헤르페스바이러스과
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
헤르페스바이러스과는 DNA를 게놈으로 가지는 바이러스들의 과로, 알파, 베타, 감마헤르페스바이러스아과로 분류된다. 이들은 숙주 세포의 핵 내에서 복제되며, 감염된 세포는 용해성 바이러스 유전자를 전사하거나 잠복 상태를 유지할 수 있다. 헤르페스바이러스는 면역 회피 기전을 통해 평생 감염을 유지하며, 사람에게 질병을 일으키는 9가지 종류의 사람 헤르페스바이러스가 존재한다. 헤르페스바이러스 감염에 대한 치료제로는 아시클로비르 등이 있으며, 알츠하이머병 등 다양한 질병과의 연관성에 대한 연구가 진행 중이다.
더 읽어볼만한 페이지
- 헤르페스바이러스과 - 오제스키병
오제스키병은 아러더 오제스키가 발견한 돼지에서 주로 발생하는 바이러스성 질병으로, 다른 포유류에도 감염될 수 있으며 돼지에서는 호흡기 감염, 다른 동물에서는 가려움증과 신경계 증상을 유발하고, 국내 발생 사례는 없으나 백신 접종으로 통제 가능하며 바이러스 신경 추적자로도 활용된다. - 헤르페스바이러스과 - 고양이 바이러스성 비기관지염
고양이 바이러스성 비기관지염은 고양이 허피스 바이러스 타입 1에 의해 발생하는 상부 호흡기 질환으로, 감염된 고양이와의 접촉이나 오염된 물건을 통해 전파되며 기침, 재채기, 콧물, 결막염 등의 증상을 보이고, 백신이 존재하나 감염을 완전히 예방하지는 못하며, 항생제, 항바이러스제, 지지요법 등으로 치료하고 위생 관리가 중요하다. - 바이러스 과 - 코로나바이러스과
코로나바이러스과는 표면의 스파이크 단백질 모양이 코로나와 유사한 RNA 바이러스로, 숙주 세포 침투 및 복제 과정을 거쳐 유전적 재조합을 통해 진화하며, 레토바이러스아과와 코로나바이러스아과로 나뉘고 다양한 속과 종으로 분류되며, 사람에게 감기부터 SARS, MERS, 코로나19와 같은 심각한 호흡기 질환을 일으킨다. - 바이러스 과 - 파르보바이러스
파르보바이러스는 선형 단일 가닥 DNA 게놈을 가진 바이러스로, NS 유전자와 VP 유전자를 포함하며, 롤링 헤어핀 복제 방식으로 게놈을 복제하고, 질병을 유발하거나 유전자 치료에 활용된다. - 감염성 암원 - 결핵
결핵은 결핵균에 의해 발생하는 감염성 질환으로, 주로 폐에 영향을 미치며 기침, 객혈 등의 증상을 보이지만 무증상일 수도 있고, 호흡기 비말 전파를 통해 감염되며, 항생제 치료를 하지만 약물 내성 문제로 인해 공중 보건 문제로 남아있다. - 감염성 암원 - 자궁경부암
자궁경부암은 인유두종바이러스 감염이 주원인인 자궁 경부의 악성 종양으로, 초기에는 증상이 미미하지만 진행 시 질 분비물 증가, 성교통, 골반통이 나타날 수 있으며, 자궁경부 세포진 검사, 조직 생검 등으로 진단하고 수술, 방사선 치료, 항암 치료 등으로 치료하며, HPV 백신 접종과 정기 검진으로 예방이 가능하다.

2. 분류
헤르페스바이러스과는 헤르페스바이러스목에 속하는 바이러스의 과이다. 바이러스학적으로는 크게 세 가지 아과로 분류된다.
2. 1. 알파헤르페스바이러스아과 (Alphaherpesvirinae)
알파헤르페스바이러스아과는 주로 점막 상피 세포를 감염시키며, 신경 세포에 잠복하는 특징을 가진다.알파헤르페스바이러스아과에는 다음 속들이 속해 있다.
- 일토바이러스속
- 마르디바이러스속
- 스쿠타바이러스속
- 심플렉스바이러스속
- 바리셀로바이러스속
종 ''이구아나 헤르페스바이러스 2''는 현재 속과 아과에 할당되지 않았다.[1]
2. 2. 베타헤르페스바이러스아과 (Betaherpesvirinae)
거대세포바이러스속, 무로메갈로바이러스속, 프로보시바이러스속, 쿠위바이러스속, 로제올로바이러스속이 있다.2. 3. 감마헤르페스바이러스아과 (Gammaherpesvirinae)
림프구, 특히 B 세포를 주로 감염시키며, 잠복하는 특징을 가진다.[1]감마헤르페스바이러스아과에는 다음 속들이 속한다.[1]
- 림포크립토바이러스속
- 마카바이러스속
- 퍼카바이러스속
- 라디노바이러스속
- 미할당
2. 4. 미할당
이구아나 헤르페스바이러스 2는 현재 속(genus)과 아과(subfamily)에 할당되지 않았다.[1]3. 구조
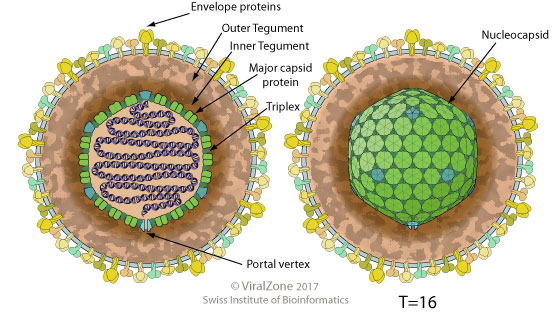
''헤르페스바이러스과''의 모든 구성원은 공통적인 구조를 가지고 있다. 상대적으로 크고 단일 분절이며, 이중 가닥의 선형 DNA 게놈은 캡시드라고 하는 정20면체 단백질 껍질(T=16 대칭) 안에 갇힌 100~200개의 유전자를 암호화한다. 캡시드는 바이러스 단백질과 바이러스 mRNA를 모두 포함하는 바이러스 외피라고 하는 단백질 층과 지질 이중층 막인 외피로 둘러싸여 있다. 이 전체 입자를 비리온이라고 한다.
전형적인 HSV 비리온의 구조적 구성 요소는 지질 이중층 외피, 외피, DNA, 당단백질 스파이크 및 핵캡시드이다. 4가지 구성 요소로 이루어진 헤르페스 단순 바이러스 비리온은 이중 가닥 DNA 게놈을 정이십면체 핵캡시드로 캡슐화한다. 그 주변에는 외피가 있다. 외피는 각 7nm 너비의 필라멘트를 포함하며, 구조화된 영역이 있는 비정질 층이다. 이것은 지단백질 외피로 덮여 있으며, 각 비리온에서 튀어나온 당단백질로 만들어진 스파이크가 있다. 이들은 바이러스의 직경을 225nm까지 확장할 수 있다. 스파이크가 없는 비리온의 직경은 약 186nm이다. 비리온의 외부 외피에는 최소한 두 개의 비당화 막 단백질이 있다. 또한 gB, gC, gD, gE, gG, gH, gI, gJ, gK, gL 및 gM을 포함한 11개의 당단백질이 있다. 외피는 26개의 단백질을 포함한다. 이들은 캡시드의 핵 및 기타 소기관으로의 수송, 초기 유전자 전사 활성화 및 mRNA 분해와 같은 역할을 한다. 정이십면체 핵캡시드는 꼬리파지목의 꼬리 파지의 핵캡시드와 유사하다. 이 캡시드는 150개의 헥손과 11개의 펜톤, 그리고 DNA가 캡시드 안으로 들어가고 나가는 것을 허용하는 포털 복합체로 구성된 161개의 캡소머를 가지고 있다.
4. 생활사
모든 헤르페스바이러스과는 핵 내에서 복제된다. 바이러스 DNA는 감염된 세포의 핵 내에서 전사되어 mRNA로 변환된다.[14]
감염은 바이러스 입자가 세포 표면에 특정 유형의 세포막 수용체 분자를 가진 세포와 접촉할 때 시작된다. 바이러스 외피 당단백질이 세포막 수용체에 결합한 후, 바이러스 입자가 내부로 들어가 해체되면서 바이러스 DNA가 세포 핵으로 이동할 수 있게 된다. 핵 내에서 바이러스 DNA의 복제와 바이러스 유전자의 전사가 일어난다.[14]
증상이 나타나는 감염 동안, 감염된 세포는 용해성 바이러스 유전자를 전사한다. 일부 숙주 세포에서는 소수의 잠복 관련 전사체 (LAT)라고 하는 바이러스 유전자가 축적된다. 이러한 방식으로 바이러스는 세포(그리고 숙주) 내에서 무기한으로 생존할 수 있다. 일차 감염은 종종 임상 질병의 자가 제한 기간을 동반하지만, 장기적인 잠복기는 증상이 없다.[14]
염색질 역학은 전체 헤르페스 바이러스 유전체의 전사 능력을 조절한다. 바이러스가 세포에 들어가면 세포 면역 반응은 세포를 보호한다. 세포는 바이러스 DNA를 히스톤 주위에 감싸고 염색질로 응축시켜 바이러스가 휴면 상태, 즉 잠복 상태가 되도록 한다. 세포가 실패하고 염색질이 느슨하게 묶여 있으면 바이러스 DNA는 여전히 접근 가능하다. 바이러스 입자는 유전자를 켜고 세포 기계를 사용하여 복제하여 재활성화하고 용해성 감염을 시작할 수 있다.[14]
잠복 바이러스의 재활성화는 대상 포진, 장미색 비강진과 같은 여러 질병과 관련이 있다. 활성화 후, 바이러스 유전자 전사는 LAT에서 여러 용해성 유전자로 전환되며, 이는 복제 및 바이러스 생산을 증가시킨다. 종종 용해성 활성화는 세포 사멸로 이어진다. 임상적으로, 용해성 활성화는 종종 미열, 두통, 인후통, 불쾌감, 발진과 같은 비특이적 증상의 출현과 부어오르거나 압통이 있는 림프절과 같은 임상 징후, 그리고 자연 살해 세포 수 감소와 같은 면역학적 소견을 동반한다.[14]
동물 모델에서 국소 외상 및 시스템 스트레스는 잠복 헤르페스바이러스 감염의 재활성을 유도하는 것으로 밝혀졌다. 단백질 합성을 일시적으로 중단하거나 저산소증과 같은 세포 스트레스 요인도 바이러스 재활성을 유도하기에 충분하다.[15]
5. 면역 회피
헤르페스바이러스는 평생 감염을 유지하는 능력이 있는데, 이는 면역 회피를 통해 가능하다. 헤르페스바이러스는 다양한 방법으로 면역 체계를 회피한다. 그중 하나는 사람의 인터류킨 10(hIL-10)을 모방하는 단백질을 암호화하는 것이고, 다른 하나는 감염된 세포에서 주요 조직 적합성 복합체 II (MHC II)의 발현을 줄이는 것이다.[18]
거대세포바이러스(CMV) 연구에 따르면, 바이러스성 인간 IL-10 유사체인 cmvIL-10은 프로 염증성 사이토카인 합성을 억제하는 데 중요하다. cmvIL-10 단백질은 hIL-10과 27%의 동일성을 가지며, hIL-10에서 사이토카인 합성 억제 기능 부위를 구성하는 아홉 개의 아미노산 중 하나의 보존된 잔기만 가지고 있다. 그러나 hIL-10과 cmvIL-10의 기능은 매우 유사하다. 둘 다 IFN-γ, IL-1α, GM-CSF, IL-6 및 TNF-α를 감소시키는데, 이들은 모두 프로 염증성 사이토카인이다. 또한 MHC I 및 MHC II를 감소시키고 HLA-G(비고전적 MHC I)를 증가시키는 역할을 한다. 이 두 가지 현상은 각각 세포 매개 면역 반응과 자연 살해 세포 반응을 억제하여 면역 회피를 가능하게 한다. hIL-10과 cmvIL-10의 유사성은 hIL-10과 cmvIL-10이 모두 동일한 세포 표면 수용체인 hIL-10 수용체를 사용한다는 사실로 설명될 수 있다. hIL-10과 cmvIL-10의 기능 중 한 가지 차이점은 hIL-10은 인간 말초 혈액 단핵 세포(PBMC)의 증식을 증가시키고 감소시키는 반면, cmvIL-10은 PBMC의 증식만 감소시킨다는 것이다. 이는 cmvIL-10이 이러한 세포에 대한 hIL-10의 자극 효과가 부족할 수 있음을 나타낸다.[18]
cmvIL-10은 Stat3 단백질의 인산화를 통해 기능한다. 원래는 이 인산화가 JAK-STAT 경로의 결과라고 생각되었다. 그러나 JAK가 실제로 Stat3를 인산화한다는 증거에도 불구하고, 그 억제는 사이토카인 합성 억제에 유의미한 영향을 미치지 않았다. 또 다른 단백질인 PI3K도 Stat3를 인산화하는 것으로 밝혀졌다. JAK 억제와 달리 PI3K 억제는 사이토카인 합성에 상당한 영향을 미쳤다. Stat3 인산화에서 PI3K와 JAK의 차이점은 PI3K는 S727 잔기에서 Stat3를 인산화하는 반면 JAK는 Y705 잔기에서 Stat3를 인산화한다는 것이다. 이러한 인산화 위치의 차이가 프로 염증성 사이토카인 합성을 억제하는 Stat3 활성화의 핵심 요소인 것으로 보인다. 실제로 PI3K 억제제를 세포에 첨가하면 사이토카인 합성 수준이 유의하게 회복된다. 사이토카인 수준이 완전히 회복되지 않는다는 사실은 cmvIL-10에 의해 활성화되어 사이토카인 시스템 합성을 억제하는 또 다른 경로가 있음을 나타낸다. 제안된 메커니즘은 cmvIL-10이 PI3K를 활성화하고, PI3K는 차례로 PKB(Akt)를 활성화하며, PKB는 mTOR을 활성화하고, mTOR은 S727 잔기에서 인산화를 위해 Stat3를 표적으로 할 수 있다는 것이다.[19]
헤르페스 바이러스가 면역 체계를 회피하는 또 다른 방법은 MHC I 및 MHC II의 발현을 줄이는 것이다. 이는 거의 모든 사람 헤르페스 바이러스에서 관찰된다. MHC I 및 MHC II의 하향 조절은 여러 가지 다른 메커니즘에 의해 발생할 수 있으며, 대부분 MHC가 세포 표면에 존재하지 않게 한다. 위에서 언급했듯이 한 가지 방법은 IL-10과 같은 바이러스성 케모카인 유사체를 이용하는 것이다. MHC를 하향 조절하는 또 다른 메커니즘은 새로 형성된 MHC를 소포체(ER)에 억류하는 바이러스 단백질을 암호화하는 것이다. MHC는 세포 표면에 도달할 수 없으므로 T 세포 반응을 활성화할 수 없다. MHC는 또한 프로테아좀 또는 리소좀에서 파괴되도록 표적이 될 수 있다. ER 단백질인 TAP 역시 MHC 하향 조절에 역할을 한다. 바이러스 단백질은 TAP를 억제하여 MHC가 바이러스 항원 펩타이드를 포착하는 것을 막는다. 이는 MHC의 적절한 접힘을 방지하므로 MHC는 세포 표면에 도달하지 못한다.[20]
6. 사람 헤르페스바이러스
사람이 감염되는 사람 헤르페스 바이러스(HHV)에는 다음과 같은 종류가 있다.[21][22][23]
- 단순 헤르페스 바이러스 속 (''Simplexvirus'')[30]
- * 학명: ''Human herpesvirus 1'' (HHV-1) = 일반명 단순 헤르페스 바이러스 1형 (HSV-1)
- * 학명: ''Human herpesvirus 2'' (HHV-2) = 일반명 단순 헤르페스 바이러스 2형 (HSV-2)
- * 학명: ''Human herpesvirus 3'' (HHV-3) = 일반명 수두-대상포진 바이러스 (VZV)[31]
- 림포크립토바이러스 속 (''Lymphocryptovirus'')
- * 학명: ''Human herpesvirus 4'' (HHV-4) = 일반명 엡스타인-바 바이러스 (EBV)[32]
- 거대세포바이러스 속 (''Cytomegalovirus'')
- * 학명: ''Human herpesvirus 5'' (HHV-5) = 일반명 사람 거대세포바이러스 (HCMV)[33]
- 로제올로바이러스 속 (''Roseolovirus'')
- * 학명: ''Human herpesvirus 6'' (HHV-6) = 일반명 사람 헤르페스 바이러스 6
- *: 1986년 미국 베스 이스라엘 디코니스 의료 센터의 Saira Salahuddin 등에 의해 발견되었고, 1988년 오사카 대학 의학부의 야마니시 코이치 등에 의해 HHV-6B가 돌발성 발진의 원인이라는 것이 보고되었다. 약물 과민성 증후군의 관련 요인으로 생각되고 있다.[34]
- * 학명: ''Human herpesvirus 7'' (HHV-7)[35]
- *: 1990년 미국 국립 보건원의 Niza Frenkel 등에 의해 보고되었고, 1994년 오사카 대학 의학부의 타나카 케이코 등에 의해 돌발성 발진의 원인이라는 것이 보고되었다.
- 라디노바이러스 속 (''Rhadinovirus'')
- * 학명: ''Human herpesvirus 8'' (HHV-8) = 일반명 카포시 육종 관련 헤르페스 바이러스 (KSHV)
7. 동물원성 헤르페스바이러스
마카크 원숭이에서 주로 발견되는 원숭이 B 바이러스(CeHV-1)는 사람에게 감염될 경우 치명적인 뇌척수염을 유발할 수 있다. 현재까지 약 25건의 인간 감염 사례가 보고되었으며, 치료하지 않으면 사망에 이를 수 있다.[25] 감염된 사람 중 16명이 뇌척수염으로 사망했고, 최소 4명은 심각한 신경학적 손상을 입고 생존했다.[25][26] 따라서 마카크 원숭이를 다루는 실험실 작업자는 바이러스 노출에 대한 증상 인식 및 조기 치료가 중요하다.[27]
쥐에서 발견되는 쥐 헤르페스바이러스 68(MuHV-4)은 감염된 쥐를 다루는 실험실 작업자에게서 동물원성 감염을 일으킬 수 있다.[29][28] 이 바이러스는 ELISA 검사에서 다른 헤르페스바이러스와의 항체 교차 반응을 일으켜 위양성 결과를 4배나 증가시킬 수 있다.[29]
8. 동물 헤르페스바이러스
동물 바이러스학에서 가장 잘 알려진 헤르페스바이러스는 ''알파헤르페스바이러스아과''에 속한다. 돼지 가성 광견병 바이러스(PrV) 연구는 돼지에서 오제스키병의 원인 물질로, 유전자 변형 백신을 이용한 동물 질병 통제의 선구적인 역할을 했다.[2][3] PrV는 현재 용해성 헤르페스바이러스 감염 중 기본적인 과정을 연구하고, 헤르페스바이러스 신경친화성의 분자적 기전을 밝히기 위한 모델로 광범위하게 연구되고 있다. 소 헤르페스바이러스 1형(BHV-1)은 소 전염성 비기관염 및 농포성 질염의 원인 물질로, 잠복기의 분자적 기전을 규명하기 위해 분석된다.[2][3] 조류 감염성 후두기관염 바이러스는 이 두 바이러스와 계통발생학적으로 거리가 멀며, ''알파헤르페스바이러스아과'' 내의 유사성과 다양성을 강조하는 역할을 한다.[2][3]
현재까지 포유류, 조류, 양서류, 파충류, 어류 등 많은 동물을 고유 숙주로 하는 150종 이상의 바이러스가 분리되어 있다.
- 소 헤르페스바이러스 1형(BHV-1)
- 칠면조 헤르페스바이러스 = 일반명 마레크병 바이러스(MDV)
9. 항바이러스제
- 아시클로비르
- 발라시클로비르
- 펜시클로비르 (일본 미승인)
- 팜시클로비르
- 비다라빈
- 아이오도데옥시우리딘
- 소리부딘 (승인 취하)
- 브리부딘
거대세포바이러스 참고.
10. 연구 동향
현재 헤르페스바이러스와 관련된 다양한 부작용 또는 동반 질환에 대한 연구가 진행 중이다. 여기에는 다음 질환들이 포함된다.
참조
[1]
서적
Sherris Medical Microbiology
McGraw Hill
[2]
서적
Animal Viruses: Molecular Biology
Caister Academic Press
[3]
서적
Alpha Herpesviruses: Molecular and Cellular Biology
http://www.horizonpr[...]
Caister Academic Press
[4]
논문
The Origin and the Use of the Word Herpes
1962
[5]
논문
Classification and nomenclature of viruses. First report of the International Committee on Nomenclature of Viruses.
1971
[6]
웹사이트
Virus Taxonomy: 2020 Release
https://ictv.global/[...]
International Committee on Taxonomy of Viruses (ICTV)
2021-03
[7]
논문
Herpes simplex
2009-04
[8]
웹사이트
In the United States, as many as 15% of adults between 35 and 72 years of age have been infected.
https://www.cdc.gov/[...]
National Center for Infectious Diseases
[9]
논문
Seroprevalence of cytomegalovirus infection in the United States, 1988–1994
2006-11
[10]
서적
Virology, Principles and Applications
John Wiley & Sons
2007-08-15
[11]
논문
Herpesvirus capsid assembly: insights from structural analysis
2011-08
[12]
논문
Cryo-EM structures of herpes simplex virus type 1 portal vertex and packaged genome
https://doi.org/10.1[...]
2019
[13]
서적
Herpesvirus Polymerase Inhibitors
https://doi.org/10.1[...]
Elsevier
2019
[14]
논문
Chromatin dynamics and the transcriptional competence of HSV-1 genomes during lytic infections
2019-11
[15]
논문
Herpesviruses: latency and reactivation – viral strategies and host response
2013-10
[16]
논문
Molecular phylogeny and evolutionary timescale for the family of mammalian herpesviruses
1995-03
[17]
논문
Topics in herpesvirus genomics and evolution
2006-04
[18]
논문
Potent immunosuppressive activities of cytomegalovirus-encoded interleukin-10
2002-02
[19]
논문
The cytomegalovirus homolog of interleukin-10 requires phosphatidylinositol 3-kinase activity for inhibition of cytokine synthesis in monocytes
http://repository.us[...]
2007-02
[20]
논문
Modulation of HLA expression in human cytomegalovirus immune evasion
2007-04
[21]
논문
Ratification vote on taxonomic proposals to the International Committee on Taxonomy of Viruses (2012)
2012-07
[22]
서적
Herpesviruses. ''in:'' Baron's Medical Microbiology
https://www.ncbi.nlm[...]
Univ of Texas Medical Branch
[23]
서적
Medical Microbiology
Elsevier Mosby
[24]
논문
Infectious mononucleosis-like syndromes in febrile travelers returning from the tropics
[25]
논문
Biology of B virus in macaque and human hosts: a review
1992-02
[26]
논문
B-virus (Cercopithecine herpesvirus 1) infection in humans and macaques: potential for zoonotic disease
2003-02
[27]
웹사이트
Herpes-B Fact Sheet
http://dcminfo.wustl[...]
[28]
논문
Conquering the Host: Determinants of Pathogenesis Learned from Murine Gammaherpesvirus 68
2021-09-29
[29]
논문
Murine gammaherpesvirus 68 serum antibodies in general human population
[30]
논문
単純ヘルペスウイルス(HSV)
https://doi.org/10.2[...]
[31]
논문
水痘帯状疱疹ウイルス(VZV)
https://doi.org/10.2[...]
[32]
논문
Epstein-Barrウイルス(EBV)とカポジ肉腫関連ヘルペスウイルス(KSHV, HHV-8)
https://doi.org/10.2[...]
[33]
논문
ヘルペスウイルス(HHV1‐8)のウイルス学 3.サイトメガロウイルス(CMV)
https://doi.org/10.2[...]
日本ウイルス学会
[34]
논문
薬剤性過敏症症候群とHHV-6の再活性化について
https://doi.org/10.2[...]
[35]
논문
ヒトヘルペスウイルス6とヒトヘルペスウイルス7(HHV-6HHV-7)
https://doi.org/10.2[...]
[36]
서적
Sherris Medical Microbiology
https://archive.org/[...]
McGraw Hill
[37]
서적
Animal Viruses: Molecular Biology
Caister Academic Press
[38]
서적
Alpha Herpesviruses: Molecular and Cellular Biology
http://www.horizonpr[...]
Caister Academic Press
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com