic carbon budget in coral tissue: a new 13C-based model of photosynthate translocation
2012-04-15
[69]
논문
Enhancement of pico- and nanoplankton growth by coral exudates
2000-00-00
[70]
논문
Organic matter release by Red Sea coral reef organisms—potential effects on microbial activity and in situ O2 availability.
2010-07-00
[71]
논문
Production and bacterial decomposition of dissolved organic matter in a fringing coral reef
2011
[72]
논문
Reef sponges facilitate the transfer of coral-derived organic matter to their associated fauna via the sponge loop
2018-02-23
[73]
논문
Surviving in a Marine Desert: The Sponge Loop Retains Resources Within Coral Reefs
2013-10-04
[74]
논문
The sponge holobiont in a changing ocean: from microbes to ecosystems
2018
[75]
논문
Editorial for: Microbial symbiosis of marine sessile hosts- diversity and function
2015-06-16
[76]
논문
The Sponge Hologenome
2016-04
[77]
웹사이트
Spongia Linnaeus, 1759
http://www.marinespe[...]
World Register of Marine Species
2012-07-18
[78]
논문
Jamaican coralline sponges: Their morphology, ecology and fossil relatives
http://mgg.rsmas.mia[...]
[79]
서적
The Origins and Relationships of Lower Invertebrates
Oxford University Press
[80]
논문
Molecular phylogeny restores the supra-generic subdivision of homoscleromorph sponges (Porifera, Homoscleromorpha)
2010-12
[81]
논문
No longer Demospongiae: Homoscleromorpha formal nomination as a fourth class of Porifera
https://hal.archives[...]
2012-05
[82]
서적
Invertebrate Zoology
Oxford University Press
[83]
논문
Solenoid: a new aquiferous system to Porifera
[84]
서적
The Invertebrate Tree of Life
Princeton University Press
2020-03-03
[85]
논문
Ancient gene linkages support ctenophores as sister to other animals
https://www.nature.c[...]
2023-05-17
[86]
논문
Phylogenetic-Signal Dissection of Nuclear Housekeeping Genes Supports the Paraphyly of Sponges and the Monophyly of Eumetazoa
https://academic.oup[...]
2009-10
[87]
논문
Sponge paraphyly and the origin of Metazoa
https://academic.oup[...]
2001-01-01
[88]
논문
Where's the glass? Biomarkers, molecular clocks, and microRNAs suggest a 200 Myr missing Precambrian fossil record of siliceous sponge spicules
2010-01
[89]
서적
Systema Porifera: A Guide to the Classification of Sponges
http://webdoc.sub.gw[...]
Kluwer Academic Plenum
[90]
논문
Precambrian sponges with cellular structures
1998-02
[91]
논문
The unique skeleton of siliceous sponges (Porifera; Hexactinellida and Demospongiae) that evolved first from the Urmetazoa during the Proterozoic: a review
[92]
논문
Early Cambrian sponge spicules from the Cerro Clemente and Cerro Rajón, Sonora, México
[93]
논문
Precambrian sponges with cellular structures
1998-02
[94]
논문
Possible poriferan body fossils in early Neoproterozoic microbial reefs
2021-08
[95]
웹사이트
Demospongia
http://www.ucmp.berk[...]
U.C. Berkeley
[96]
논문
Earliest known rugosan-stromatoporoid symbiosis from the Llandovery of Estonia (Baltica)
https://www.research[...]
[97]
논문
Sterols in a unicellular relative of the metazoans
2008-07
[98]
논문
Sponges: New views of old animals
2005-04
[99]
논문
Fossil steroids record the appearance of Demospongiae during the Cryogenian period
https://authors.libr[...]
2009-02
[100]
논문
Questioning the evidence of organic compounds called sponge biomarkers
[101]
논문
The slow rise of complex life as revealed through biomarker genetics
2018-09
[102]
논문
Sterol and genomic analyses validate the sponge biomarker hypothesis
2016-03
[103]
논문
Archaeocyatha: A history of phylogenetic interpretation
[104]
논문
Burgess Shale-type preservation of both non-mineralizing and "shelly" Cambrian organisms from the Mackenzie Mountains, northwestern Canada
[105]
논문
New well-preserved scleritomes of Chancelloridae from the early Cambrian Yuanshan Formation (Chengjiang, China) and the middle Cambrian Wheeler Shale (Utah, USA) and paleobiological implications
[106]
논문
Skeletal microstructure indicates Chancelloriids and Halkieriids are closely related
[107]
논문
Sponge paraphyly and the origin of Metazoa
2001-01-00
[108]
논문
Evaluating hypotheses of basal animal phylogeny using complete sequences of large and small subunit rRNA
2001-08-00
[109]
논문
Poriferan paraphyly and its implications for Precambrian paleobiology
http://www.dartmouth[...]
2008-11-04
[110]
논문
Evolution of collagens
2002-11-00
[111]
논문
Broad phylogenomic sampling improves resolution of the animal tree of life
2008-04-00
[112]
논문
Assessing the root of bilaterian animals with scalable phylogenomic methods
2009-12-00
[113]
논문
The genome of the ctenophore Mnemiopsis leidyi and its implications for cell type evolution
2013-12-00
[114]
논문
The ctenophore genome and the evolutionary origins of neural systems
2014-00-00
[115]
논문
Genomic data do not support comb jellies as the sister group to all other animals
2015-12-00
[116]
서적
Spineless: the science of jellyfish and the art of growing a backbone
Riverhead Books
2018-10-00
[117]
논문
Concatenated analysis sheds light on early metazoan evolution and fuels a modern "urmetazoon" hypothesis
2009-01-00
[118]
논문
Topology-dependent asymmetry in systematic errors affects phylogenetic placement of Ctenophora and Xenacoelomorpha
2020-12-00
[119]
Submitted manuscript
A Large and Consistent Phylogenomic Dataset Supports Sponges as the Sister Group to All Other Animals
https://hal.archives[...]
2018-11-04
[120]
논문
Evidence for sponges as sister to all other animals from partitioned phylogenomics with mixture models and recoding
2021-03-00
[121]
논문
Ancient gene linkages support ctenophores as sister to other animals
2023-06-00
[122]
논문
Sponge-carrying by Indian Ocean bottlenose dolphins: Possible tool-use by a delphinid
[123]
논문
Cultural transmission of tool use in bottlenose dolphins
2005-06-00
[124]
서적
Oceans Past: Management Insights from the History of Marine Animal Populations
Earthscan
[125]
서적
Cleavage
https://books.google[...]
Rutgers University Press
[126]
웹사이트
Sponges
http://www.cervicalb[...]
2006-09-17
[127]
논문
Loofah — The sponge gourd
[128]
서적
Sponges (Porifera): Porifera
Springer
[129]
논문
Antibiotic and cytotoxic activity of brominated compounds from the marine sponge Verongia aerophoba
1993-00-00
[130]
논문
Plakoridine A, a new tyramine-containing pyrrolidine alkaloid from the ''Okinawan marine'' sponge Plakortis sp
[131]
논문
A synthetic approach to the plakoridines modeled on a biogenetic theory
2005-04-18
[132]
논문
The Inner Lives of Sponges
[133]
논문
Discovery of the Novel Candidate Phylum “Poribacteria” in Marine Sponges
[134]
문서
Vacelet
[135]
뉴스
Liquid Gold Mine
http://pubs.acs.org/[...]
[136]
웹사이트
海洋生物由来の“切れ者分子”の謎
http://www.sci.nagoy[...]
[137]
간행물
海産環状グアニジン系天然物の全合成及び新規グアニジン型不斉有機分子触媒の開発
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com