모악동물
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
모악동물은 큐티클로 덮인 투명하거나 반투명한 다트 모양의 동물로, 머리, 몸통, 꼬리로 구분되며, 크기는 1.5mm에서 105mm까지 다양하다. 지느러미와 갈고리 모양의 가시, 이빨을 가지고 있으며, 신경계는 있지만 호흡기, 순환기, 배출기는 없다. 자웅동체로 난자와 정자를 모두 가지며, 전 세계 해양에서 발견되고 플랑크톤을 잡아먹는 매복 포식자이다. 미토콘드리아 유전체와 계통 분류, 화석 기록을 통해 연구되며, 무막근목과 유막근목으로 나뉜다.

더 읽어볼만한 페이지
- 표시 이름과 문서 제목이 같은 위키공용분류 - 라우토카
라우토카는 피지 비치레부섬 서부에 위치한 피지에서 두 번째로 큰 도시이자 서부 지방의 행정 중심지로, 사탕수수 산업이 발달하여 "설탕 도시"로 알려져 있으며, 인도에서 온 계약 노동자들의 거주와 미 해군 기지 건설의 역사를 가지고 있고, 피지 산업 생산의 상당 부분을 담당하는 주요 기관들이 위치해 있다. - 표시 이름과 문서 제목이 같은 위키공용분류 - 코코넛
코코넛은 코코넛 야자나무의 열매로 식용 및 유지로 사용되며, 조리되지 않은 과육은 100g당 354kcal의 열량을 내는 다양한 영양 성분으로 구성되어 있고, 코코넛 파우더의 식이섬유는 대부분 불용성 식이섬유인 셀룰로오스이며, 태국 일부 지역에서는 코코넛 수확에 훈련된 원숭이를 이용하는 동물 학대 문제가 있다. - 한국어 위키백과의 링크가 위키데이터와 같은 위키공용분류 - 라우토카
라우토카는 피지 비치레부섬 서부에 위치한 피지에서 두 번째로 큰 도시이자 서부 지방의 행정 중심지로, 사탕수수 산업이 발달하여 "설탕 도시"로 알려져 있으며, 인도에서 온 계약 노동자들의 거주와 미 해군 기지 건설의 역사를 가지고 있고, 피지 산업 생산의 상당 부분을 담당하는 주요 기관들이 위치해 있다. - 한국어 위키백과의 링크가 위키데이터와 같은 위키공용분류 - 코코넛
코코넛은 코코넛 야자나무의 열매로 식용 및 유지로 사용되며, 조리되지 않은 과육은 100g당 354kcal의 열량을 내는 다양한 영양 성분으로 구성되어 있고, 코코넛 파우더의 식이섬유는 대부분 불용성 식이섬유인 셀룰로오스이며, 태국 일부 지역에서는 코코넛 수확에 훈련된 원숭이를 이용하는 동물 학대 문제가 있다. - 한국어 위키백과의 링크가 위키데이터와 다른 위키공용분류 - 라우토카
라우토카는 피지 비치레부섬 서부에 위치한 피지에서 두 번째로 큰 도시이자 서부 지방의 행정 중심지로, 사탕수수 산업이 발달하여 "설탕 도시"로 알려져 있으며, 인도에서 온 계약 노동자들의 거주와 미 해군 기지 건설의 역사를 가지고 있고, 피지 산업 생산의 상당 부분을 담당하는 주요 기관들이 위치해 있다. - 한국어 위키백과의 링크가 위키데이터와 다른 위키공용분류 - 코코넛
코코넛은 코코넛 야자나무의 열매로 식용 및 유지로 사용되며, 조리되지 않은 과육은 100g당 354kcal의 열량을 내는 다양한 영양 성분으로 구성되어 있고, 코코넛 파우더의 식이섬유는 대부분 불용성 식이섬유인 셀룰로오스이며, 태국 일부 지역에서는 코코넛 수확에 훈련된 원숭이를 이용하는 동물 학대 문제가 있다.

2. 해부학적 특징
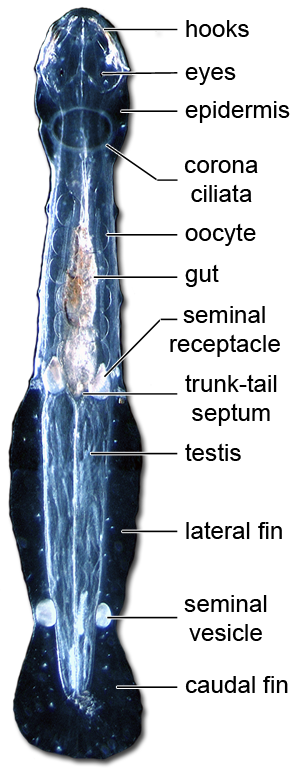
모악동물은 투명하거나 반투명한 다트 모양의 동물이며, 몸은 머리, 몸통, 꼬리의 세 부분으로 나뉜다. 몸 표면은 큐티클로 덮여 있고, 몸의 약 80%는 주요 종단 근육이 차지한다.[39]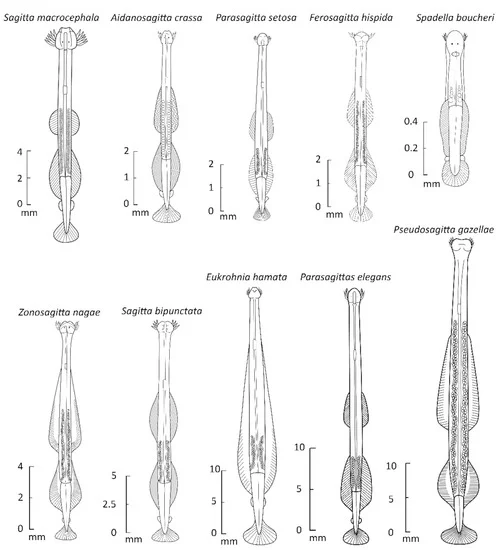
현생종의 체장은 3cm에서 12cm 정도이며, 일부 종은 부분적으로 착색되어 있다. 머리 좌우에는 키틴질 갈고리 모양 가시가 있어 먹이를 잡는 데 사용하며, 입에는 1~2열의 작은 이빨이 있어 먹이를 물어 삼킨다. 공생 세균이 생산한 테트로도톡신을 주입하기도 한다.
눈과 신경계는 있지만, 호흡기, 순환기, 배출기는 없다. 중추 신경계는 머리 등쪽의 뇌, 식도 좌우와 후방 복면의 신경절, 몸통 복면의 신경절로 구성된다.[58] 배출은 피부를 통해 이루어진다. 자웅동체로 난자와 정자를 모두 가지며, 알을 물속에 낳거나 품는 종도 있다. 발광하는 종도 있다.
2. 1. 형태
모악동물은 큐티클로 덮인 투명하거나 반투명한 다트 모양의 동물이다. 크기는 1.5mm에서 남극 종인 ''Pseudosagitta gazellae''의 105mm까지 다양하다.[15] 종 내 개체 간 또는 다른 종 간의 신체 크기는 온도가 낮아짐에 따라 증가하는 것으로 보인다.[15] 몸은 뚜렷한 머리, 몸통, 꼬리로 나뉜다. 몸의 약 80%는 주요 종단 근육이 차지한다.[39]현생종의 체장은 3cm - 12cm 정도이다. 표면은 큐티클로 덮여 있으며, 부분적으로 착색된 종도 있다. 경부 횡격막과 꼬리 횡격막에 의해 몸은 머리, 몸통, 꼬리로 나뉜다. 지느러미 (1쌍 또는 2쌍의 측면 지느러미와 1개의 꼬리 지느러미)가 있으며, 꼬리 지느러미로 전진하고, 측면 지느러미로 정지 및 방향 전환을 한다. 지느러미는 피부가 신장된 것으로 지느러미 줄기에 의해 지지된다는 점은 척추동물과도 유사하지만, 발생하는 위치가 수평 방향이라는 점에서 척추동물과는 결정적으로 다르다. 또한 머리 좌우에는 키틴질의 갈고리 모양의 가시가 있어 먹이를 잡는 데 사용한다. 입에는 소형의 이빨이 1개 또는 2열 있으며, 이것으로 먹이를 물어 삼킨다. 물었을 때, 공생 세균이 생산한 테트로도톡신을 주입하는 것도 있다. 눈과 신경계가 있지만, 호흡기, 순환기, 배출기는 없다. 중추 신경계는 머리 등쪽의 뇌, 식도 좌우와 후방 복면의 신경절, 및 몸통 복면의 신경절로 구성된다.[58] 배출은 피부를 통해 이루어진다. 자웅동체이며, 난자와 정자를 모두 가진다. 알을 물 속에 낳는 것과, 포란하는 것이 있다. 발광하는 종도 있다.
2. 2. 소화계
입은 음식의 통과를 윤활시키는 샘을 포함하는 근육질의 인두로 열린다. 여기에서 곧은 창자가 몸통의 길이를 따라 꼬리 바로 앞의 항문까지 이어진다. 창자는 소화의 주요 부위이며, 앞쪽 끝 근처에 한 쌍의 게실을 포함한다.[16] 물질은 섬모에 의해 몸통 내강으로 이동한다. 폐기물은 단순히 피부와 항문을 통해 배설된다. 유크로니아류 종은 내장과 밀접하게 관련된 오일 액포를 가지고 있는데, 이 기관에는 북극해에서 ''Eukrohnia hamata''의 번식기 외에 번식과 성장을 돕는 왁스 에스테르가 포함되어 있다.[4] 오일 액포가 소화관 중앙에 위치하기 때문에 이 기관은 부력, 트림 및 이동에도 영향을 미칠 수 있다.[5]2. 3. 신경계 및 감각 기관
신경계는 비교적 단순하며 전형적인 원구동물 해부학적 구조를 보인다.[39] 인두를 둘러싼 신경절 신경환으로 구성되어 있다. 뇌는 전방 신경총 영역과 후방 신경총 영역의 두 가지 뚜렷한 기능적 영역으로 구성된다. 전방 신경총 영역은 가시를 움직이는 머리 근육과 소화계를 제어하는 것으로 추정된다. 후방 신경총 영역은 눈과 관상 섬모와 연결되어 있다. 기능이 알려지지 않은 후뇌 기관이라는 추정 감각 구조도 후방 신경총 영역에 있다.[39] 등쪽 신경절이 가장 크지만, 신경은 몸 전체의 모든 신경절에서 뻗어 나온다.모악동물은 각각 여러 개의 색소 컵 단안이 융합된 두 개의 복안을 가지고 있다. 일부 심해 및 동굴생물 종은 색소가 없거나 눈이 없다.[39] 또한 몸 측면을 따라 여러 줄의 감각 강모가 배열되어 있는데, 이는 물고기의 측선과 유사한 기능을 수행하는 것으로 보인다. 머리와 목 위에는 추가로 곡선 모양의 감각 강모 띠가 있다.[16] 거의 모든 모악동물은 광수용체 세포의 방향에 따라 "간접" 또는 "반전" 눈을 가지고 있다. 일부 Eukhroniidae 종만 "직접" 또는 "외전" 눈을 가지고 있다.[39] 모악동물 눈의 독특한 특징은 35–55 nm 폭의 원형 기공 격자를 포함하는 광수용체 막의 층상 구조이다.[39]
섬모 수용체 기관으로 구성된 상당한 기계 감각 시스템은 진동을 감지하여 모악동물이 잠재적 먹이의 헤엄치는 움직임을 감지할 수 있게 한다. 목의 등쪽 부분에 있는 또 다른 기관인 관상 섬모는 화학 수용에 관여하는 것으로 추정된다.[39]
2. 4. 근육 및 운동
털꼬리벌레는 배쪽으로 굽이치는 몸의 움직임을 사용하여 짧게 헤엄치며, 꼬리 지느러미는 추진에, 몸의 지느러미는 안정과 조향에 도움을 준다.[10] 근육의 움직임은 후생동물 중에서 가장 빠르다고 묘사되었다.[39] 근육은 전기 전류 또는 강한 K+ 용액에 의해 직접 흥분할 수 있으며, 주요 신경근 전달 물질은 아세틸콜린이다.[39]2. 5. 기타 기관
체강은 복막으로 덮여 있으며, 진정한 체강을 나타낸다. 체강은 몸통의 각 측면에 하나의 구획과 머리와 꼬리 내부의 추가 구획으로 나뉘며, 이 모든 구획은 격막에 의해 완전히 분리되어 있다. 한두 줄의 작은 이빨, 겹눈, 그리고 신경계를 가지고 있지만, 배설계나 호흡계는 없다.[6][39] 순환계가 없다고 흔히 말하지만, 모악동물은 환형동물과 유사한 초보적인 혈관계를 가지고 있다.[39] 중추 신경계는 머리 등쪽의 뇌, 식도 좌우와 후방 복면의 신경절, 및 몸통 복면의 신경절로 구성된다.[58] 배출은 피부를 통해 이루어진다.3. 생식과 생활사
모악동물은 자웅동체로 난자와 정자를 모두 가지고 있으며, 짝짓기를 통해 수정을 한다.[3][16] 수정된 알은 대부분 물에 떠다니며, 1~3일 후에 부화한다.[39][11] 모악동물은 변태를 하지 않고, 잘 정의된 유충 단계가 없는 것이 특징이다.[16][39][24] 수명은 짧은 편이며, ''Sagitta friderici''의 경우 최대 15개월까지 기록되었다.[12]
3. 1. 생식
모든 종은 자웅동체로, 난자와 정자를 모두 가지고 있다.[3] 각 동물은 꼬리 내부에 한 쌍의 고환과 몸통 후방 부위에 한 쌍의 난소를 가지고 있다. 미성숙한 정자는 고환에서 방출되어 꼬리 내부에서 성숙하며, 짧은 관을 통해 정낭으로 이동하여 정자낭으로 포장된다.[16]짝짓기 동안, 각 개체는 정낭이 파열된 후 파트너의 목에 정자낭을 놓는다. 정자는 정자낭에서 빠르게 탈출하여 동물의 정중선을 따라 헤엄쳐 꼬리 바로 앞에 있는 작은 구멍 한 쌍에 도달한다. 이 구멍은 난관과 연결되며, 이곳으로 이미 발달된 난자가 난소에서 통과했으며, 수정이 일어나는 곳이다.[16] 정낭과 난관은 정자를 축적하고 저장하여 여러 번의 수정 주기를 수행한다.[39] Spadellidae의 일부 저서생물은 교미 전에 정교한 구애 행동을 하는 것으로 알려져 있으며,[39] 예를 들어 ''Paraspadella gotoi''가 있다.[24]
난자는 대부분 부유성이며, 기질에 난자를 부착하는 ''Ferosagitta hispida''와 같은 몇몇 종을 제외하고는 그렇다.[39] ''Eukrohnia''에서는 난자가 육아낭에서 발달하거나 조류에 부착된다.[11] 난자는 보통 1~3일 후에 부화한다. 모악동물은 변태를 거치지 않으며, 잘 정의된 유충 단계를 가지지 않는다.[16][39] 이는 해양 무척추동물에서 드문 특징이다.[24] 그러나 비율, 키틴질 구조 및 지느러미 발달과 관련하여 신생 동물과 성체 간에는 상당한 형태적 차이가 있다.[39][12]
3. 2. 생활사
모든 종은 자웅동체로, 난자와 정자를 모두 가지고 있다.[3] 각 동물은 꼬리 내부에 한 쌍의 고환과 몸통 후방 부위에 한 쌍의 난소를 가지고 있다. 미성숙한 정자는 고환에서 방출되어 꼬리 내부에서 성숙하며, 짧은 관을 통해 정낭으로 이동하여 정자낭으로 포장된다.[16]짝짓기 동안, 각 개체는 정낭이 파열된 후 파트너의 목에 정자낭을 놓는다. 정자는 정자낭에서 빠르게 탈출하여 동물의 정중선을 따라 헤엄쳐 꼬리 바로 앞에 있는 작은 구멍 한 쌍에 도달한다. 이 구멍은 난관과 연결되며, 이곳으로 이미 발달된 난자가 난소에서 통과했으며, 수정이 일어나는 곳이다.[16] 정낭과 난관은 정자를 축적하고 저장하여 여러 번의 수정 주기를 수행한다.[39] Spadellidae의 일부 저서생물은 교미 전에 정교한 구애 행동을 하는 것으로 알려져 있으며,[39] 예를 들어 ''Paraspadella gotoi''가 있다.[24]
난자는 대부분 부유성이며, 기질에 난자를 부착하는 ''Ferosagitta hispida''와 같은 몇몇 종을 제외하고는 그렇다.[39] ''Eukrohnia''에서는 난자가 육아낭에서 발달하거나 조류에 부착된다.[11] 난자는 보통 1~3일 후에 부화한다. 모악동물은 변태를 거치지 않으며, 잘 정의된 유충 단계를 가지지 않는다.[16][39] 이는 해양 무척추동물에서 드문 특징이다.[24] 그러나 비율, 키틴질 구조 및 지느러미 발달과 관련하여 신생 동물과 성체 간에는 상당한 형태적 차이가 있다.[39][12]
모악동물의 수명은 가변적이지만 짧다. 가장 긴 기록은 ''Sagitta friderici''에서 15개월이었다.[12]
4. 생태
모악동물은 주로 요각류와 가지뿔새우를 먹는 매복 포식자이지만, 단각류, 크릴, 물고기 유생도 먹는다.[14] 같은 종의 어린 개체를 먹기도 하며,[17] 일부는 조류와 부스러기를 먹는 잡식성이다.[18] 먹이를 제압하기 위해 신경독인 테트로도톡신을 사용하는데,[19] 이는 ''비브리오'' 박테리아 종에 의해 합성되는 것으로 추정된다.[39]
4. 1. 서식 환경
모악동물은 극지방에서 열대 지방에 이르기까지 전 세계의 모든 대양에서 발견되며, 기수 및 강기슭에서도 발견된다. 이들은 열수 분출구에서 심해 해저, 해초류 서식지 및 해양 동굴에 이르기까지 매우 다양한 환경에 서식한다.[39] 대부분 플랑크톤이며, 종종 동물성 플랑크톤의 두 번째로 흔한 구성 요소로, 요각류 생물량의 10~30% 범위를 차지한다.[39] 캐나다 분지에서 모악동물은 동물성 플랑크톤 생물량의 약 13%를 차지한다.[13] 따라서 이들은 생태학적으로 중요하며 고등어 또는 정어리와 같이 상업적으로 관련된 물고기를 포함하여 물고기와 다른 포식자의 주요 식량원이다.[14] 알려진 종의 58%는 원양생물이며,[15] 종의 약 3분의 1은 저서생물 또는 미세저서생물이거나 기질 바로 근처에 서식한다.[39] 모악동물은 최대 5000m, 어쩌면 6000m 깊이까지 기록되었다.[15]모악동물의 가장 높은 밀도는 얕은 물의 유광층에서 관찰된다.[39] 더 큰 모악동물 종은 물속 더 깊이 사는 경향이 있지만 유생 단계는 물기둥에서 더 높이 보낸다.[24] 그러나 화살벌레는 일주기 수직 이동을 하여 포식자를 피하기 위해 낮에는 더 낮은 깊이에서 보내고 밤에는 표면에 가까이 온다. 물기둥에서의 위치는 빛, 온도, 염분, 나이 및 식량 공급에 따라 달라질 수 있다. 이들은 해류에 거슬러 헤엄칠 수 없으며, 해류와 수괴의 수문학적 지표로 사용된다.[39]
4. 2. 행동
모악동물은 배양이 복잡하고 자연 서식지를 재현하기 어렵기 때문에 행동과 생리에 대해 알려진 바가 거의 없다.[39] 온도가 높을수록 더 자주 먹이를 섭취하는 것으로 알려져 있다. 플랑크톤성 모악동물은 종종 "뛰어오르기 및 가라앉기" 행동으로 지속적으로 헤엄쳐서 수층에서 원하는 위치에 머물러야 하며, 먹이를 잡기 위해 적극적으로 헤엄친다. 이들은 머리를 아래로 향하게 하여 몸을 약간 기울이는 경향이 있다.[39] 종종 "활강" 행동을 보이며, 잠시 천천히 가라앉다가 지느러미의 빠른 움직임으로 따라잡는다.[24] 저서성 모악동물은 일반적으로 바위, 해조류 또는 해초와 같은 기질에 부착되어 있으며, 모래 알갱이 위나 사이에 드물게 위치하며, 먹이가 지나갈 때까지 가만히 있는 매복 포식자 역할을 한다.[39] 먹이는 섬모 울타리와 털 기관을 통해 진동을 감지하여 감지된다.[39] 예를 들어, ''Spadella cephaloptera'' 개체는 적절한 주파수로 진동하는 유리 또는 금속 프로브를 공격한다.[24] 먹이를 잡기 위해 모악동물은 꼬리 지느러미의 강한 움직임으로 앞으로 점프한다.[39] 먹이와 접촉하면, 움켜쥐는 가시 위로 후드를 당겨서 먹이 주변에 가둔 후 입으로 가져간다. 이들은 먹이를 통째로 삼킨다.[24]4. 3. 먹이와 포식자
모든 모악동물은 다른 플랑크톤 동물, 주로 요각류와 가지뿔새우를 잡아먹는 매복 포식자이지만,[16][39] 단각류, 크릴, 물고기 유생도 잡아먹는다.[14] 성체는 같은 종의 어린 개체를 먹을 수 있다.[17] 일부 종은 조류와 부스러기를 먹는 잡식성 동물로도 보고된다.[18] 모악동물은 먹이를 제압하기 위해 신경독 테트로도톡신을 사용하는 것으로 알려져 있으며,[19] 아마도 ''비브리오'' 박테리아 종에 의해 합성될 것이다.[39]4. 4. 생태학적 중요성
모악동물은 플랑크톤이며, 종종 동물성 플랑크톤에서 요각류 다음으로 흔한 구성 요소로, 요각류 생물량의 10~30% 범위를 차지한다.[39] 캐나다 분지에서 모악동물은 동물성 플랑크톤 생물량의 약 13%를 차지한다.[13] 따라서 이들은 생태학적으로 중요하며 고등어, 정어리와 같이 상업적으로 관련된 물고기를 포함하여 물고기와 다른 포식자의 주요 먹이가 된다.[14]모악동물은 해류에 거슬러 헤엄칠 수 없으므로, 해류와 수괴의 수문학적 지표로 사용된다.[39]
5. 유전학
모악동물은 같은 종 안에서도 개체마다 미토콘드리아 유전체에 차이를 보인다.[22]
5. 1. 미토콘드리아 유전체
화살벌레( ''Spadella cephaloptera'')의 mtDNA는 2004년에 염기 서열이 밝혀졌으며, 당시 알려진 가장 작은 후생동물 미토콘드리아 유전체로 11,905 염기쌍(bp) 길이를 가졌다.[53] (현재는 빗해파리 ''Mnemiopsis leidyi''의 미토콘드리아 유전체(10,326 bp)에 의해 기록이 깨졌다).[20] 모든 미토콘드리아 tRNA 유전자는 존재하지 않으며, MT-ATP8 및 MT-ATP6 유전자도 없다.[53] 2004년에 염기 서열이 밝혀진 ''Paraspadella gotoi''의 mtDNA는 11,403 bp로 더 작고 유사한 패턴을 보인다. 일반적으로 존재하는 22개의 tRNA 유전자 중 21개가 없고, 37개의 유전자 중 14개만 존재한다.[21]모악동물은 동일 종 내 개체 간에 독특한 미토콘드리아 유전체 다양성을 보인다.[22]
6. 계통 분류
모악동물은 전통적으로 배아 발생 특징에 근거하여 후구동물로 분류되었으나, 분자계통학적 연구와 기타 형질들은 선구동물에 속한다는 것을 보여준다.[65][66][67][68][69][70][71] 발생학적으로는 선형동물과 유사하지만, 분자계통학 및 고생물학적 증거는 악구동물, 미악동물, 윤형동물을 포함하는 담악동물과의 연관성을 시사한다.[59] 이에 따라 모악동물은 담악동물 내에 분류되거나,[59] 모악동물과 기존 담악동물을 합쳐 새로운 계통군으로 칭하기도 한다.[60]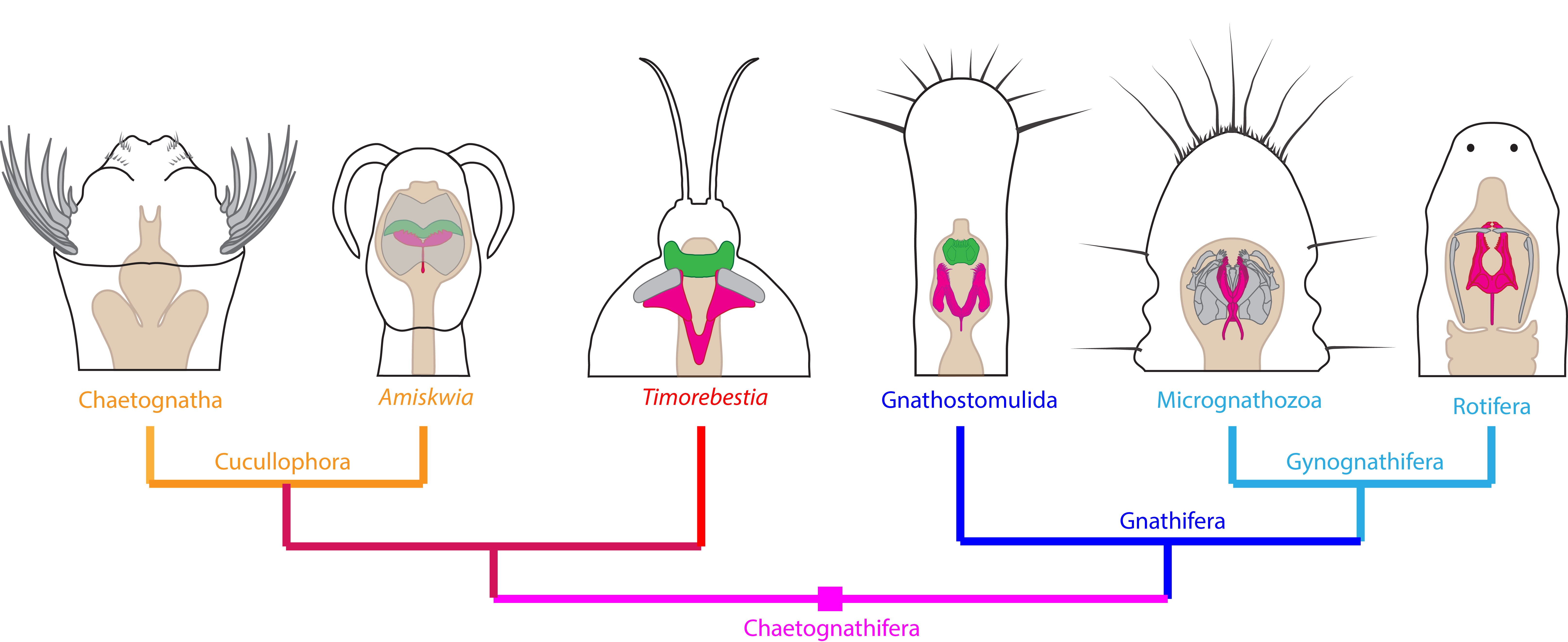
좌우대칭동물 내에서 모악동물의 계통발생학적 위치는 다음과 같다.
{{분기도
| 캡션 = 좌우대칭동물에서의 모악동물의 계통 위치.
| 분기도 =
{{분기
| 스타일 = width:30em;font-size:100%;line-height:100%
| 라벨1 = | 1 = {{분기
| 라벨1 = 후구동물 | 1 = 극피동물・척삭동물 등
| 라벨2 = 선구동물 | 2 = {{분기
| 라벨1 = 탈피동물 | 1 = 절지동물・선형동물 등
| 라벨2 = 나선동물 | 2 = {{분기
| 라벨1 = 협의의 환형동물 | 1 = 연체동물・환형동물 등
| 라벨2 = 모악동물 | 2 = {{분기
| 라벨1 = 담악동물 | 1 = 악구동물・미악동물・윤형동물
| 2 = '''모악동물'''
}} }} }} }} }}
6. 1. 내부 계통
현생 모악동물의 분류 체계는 다음과 같다.
다음은 2021년 현재 형태학적 및 분자 데이터를 모두 기반으로 한 화살벌레의 진화 계통수이다.[39]
- Sagittoideala 현생 화살벌레강
- * Biphragmophorala 양막근목
- * Monophragmophorala 단막근목
- * Aphragmophorala 무막근목
- †Archisagittoideala 고(古)화살벌레강
- †Protoconodontala 프로토코노돈트[61]
6. 2. 화석 기록
모악동물은 부드러운 몸체 때문에 화석으로 잘 보존되지 않지만, 몇몇 화석 종들이 보고되었다.[1] 모악동물은 캄브리아기에 처음 나타났다. 중국 윈난성의 하부 캄브리아기 마오톈산 셰일에서 발견된 ''Eognathacantha ercainella''[40]와 ''Protosagitta spinosa''[41], 브리티시컬럼비아의 중부 캄브리아기 버제스 셰일에서 발견된 ''Capinatator praetermissus'' 등이 완전한 몸체 화석으로 보고되었다. 2024년에 처음 보고된 캄브리아기 줄기군 모악동물인 ''Timorebestia''는 현생 종보다 훨씬 컸으며, 이는 모악동물이 오늘날과 비교하여 해양 생태계에서 다른 역할을 수행했음을 보여준다.[42]모악동물은 한때 코노돈트와 관련이 있을 것으로 생각되었으나, 코노돈트는 척추동물의 치아 요소로 밝혀졌다. 현재는 원시코노돈트 요소(예: ''Protohertzina anabarica'')가 코노돈트의 이빨이 아니라 모악동물의 붙잡는 가시일 가능성이 높다고 여겨진다. 이전에는 초기 캄브리아기의 모악동물이 원시코노돈트 요소로만 추정되었지만, 최근 몸체 화석의 발견으로 당시의 존재가 확인되었다.[43] 모악동물이 이미 초기 캄브리아기부터 해양 먹이 그물의 중요한 구성 요소였다는 증거도 있다.[44]
화석은 적지만, 캄브리아기에 발생한 것으로 여겨진다. 드물게 구기(口器)의 이빨이 고생대 후기 이후의 화석으로 발견된다. 완전한 몸의 화석은, 중국 윈난성의 청장 동물군과 그린란드의 시리우스 파셋 동물군 (둘 다 캄브리아기 제3기, 약 5억 1800만 년 전)에서 보고된 것이 가장 오래된 것으로 여겨진다.
과거 코노돈트 (나중에 척추동물로 여겨짐)의 기원으로 생각되었으며, 미소 경골격 화석군의 가시만 알려진 프로토코노돈트 (Protoconodont)는, 실제로는 코노돈트가 아닌 털벌레동물로 재검토되었다.[61] 이에 따르면, 털벌레동물의 화석 기록은 더 오래된 시기 (포르투니안기, 약 5억 3800만 ~ 약 5억 3500만 년 전)까지 거슬러 올라간다.[63]
같은 캄브리아기의 아미스퀴아와 티모레베스티아는 털벌레동물과 악구동물 (특히 턱구동물)의 중간적인 성질을 가지며, 기저 (스템 그룹) 털벌레동물로 여겨진다.[62][60][63] 특히 티모레베스티아는 현생종을 크게 웃도는 20 cm의 대형이며, 당시의 털벌레동물은 현생종과 같은 부유성의 2차 소비자뿐만 아니라, 유영성이며 최상위 포식자의 틈새 시장도 차지했음이 시사된다.[63]
7. 하위 분류
참조
[1]
논문
Early Cambrian origin of modern food webs: evidence from predator arrow worms
2007-03
[2]
간행물
2012-02-08
[3]
서적
The Biology of Chaetognaths
Oxford University Press
[4]
논문
Growth and reproduction of the chaetognaths Eukrohnia hamata and Parasagitta elegans in the Canadian Arctic Ocean: capital breeding versus income breeding
https://academic.oup[...]
2017-11-01
[5]
논문
The physical properties of lipids and their role in controlling the distribution of zooplankton in the oceans
https://academic.oup[...]
2012-06-01
[6]
서적
Kingdoms and Domains: An Illustrated Guide to the Phyla of Life on Earth
https://books.google[...]
[7]
문서
Photoreception
Encyclopædia Britannica
2009
[8]
논문
A bioluminescent chaetognath
1994-01-20
[9]
논문
Bioluminescent organs of two deep-sea arrow worms, Eukrohnia fowleri and Caecosagitta macrocephala, with further observations on Bioluminescence in chaetognaths
2010-10
[10]
논문
A model of rapid-start swimming at intermediate Reynolds number: undulatory locomotion in the chaetognath ''Sagitta elegans''
http://jeb.biologist[...]
[11]
논문
Reproduction of meso- and bathypelagic chaetognaths in the genus Eukrohnia
1982-11-01
[12]
논문
Sagitta friderici Ritter-Záhony (Chaetognatha) from South Atlantic waters: Abundance, population structure, and life cycle
2004
[13]
논문
Diversity and vertical distribution of mesozooplankton in the Arctic's Canada Basin
2010
[14]
논문
Taxonomic reassessment of chaetognaths (Chaetognatha, Sagittoidea, Aphragmophora) from Korean waters
2022
[15]
논문
The Sizes, Growth and Reproduction of Arrow Worms (Chaetognatha) in Light of the Gill-Oxygen Limitation Theory (GOLT)
2021
[16]
서적
Invertebrate Zoology
Holt-Saunders International
[17]
논문
In situ feeding behavior of Sagitta elegans and Eukrohnia hamata (Chaetognatha) in relation to the vertical distribution and abundance of prey at Ocean Station "P"1
1980
[18]
논문
Non-carnivorous feeding in Arctic chaetognaths
http://www.sciencedi[...]
2020-07-01
[19]
서적
The Biology of chaetognaths
Oxford University Press
1991
[20]
논문
Mitochondrial Genomic Landscape: A Portrait of the Mitochondrial Genome 40 Years after the First Complete Sequence
2021
[21]
논문
The mitochondrial genome of ''Paraspadella gotoi'' is highly reduced and reveals that chaetognaths are a sister group to protostomes
2004
[22]
논문
Extreme Mitogenomic Variation in Natural Populations of Chaetognaths
2017
[23]
논문
Careful amendment of morphological data sets improves phylogenetic frameworks: re-evaluating placement of the fossil ''Amiskwia sagittiformis''
https://hal.sorbonne[...]
2022
[24]
논문
Phylogeny: The Continuing Classificatory Conundrum of Chaetognaths
2006
[25]
웹사이트
Systema Naturae 2000 Taxon: Phylum Chaetognatha per Margulis and Schwartz
http://sn2000.taxono[...]
2006-11-25
[26]
논문
A new spiralian phylogeny places the enigmatic arrow worms among gnathiferans
https://discovery.uc[...]
[27]
논문
Bilateral jaw elements in Amiskwia sagittiformis bridge the morphological gap between gnathiferans and chaetognaths
https://research-inf[...]
[28]
논문
Rotiferan Hox genes give new insights into the evolution of metazoan bodyplans
[29]
논문
Spiralian phylogeny informs the evolution of microscopic lineages
[30]
논문
Phylogeny of Syndermata (syn. Rotifera): Mitochondrial gene order verifies epizoic Seisonidea as sister to endoparasitic Acanthocephala within monophyletic Hemirotifera
[31]
웹사이트
Systema Naturae 2000 Taxon: Phylum Chaetognatha per Cavalier-Smith
http://sn2000.taxono[...]
2006-11-25
[32]
논문
Broad taxon and gene sampling indicate that chaetognaths are protostomes
2006-08
[33]
논문
Chaetognath phylogenomics: a protostome with deuterostome-like development
2006-08
[34]
논문
Identification of chaetognaths as protostomes is supported by the analysis of their mitochondrial genome
2004-11
[35]
논문
Immunohistochemical analysis and 3D reconstruction of the cephalic nervous system in Chaetognatha: Insights into the evolution of an early bilaterian brain?
2010-02
[36]
논문
A new look at the ventral nerve centre of Sagitta: implications for the phylogenetic position of Chaetognatha (arrow worms) and the evolution of the bilaterian nervous system
2007-05
[37]
논문
Rotiferan Hox genes give new insights into the evolution of metazoan bodyplans
2017-04
[38]
논문
A New Spiralian Phylogeny Places the Enigmatic Arrow Worms among Gnathiferans
https://discovery.uc[...]
2019-01
[39]
서적
Invertebrate Zoology: A Tree of Life Approach
CRC Press
2023-08-14
[40]
논문
A possible Lower Cambrian chaetognath (arrow worm)
2002-10
[41]
논문
Taphonomy and palaeoecology of the Early Cambrian Chengjiang Biota from Eastern Yunnan, China
[42]
논문
A giant stem-group chaetognath
2024
[43]
논문
New evidence for the protoconodont origin of chaetognaths
http://www.app.pan.p[...]
[44]
논문
Early Cambrian origin of modern food webs: Evidence from predator arrow worms
2007
[45]
서적
Natuurkundige Verlustigingen, Behelzende Microscopise Waarneemingen Van de in—En Uitlandse Water—En Land-Dieren
https://books.google[...]
J. Bosch
1778
[46]
간행물
"Observations Zoologiques Faites à Bord de l’Astrolabe, en Mai 1826, dans le Détroit de Gibraltar (suite et fin). Description des genres Biphore, Carinaire, Hyale, Flèche, Cléodore, Anatife et Briarée."
1827
[47]
간행물
"Observations on the Structure and Propagation of the Genus Sagitta."
1844
[48]
논문
XXXI.— Anatomical and physiological observations on Sagitta bipunctata
https://www.biodiver[...]
1845
[49]
서적
Nachträge und Berichtigungen zu dem ersten Bande von J. van Der Hoeven’s Handbuch der Zoologie. Eine Systematisch Geordnete Übersicht der Hauptsächlichste Neueren Leistungen:über die Zoologie der Wirbellosen Thiere
L. Voss
1856
[50]
간행물
"Revision der Chaetognathan."
1911
[51]
논문
The Taxonomical Outline of Chaetognatha
1965
[52]
간행물
"Systematics of the Chaetognatha."
1991
[53]
논문
Identification of Chaetognaths as Protostomes is Supported by the Analysis of Their Mitochondrial Genome
2004
[54]
논문
Ultrastructure of Meelsvirus: A nuclear virus of arrow worms (phylum Chaetognatha) producing giant "tailed" virions
2018-09-19
[55]
논문
Prey detection by Chaetognatha via a vibration sense
1967-11-14
[56]
논문
Ultrastructural study and ontogenesis of the appendages and related musculature of Paraspadella (Chaetognatha)
2003-10
[57]
간행물
Serendipitous Discovery in a Marine Invertebrate (Phylum Chaetognatha) of the Longest Giant Viruses Reported till Date
https://www.semantic[...]
[58]
논문
Development of the nervous system in hatchlings of Spadella cephaloptera (Chaetognatha), and implications for nervous system evolution in Bilateria: Nervous system development in S. cephaloptera
https://onlinelibrar[...]
2011-06
[59]
논문
A New Spiralian Phylogeny Places the Enigmatic Arrow Worms among Gnathiferans
https://doi.org/10.1[...]
2019-01
[60]
논문
Careful amendment of morphological data sets improves phylogenetic frameworks: re-evaluating placement of the fossil Amiskwia sagittiformis
https://www.tandfonl[...]
2022-12-31
[61]
논문
Zooproblematica and mollusca from the Lower Cambrian Meishucun section (Yunnan, China) and taxonomy and systematics of the Cambrian small shelly fossils of China
https://www.research[...]
2010-12-01
[62]
논문
Bilateral Jaw Elements in Amiskwia sagittiformis Bridge the Morphological Gap between Gnathiferans and Chaetognaths
https://doi.org/10.1[...]
2019-03
[63]
논문
A giant stem-group chaetognath
https://www.science.[...]
2024-01-05
[64]
ITIS
2012-02-08
[65]
저널 인용
Higher-level metazoan relationships: recent progress and remaining questions
2011-06
[66]
저널 인용
Rotiferan Hox genes give new insights into the evolution of metazoan bodyplans
http://www.nature.co[...]
2017-04-04
[67]
저널 인용
Hallucigenia’s onychophoran-like claws and the case for Tactopoda
https://doi.org/10.1[...]
[68]
웹인용
Palaeos Metazoa: Ecdysozoa
http://palaeos.com/m[...]
2017-09-02
[69]
저널 인용
Phylogenetic position of Loricifera inferred from nearly complete 18S and 28S rRNA gene sequences
https://doi.org/10.1[...]
2015-06
[70]
서적 인용
Animal Evolution: Interrelationships of the Living Phyla
Oxford University Press
[71]
웹인용
Bilateria
http://tolweb.org/Bi[...]
2001-01-01
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com