은편모조류
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
은편모조류는 특징적인 분출소체를 가지는 단세포 진핵생물로, 전 세계 담수 및 해양 환경에 널리 분포하며, 엽록체를 통해 광합성을 한다. 엽록체는 홍조류 유래이며, 클로로필 a와 c, 피코빌리단백질 등을 포함한다. 일부 종은 세포 내 공생체로 존재하며, 와편모조류나 섬모충류의 엽록체로 활용되기도 한다. 은편모조류는 이분열을 통해 증식하며, 과거에는 다양한 분류 체계로 다루어졌으나, 현재는 크립토식물문 또는 크립토조강으로 분류된다. 한국 연안에서도 발견되며, 해양 생태계의 중요한 구성원이다.
더 읽어볼만한 페이지
- 은편모조류 - 스토레아툴라속
스토레아툴라속은 세균에 속하는 속이다. - 은편모조류 - 은편모조강
은편모조강은 분출소체라는 압출체를 가진 조류의 한 종류로, 다양한 색깔의 엽록체와 핵형질체를 가지며, 무성 생식과 유성 생식을 모두 수행하고, 다양한 파장의 빛을 흡수할 수 있어 생태적 틈새에서 중요한 역할을 한다. - 토머스 캐빌리어 스미스가 명명한 분류군 - 코아노조아
코아노조아는 칼라 구조를 특징으로 하는 깃편모충류와 동물을 포함하는 계통 분류군으로, 동물의 기원과 밀접한 관련이 있으며, 동물 진화 연구에 중요한 연결고리를 제공한다. - 토머스 캐빌리어 스미스가 명명한 분류군 - 합티스타
합티스타는 2015년 캐빌리어-스미스에 의해 명명된 생물 분류군으로, 엑스카바타 생물군 중 디스코바에 속하며, 하위 분류로 알베이다아문, 육질태양충아문, 착편모조아문 등이 있다. - 조류 - 스토레아툴라속
스토레아툴라속은 세균에 속하는 속이다. - 조류 - 규조토
규조토는 규조류의 화석 잔해로 구성된 퇴적암으로, 여과 보조제, 연마재, 단열재, 살충제 등 다양한 용도로 사용되며 전 세계적으로 분포한다.

2. 특징
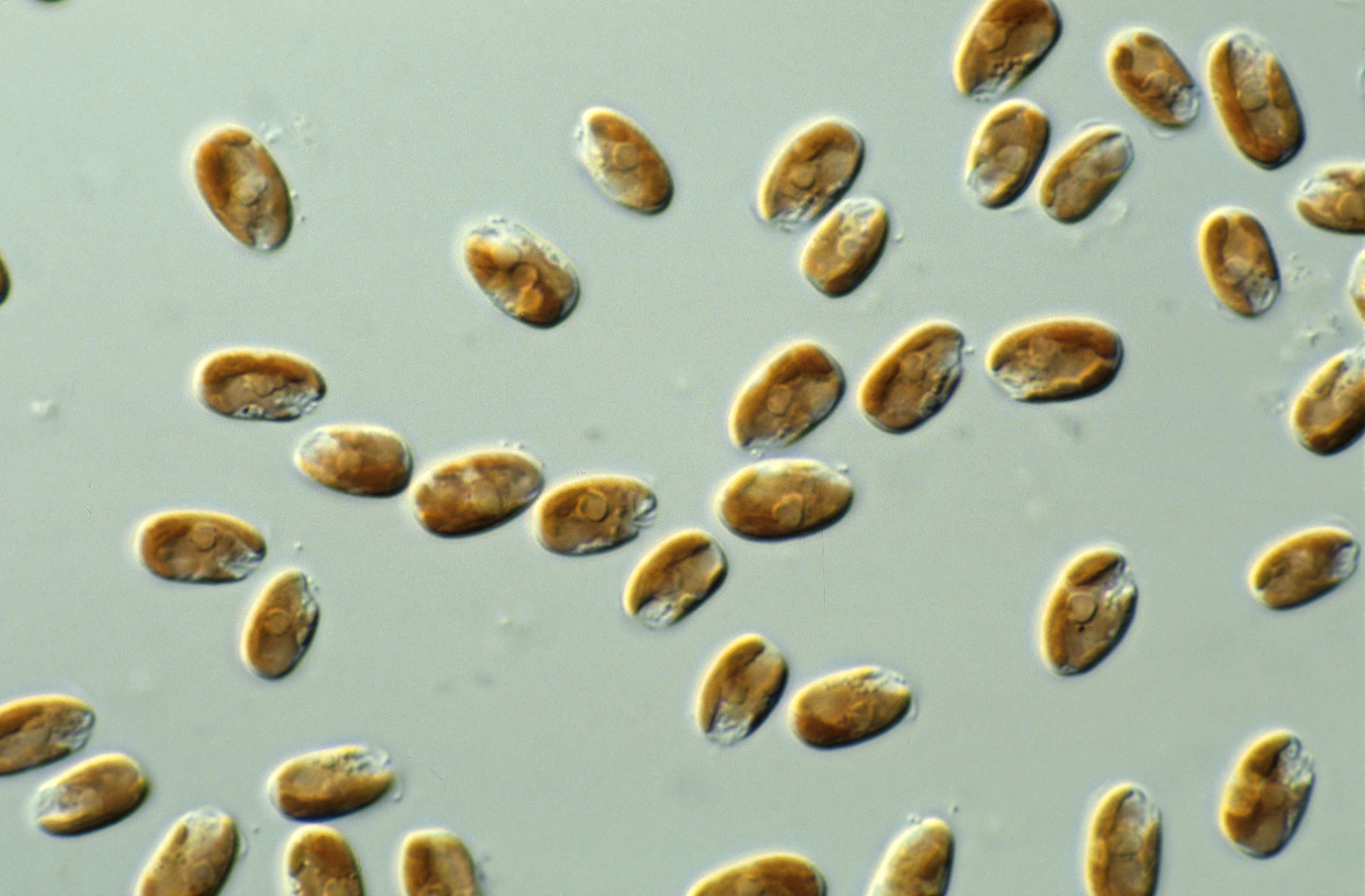
은편모조류의 가장 두드러진 특징은 압출체의 일종인 분출소체를 가진다는 점이다. 이 구조는 외부 자극에 반응하여 내용물을 방출함으로써 세포가 빠르게 이동하도록 돕는다.[9] 큰 분출소체는 세포 내 특정 주머니와 연관되어 있고, 더 작은 것들은 세포 외피인 주피막 아래에 분포한다.[10][11]
또한, 대부분의 은편모조류는 엽록체를 가지고 있는데, 이는 홍조류가 세포 내 공생을 통해 통합된 결과로 여겨진다.[13] 이 엽록체는 4개의 막으로 둘러싸여 있으며, 중간 두 막 사이에는 공생 과정에서 축소된 홍조류의 핵 흔적인 핵상체가 남아있는 독특한 구조를 보인다.
2. 1. 세포 구조
세포는 대략 납작한 쌀알 모양이며, 쌀로 말하면 배아 부위에 2개의 편모를 갖는다. 편모는 서로 평행하게 삽입되며, 소포체 내에서 형성되어 세포 표면으로 운반되는 마스티고네메라고 불리는 이분 구조의 털로 덮여 있다. 작은 비늘도 편모와 세포체에 존재할 수 있다. 편모 기부 근처에는 갈레트(인두부)라고 불리는 함입부가 있으며, 갈레트의 내벽에는 이를 둘러싸듯이 트리코시스트(trichocyst; 모포)라고 불리는 사출 장치가 배치되어 있다. 이 트리코시스트는 마치 화장지가 말려 있는 상태로 수납되어 있으며, 사출 시에는 나선형으로 풀려 펼쳐진다. 갈레트 주변에는 안점이나 반사체(refractile body)와 같은 광학계 기관을 갖는 종도 있다.은편모조류는 특징적인 압출체인 분출소체의 존재로 구별되는데, 이는 긴장 상태에서 유지되는 두 개의 연결된 나선형 리본으로 구성된다.[9] 세포가 기계적, 화학적 또는 광학적 스트레스에 의해 자극을 받으면 방출되어 교란으로부터 지그재그로 세포를 추진한다. 광학 현미경으로 관찰 가능한 큰 분출소체는 주머니와 관련되어 있으며, 더 작은 분출소체는 은편모조류 특유의 세포 외피인 주피막 아래에서 발생한다.[10][11]

전적으로 엽록체를 갖지 않는 Goniomonadea 강[12]과 백색체를 갖는 ''Cryptomonas paramecium'' (이전 명칭: ''Chilomonas paramecium'')을 제외하고, 은편모조류는 하나 또는 두 개의 엽록체를 가지고 있다. 이들은 클로로필 ''a''와 ''c'', 피코빌리단백질 및 기타 색소를 함유하며 색상이 다양하다 (갈색, 적색에서 청록색). 각 엽록체는 4개의 막으로 둘러싸여 있으며, 중간 두 막 사이에는 핵상체라고 불리는 축소된 핵이 있다. 이는 엽록체가 진핵생물 공생체에서 유래되었음을 나타내며, 유전 연구를 통해 홍조류였음이 밝혀졌다.[13] 그러나 엽록체는 홍조류 엽록체와 매우 다르다. 피코빌리단백질은 존재하지만 틸라코이드 루멘에만 존재하며, 피코에리트린 또는 피코시아닌으로만 존재한다. ''Rhodomonas''의 경우 결정 구조가 1.63Å로 결정되었으며,[14] 알파 서브유닛은 다른 알려진 피코빌리단백질과 관련이 없음이 밝혀졌다.
미토콘드리아는 편평한 크리스타를 가지고 있으며, 세포 분열은 열려 있고, 유성 생식도 보고되었다.
''Cryptomonas''와 같은 몇몇 은편모조류는 팜멜로이드 단계를 형성할 수 있지만, 주변의 점액에서 쉽게 탈출하여 다시 자유 생활하는 편모충이 된다. 일부 ''Cryptomonas'' 종은 또한 불리한 조건을 견디기 위해 경직된 세포벽을 가진 휴면 단계인 비운동성 미생물 낭종을 형성할 수 있다.
2. 2. 편모
은편모조류의 편모는 종에 따라 길이가 다른 부등장(不等長) 또는 거의 같은 아등장(亞等長)이다. 편모는 서로 평행하게 삽입되며,[11] 마스티고네메(mastigoneme)라고 불리는 이분 구조의 털로 덮여 있다. 이 털은 소포체 내에서 형성되어 세포 표면으로 운반된다.[11]편모 소모(mastigoneme)는 편모 축을 기준으로 양쪽에 돋아 있는 양우형(兩羽型)과 한쪽에만 돋아 있는 편우형(偏羽型)이 있다. 2개의 편모에서 양우형과 편우형의 조합은 다양하다. 은편모조류의 소모는 축 부분과 선단모(先端毛)로 이루어진 2부분 구성으로, 기부를 포함한 3부분으로 구성된 황색조류의 소모와는 다르다. 이 소모가 황색조류처럼 추진력 역전 효과를 가진다고 알려져 있지만, 편우형 편모의 존재나 조합의 문제 때문에 역학적인 근거는 아직 명확하지 않다.
편모와 세포체에는 작은 인편(鱗片)이 존재할 수도 있다.[11] 인편이 있는 경우, 로제트 모양의 특징적인 형태를 보인다.
2. 3. 엽록체
Goniomonadea 강[12]과 같이 엽록체가 없거나, ''Cryptomonas paramecium''처럼 백색체만 가진 일부 종을 제외하면, 대부분의 은편모조류는 하나 또는 두 개의 엽록체를 가진다. 이 엽록체는 보통 세포핵이나 다른 세포 소기관을 둘러싸듯 세포 표면 가까이에 위치한다.광합성 색소로는 클로로필 ''a''와 ''c'', 카로테노이드류, 그리고 피코빌린 단백질(주로 피코시아닌 또는 피코에리트린)을 가진다.[13] 이 때문에 은편모조류는 갈색, 붉은색, 청록색 등 다양한 색을 띤다. 특히 피코시아닌을 주로 가진 ''Chroomonas'' 속은 남세균과 비슷한 청록색을 나타낸다. 피코빌린 단백질은 홍조류나 남세균처럼 피코빌리솜 구조를 이루지 않고 틸라코이드 내부 공간(루멘)에 채워져 있어, 다른 조류에 비해 틸라코이드가 더 두껍게 보인다.
엽록체는 4개의 막으로 둘러싸여 있으며, 가장 바깥쪽 막은 세포핵의 막과 연결되어 있다. 이는 은편모조류의 엽록체가 홍조류를 세포 내 공생으로 받아들이는 2차 내부 공생 과정에서 유래했음을 시사한다.[13] 이 과정에서 흡수된 홍조류의 세포핵은 크게 축소되어 핵상체(누클레오모프)라는 형태로 엽록체 내에 남아 있다. 핵상체는 엽록체의 4중막 중 바깥쪽 두 막과 안쪽 두 막 사이의 공간(periplastidal compartment)에 존재하며, 종에 따라서는 탄소 고정의 중심인 피레노이드에 파묻혀 있기도 하다. 이 공간에는 핵상체 외에도 진핵생물 유래의 80S rRNA와 광합성 산물인 녹말 입자 등도 발견된다.
은편모조류의 엽록체는 유래가 된 홍조류의 엽록체와 몇 가지 차이점을 보인다. 앞서 언급했듯이 피코빌린 단백질이 틸라코이드 막 표면의 피코빌리솜 구조 대신 틸라코이드 내부 공간에 존재한다. 또한, ''Rhodomonas'' 종에서 연구된 피코빌리단백질의 결정 구조(1.63Å)는 다른 조류의 피코빌리단백질과는 다른 독특한 구조적 특징을 가지는 것으로 밝혀졌다.[14]
2. 4. 세포외피
은편모조류의 세포외피는 페리플라스트(periplast)라고 불린다. 이는 세포막과 이를 뒷받침하는 판상 구조로 이루어져 있다. 판상 구조는 단백질로 구성되어 있으며, 그 규칙적인 패턴은 분류 형질로도 유용하다.3. 세포 내 공생
은편모조류는 세포 내에 누클레오모프를 가지고 있어 세포 내 공생설 연구에서 중요한 대상으로 주목받아 왔다. 더 나아가, 은편모조류 자체가 다른 원생생물의 세포 내로 들어가 공생하는 현상도 관찰된다. 이렇게 은편모조류를 세포 내에 받아들여 엽록체처럼 활용하는 생물로는 와편모조류나 섬모충류와 같은 알베올라타 그룹에 속하는 생물들이 알려져 있다.
3. 1. 와편모조류
와편모조류 중 일부는 은편모조류를 세포 내에 섭취하여 엽록체처럼 활용하는 것으로 알려져 있다. 이는 세포 내 공생설과 관련하여 연구자들의 주목을 받는 현상이다. 은편모조류를 활용하는 대표적인 와편모조류 속은 다음과 같다.- Amphidinium: A. wigrense, A. latum 등의 종이 해당된다. 이들은 여러 종류의 은편모조류를 섭취하며, 때로는 색소 조성이 다른 은편모조류를 세포의 좌우에 각각 유지하는 독특한 사례도 보고되었다.
- Dinophysis
- Gymnodinium: G. acidotum 종이 주로 Chroomonas 속의 은편모조류를 섭취하여 이용한다.
3. 2. 섬모충류
섬모충류의 일종인 ''Mesodinium'' 속, 특히 ''M. rubrum'' 종은 세포 내에 은편모조류를 유지하는 것으로 알려져 있다. 이들이 유지하는 은편모조류는 정확한 속(屬)까지는 밝혀지지 않았지만, 여러 마리의 피코에리스린 계열 붉은색 은편모조류인 것으로 확인되었다.4. 분포
5. 생활사
일반적으로 이분열을 통한 무성 생식으로 증식한다. 드물지만, 동형 배우자에 의한 유성 생식도 알려져 있다.
오래전부터 은편모조류는 생활환 중에 서로 다른 형태의 편모충으로 살아가는 모습이 보고되었다. 예를 들어, ''Proteomonas sulcata''는 복상 세대일 때는 큰 편모충 형태를, 단상 세대일 때는 작은 편모충 형태를 띤다. 각 단계에서는 페리플라스트의 패턴도 다르다(Hill and Wetherbee 1986). 이러한 특징은 과거 은편모조류 중 극히 일부에서만 나타나는 것으로 여겨졌으나, 최근 연구(Hoef-Emden and Melkonian 2003)에 따르면 대부분의 은편모조류에 해당하는 현상일 가능성이 제기되었다. 이로 인해 단상 세대와 복상 세대에 같은 종이 다른 이름으로 불리는 경우가 밝혀졌으며, 분류 체계를 정리해야 할 필요성이 생겼다. 이는 갈조류에서도 비슷한 문제가 나타났는데, 배양 기술과 분자 계통 분석 방법의 발달 덕분에 밝혀진 사실이다.
6. 분류
과거 일부 분류에서는 겉보기 색소가 비슷하다는 이유로 와편모조류와 가까운 관계로 보아 피루스조류로 묶거나, 와편모식물문과 함께 붉은색조식물문(Division Pyrrophyta)으로 분류하기도 했으나, 이러한 분류 체계는 현재 사용되지 않는다.
최근에는 독립된 문(門) 수준인 크립토식물문(Division Cryptophyta, Cavalier-Smith 1986)으로 분류하거나, 강(綱) 수준인 크립토조강(Class Cryptophyceae, Pascher 1913, emend. Schoenichen, 1925)으로 분류하는 것이 일반적이다. 하위 분류 체계는 지속적인 연구를 통해 재편되고 있다.
6. 1. 하위 분류
- 은편모조강 (Cryptophyceae)
- * 은편모충목 (Cryptomonadales)
- ** Baffinellaceae
- ** Butschliellaceae
- ** Cryptochrysidaceae
- ** 크립토모나스과 또는 밤색초리말과[21] (Cryptomonadaceae)
- ** 키아토모나스과 (Cyathomonadaceae)
- ** 헤미셀미스과 (Hemiselmidaceae)
- ** 힐레아과 (Hilleaceae)
- * 피레노모나스목 (Pyrenomonadales)
- ** 크루모나스과 (Chroomonadaceae)
- ** 게미니게라과 (Geminigeraceae)
- ** 피레노모나스과 (Pyrenomonadaceae)
- * 테트라고니디움목 (Tetragonidiales)
- ** 테트라고니디움과 (Tetragonidiaceae)
- 고니오모나스강 (Goniomonadea)
- * 고니오모나스목 (Goniomonadales)
- * 헤미아르마목 (Hemiarmida)

6. 2. 계통 분류
2016년 실라(Silar)의 하크로비아 계통 분류에 따르면, 은편모조류는 넓은 의미의 크립티스타에 속한다.[24][25] 넓은 의미의 크립티스타는 코르비헬리아와 좁은 의미의 크립티스타로 나뉜다. 좁은 의미의 크립티스타는 다시 팔피토아문과 롤로모나스아문으로 구성된다. 롤로모나스아문은 레우크립토스상강과 은편모조류로 나뉘는데, 이때 레우크립토스상강에 속하는 카타블레파리스류는 은편모조류의 자매군으로 여겨진다.[19] 은편모조류는 고니오모나스강과 은편모조강으로 분류된다.은편모조류에 대한 최초의 기록은 1831년 크리스티안 고트프리트 에렌베르크가 섬모충류를 연구하면서 남긴 것이다.[15] 이후 식물학자들은 은편모조류를 별도의 조류 그룹인 크립토조강(Cryptophyceae) 또는 크립토식물문(Cryptophyta)으로 분류했고, 동물학자들은 이를 편모충 원생동물의 한 목인 크립토모나스목(Cryptomonadina)으로 취급했다. 과거 일부 분류에서는 겉보기 색소가 비슷하다는 이유로 와편모조류와 가까운 관계로 보아 피루스조류로 묶기도 했다.
은편모조류의 엽록체는 이형편모조류 및 갈조류의 엽록체와 밀접한 관련이 있는 것으로 밝혀져, 캐빌리어-스미스(Cavalier-Smith)는 이 세 그룹을 색소체생물로 통합하기도 했다. 그러나 세포의 미세 구조 차이 등은 이들이 독립적으로 색소체를 획득했을 가능성을 시사하며, 따라서 색소체생물이라는 분류가 다계통군일 수 있다는 반론도 제기되었다. 분자생물학적 증거들은 은편모조류가 원래 종속영양생물이었으며 이차적으로 엽록체를 획득했다는 가설을 지지한다.[16] 파프리(Parfrey) 연구팀과 부르키(Burki) 연구팀 등은 은편모조류를 녹조류 또는 녹조류와 남조식물의 자매 그룹으로 분류하기도 했다.[17][18]
제안된 분류는 다음과 같다.[20]
- ''Cryptomonas''
- ''Chroomonas'' / ''Komma'' 및 ''Hemiselmis''
- ''Rhodomonas'' / ''Rhinomonas'' / ''Storeatula''
- ''Guillardia'' / ''Hanusia''
- ''Geminigera'' / ''Plagioselmis'' / ''Teleaulax''
- ''Proteomonas sulcata''
- ''Falcomonas daucoides''
현재는 독립된 문(門) 수준인 크립토식물문(Division Cryptophyta, Cavalier-Smith 1986)으로 분류하거나, 강(綱) 수준인 크립토조강(Class Cryptophyceae, Pascher 1913, emend. Schoenichen, 1925)으로 분류하는 것이 일반적이다. 하위 분류는 생활환 연구 등에 따라 계속 재편되고 있으며, 주요 분류군은 다음과 같다.
;크립토조강 (Class Cryptophyceae)
:;크립토모나스과 (Family Cryptomonadaceae)
::* 대표 속인 ''Cryptomonas''를 포함하며, ''Chroomonas'', ''Rhodomonas'', ''Plagioselmis'' 등이 속한다. ''Chilomonas'' 속은 엽록체가 이차적으로 소실된 것으로 알려져 있다.
:;크립토크리시스과 (Family Cryptochrysidaceae)
::* ''Cryptochrysis'' 속을 포함한다.
:;고니오모나스과 (Family Goniomonadaceae)
::* ''Goniomonas'' 속과 ''Cyathomonas'' 속(''Goniomonas'' 속으로 통합됨)을 포함한다. 이들은 색소가 없는 무색 크립토조류로, 엽록체를 획득하기 이전에 분기된 그룹으로 여겨진다.
:;헤미셀미스과 (Family Hemiselmidaceae)
::* ''Hemiselmis'' 속을 포함한다.
:;카타블레파리스과 (Family Katablepharidaceae)
::* ''Katablepharis'' 속과 ''Leucocryptos'' 속을 포함하는 무색 크립토조류이다. 두 개의 편모는 각각 유영 편모와 견인 편모로 구분된다. 이 과는 독립된 문(門)인 카타블레파리스식물문(Division Katablepharidophyta) 또는 카타블레파리스문(Phylum Kathablepharida)으로 분류해야 한다는 의견도 있다.
참조
[1]
논문
Kingdom Chromista and its eight phyla: A new synthesis emphasising periplastid protein targeting, cytoskeletal and periplastid evolution, and ancient divergences
2018
[2]
서적
The Invertebrates: A Synthesis
Wiley-Blackwell
[3]
논문
Lateral transfer of introns in the cryptophyte plastid genome
2008-05
[4]
논문
The Katablepharids are a Distant Sister Group of the Cryptophyta: A Proposal for Katablepharidophyta Divisio Nova/Kathablepharida Phylum Novum Based on SSU rDNA and Beta-Tubulin Phylogeny
2005
[5]
웹사이트
Cryptophyta - the cryptomonads
http://www.life.umd.[...]
2009-06-02
[6]
서적
Organelles, Genomes, and Eukaryote Phylogeny
[7]
논문
Goniomonas: rRNA sequences indicate that this phagotrophic flagellate is a close relative of the host component of cryptomonads
[8]
논문
Nuclear genome sequence of the plastid-lacking cryptomonad Goniomonas avonlea provides insights into the evolution of secondary plastids
[9]
서적
Algae
Benjamin Cummings (Pearson)
[10]
논문
A comparison of the periodic sub-structures of the trichocysts of the Cryptophyceae and Prasinophyceae
[11]
논문
The ejectisomes of the flagellate ''Chilomonas paramecium'' - Visualization by freeze-fracture and isolation techniques
[12]
뉴스
Nuclear genome sequence of the plastid-lacking cryptomonad Goniomonas avonlea provides insights into the evolution of secondary plastids
https://bmcbiol.biom[...]
[13]
논문
The highly reduced genome of an enslaved algal nucleus
[14]
논문
Evolution of a light-harvesting protein by addition of new subunits and rearrangement of conserved elements: Crystal structure of a cryptophyte phycoerythrin at 1.63Å resolution.
[15]
논문
Cryptomonad taxonomy in the 21st century: The first 200 years
https://www.research[...]
2018-10-16
[16]
논문
Nuclear genome sequence of the plastid-lacking cryptomonad Goniomonas avonlea provides insights into the evolution of secondary plastids
2018
[17]
논문
Estimating the timing of early eukaryotic diversification with multigene molecular clocks
2011-08-16
[18]
논문
Untangling the early diversification of eukaryotes: a phylogenomic study of the evolutionary origins of Centrohelida, Haptophyta and Cryptista
2016-01-27
[19]
논문
Mitochondrial Genomes of Hemiarma marina and Leucocryptos marina Revised the Evolution of Cytochrome c Maturation in Cryptista
2020
[20]
웹사이트
Cryptomonads
http://tolweb.org/Cr[...]
2009-06-24
[21]
서적
조선포자식물 조류편3
과학백과사전종합출판사
[22]
저널
Lateral transfer of introns in the cryptophyte plastid genome
http://nar.oxfordjou[...]
[23]
웹인용
Cryptophyta - the cryptomonads
http://www.life.umd.[...]
2009-06-02
[24]
인용
Protistes Eucaryotes: Origine, Evolution et Biologie des Microbes Eucaryotes
https://hal.archives[...]
2016
[25]
인용
Higher Level Classification of All Living Organisms
http://journals.plos[...]
2015
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com