도에디쿠루스
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
도에디쿠루스는 멸종된 글립토돈트의 일종으로, 몸길이가 최대 4m에 달하며, 꼬리 끝에 가시가 달린 뼈 덩어리를 가진 것이 특징이다. 현대 아르마딜로와 가장 가까운 친척으로, 남미에 고유한 빈치류의 일원이었으며, 약 3,400만 년 전 아르마딜로와 분기되었다. 초식 동물로, 꼬리 곤봉을 이용하여 방어하거나 종내 경쟁을 했을 것으로 추정된다. 플라이스토세와 홀로세에 걸쳐 생존했으며, 인간의 사냥과 기후 변화로 인해 제4기 대멸종 때 멸종된 것으로 보인다.
더 읽어볼만한 페이지
- 고대 빈치류 - 글립토돈
남아메리카에 서식했던 글립토돈은 갑옷 같은 등갑과 꼬리 갑옷을 가진 거대한 멸종 포유류로, 리처드 오웬에 의해 명명되었으며 기후 변화와 인간의 영향으로 멸종했을 것이라는 가설이 있다. - 고대 빈치류 - 땅늘보
땅늘보는 올리고세에 등장하여 마이오세에 다양성이 증가한 멸종된 동물로, 크기는 100kg 미만에서 4000kg 이상까지 다양하며, 초식성이지만 일부는 잡식성이었을 가능성이 있고, 인간의 사냥으로 인해 약 11,000년 전에 멸종되었다. - 남아메리카의 고대 포유류 - 메가테리움
메가테리움은 최대 6m에 달하는 멸종된 거대한 지상 나무늘보로, 1,200만 년 전 파타고니아에서 처음 나타나 초식 생활을 했으며 플라이스토세 말에 멸종되었다. - 남아메리카의 고대 포유류 - 땅늘보
땅늘보는 올리고세에 등장하여 마이오세에 다양성이 증가한 멸종된 동물로, 크기는 100kg 미만에서 4000kg 이상까지 다양하며, 초식성이지만 일부는 잡식성이었을 가능성이 있고, 인간의 사냥으로 인해 약 11,000년 전에 멸종되었다. - 1874년 기재된 화석 분류군 - 폴리오낙스
폴리오낙스는 1873년 에드워드 드링커 코프가 명명한 각룡류 공룡의 한 속으로, 불분명한 분류의 각룡류로 여겨지며, 뿔과 프릴을 가진 사족보행 초식 공룡으로 추정된다. - 1874년 기재된 화석 분류군 - 마우이사우루스
마우이사우루스는 뉴질랜드에서 발견된 긴 목을 가진 멸종된 해양 파충류로, 현재는 유효성에 의문이 제기되어 무효명으로 취급되지만 뉴질랜드 문화적으로 특별한 의미를 가진다.

2. 분류 및 진화

도에디쿠루스는 글립토돈트의 일종으로, 19세기와 20세기를 거치면서 다양한 종과 속이 기술되어 총 65개 속과 220개 종에 달했다.[3][4] 글립토돈트는 과 글립토돈티데에 분류되었다.
2022년 Barasoain 외 연구진은 글립토돈트의 분기도를 제시했다.[8]
2. 1. 진화 분기도
이 동물은 1847년 영국의 고생물학자 리처드 오언에 의해 처음 기술되었으며, ''글립토돈 클라비페스'' 등에 이어 기술된 다섯 번째 글립토돈트 종이었다. 모식 표본은 거대한 곤봉을 나타내는 것으로 보이는 부분적인 꼬리였으며, 오언은 ''G. 클라비카우다투스''(종명은 "곤봉 꼬리"를 의미하는 라틴어에서 유래)라는 이름을 붙였다. 1874년, 독일의 동물학자 헤르만 부르마이스터는 이를 ''도에디쿠루스 클라비카우다투스''로 자체 속으로 분류했으며, 속명은 고대 그리스어의 δοῖδυξ "절구"와 oυρά "꼬리"에서 유래했다.[2]''도에디쿠루스''는 현대의 아르마딜로와 가장 밀접하게 관련된 글립토돈트였으며, 남아메리카에 고유종인 상목 빈치류(나무늘보와 개미핥기와 함께)의 일원이었다. 1997년, 말콤 C. 맥케나와 수잔 K. 벨은 모든 글립토돈트를 상과 글립토돈토이데아에 할당했으며, 여기에는 팜파테리과, 팔레오펠타과, 글립토돈티데과가 포함되었다. ''도에디쿠루스''는 아과 도에디쿠리네에 분류되었다.[5]
2016년, 12,000년 된 ''도에디쿠루스'' 표본의 등갑에서 고대 DNA가 추출되었고, 거의 완전한 미토콘드리아 유전체가 재구성되었다(76x 커버리지). 현대 아르마딜로의 유전체와 비교한 결과, 글립토돈트가 약 3,400만 년 전 프리아보니아절인 후기 에오세에 톨리페우티나 및 클라미포리나 아르마딜로와 분기된 것으로 나타났다.[7][6] 이로 인해 자체 과인 글립토돈티데에서 현존하는 클라미포리데 내 아과인 글립토돈티네으로 이동하게 되었다.[6] 이를 바탕으로 화석 기록을 보면, 글립토돈트는 냉각, 건조한 기후와 열린 사바나의 확장에 대한 반응으로 특징적인 형태와 큰 크기(거대증)를 상당히 빠르게 진화했을 것이다.[7]
3. 형태
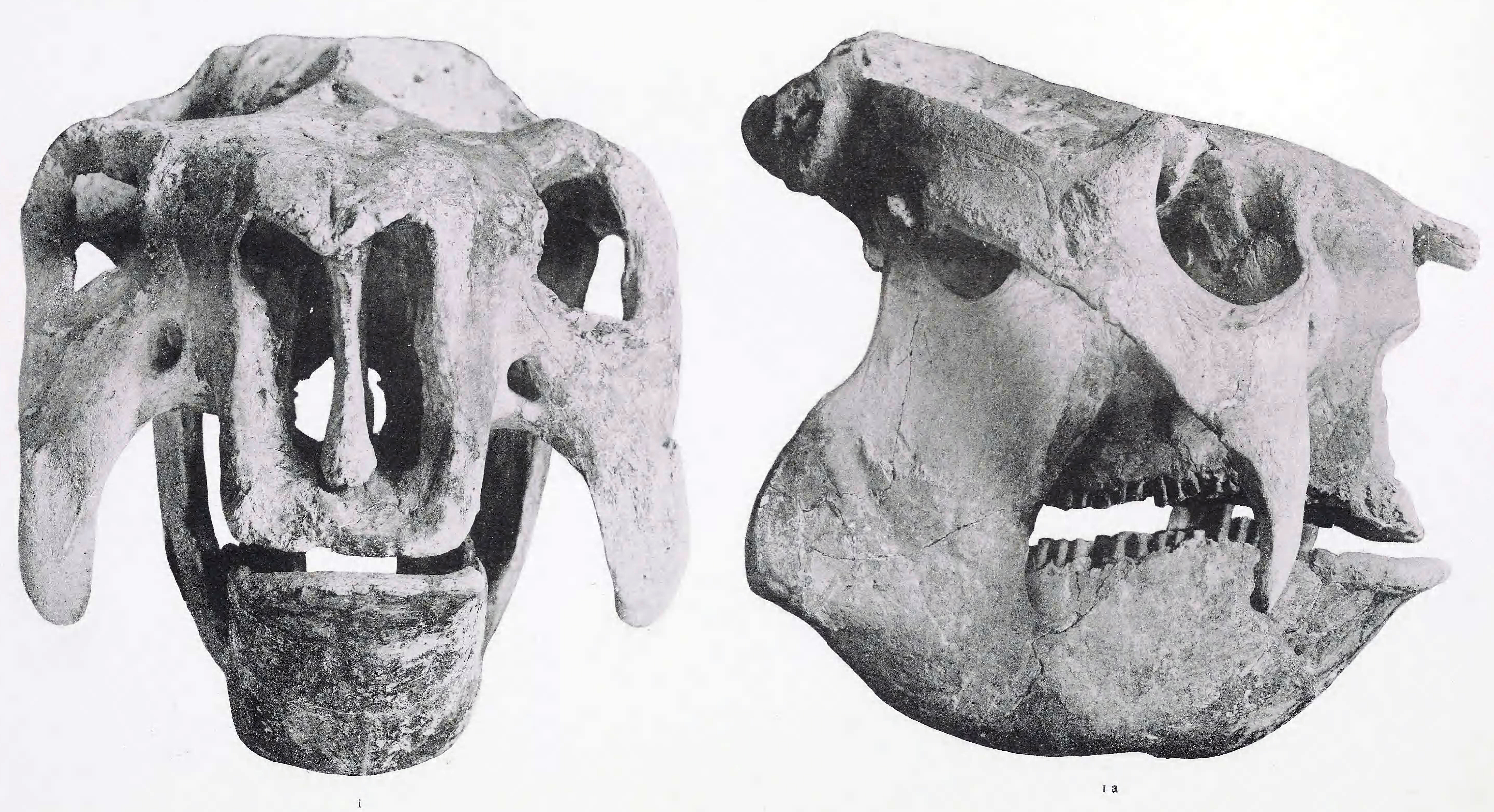
홍적세 후기에 서식했으며, 글립토돈과에서는 최대급의 동물이다. 몸길이는 최대 4m에 달하며, 등갑 뒤쪽에 혹이 있는 것이 특징이다. 꼬리는 막대 모양이며, 끝부분에는 가시 모양의 돌기가 여러 개 존재했다.
''도에디쿠루스''는 평균적으로 키가 1.5m, 전체 길이는 약 3.6m이며,[10] 몸무게는 약 1400kg였다. 8,000년 된 표본은 1900kg에서 2370kg으로 계산되었는데, 이는 멸종 직전인 홀로세에 ''도에디쿠루스''가 훨씬 더 커졌을 수 있음을 나타낸다. ''Pa. intermedius'', ''Pa. subintermedius'', ''G. munizi'', ''G. elongatus'', ''Plaxhaplous''와 함께 알려진 가장 무거운 글립토돈 종 중 하나이다.[11]
''도에디쿠루스''는 오늘날의 친척인 아르마딜로와 다소 유사한, 촘촘하게 맞물린 많은 골판으로 만들어진 거대한 돔 형태의 등갑을 가지고 있었다. 등갑은 골반에 단단히 고정되어 있었지만 어깨 주위는 느슨했다. 등갑에는 낙타의 혹과 유사한 지방이 채워진 공간이 있었을 것으로 추정된다.[1]
글립토돈트와 빈치류처럼, 질량 중심은 앞다리보다 뒷다리에 더 가까웠던 것으로 보이며, 이는 무게의 대부분이 뒷다리에 실렸음을 나타낸다.[13][14]
3. 1. 이빨
검치목은 고치형 치아를 가지고 있으며, 평생 동안 이가 자라기를 멈추지 않아 주로 풀을 먹고 살았을 것으로 추정된다. 다른 포유류의 치아와 비교하여 특이한 점이 있는데, 처음 두 개의 치아를 제외하고 세 개의 엽(lobe)이 있다는 것이다. 치아의 핵심은 골상아질로 이루어져 있으며, 정상아질 층으로 둘러싸여 있고, 법랑질 대신 백악질로 덮여 있다. 시간이 지남에 따라 백악질이 마모되면서 일부 정상아질이 노출되어, 말과 소의 이빨에서 법랑질을 통해 단단한 상아질과 백악질이 결국 튀어나오는 것과 유사하게, 풀을 더 잘 처리할 수 있는 줄 모양의 표면을 만들어낸다.[9]검치목은 8개의 볼 치아를 가지고 있으며, 소와 마찬가지로 송곳니와 앞니가 완전히 없다. ''도에디쿠루스''와 다른 큰 검치목은 눈에 띄게 좁아진 입을 가지고 있었으며, 치아의 연마 면이 비교적 작아 음식을 완전히 씹을 수 없었음을 나타낸다. 이는 이 부위의 갑옷이 점점 더 무거워지면서 머리와 목을 지탱하는 근육의 크기가 커지고, 씹는 근육이 기계적으로 덜 효율적인 위치로 이동했기 때문일 수 있다. 풀을 완전히 갈아서 영양 흡수를 극대화하는 것이 매우 중요한데, 이러한 비효율성은 느린 신진대사를 나타낼 수 있다는 점에서 이상하다. 겉보기에 강한 혀는 불완전하게 씹힌 음식을 부분적으로 재가공하여 위 또는 아마도 맹장으로 밀어 넣었을 수 있다.[9]
3. 2. 몸통
''도에디쿠루스''는 평균적으로 키가 1.5m, 전체 길이는 약 3.6m,[10] 몸무게는 약 1400kg였다. 8,000년 된 표본은 1900kg 에서 2370kg으로 계산되었는데, 이는 멸종 직전인 홀로세에 ''도에디쿠루스''가 훨씬 더 커졌을 수 있음을 나타낸다. ''Pa. intermedius'', ''Pa. subintermedius'', ''G. munizi'', ''G. elongatus'', ''Plaxhaplous''와 함께 알려진 가장 무거운 글립토돈 종 중 하나이다.[11] ''도에디쿠루스''는 오늘날의 친척인 아르마딜로와 다소 유사한, 촘촘하게 맞물린 많은 골판으로 만들어진 거대한 돔 형태의 등갑을 가지고 있었다. 등갑은 골반에 단단히 고정되어 있었지만 어깨 주위는 느슨했다. 등갑은 낙타의 혹과 유사한 지방이 채워진 공간일 수 있는 돔 형태를 특징으로 했다.[1]글립토돈에 비해 긴 꼬리 끝에는 중세 시대의 무기인 모닝스타를 연상시키는 가시가 달린 뼈 덩어리를 가지고 있어, 포식자에게 이것을 사용하여 적극적인 방어 수단을 취했다고 생각된다.
3. 3. 꼬리
''도에디쿠루스''의 꼬리는 유연한 뼈로 둘러싸여 있었고, 가장자리를 따라 얕은 함몰부가 있었는데, 이는 살아있을 때 가시였을 수 있다.[1] 꼬리 곤봉은 길이가 최대 1m에 달할 수 있었다. 최대 변형률이 0.25(전형적인 척추동물)이고, 응력 발휘가 3x105 N m−2(최근 죽은 동물의 근육에서 측정된 것을 기반으로 함)이며, 꼬리 근육의 부피가 100L라고 가정하면, ''도에디쿠루스''는 약 2.5kJ의 타격을 가할 수 있었을 것이지만, 이는 과소평가일 수 있다. 곤봉의 전체 질량이 살아있을 때 40kg이라고 가정하면, 이는 최대 속도 11m/s와 같다.[12] 꼬리 끝은 15m/s에 달했을 수 있다. 곤봉이 살아있을 때 65kg이라고 가정하면, 타격점 (최대 힘을 발휘하고 자체 손상을 최소화했을 곤봉의 충격 지점)은 끝에서 약 77cm였을 것이다.[17]글립토돈에 비해 긴 꼬리 끝에는 중세 시대의 무기인 모닝스타를 연상시키는 가시가 달린 뼈 덩어리를 가지고 있다. 포식자에게는 이것을 사용하여 적극적인 방어 수단을 취했다고 생각된다.
3. 4. 다리
다른 글립토돈트와 빈치류처럼, 질량 중심은 앞다리보다 뒷다리에 더 가까웠던 것으로 보이며, 이는 무게의 대부분, 어떤 경우에는 거의 전부가 뒷다리에 실렸음을 나타낸다.[13][14] 현대 빈치류는 방어, 관찰 또는 먹이를 먹기 위해 흔히 이런 식으로 일어선다. 튼튼한 뒷다리는 또한 꼬리 곤봉을 가속하고 타격을 받은 후 자세를 유지하는 데 중요했을 것이다.[14]그럼에도 불구하고, 글립토돈트는 또한 강력한 앞팔을 가지고 있었다. 앞다리가 무게를 지탱할 필요가 없었기 때문에, 이것들은 현대 아르마딜로처럼 팠을 수 있지만, 등딱지와 척추는 아르마딜로보다 훨씬 더 뻣뻣했다. 또는, 앞다리는 꼬리 곤봉을 휘두르기 위해 몸을 회전시키는 동안 사용되었을 수 있다.[14] 더 작았던 초기의 글립토돈트가 비슷한 무게 분포를 공유하지 않기 때문에, 두 발로 서는 자세를 채택한 것은 몸 크기가 커지는 것과 관련이 있을 수 있다.[13][14]
4. 생태
아메리카 대교환은 약 280만 년 전 파나마 지협이 형성된 후, 남아메리카가 오랫동안 고립되었던 시기를 끝내고 북아메리카 종들의 침입을 받으면서 시작되었다. 이로 인해 글립토돈트는 단두 곰, ''스밀로돈''과 ''호모테리움''과 같은 검치호, 재규어 등 새로운 대형 포유류 육식동물을 만나게 되었다.[7][1] 이들은 세베코수스 악어, 마드소이드 뱀, 테러새, 유대류와 유사한 스파라소돈트 등 이전의 고유한 최상위 포식자를 대체했다.[18] 남아메리카로 이주해 온 다른 종으로는 말, 낙타, 사슴, 코끼리 (곰포테리움), 맥, 신세계 쥐 등이 있었다. 플라이스토세 시대 남아메리카 토착 포유류에는 글립토돈트, 지상늘보, 개미핥기, 아르마딜로와 같은 이빨없는 포유류, 유대류, 대형 톡소돈과 뉴월드호저 같은 토착 설치류가 있었다.[19]
''도에디쿠루스''는 플라이스토세 시대의 글립토돈트 속 중에서 ''글립토돈'', ''네오스클레로칼립투스'', ''호플로포루스'', ''뉴리우루스'', ''파노크투스''와 함께 가장 흔하게 확인되는 속에 속한다.[20] 글립토돈트는 일반적으로 온대에서 서늘한 기후의 개방된 초원에 서식했으며,[1] 차코-팜파 평원 북동부 파타고니아의 춥고 습한 지역에 제한적으로 서식했던 것으로 보인다.[21] 화석은 아르헨티나, 브라질, 우루과이에서 발견되었다.[22] 플라이스토세는 빙하기와 간빙기의 주기적인 변화로 특징지어졌으며, 파타고니아의 층서는 15번 이상의 빙하 주기를 기록하는데, 이는 뢰스 (빙하기 동안 퇴적)에서 고토양 (간빙기 동안)으로의 전환을 통해 나타난다.[23] 빙하기에는 사바나가 증가했고, 간빙기(오늘날 포함)에는 열대 우림이 확장되는 특징을 보였다.[19]
4. 1. 포식자와의 관계
''도에디쿠루스''는 초식 동물로 여겨지며, 높은 고치상 치아와 넓은 주둥이는 대량 섭식을 했음을 나타낼 수 있다.[21]글립토돈트 종은 아메리카 대교환 이후 이전에 고립되었던 대륙으로 새로운 포유류가 이주하면서 특히 크기가 커졌으며, ''도에디쿠루스''를 포함한 가장 큰 글립토돈트 중 일부는 이 사건 이후의 플라이스토세에서 알려져 있다. 이는 증가하는 거대화가 새로운 포유류 육식 동물에 대한 포식자 방어 적응이었음을 나타낼 수 있다.[7][1] ''스밀로돈''이 ''도에디쿠루스''를 사냥했다는 증거가 있다.[15] 후기 플라이스토세와 홀로세에 크기가 극적으로 증가했는데, 이는 기후 냉각에 대한 반응(이는 신진대사를 감소시켜 크기를 증가시켰을 것)이거나 최근 이주해 온 인간 사냥꾼에 대한 방어 때문일 수 있다.[1]
그러나 갑옷과 체중의 증가는 ''도에디쿠루스'' 개체 간의 싸움에서 종내 경쟁에 의해 주로 유발되었을 수 있다. 만약 그렇다면, 수컷이 암컷보다 훨씬 더 튼튼하게 만들어졌을 것이다. 꼬리 곤봉의 충격으로 계산된 힘과 일치하는 등갑 파편의 증거가 발견되었다. ''도에디쿠루스''의 시력은 포식자 방어에 꼬리 곤봉을 사용하기에는 너무 좋지 않았을 수 있다.[16] 곤봉으로 표적을 타격하는 데 필요한 정확성은 정지된 상대를 통해서만 얻을 수 있었을 수 있으며, 이는 포식자 방어보다는 의례적인 전투에서의 사용을 뒷받침한다.[17]
글립토돈에 비해 긴 꼬리 끝에는 중세 시대의 무기인 모닝스타를 연상시키는 가시가 달린 뼈 덩어리를 가지고 있었다. 포식자에게는 이것을 사용하여 적극적인 방어 수단을 취했다고 생각된다.
4. 2. 경쟁
''도에디쿠루스''는 초식 동물로 여겨지며, 높은 수준의 고치상 치아와 넓은 주둥이는 대량 섭식을 했음을 나타낼 수 있다.[21]글립토돈트 종은 아메리카 대교환 이후 이전에 고립되었던 대륙으로 새로운 포유류가 이주하면서 특히 크기가 커졌으며, ''도에디쿠루스''를 포함한 가장 큰 글립토돈트 중 일부는 이 사건 이후의 플라이스토세에서 알려져 있다. 이는 증가하는 거대화가 새로운 포유류 육식 동물에 대한 포식자 방어 적응이었음을 나타낼 수 있다.[7][1] ''스밀로돈''이 ''도에디쿠루스''를 사냥했다는 증거가 있다.[15] 후기 플라이스토세와 홀로세에 크기가 극적으로 증가했는데, 이는 기후 냉각에 대한 반응(이는 신진대사를 감소시켜 크기를 증가시켰을 것)이거나 최근 이주해 온 인간 사냥꾼에 대한 방어 때문일 수 있다.[1]
그러나 갑옷과 체중 증가는 ''도에디쿠루스'' 개체 간의 싸움에서 종내 경쟁에 의해 주로 유발되었을 수 있다. 만약 그렇다면, 수컷이 암컷보다 훨씬 더 튼튼하게 만들어졌을 것이다. 꼬리 곤봉의 충격으로 계산된 힘과 일치하는 등갑 파편의 증거가 발견되었다. ''도에디쿠루스''의 시력은 포식자 방어에 꼬리 곤봉을 사용하기에는 너무 좋지 않았을 수 있다.[16] 곤봉으로 표적을 타격하는 데 필요한 정확성은 정지된 상대를 통해서만 얻을 수 있었을 수 있으며, 이는 포식자 방어보다는 의례적인 전투에서의 사용을 뒷받침한다.[17]
5. 서식 환경

약 280만 년 전 파나마 지협이 형성된 후, 남아메리카는 오랫동안 세계와 격리되었던 시기를 끝내고 북아메리카 종들의 침입을 받았다. 이는 대규모 아메리카 교환의 일부였다. 당시 글립토돈트는 단두 곰, ''스밀로돈''과 ''호모테리움''과 같은 검치호, 재규어와 같은 새로운 대형 포유류 육식동물을 만났을 것이다.[7][1] 이들은 이전의 고유한 최상위 포식자인 세베코수스 악어, 마드소이드 뱀, 테러새, 유대류와 유사한 스파라소돈트를 대체했다.[18] 곰과 고양이 외에도 남아메리카로 이주해 온 다른 종으로는 말, 낙타, 사슴, 코끼리 (곰포테리움), 맥, 신세계 쥐가 있다. 플라이스토세 시대의 남아메리카 토착 포유류에는 글립토돈트, 지상늘보, 개미핥기, 아르마딜로와 같은 이빨없는 포유류가 있었으며, 유대류, 대형 톡소돈과 뉴월드호저와 같은 토착 설치류도 있었다.[19]
''도에디쿠루스''는 플라이스토세 시대의 글립토돈트 속 중에서 ''글립토돈'', ''네오스클레로칼립투스'', ''호플로포루스'', ''뉴리우루스'', ''파노크투스''와 함께 가장 흔하게 확인되는 속에 속한다.[20] 글립토돈트는 일반적으로 온대에서 서늘한 기후의 개방된 초원에 서식했다.[1] 차코-팜파 평원 북동부 파타고니아의 춥고 습한 지역에 제한적으로 서식했던 것으로 보인다.[21] 화석은 아르헨티나, 브라질, 우루과이에서 발견되었다.[22] 플라이스토세는 빈번한 한랭/온난 주기(빙하기와 간빙기)로 특징지어졌으며, 파타고니아의 층서는 15번 이상의 빙하 주기를 기록하는데, 이는 뢰스 (빙하기 동안 퇴적)에서 고토양 (간빙기 동안)으로의 전환을 통해 나타난다.[23] 빙하기에는 사바나가 증가했을 수 있으며, 간빙기(오늘날 포함)에는 열대 우림이 확장되는 특징을 보였다.[19]
6. 멸종
''도에디쿠루스''는 가장 최근까지 생존한 글립토돈트 종일 수 있으며, 가장 늦은 화석은 팜파스에서 약 8,000~7,000년 전으로 추정된다. 그러나 G. claviceps 표본은 약 4,300년 전으로 연대가 추정되어 논란이 있다.[1][24] 2019년 연구에 따르면 팜파스 지역의 이러한 홀로세 연대는 부식산에 의한 오염으로 인해 과소평가되었으며, 후기 플라이스토세에 해당할 가능성이 더 높다.[25]
''도에디쿠루스''는 전 세계의 다른 많은 대형 동물과 마찬가지로, 인간의 과도한 사냥과 기후 변화의 복합적인 요인으로 인해 발생했을 가능성이 있는 제4기 대멸종 때 멸종되었다. 아르헨티나 라 모데르나 유적지의 늪지 가장자리에서 발견된 약 7,500~7,000년 전으로 거슬러 올라가는 도살된 표본은 ''도에디쿠루스''가 아메리카 대륙 정착의 남아메리카 최초의 인간 정착민들에 의해 사냥되었고, 그들과 수천 년 동안 공존했음을 보여준다. 연대가 정확하다면 팜파스는 대륙 다른 곳에서 쇠퇴했을 가능성이 있는 생산적인 초원을 제공하는 생물 피난처 역할을 했을 수 있는데, 다른 많은 남아메리카 대형 동물(예: 아르마딜로 ''에우타투스'', 거대한 땅늘보 ''메가테리움'', 개 ''두시키온 아부스'')도 이 지역에서 플라이스토세 종료 후 얼마 동안 생존했던 것으로 보이기 때문이다.[26] 이들의 최종적인 멸망은 인간의 사냥에 의해 촉발되었거나 단순히 가속화되었을 수 있다.[27] 그러나 이후 연구에서는 라 모데르나의 ''도에디쿠루스''의 늦은 연대와 다른 팜파스 지역의 홀로세로 추정되는 다른 대형 동물들의 연대가 오염 또는 기타 오류로 인해 발생했을 가능성이 있어, 이들의 홀로세 생존에 대한 의문을 제기했다.[28]
참조
[1]
서적
Catalogue of the Fossil Mammalia in the British Museum, (Natural History): The group Tillodontia, the orders Sirenia, Cetacea, Edentata, Marsupialia, Monotremata, and Supplement
https://google.com/b[...]
Natural History Museum, London
[2]
서적
Megafauna: Giant Beasts of Pleistocene South America
Indiana University Press
[3]
논문
On the taxonomic status of some Glyptodontidae (Mammalia, Xenarthra, Cingulata) from the Pleistocene of South America
[4]
논문
The phylogenetic affinities of the extinct glyptodonts
[5]
서적
Classification of Mammals: Above the Species Level
Columbia University Press
[6]
논문
The phylogenetic affinities of the extinct glyptodonts
[7]
논문
Ancient DNA from the extinct South American giant glyptodont ''Doedicurus'' sp. (Xenarthra: Glyptodontidae) reveals that glyptodonts evolved from Eocene armadillos
http://sedici.unlp.e[...]
[8]
논문
A New Glyptodont (Xenarthra: Cingulata) from the Late Miocene of Argentina: New Clues About the Oldest Extra-Patagonian Radiation in Southern South America
https://link.springe[...]
2022-06
[9]
논문
Carved teeth and strange jaws: How glyptodonts masticated
https://www.app.pan.[...]
[10]
서적
History of Terrestrial Mammals in South America
Springer
[11]
논문
Un Glyptodontidae de gran tamaño en el Holoceno temprano de la Región Pampeana, Argentina
http://www.sbpbrasil[...]
[12]
논문
Mechanics of fighting
[13]
논문
Limb bone strength and habits in large glyptodonts
[14]
논문
Proportions and function of the limbs of glyptodonts
[15]
논문
New Late Pleistocene megafaunal assemblage with well-supported chronology from the Pampas of southern South America
[16]
논문
Tail blow energy and carapace fractures in a large glyptodont (Mammalia, Xenarthra)
1999-05
[17]
논문
The sweet spot of a biological hammer: the centre of percussion of glyptodont (Mammalia: Xenarthra) tail clubs
[18]
논문
The Evolution of the Cenozoic Terrestrial Mammalian Predator Guild in South America: Competition or Replacement?
[19]
논문
The Great American Biotic Interchange: Dispersals, Tectonics, Climate, Sea Level and Holding Pens
[20]
논문
Accessory protection structures in Glyptodon Owen (Xenarthra, Cingulata, Glyptodontidae)
http://sedici.unlp.e[...]
[21]
논문
Potential Distribution of Fossil Xenarthrans in South America during the Late Pleistocene: co-Occurrence and Provincialism
[22]
웹사이트
''Doedicurus'' in the Paleobiology Database
https://paleobiodb.o[...]
2021-12-17
[23]
서적
Paleoenvironments: Vertebrates and Invertebrates
[24]
논문
Megafauna extinction in South America: A new chronology for the Argentine Pampas
https://www.research[...]
[25]
논문
Campo Laborde: A Late Pleistocene giant ground sloth kill and butchering site in the Pampas
2019-03
[26]
논문
Gliptodontes y Cazadores-Recolectores de la Region Pampeana (Argentina)
https://www.research[...]
[27]
논문
The Pleistocene-Holocene transition and human occupations in the Southern Cone of South America
[28]
논문
Site formation process and megamammal bone radiocarbon dates in Campo Laborde (Pampas of Argentina): Contribution towards a research methodology
https://linkinghub.e[...]
2021-06
[29]
논문
Shotgun Mitogenomics Provides a Reference Phylogenetic Framework and Timescale for Living Xenarthrans
https://www.ncbi.nlm[...]
2016-03
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com