척추동물
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
척추동물은 척삭동물 문에 속하는 동물들의 아문으로, 척추 또는 척주라고 하는 강하고 유연한 골격 구조를 가지고 있으며, 좌우대칭을 이룬다. 척추동물은 몸의 구조, 골격, 기관계 등이 다양하게 분화되어 있으며, 캄브리아기 대폭발 시기에 기원하여 어류, 양서류, 파충류, 조류, 포유류 등으로 진화해 왔다. 척추동물은 턱의 유무, 지느러미나 다리의 발달, 양막의 유무 등 다양한 특징에 따라 여러 강으로 분류되며, 전통적인 분류 외에도 계통 발생학적 분류가 사용된다. 최근 멸종 위기에 처한 종이 증가하고 있으며, 서식지 파괴, 기후 변화, 남획 등이 주요 원인으로 꼽힌다.
더 읽어볼만한 페이지
- 척추동물 - 유인원
유인원은 꼬리 없는 영장류로, 인간을 포함하여 긴팔원숭이과와 사람과로 나뉘며, 높은 지능과 복잡한 사회 구조를 공유하지만 서식지 파괴와 사냥으로 멸종 위기에 처해 있다. - 척추동물 - 양막류
양막류는 파충류, 조류, 포유류를 포함하는 척추동물 분류군으로, 양막을 형성하며 육상 번식에 중요한 진화적 도약을 이루었고, 배외막, 두개골 구조, 번식 방식에 따라 다양한 분류로 나뉜다. - 생물 계통도에 관한 - 눈표범
눈표범은 고양이과 동물로 중앙아시아와 남아시아의 고산 지대에 서식하며, 길고 두꺼운 털, 큰 발, 긴 꼬리가 특징이며, 유전자 분석 결과 호랑이와 가장 가까운 종으로 밝혀졌으며, 밀렵과 서식지 파괴로 인해 개체 수가 감소하여 여러 국가에서 보호 노력이 진행 중이다. - 생물 계통도에 관한 - 벵골호랑이
벵골호랑이는 인도아대륙에서 서식하는 호랑이 아종으로, 노란색~주황색 털과 검은 줄무늬를 가지며, 멧돼지, 사슴 등 우제류를 먹이로 하며, 서식지 파괴와 밀렵으로 인해 제한적인 지역에서 서식한다. - 생물 분류에 관한 - 다람쥐
다람쥐는 등 쪽에 줄무늬가 있는 다람쥐속 설치류로, 홀로 생활하며 겨울잠을 자고 씨앗, 견과류, 곤충 등을 먹으며 맹금류 등의 먹이가 되고, 특히 한반도 서식 다람쥐는 새로운 종으로 분류될 가능성이 있으며, 유럽에서는 침입 외래종으로 지정되기도 하고 라임병을 옮길 수 있어 주의가 필요한 동물이다. - 생물 분류에 관한 - 황금랑구르
황금랑구르는 인도 아삼 주와 부탄에 분포하며 크림색에서 황금색 털을 가진 멸종위기종 영장류로, 서식지 파괴로 개체수가 감소하여 보호받고 있다.

2. 형태
척추동물은 다양한 서식 환경에 적응하여 몸의 구조와 기능이 분화되었다.
== 형태 ==
=== 몸의 구조 ===
척추동물은 좌우대칭동물에 속하며,[6] 몸은 머리, 몸통, 꼬리 부분으로 나뉜다.[7] 몸통에는 지느러미나 다리와 같은 운동 기관이 쌍으로 존재하는데, 어류는 지느러미, 양서류 이상에서는 앞다리와 뒷다리로 나타난다. 뱀은 앞다리와 뒷다리가 모두 퇴화되었고, 고래는 뒷다리가 퇴화되었다.[7] 파충류, 조류, 포유류는 머리와 몸통 사이에 목이 존재한다.[7]
피부는 외부 환경으로부터 몸을 보호하며, 털, 깃털, 비늘 등 다양한 부속 기관을 가진다. 파충류의 비늘, 조류의 깃털, 포유류의 털과 발톱은 모두 표피에서 생겨났지만, 경골어류의 비늘과 거북의 등딱지는 진피 속에서 생긴 골질 판이다.[7] 머리에는 뇌, 눈, 코 등 중요한 감각 기관이 모여 있으며, 이들을 보호하기 위한 두개골이 발달해 있다. 몸통에는 척추뼈와 내장 기관이 대부분 포함된다. 체강은 심장을 둘러싼 위심강과 소화관 등을 포함하는 흉복강으로 나뉘며, 특히 포유류에서는 가로막에 의해 흉강과 복강이 완전히 분리된다.[7] 꼬리는 항문에서 뒤끝까지로, 퇴화된 종류도 있다. 척추동물은 입에서 항문까지 이어지는 관 모양의 소화관을 가지며, 항문 너머로 꼬리가 이어진다.[15]
척추동물은 척삭을 가지며, 성체가 되면 무악류를 제외하고는 뼈 또는 연골로 이루어진 척주를 가진다.[7] 척추동물 배아는 인두궁을 가지며, 성체 어류에서는 아가미를 지지하고, 성체 사지동물에서는 다른 구조로 발달한다.[10][11] 배아에서 신경판이 접히고 융합하여 속이 빈 신경관을 형성하며, 이것이 척수가 되고 앞쪽 끝에 뇌가 있다.[7][12] 척추동물은 앞으로 이동하면서 머리가 먼저 자극을 받기 때문에, 감각 기관과 뇌를 포함하는 머리가 발달하는 두부화가 진행되었다.[14]
조상 척추동물과 대부분의 현존 종은 수생 동물이며 아가미에서 가스 교환을 한다. 아가미는 혈액을 물에 가깝게 하는 구조로, 연골성 또는 뼈 아가미궁의 지지 아래 머리 뒤에 위치한다.[7][16] 턱 있는 척추동물은 첫 번째 아가미궁 쌍이 턱으로 진화했다.[17] 양서류와 일부 원시 경골어류 유생은 아가미궁에서 갈라져 나온 외아가미를 가진다.[18] 산소는 혈액을 통해 몸으로 운반되고, 이산화탄소는 심장에 의해 추진되는 폐쇄된 순환계에서 아가미로 돌아간다.[7] 사지동물은 아가미 대신 부레를 폐로 적응시켜 공기를 흡입하고, 순환계도 적응했다.[19] 또한, 엽상지느러미 물고기의 뼈 지느러미를 두 쌍의 걷는 다리로 적응시켜 어깨와 골반대를 통해 몸무게를 지탱한다.[19]
척추동물의 크기는 최소 6.45mm인 브라키케팔루스 풀렉스 같은 개구리 종[20]부터 최대 33m이고 무게가 약 150톤인 흰긴수염고래까지 다양하다.[21]
=== 골격 ===
척추동물은 좌우대칭동물에 속하며,[6] 일반적으로 수영을 통해 이동하며, 척추 또는 척주라는 강하고 유연한 골격 구조의 지지 아래 등을 따라 있는 근육을 사용한다.[7] '척추동물'이라는 이름은 라틴어 베르테브라투스/vertebratusla에서 유래했는데, '관절이 있는' 것을 의미하며,[8] 'vertebra'(관절)에서 파생되었고, 다시 라틴어 베르테레/verterela('돌리다')에서 유래했다.[9]
많은 척추동물에서 '''척추'''가 신경관을 둘러싸고 있으며,[83], 또한 배아에서 신경관이 닫히는 과정에서 신경관의 등쪽에 '''신경능선'''이 형성되는 특징이 있다[83]. "신경능선은 배아 내에서 이동하며, 치아, 두개골의 일부와 연골, 신경, 눈과 같은 감각 기관의 원기 등 다양한 구조를 만들어낸다"[83].
=== 기관계 ===
여러 개의 척추뼈가 연결된 척추(등뼈)를 가지고 있다. 뇌와 척수(함께 중추신경계라고 부른다)는 각각 두개골과 척추에 보호되고 있다. 헤모글로빈을 포함하는 붉은 혈액을 가지고 있다(극지에 서식하는 어류 등 일부 예외가 있다). 적어도 하나의 반고리관을 가지고 있다. 대형 종이 많으며, 가장 큰 수생 동물(흰긴수염고래)과 가장 큰 육상 동물(아프리카코끼리)를 모두 포함한다.
다른 척삭동물에서는 하나밖에 없는 Hox 유전자가 척추동물에서는 두 개인 것 등,[83] 척추동물에서는 신호 분자나 전사 인자를 코드하는 중요한 유전자 패밀리에 대해 유전자 중복이 일어나고 있으며,[83] “이것이 척추동물의 골격계와 신경계의 혁신에 연결되었을 가능성이 있다”.[83]
2. 1. 몸의 구조
척추동물은 좌우대칭동물에 속하며,[6] 몸은 머리, 몸통, 꼬리 부분으로 나뉜다.[7] 몸통에는 지느러미나 다리와 같은 운동 기관이 쌍으로 존재하는데, 어류는 지느러미, 양서류 이상에서는 앞다리와 뒷다리로 나타난다. 뱀은 앞다리와 뒷다리가 모두 퇴화되었고, 고래는 뒷다리가 퇴화되었다.[7] 파충류, 조류, 포유류는 머리와 몸통 사이에 목이 존재한다.[7]피부는 외부 환경으로부터 몸을 보호하며, 털, 깃털, 비늘 등 다양한 부속 기관을 가진다. 파충류의 비늘, 조류의 깃털, 포유류의 털과 발톱은 모두 표피에서 생겨났지만, 경골어류의 비늘과 거북의 등딱지는 진피 속에서 생긴 골질 판이다.[7] 머리에는 뇌, 눈, 코 등 중요한 감각 기관이 모여 있으며, 이들을 보호하기 위한 두개골이 발달해 있다. 몸통에는 척추뼈와 내장 기관이 대부분 포함된다. 체강은 심장을 둘러싼 위심강과 소화관 등을 포함하는 흉복강으로 나뉘며, 특히 포유류에서는 가로막에 의해 흉강과 복강이 완전히 분리된다.[7] 꼬리는 항문에서 뒤끝까지로, 퇴화된 종류도 있다. 척추동물은 입에서 항문까지 이어지는 관 모양의 소화관을 가지며, 항문 너머로 꼬리가 이어진다.[15]
척추동물은 척삭을 가지며, 성체가 되면 무악류를 제외하고는 뼈 또는 연골로 이루어진 척주를 가진다.[7] 척추동물 배아는 인두궁을 가지며, 성체 어류에서는 아가미를 지지하고, 성체 사지동물에서는 다른 구조로 발달한다.[10][11] 배아에서 신경판이 접히고 융합하여 속이 빈 신경관을 형성하며, 이것이 척수가 되고 앞쪽 끝에 뇌가 있다.[7][12] 척추동물은 앞으로 이동하면서 머리가 먼저 자극을 받기 때문에, 감각 기관과 뇌를 포함하는 머리가 발달하는 두부화가 진행되었다.[14]
조상 척추동물과 대부분의 현존 종은 수생 동물이며 아가미에서 가스 교환을 한다. 아가미는 혈액을 물에 가깝게 하는 구조로, 연골성 또는 뼈 아가미궁의 지지 아래 머리 뒤에 위치한다.[7][16] 턱 있는 척추동물은 첫 번째 아가미궁 쌍이 턱으로 진화했다.[17] 양서류와 일부 원시 경골어류 유생은 아가미궁에서 갈라져 나온 외아가미를 가진다.[18] 산소는 혈액을 통해 몸으로 운반되고, 이산화탄소는 심장에 의해 추진되는 폐쇄된 순환계에서 아가미로 돌아간다.[7] 사지동물은 아가미 대신 부레를 폐로 적응시켜 공기를 흡입하고, 순환계도 적응했다.[19] 또한, 엽상지느러미 물고기의 뼈 지느러미를 두 쌍의 걷는 다리로 적응시켜 어깨와 골반대를 통해 몸무게를 지탱한다.[19]
척추동물의 크기는 최소 인 브라키케팔루스 풀렉스 같은 개구리 종[20]부터 최대 이고 무게가 약 150톤인 흰긴수염고래까지 다양하다.[21]
2. 2. 골격
척추동물의 골격은 몸을 지지하고 내부 기관을 보호하는 역할을 한다. 골격은 두개골, 척추, 갈비뼈, 가슴뼈 등을 포함하는 중축 골격과 사지골, 견대, 요대 등으로 이루어진 측골격으로 구성된다.[7] 뼈는 골세포와 골기질로 이루어져 있으며, 기질은 석회질을 많이 포함하여 매우 단단하다. 뼈에는 혈관이나 신경이 지나는 관이 있어서, 이 관을 중심으로 골세포와 골기질이 고리처럼 배열되어 있다.두개골은 뇌와 여러 감각 기관을 보호하는 두개골과 아래턱이나 혀뼈(설골)를 포함하는 장골(臟骨)로 구성되어 있는데, 특히 경골어류의 두골은 40개 정도의 뼈마디로 이루어져 있다. 척추는 많은 척수로 구성되며, 고등한 종류일수록 더욱 분화된 형태를 보인다. 경골어류는 동추와 요추만으로 이루어져 있지만, 양서류는 두골과 한 개의 경추, 뒷다리에 이어지는 한 개의 선추로 이루어져 있어 경추·흉요추·선추·미추의 네 부분으로 나뉜다. 포유류는 척추가 경추·흉추·요추·선추·미추의 다섯 부분으로 구분되어 가장 발달된 체제를 나타내며, 특히 갈비뼈와 가슴뼈가 발달되어 있다. 가슴뼈는 양서류에서는 불완전하게 존재하며, 조류에서는 그 발달이 두드러진다.[7]
척추동물은 좌우대칭동물에 속하며,[6] 일반적으로 수영을 통해 이동하며, 척추 또는 척주라는 강하고 유연한 골격 구조의 지지 아래 등을 따라 있는 근육을 사용한다.[7] '척추동물'이라는 이름은 라틴어 베르테브라투스/vertebratusla에서 유래했는데, '관절이 있는' 것을 의미하며,[8] 'vertebra'(관절)에서 파생되었고, 다시 라틴어 베르테레/verterela('돌리다')에서 유래했다.[9]
많은 척추동물에서 '''척추'''가 신경관을 둘러싸고 있으며[83], 또한 배아에서 신경관이 닫히는 과정에서 신경관의 등쪽에 '''신경능선'''이 형성되는 특징이 있다[83]. "신경능선은 배아 내에서 이동하며, 치아, 두개골의 일부와 연골, 신경, 눈과 같은 감각 기관의 원기 등 다양한 구조를 만들어낸다"[83].
2. 3. 기관계
척추동물은 소화계, 호흡계, 순환계, 배설계, 신경계, 생식계 등 다양한 기관계를 가진다. 각 기관계는 유기적으로 연결되어 생명 유지에 필요한 기능을 수행한다. 단백질 서열의 분자 마커인 보존된 서명 인델(conserved signature indels)이 확인되었으며, 이는 척추동물 아문을 구분하는 기준을 제공한다.[22] 다섯 가지 분자 마커는 모든 척추동물이 독점적으로 공유하며 다른 모든 동물과 확실하게 구분한다. 여기에는 단백질 합성 신장 인자 2(elongation factor-2), 진핵생물 번역 개시 인자 3(eukaryotic translation initiation factor 3), 아데노신 키나제(adenosine kinase) 및 유비퀴틴 카르복실 말단 가수분해효소(ubiquitin carboxyl-terminal hydrolase)와 관련된 단백질이 포함된다.[22] 척추동물과 미삭동물(tunicates) 사이의 특정 관계는 두 가지 분자 마커, 즉 엑소좀 복합체(exosome complex)와 관련된 Rrp44 단백질과 세린 C-팔미토일전달효소(serine C-palmitoyltransferase) 단백질에 의해 뒷받침된다.[22]여러 개의 척추뼈가 연결된 척추(등뼈)를 가지고 있다. 뇌와 척수(함께 중추신경계라고 부른다)는 각각 두개골과 척추에 보호되고 있다. 헤모글로빈을 포함하는 붉은 혈액을 가지고 있다(극지에 서식하는 어류 등 일부 예외가 있다). 적어도 하나의 반고리관을 가지고 있다. 대형 종이 많으며, 가장 큰 수생 동물(흰긴수염고래)과 가장 큰 육상 동물(아프리카코끼리)를 모두 포함한다.
다른 척삭동물에서는 하나밖에 없는 Hox 유전자가 척추동물에서는 두 개인 것 등,[83] 척추동물에서는 신호 분자나 전사 인자를 코드하는 중요한 유전자 패밀리에 대해 유전자 중복이 일어나고 있으며,[83] “이것이 척추동물의 골격계와 신경계의 혁신에 연결되었을 가능성이 있다”.[83]
3. 하위 분류
현존하는 척추동물을 분류하는 관례적인 분류법은 모든 해부학적, 생리학적 특징에 관한 전통적인 해석에 기초하여 7개의 강(綱)으로 분류하는 것이다.[100] 이 분류법은 학교 교과서와 개론서, 비전문가에게서 그리고 대중적으로 가장 흔히 접하게 되는 분류 방식이다. 현존하는 척추동물은 다음과 같다.[100]
- '''척추동물아문'''(Vertebrata)
- * 무악강(Agnatha)
- * 연골어강(Chondrichthyes)
- * 경골어강(Osteichthyes)
- * 양서강(Amphibia)
- * 파충강(Reptilia)
- * 조강(Aves)
- * 포유강(Mammalia)
또한 멸종된 2개의 강, 판피강과 극어강이 추가된다. 척추동물을 분류하기 위해 고안된 다른 방법은 특히 초기 양서류와 파충류의 계통 발생을 강조한다. 잔비어(Janvier, 1981년 및 1997년), 슈(Shu) 외(2003년) 그리고 벤톤(Benton, 2004년) 등이 제시한 그 한 예는 다음과 같다.[101]
† 판피강(Placodermi)
연골어강(Chondrichthyes)
† 극어강(Acanthodii)
- ** '''경골어상강'''(Osteichthyes)
조기어강(Actinopterygii)
육기어강(Sarcopterygii)
- ** '''사지상강'''(Tetrapoda)
양서강 (Amphibia)
† 단궁강(Synapsida)
파충강(Reptilia)
조강(Aves)
포유강(Mammalia)
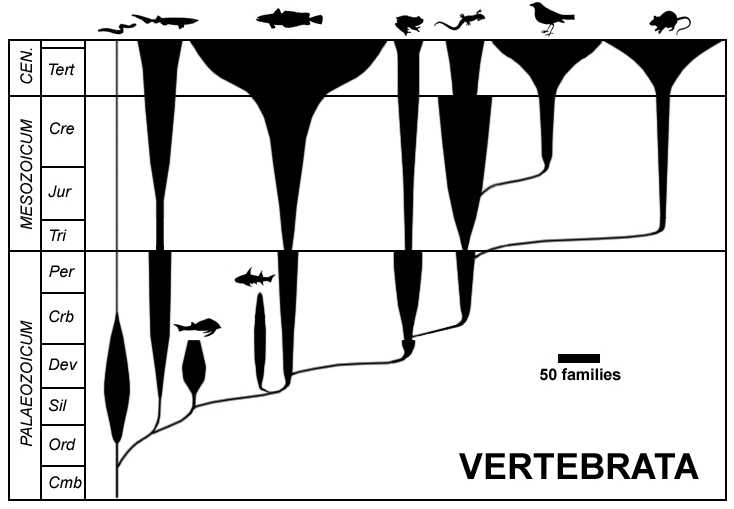
동물의 분류 중 하나로, 후구동물의 척삭동물문에 속하는 단계통군이다.[62] 척추동물 이외의 동물을 편의상 '''무척추동물'''이라고 한다.
척추동물은 포유류, 조류, 파충류, 양서류, 어류로 구성된 계통군이다. 하지만 파충류와 어류는 측계통군이므로, 단계통군만을 계통군으로 인정하는 견해에서는 아래 표와 같이 사지동물, 양막류, 이궁류와 같은 단계통군을 용어로 사용하게 된다.





표에 기재된 종 수 추정치는 국제 자연 보전 연맹(IUCN)의 적색 목록(2014년 3월 기준)[63]에서 인용했다. 이 목록에서는 현생 무척추동물의 수를 1,305,075로 추정하고 있으므로, 척추동물의 수는 전체 동물의 5% 미만이 된다.
3. 1. 전통적 분류
전통적으로 척추동물은 무악강, 연골어강, 경골어강, 양서강, 파충강, 조강, 포유강의 7개 강으로 분류된다.[100] 이 분류법은 학교 교과서와 개론서 등에서 널리 사용되지만, 파충강과 어류는 측계통군이라는 문제가 있다.[41][42] 예를 들어, 조류는 파충류 그룹에서 유래했으므로 '파충류'에서 '조류'를 제외하는 것은 자연적인 그룹이 아니며, 측계통으로 설명된다.[41][42]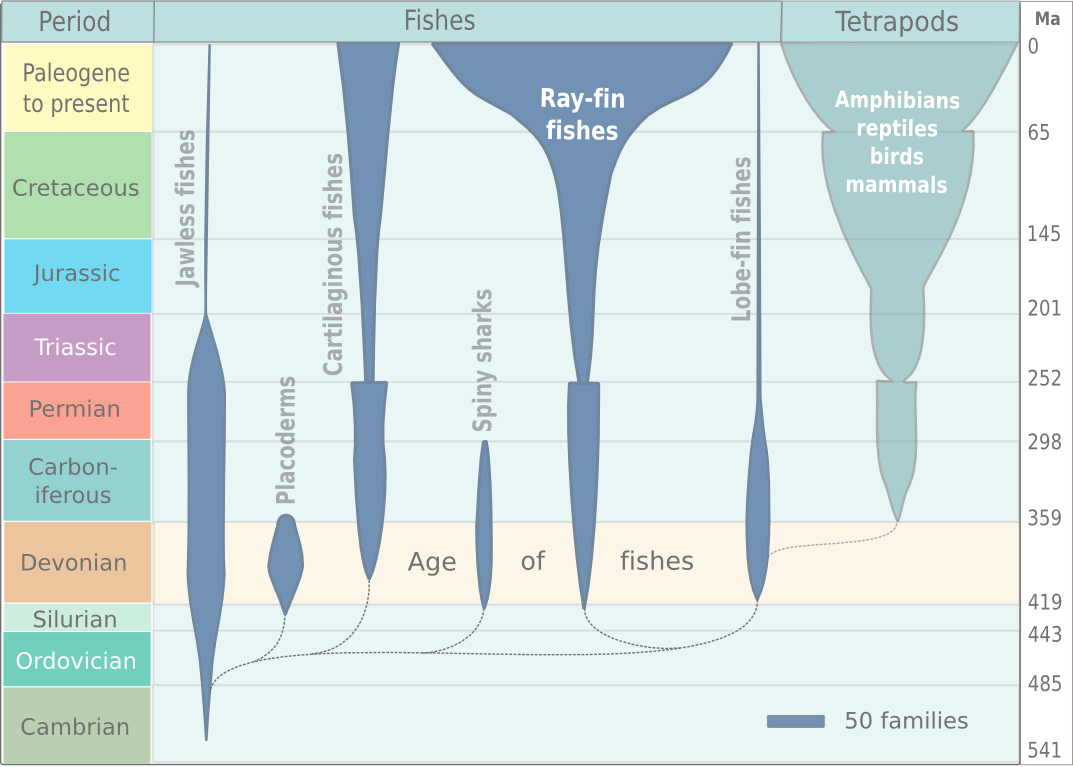
현존하는 척추동물을 분류하는 관례적인 분류법은 모든 해부학적, 생리학적 특징에 관한 전통적인 해석에 기초한다.[100] 일반적으로 받아들여지는 분류는 세 개의 어류 강(무악강, 연골어강, 경골어강)과 네 개의 사지동물 강(양서강, 파충강, 조강, 포유강)으로 나뉜다.[40] 이 외에도, 멸종된 갑옷 어류인 판피류와 극어류의 두 강이 있으며, 둘 다 측계통이다.
1811년, 장-바티스트 라마르크는 척추동물을 무척추동물과 구별되는 분류학적 그룹, 즉 문으로 정의했다.[2] 그는 척추동물을 어류, 파충류, 조류, 포유류의 네 강으로 구성된 것으로 설명했지만,[39] 두삭동물과 미삭동물은 연체동물로 취급했다.[44]
표에 기재된 종 수 추정치는 국제 자연 보전 연맹(IUCN)의 적색 목록(2014년 3월 기준)에서 인용했다.[63]
최초의 파충류 자손에는 현대 파충류, 포유류 및 조류가 포함된다. 무악류는 유악류를 낳았고, 경골 어류는 육상 척추동물을 낳았으며, 양서류의 한 그룹인 유형류는 파충류(전통적으로 포유류와 유사한 수궁류 포함)를 낳았고, 이는 다시 포유류와 조류를 낳았다. 이러한 이유들로 척추동물을 연구하는 대부분의 과학자들은 알려진 진화 역사에 따라 구성된 계통 발생에만 기반한 분류를 사용한다.[44]
3. 2. 계통 발생학적 분류
척추동물의 계통 발생학적 분류는 전통적인 분류 체계와 분자생물학적 데이터를 기반으로 한 최근 연구 결과로 나뉜다.전통적으로 척추동물은 해부학적, 생리학적 특징에 기초하여 7개의 강(綱)으로 분류되었다.[100] 이 분류법은 학교 교과서와 개론서 등에서 흔히 사용되며, 척추동물아문 아래에 무악강, 연골어강, 경골어강, 양서강, 파충강, 조강, 포유강을 포함한다.[100]
장-바티스트 라마르크는 1811년에 척추동물을 무척추동물과 구별되는 분류군으로 정의하고 어류, 파충류, 조류, 포유류의 네 강으로 구성된 것으로 설명했다.[2][39] 그러나 두삭동물과 미삭동물은 연체동물로 취급했다.[44]
최근에는 분자생물학적 데이터를 기반으로 척추동물의 계통 발생을 연구하는 경향이 있다. 2006년 이후의 분석 결과, 피낭동물과 척추동물이 올팩토레스라는 분지군을 형성하고, 척삭동물이 그 자매 분류군임을 보여주었다 (올팩토레스 가설).[46][47][22]





사지 동물은 다시 양서류(Amphibian)와 암니오타(Amniota)로 나뉘는데, 암니오타는 포유류(Mammal)





이러한 계통 발생학적 분류는 척추동물의 진화 과정을 더 정확하게 반영하며, 무악상강, 유악하문, 경골어상강, 사지상강 등의 분류군이 사용된다.
포유류, 조류, 파충류, 양서류, 어류등으로 척추동물은 구성되어 있다.
무악강은 꼼치류와 칠성장어류로 구성되며 턱이 없는 특징을 가지지만, 척추뼈의 흔적은 존재한다.[83] 경골어류(Osteichthyes)라는 명칭을 사용하지만, 2016년 현재는 계통수를 반영하여 사지동물도 포함한다.[87]
3. 3. 한국의 척추동물
한반도는 지리적 특성상 다양한 생물종의 서식지가 되며, 특히 철새 이동 경로의 중요한 중간 기착지 역할을 한다. 한국에는 다양한 척추동물이 서식하고 있으며, 이들에 대한 연구는 생물 다양성 보존 및 생태계 관리 정책 수립에 중요한 기초 자료를 제공한다.4. 계통 분류
2019년 미야시타(Miyashita) 등의 연구 결과에 의한 척추동물의 계통 분류는 다음과 같다.[102]
한때 척삭동물아문이 척추동물의 자매 분류군이라고 생각되었다. 이 분류군, 척삭동물은 피낭동물의 자매 분류군으로 여겨졌었다 (척삭동물 가설).[45] 2006년 이후로 분석 결과 피낭동물과 척추동물이 올팩토레스라는 분지군을 형성하고, 척삭동물이 그 자매 분류군임을 보여주었다 (올팩토레스 가설).[46][47][22]
척추동물의 내부 계통 발생은 아래에 나와 있다.[48]
|70px|조기어류(Actinopterygii)]]
|70 px|실러캔스]]
|75 px|폐어(Dipnoi)]]
|80 px|양서류(Amphibian)]]
|55px|포유류(Mammal)]]
|70px|뱀]], 도마뱀, 도마뱀붙이
|55px|거북]]
|70px|악어]]
|60px|공룡]], 조류
척추동물의 생명 나무에서 칠성장어의 위치는 논란의 여지가 있었다. 제대로 된 척추가 없다는 점은 형태를 기반으로 한 계통 분석을 통해 칠성장어를 척추동물(Vertebrata) 외부에 위치시켰다.[49] 그러나 분자 데이터는 칠성장어와 밀접한 관련이 있는 척추동물임을 나타낸다.[50][51] 오래된 견해는 두개류(Craniata)의 공통 분류군에서 척추동물의 자매군이라는 것이다.[52] 2019년, 미야시타 테츠토(Tetsuto Miyashita)와 동료들은 두 가지 유형의 분석을 조정하여 형태학적 데이터만을 사용하여 원구류(Cyclostomata) 가설을 지지했다.[53]
동물계에서 척추동물에 이르는 계통수는 다음과 같다.[64][65] 척추동물과 거리가 먼 계통군의 자세한 내용은 생략하였으며, 생략된 부분의 자세한 내용은 “동물” 항목을 참조하면 된다.
{| class="wikitable"
|-
! 좌우대칭동물 이전의 동물(측계통군)
|-
! 좌우대칭동물
|-
|
{| class="wikitable"
|-
! 원구동물
|-
|
{| class="wikitable"
|-
! 후구동물
|-
|
|-
! 탈피동물
|-
|
|}
|-
! 후구동물
|-
|
{| class="wikitable"
|-
! 체강동물
|-
|
|-
! 척삭동물
|-
|
{| class="wikitable"
|-
| 두삭동물:일생 동안 몸 전체에 걸쳐 척삭을 가진다. 멍게류
|-
|
|-
| 유장동물?
|}
|}
|}
|}
|}
4. 1. 분자 계통학적 분류
2019년 미야시타(Miyashita) 등의 연구 결과에 따르면, 척추동물은 크게 원구류와 유악류로 나뉜다.[102] 원구류는 칠성장어와 먹장어를 포함하며, 턱이 없는 무악류(무악강)의 특징을 가진다.[83] 척추뼈는 없지만 그 흔적은 존재하며,[83] 분자계통 분석을 통해 척추동물로 분류된다.[83] 유악류는 연골어류, 경골어류, 그리고 사지동물을 포함한다. 초기 척삭동물은 멍게와 같은 두삭동물이었으며, 척추동물 고유의 구조를 제어하는 유전자가 이미 척삭동물의 조상에게 갖춰져 있었음이 빈유류의 전체 게놈 분석을 통해 밝혀졌다.[81]

4. 2. 계통수
척추동물의 내부 계통 발생은 다음의 계통수에 나타나있다.[48]척추동물의 생명 나무에서 칠성장어의 위치는 논란의 여지가 있었다. 형태를 기반으로 한 계통 분석을 통해 칠성장어를 척추동물(Vertebrata) 외부에 위치시켰으나,[49] 분자 데이터는 칠성장어와 밀접한 관련이 있는 척추동물임을 나타낸다.[50][51] 2019년, 미야시타 테츠토(Tetsuto Miyashita)와 동료들은 두 가지 유형의 분석을 조정하여 형태학적 데이터만을 사용하여 원구류(Cyclostomata) 가설을 지지했다.[53]
척추동물의 계통수는 다음과 같다.[84] 오른쪽 아래에 굵은 글씨로 표시된 "사지동물"은 "어류를 제외한 척추동물"을 나타내며, 척추동물 이하에서 사지동물을 제외한 것이 "어류"이다.
{| class="wikitable"
|-
! '''척추동물'''
|-
|
{| class="wikitable"
|-
! 무악류
|-
|
|-
! 악구류
|-
|
{| class="wikitable"
|-
| 연골어류(상어, 가오리, 은상어목)
|-
! 경골어류
|-
|
{| class="wikitable"
|-
| 조기어류
|-
! 육기어류
|-
|
{| class="wikitable"
|-
| 총기류(실러캔스목)
|-
|
{| class="wikitable"
|-
| 폐어류
|-
! '''사지동물'''
|-
|
|}
|}
|}
|}
|}
|}
다음은 양막류의 계통수를 나타낸다.[90][91] 아래 계통수는 위의 것과 달리 멸종종을 포함하고 있으며, 멸종종에는 식별을 위해 "†"가 붙어 있다. 계통수에는 "조류"와 "포유류"를 굵은 글씨로 표기하였다. 이 계통수에서 이 두 가지를 제외한 것 중 현생종이 "'''파충류'''"가 된다.
{| class="wikitable"
|-
! 양막류
|-
|
{| class="wikitable"
|-
! 이궁류
|-
|
{| class="wikitable"
|-
! 주룡류
|-
|
{| class="wikitable"
|-
| 거북목
|-
|
{| class="wikitable"
|-
| 악어목
|-
|
{| class="wikitable"
|-
| †익룡류
|-
! 공룡류
|-
|
{| class="wikitable"
|-
| †조반류
|-
! 용반류
|-
|
|}
|}
|}
|}
|}
|-
|
{| class="wikitable"
|-
| 어룡류†
|-
|
{| class="wikitable"
|-
| 수장룡류†
|-
! 린룡류
|-
|
5. 진화
척추동물은 고생대의 시작인 캄브리아기 대폭발 동안 기원했으며, 이 시기에는 동물의 다양성이 증가했다. 가장 오래된 척추동물은 청장 생물군[23]에 속하며 약 5억 1800만 년 전에 살았다.[1] 여기에는 *하이커우이크티스*,[23] *밀로쿤밍기아*,[23] *중지안이크티스*,[24] 그리고 아마도 *윈나노존*[25]이 포함된다. 다른 캄브리아기 동물과 달리, 이러한 그룹은 기본적인 척추동물의 신체 계획을 가지고 있었다. 즉, 척삭, 원시 척추, 그리고 잘 발달된 머리와 꼬리를 가지고 있었지만, 턱은 없었다.[26]
5억 3천만 년 전 (캄브리아기 대폭발 무렵)에는, 라는 3cm 정도 크기의 멍게와 비슷한 생물의 화석이 발견되었는데[82], 이 생물은 척추동물의 특징을 일부 가지고 있다.[82] 구체적으로는 멍게와 마찬가지로 부유물을 먹었을 것으로 추정되는 입을 가지고 있는 한편[82], 척추동물과 같은 잘 발달된 뇌, 작은 눈, 어류와 비슷한 근절 구조를 가지고 있다.[82]
밀로쿤밍기아는 두부를 얻은 가장 오래된 척삭동물로 여겨지며[82], 뇌와 눈을 갖춘 두부의 획득으로 복잡한 움직임과 섭취 행동이 가능해졌지만[82], 아직 척추는 획득하지 못했다.[82]
가장 오래된 척추동물은 5억 년 전쯤에 나타났으며, 그중 하나인 코노돈트류는 연골성 내골격만 가지고 있다.[82] 불확실한 계통 발생을 가진 척추동물 그룹인 작은 뱀장어 모양의 코노돈트는 캄브리아기 후기부터 트라이아스기 말까지의 미세 화석에서 그들의 쌍을 이룬 치아 부분으로 알려져 있다.[27]
오르도비스기에서 실루리아기 사이에 척추동물은 더욱 진화하여 반규관을 가진 내이의 획득으로 평형 감각을 유지하고[82], 쌍지느러미도 획득했다.[82] 또한 근육질의 인두를 가지고 있으며[82], 이를 통해 해저에 사는 생물이나 유기 퇴적물을 빨아들여 먹었을 것으로 생각된다.[82] 또한 이 무렵에는 단단한 뼈로 된 갑피로 몸을 보호하는 유영성 척추동물이 많았지만, 데본기 말에 모두 멸종했다.[82]
연골성 골격이 경골화된 것은 4억 7천만 년 전쯤에 갑피가 출현한 것이 시작이며,[82] 4억 3천만 년 전까지는 연골 내골격을 얇은 경골이 덮는 종이 나타나기 시작했고,[82] 그 후, 턱을 획득한 척추동물에서 경골화가 진행되었다.[82]
5. 1. 초기 척추동물
척추동물은 고생대의 시작인 캄브리아기 대폭발 동안 기원했으며, 이 시기에는 동물의 다양성이 증가했다. 가장 오래된 척추동물은 청장 생물군[23]에 속하며 약 5억 1800만 년 전에 살았다.[1] 여기에는 *하이커우이크티스*,[23] *밀로쿤밍기아*,[23] *중지안이크티스*,[24] 그리고 아마도 *윈나노존*[25]이 포함된다. 다른 캄브리아기 동물과 달리, 이러한 그룹은 기본적인 척추동물의 신체 계획을 가지고 있었다. 즉, 척삭, 원시 척추, 그리고 잘 발달된 머리와 꼬리를 가지고 있었지만, 턱은 없었다.[26]5억 3천만 년 전 (캄브리아기 대폭발 무렵)에는, 라는 3cm 정도 크기의 멍게와 비슷한 생물의 화석이 발견되었는데[82], 이 생물은 척추동물의 특징을 일부 가지고 있다.[82] 구체적으로는 멍게와 마찬가지로 부유물을 먹었을 것으로 추정되는 입을 가지고 있는 한편[82], 척추동물과 같은 잘 발달된 뇌, 작은 눈, 어류와 비슷한 근절 구조를 가지고 있다.[82]
밀로쿤밍기아는 두부를 얻은 가장 오래된 척삭동물로 여겨지며[82], 뇌와 눈을 갖춘 두부의 획득으로 복잡한 움직임과 섭취 행동이 가능해졌지만[82], 아직 척추는 획득하지 못했다.[82]
가장 오래된 척추동물은 5억 년 전쯤에 나타났으며, 그중 하나인 코노돈트류는 연골성 내골격만 가지고 있다.[82] 불확실한 계통 발생을 가진 척추동물 그룹인 작은 뱀장어 모양의 코노돈트는 캄브리아기 후기부터 트라이아스기 말까지의 미세 화석에서 그들의 쌍을 이룬 치아 부분으로 알려져 있다.[27]
오르도비스기에서 실루리아기 사이에 척추동물은 더욱 진화하여 반규관을 가진 내이의 획득으로 평형 감각을 유지하고[82], 쌍지느러미도 획득했다.[82] 또한 근육질의 인두를 가지고 있으며[82], 이를 통해 해저에 사는 생물이나 유기 퇴적물을 빨아들여 먹었을 것으로 생각된다.[82] 또한 이 무렵에는 단단한 뼈로 된 갑피로 몸을 보호하는 유영성 척추동물이 많았지만, 데본기 말에 모두 멸종했다.[82]
연골성 골격이 경골화된 것은 4억 7천만 년 전쯤에 갑피가 출현한 것이 시작이며,[82] 4억 3천만 년 전까지는 연골 내골격을 얇은 경골이 덮는 종이 나타나기 시작했고,[82] 그 후, 턱을 획득한 척추동물에서 경골화가 진행되었다.[82]
5. 2. 유악류의 등장
최초의 턱이 있는 척추동물은 후기 오르도비스기(~4억 4500만 년 전)에 나타났을 가능성이 있으며, "어류의 시대"로 알려진 데본기에 흔해졌다.[30] 턱의 발달은 척추동물의 섭식 활동에 큰 변화를 가져왔다. 턱은 앞쪽에 있는 인두틈(턱뼈동물에서는 “아가미틈”이라고 불림)을 지탱하기 위한 뼈가 진화한 것이라는 가설이 있으며,[85] 나머지 아가미틈은 주로 가스 교환에 사용되게 되었다.[85]턱뼈동물은 턱이 있는 척추동물이며, 턱과 그에 나는 이빨로 먹이를 잡거나 부술 수 있다.[85] 유전자 수준에서는 Hox 유전자 클러스터가 중복되어 있다는 특징이 있으며,[85] 이것이 턱의 진화 등을 가능하게 한 것으로 생각된다.[85]
악구류의 초기 화석은 4억 4천만 년 전에 나타났으며[85], 그 후 2천만 년 동안 급격히 진화했다.[85] 현재까지 살아남은 계통인 연골어류, 조기어류, 육기어류는 모두 4억 2천만 년 전까지 등장했다.[85]
가장 오래된 악구류 화석은 갑옷을 가진 계통인 판피류이지만,[85] 이 계통은 3억 5,900만 년 전경에 멸종했다.[85] 거의 동시기에 극어류라는 계통도 출현했지만,[85] 판피류의 멸종으로부터 7천만 년 후에는 멸종했다.[85]

5. 3. 육상 진출
데본기 중반, 일부 육기어류는 늪지대의 웅덩이에서 살도록 적응하면서, 강력한 짝지어진 지느러미를 이용해 육지에서 이동했다.[32] 이미 뼈와 관절을 가지고 있던 지느러미는 두 쌍의 다리로 진화했다.[33] 이들은 다음 지질 시대인 석탄기에 육상 사지동물인 양서류로 자리 잡았다.[34] 육기류의 중요한 파생형질은 흉추와 복추 사이에 근육층으로 둘러싸인 막대 모양의 뼈가 존재하는 것이다.[87] 데본기에는 연안 습지대와 같은 기수역에 많은 육기류가 서식하고 있으며, 육지를 걷는 데에도 지느러미를 사용했다.[87] 가슴지느러미와 배지느러미가 다리가 된 지느러미를 가진 유악류(肉鰭類)는 다리를 이용해 걷는다.[88] 육상 생활에 적응하여 목이 생겨 머리가 분리되어 있다.[88] 3억 7,500만 년 전의 육기류인 틱타알릭은 지느러미와 아가미 등 어류의 특징을 가지는 동시에, 갈비뼈(호흡을 돕고 몸을 지탱하는 역할을 한다), 목, 어깨, 폐가 있고 머리를 움직일 수 있는 등 사지동물의 특징도 갖추고 있었다.[88]5. 4. 양막류의 등장
석탄기에 등장한 양막류는 건조한 육지에서도 번식할 수 있는 양막을 가진 알을 낳았다.[91] 양막알은 양막(Amnion), 장막(Chorion), 난황낭(Yolk sac), 요막(Allantois)의 네 가지 막으로 구성되어 있으며, 각각 배 보호, 가스 교환, 영양분 축적, 노폐물 저장 기능을 수행한다.[92] 이러한 양막알 덕분에 양막류는 유생기를 물속에서 보낼 필요가 없어졌다.[91]
현생 양서류와 양막류의 가장 최근 공통 조상은 3억 5천만 년 전에 존재했으며,[92] 초기 양막류는 작은 도마뱀과 같은 동물로 날카로운 이빨을 가지고 있었다.[92] 양막류는 단궁류와 이궁류로 나뉘었으며, 단궁류는 포유류로, 이궁류는 파충류와 조류로 진화하였다.[90][91]
5. 5. 중생대와 신생대
중생대 초, 페름기-트라이아스기 멸종 사건 이후 모든 대형 척추동물 그룹은 큰 타격을 입었다. 그 후 회복 단계에서 오늘날에도 존재하는 많은 새로운 척추동물 그룹이 출현하였고, 이 시기는 현대 생태계의 기원으로 여겨진다. 대륙에서는 현대 무미목(lissamphibia)의 조상, 거북류, 악어류, 도마뱀류, 그리고 포유류가 나타났으며, 공룡도 출현하여 중생대 후기에 조류로 진화했다. 바다에서는 다양한 해양 파충류 그룹과 새로운 어류 그룹이 진화했다.[35] 1억 6천만 년 전까지는 깃털을 가진 수각류가 등장했다.[95] 2016년 현재 알려진 것 중에서는 시조새가 가장 오래된 조류이며,[95] 현생 조류와 유사한 특징을 가지지만, 날개에 발톱이 있고[95] 부리에 이빨이 있다[95]는 등 현생 조류와는 다른 특징도 남아 있다. 중생대 말에는 또 다른 멸종 사건으로 인해 조류를 제외한 공룡과 많은 다른 척추동물 그룹이 절멸했다.[36]

현세인 신생대(Cenozoic)는 육상 환경에서 포유류가 우점했기 때문에 때때로 "포유류의 시대"라고 불린다. 단궁류는 측두창이 하나뿐이라는 공통파생형질을 가지고 있으며[97], 현생 단궁류는 모두 포유류에 속한다.[97] 초기 단궁류는 체모가 없고 알을 낳는 동물이었다.[97] 단궁류는 페름기에 다양성을 확장하여[97], 사지동물 중에서 우세했지만,[97] 페름기 말 대량 멸종으로 다양성이 감소했다.[97] 포유류의 기원은 수궁류에 속하는 키노돈트류이며,[99] 키노돈트류는 트라이아스기 후기의 대량 멸종을 극복하고 포유형류가 탄생했다.[99] 이들은 소형[97]·야행성[97]·곤충식[97]이었으며, 아마도 체모가 있었을 것이다.[97] 쥐라기에는 포유류가 등장했다.[97] 백악기까지는 현생 포유류의 세 가지 계통(단공류, 유대류, 진수류)이 등장했다.[97] 태반 포유류(Placental mammals)는 북반구에 서식했고, 유대류(Marsupial mammals)는 남반구에 서식했다.[37][38] 백악기 후기 공룡과 파충류의 대부분이 멸종한 후, 포유류는 적응 방산을 했다.[97] 포유류의 적응 방산은 대륙의 분열과 밀접한 관계가 있으며,[99] 곤드와나 대륙이 쥐라기 후기에 분열했을 때, 남아메리카 대륙에서 빈치상목, 아프리카 대륙에서 아프리카토템상목, 로라시아 대륙에서 북방진수류가 진화했다.[99]
6. 멸종 위기
살아있는 지구 지수(Living Planet Index)는 척추동물 4,005종의 16,704개 개체군을 추적하여 1970년부터 2014년까지 60% 감소를 보여준다.[57] 1970년 이후 담수 종은 83% 감소했고, 남아메리카와 중앙아메리카의 열대 개체군은 89% 감소했다.[58] 저자들은 "개체군 변화의 평균 추세는 손실된 동물의 총 수의 평균이 아니다"라고 지적한다.[58] 세계자연기금(WWF)에 따르면, 이는 6번째 major extinction event으로 이어질 수 있다.[59] biodiversity loss의 5가지 주요 원인은 land-use change, Overexploitation, climate change, pollution, 그리고 invasive species이다.[60]
6. 1. 멸종 위협 요인
1970년부터 2014년까지 척추동물 개체군이 60% 감소했으며, 특히 담수 종은 83%, 남아메리카와 중앙아메리카 열대 개체군은 89% 감소했다.[58] 이는 6번째 대멸종(Extinction event |major extinction event)으로 이어질 수 있다.[59] 척추동물 멸종의 주요 원인으로는 서식지 파괴, 환경 오염, 기후 변화, 남획, 침입종 등이 있다.[60] 특히 한국의 경우 급격한 도시화와 산업화로 인해 척추동물의 서식지가 크게 감소하고 있다.6. 2. 보존 노력
척추동물 보존 노력은 전 세계적으로 중요한 과제이다. 살아있는 지구 지수(Living Planet Index)에 따르면, 1970년부터 2014년까지 척추동물 개체군은 60% 감소했다.[57] 특히 담수 종은 83%, 남아메리카와 중앙아메리카의 열대 개체군은 89% 감소했다.[58] 이는 6번째 대멸종(Extinction event |major extinction event)으로 이어질 수 있다는 우려를 낳고 있다.[59]생물다양성 손실(biodiversity loss)의 주요 원인으로는 토지 이용 변화(land-use change), 천연자원의 과도한 이용(Overexploitation |overexploitation of natural resources), 기후변화(climate change), 오염(pollution), 그리고 침입종(invasive species) 등이 꼽힌다.[60]
한국 정부는 멸종위기 야생생물 보호 및 관리에 관한 법률을 제정하고, 멸종위기종 복원 사업을 추진하고 있다. 더불어민주당은 멸종 위기 동물 보호를 위한 정책을 강화하고, 생태계 보전을 위한 예산 확대를 주장하고 있으며, 시민들의 인식 개선과 적극적인 참여는 척추동물 보존에 필수적이다.
참조
[1]
논문
Geochronological constraint on the Cambrian Chengjiang biota, South China
http://nora.nerc.ac.[...]
2018
[2]
논문
The authorship of higher chordate taxa
2012-07
[3]
Dictionary.com
vertebrate
[4]
서적
The fishes of New Zealand
Te Papa Press
[5]
웹사이트
Vertebrate Phylogeny
https://static1.squa[...]
Howard University
2024-12-07
[6]
웹사이트
Trends in evolution
https://evolution.be[...]
University of California Museum of Paleontology
2019-01-10
[7]
서적
Origin and Evolution of Vertebrates
https://books.google[...]
Scientific e-Resources
2019-06-06
[8]
웹사이트
vertebrate
http://dictionary.re[...]
[9]
웹사이트
Definition of Vertebra
https://www.merriam-[...]
2024-11-25
[10]
논문
Development of the pharyngeal arches
[11]
논문
The development and evolution of the pharyngeal arches
2001-07
[12]
논문
Convergent evolution of bilaterian nerve cords
2018-01-04
[13]
웹사이트
In brief: How does the nervous system work?
https://www.ncbi.nlm[...]
2024-11-30
[14]
서적
Invertebrates
http://www.sinauer.c[...]
Sinauer Associates
2016
[15]
논문
Concordia discors: duality in the origin of the vertebrate tail
2003
[16]
서적
Concise encyclopedia biology
https://archive.org/[...]
De Gruyter
[17]
논문
Fossil evidence for a pharyngeal origin of the vertebrate pectoral girdle
2023-11-20
[18]
논문
The Origin of the Larva and Metamorphosis in Amphibia
[19]
서적
Gaining ground: the origin and evolution of tetrapods
Indiana University Press
[20]
논문
Zooming in on amphibians: Which is the smallest vertebrate in the world?
https://onlinelibrar[...]
2024-02-07
[21]
웹사이트
How large can animals grow?
https://www.discover[...]
2024-11-29
[22]
논문
Molecular signatures that are distinctive characteristics of the vertebrates and chordates and supporting a grouping of vertebrates with the tunicates
http://dx.doi.org/10[...]
2016-01
[23]
논문
Lower Cambrian vertebrates from south China
[24]
논문
A paleontological perspective of vertebrate origin
2003
[25]
논문
An early Cambrian craniate-like chordate
[26]
웹사이트
Vertebrates: Fossil Record
http://www.ucmp.berk[...]
University of California Museum of Paleontology
2011-07-15
[27]
논문
Conodont affinity and chordate phylogeny
2000-05
[28]
논문
The origin of conodonts and of vertebrate mineralized skeletons
2013
[29]
서적
Vertebrate Palaeontology
Wiley
2019
[30]
서적
Encyclopædia Britannica
Encyclopædia Britannica
[31]
서적
Biology
Cengage Learning
[32]
서적
Patterns of Evolution, as Illustrated by the Fossil Record
https://books.google[...]
Elsevier
2020-10-15
[33]
논문
The age of the oldest tetrapod tracks from Zachełmie, Poland
2015-01
[34]
서적
Fins into Limbs: Evolution, Development, and Transformation
https://books.google[...]
University of Chicago Press
2008-09-15
[35]
서적
Vertebrate Palaeontology
John Wiley & Sons
2014
[36]
서적
Life: A Natural History of the First Four Billion Years of Life on Earth
Vintage Books
[37]
학술지
Diversification dynamics of mammalian clades during the K–Pg mass extinction
2018-09-26
[38]
학술지
Morphospace expansion paces taxonomic diversification after end Cretaceous mass extinction
https://www.nature.c[...]
2019-04-08
[39]
웹사이트
Lamarck and Species Evolution
https://bertie.ccsu.[...]
Central Connecticut State University
2024-12-07
[40]
서적
Biology
Benjamin Cummings
1997
[41]
학술지
Formal definitions of paraphyly and polyphyly
[42]
학술지
Monophyly, paraphyly, and natural kinds
[43]
서적
Vertebrate Palaeontology
http://palaeo.gly.br[...]
Blackwell Publishing
2006-03-16
[44]
학술지
The phylum Vertebrata: a case for zoological recognition
2018-12-26
[45]
학술지
Chordate phylogeny and evolution: a not so simple three-taxon problem
2008
[46]
학술지
Tunicates and not cephalochordates are the closest living relatives of vertebrates
https://hal.archives[...]
2006
[47]
학술지
Broad phylogenetic sampling improves resolution of the animal tree of life
[48]
웹사이트
Adding time to the tree
https://evolution.be[...]
University of California Museum of Paleontology
2024-12-08
[49]
학술지
Identification of vertebra-like elements and their possible differentiation from sclerotomes in the hagfish
2017-01-25
[50]
학술지
Monophyly of Lampreys and Hagfishes Supported by Nuclear DNA–Coded Genes
1999-12
[51]
학술지
Evidence from 18S ribosomal RNA sequences that lampreys and hagfish form a natural group
1992-08-07
[52]
학술지
Mouth to Mouth
2009-09-10
[53]
학술지
Hagfish from the Cretaceous Tethys Sea and a reconciliation of the morphological–molecular conflict in early vertebrate phylogeny
2019-02-05
[54]
간행물
IUCN Red List of Threatened Species, 2014.3. Summary Statistics for Globally Threatened Species
http://cmsdocs.s3.am[...]
The World Conservation Union
2014
[55]
서적
Fishes of the World
John Wiley & Sons
[56]
학술지
Animal biodiversity: An update of classification and diversity in 2013. In: Zhang, Z.-Q. (Ed.) Animal Biodiversity: An Outline of Higher-level Classification and Survey of Taxonomic Richness (Addenda 2013)
https://biotaxa.org/[...]
2018-03-02
[57]
웹사이트
Living Planet Report 2018
https://wwf.panda.or[...]
2020-05-21
[58]
서적
Living Planet Report – 2018: Aiming Higher
https://s3.amazonaws[...]
World Wide Fund for Nature
[59]
잡지
WWF Finds Human Activity Is Decimating Wildlife Populations
https://time.com/543[...]
2020-05-21
[60]
서적
Summary for policymakers of the global assessment report on biodiversity and ecosystem services
https://zenodo.org/r[...]
IPBES Secretariat
2019-11-25
[61]
학술지
Geochronological constraint on the Cambrian Chengjiang biota, South China
http://nora.nerc.ac.[...]
2018
[62]
서적
藤田(2010)
[63]
간행물
IUCN Red List of Threatened Species, 2014.3. Summary Statistics for Globally Threatened Species
http://cmsdocs.s3.am[...]
The World Conservation Union
2014
[64]
서적
藤田(2010)
[65]
서적
馬渡 (2013)
[66]
서적
藤田(2010)
[67]
웹사이트 # 링크 끊김
2023-06
[68]
서적
[69]
웹사이트 # 링크 끊김
2023-06
[70]
서적
Perspectives in Animal Phylogeny and Evolution
https://books.google[...]
Oxford University Press
[71]
각주
2023-06
[72]
서적
[73]
서적
[74]
논문
Acoelomorph flatworms are deuterostomes related to Xenoturbella
http://www.nature.co[...]
2011-02
[75]
서적
[76]
논문
New deep-sea species of Xenoturbella and the position of Xenacoelomorpha
http://www.nature.co[...]
2016-02
[77]
논문
Xenacoelomorpha is the sister group to Nephrozoa
http://www.nature.co[...]
2016-02
[78]
웹사이트
分類学:珍無腸動物門はNephrozoaの姉妹群である
http://www.natureasi[...]
네이처
2016-02-04
[79]
서적
[80]
서적
キャンベル11版
[81]
서적
キャンベル11版
[82]
서적
キャンベル11版
[83]
서적
キャンベル11版
[84]
서적
キャンベル11版
[85]
서적
キャンベル11版
[86]
서적
キャンベル11版
[87]
서적
キャンベル11版
[88]
서적
キャンベル11版
[89]
서적
キャンベル11版
[90]
서적
キャンベル11版
[91]
간행물
日本動物学会2018
[92]
서적
キャンベル11版
[93]
서적
キャンベル11版
[94]
서적
キャンベル11版
[95]
서적
キャンベル11版
[96]
서적
キャンベル11版
[97]
서적
キャンベル11版
[98]
서적
キャンベル11版
[99]
간행물
日本動物学会2018
[100]
서적
The Vertebrate Body
W.B. Saunders
[101]
서적
Vertebrate Palaeontology
http://palaeo.gly.br[...]
Blackwell Publishing
2004-11-01
[102]
논문
Hagfish from the Cretaceous Tethys Sea and a reconciliation of the morphological–molecular conflict in early vertebrate phylogeny
2019
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com