리보스 5-인산
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
리보스 5-인산은 5탄당인 리보스의 5번 탄소에 인산기가 결합된 구조를 가지는 당의 일종이다. 세포 생장, NADPH, 리보스 5-인산, ATP의 필요에 따라 오탄당 인산 경로 및 해당과정을 통해 생합성된다. 리보스 5-인산은 DNA, RNA, ATP, 조효소 A 등 다양한 생체 분자의 전구체로 작용하며, 뉴클레오타이드 및 히스티딘 생합성에 관여한다. 리보스 5-인산의 불균형은 암, 리보스 5-인산 이성질화효소 결핍증, 통풍, 레쉬-니한 증후군 등 여러 질병과 관련이 있다.
더 읽어볼만한 페이지
- 단당류 유도체 - 1-디옥시-D-자일룰로스 5-인산
1-디옥시-D-자일룰로스 5-인산은 비메발론산 경로의 중간체로서 피루브산과 D-글리세르알데히드 3-인산으로부터 생성되어 박테리아, 식물, 기생충 등 다양한 생물에서 아이소프레노이드, 티아민, 피리독솔을 만드는 데 사용된다. - 단당류 유도체 - 뉴라민산
뉴라민산은 자연적으로 존재하지 않지만, 그 유도체인 시알산은 동물 조직, 박테리아, 당단백질, 산성 스핑고당지질 등에 널리 분포하며, N-아세틸뉴라민산이 대표적이다. - 유기 인산염 - 인산다이에스터 결합
DNA와 RNA 골격을 이루는 인산다이에스터 결합은 뉴클레오사이드 사이의 인산기를 매개하며 핵산 복제, 복구, 재조합에 중요한 역할을 하고 pH 7에서 음전하를 띠며 특정 조건 하에 분해될 수 있는 화학 결합이다. - 유기 인산염 - 실로시빈
실로시빈은 환각 작용을 일으키는 화합물로, 뇌의 세로토닌 수용체에 작용하여 시각적 환각, 시간 감각 변화 등을 유발하며, 우울증, 불안 등의 치료에 대한 연구가 진행되고 있다.
2. 구조
리보스 5-인산은 5탄당인 리보스의 5번 탄소에 인산기가 결합되어 있는 화합물이다. 리보스 5-인산은 선형 또는 푸라노스 고리 형태로 존재할 수 있으며, 주로 푸라노스 형태로 존재한다.[18]
리보스 5-인산(R5P)의 합성은 세포 생장, 니코틴아마이드 아데닌 다이뉴클레오타이드 인산(NADPH), 리보스 5-인산, 아데노신 삼인산(ATP)의 필요에 따라 크게 달라진다. 각 분자의 생성은 오탄당 인산 경로와 해당과정 두 가지 대사 경로에서 포도당 6-인산(G6P)의 흐름에 의해 조절된다. 이 두 경로 사이의 관계는 다양한 대사 상황을 통해 확인할 수 있다.[19]
리보스 5-인산은 5개의 탄소로 이루어진 당인 리보스와 5번 탄소 위치에 있는 인산기로 구성된다. 리보스 5-인산은 열린 사슬 형태 또는 푸라노스 형태로 존재할 수 있으며, 푸라노스 형태가 가장 일반적이다.[1]
3. 생합성

3. 1. 오탄당 인산 경로
리보스 5-인산은 모든 생물에서 오탄당 인산 경로에서 생성된다.[19] 오탄당 인산 경로는 해당과정과 병행해서 진행되는 대사 경로이다. 오탄당 인산 경로는 환원성 생합성[20](예: 지방산 합성)에 대한 NADPH 및 오탄당의 중요한 공급처이다. 오탄당 인산 경로는 NADPH를 생성하는 산화적 단계와 당의 상호전환을 포함하는 비산화적 단계의 두 가지 단계로 구성된다. 오탄당 인산 경로의 산화적 단계에서 포도당 6-인산(G6P)을 리불로스 5-인산(RuBP)으로 전환시키는 과정을 통해 2분자의 NADP+를 2분자의 NADPH로 환원시킨다. 오탄당 인산 경로의 비산화적 단계에서, 리불로스 5-인산은 리보스 5-인산 이성질화효소의 촉매 작용을 통해 리보스 5-인산으로 전환될 수 있다.[21]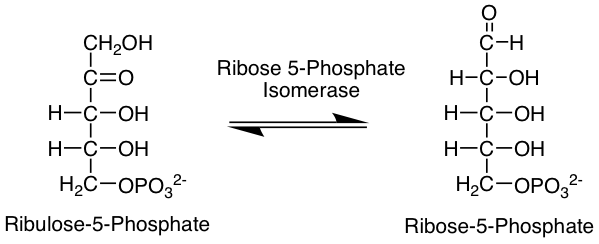
NADPH 및 리보스 5-인산에 대한 수요가 균형을 이루면 포도당 6-인산은 오탄당 인산 경로를 통해 한 분자의 리불로스 5-인산을 형성하여 두 분자의 NADPH와 한 분자의 리보스 5-인산을 생성한다.[19]
리보스-5-인산 이성질화 효소에 의해 리불로스-5-인산으로부터 생성되며, 트랜스케톨레이스에 의해 세도헵툴로스-7-인산과 글리세르알데히드-3-인산, 과당-6-인산으로 전환된다. 이 중 글리세르알데히드-3-인산과 과당-6-인산은 해당 과정의 중간 생성물이기도 하다. 또한, 리보스 인산 디포스포키나아제에 의해 포스포리보실 피로인산으로 전환된다.
3. 2. 해당과정
NADPH보다 더 많은 리보스 5-인산이 필요할 때, 리보스 5-인산은 해당과정의 대사 중간생성물을 통해 생성될 수 있다. 포도당 6-인산은 해당과정동안 과당 6-인산 및 글리세르알데하이드 3-인산으로 전환될 수 있다. 트랜스케톨레이스 및 트랜스알돌레이스는 2분자의 과당 6-인산과 1분자의 글리세르알데하이드 3-인산을 3분자의 리보스 5-인산으로 전환시킨다.[19] 빠른 세포 생장 동안 뉴클레오타이드 및 지방산 합성을 위해 더 많은 양의 리보스 5-인산 및 NADPH가 필요하다. 해당과정의 대사 중간생성물은 피루브산 키네이스 동질효소에 대한 유전자 발현에 의해 오탄당 인산 경로의 비산화적 단계로 유입될 수 있다. 피루브산 키네이스 동질효소는 해당과정에서 병목 현상을 일으켜 오탄당 인산 경로가 해당과정의 대사 중간생성물들을 사용하여 NADPH와 리보스 5-인산을 합성할 수 있도록 한다. 이러한 과정은 피루브산 키네이스 동질효소의 기질인 포스포엔올피루브산에 의한 삼탄당 인산 이성질화효소의 저해에 의해 더욱 활성화된다.[19]
글리세르알데하이드 3-인산과 과당 6-인산은 해당과정의 중간 생성물이기도 하다.
4. 기능
리보스 5-인산 및 그 유도체들은 DNA, RNA, ATP, 조효소 A, FAD, 히스티딘을 비롯한 많은 생체분자들의 전구체로 역할을 한다.[22][5] 리보스-5-인산 이성질화 효소에 의해 리불로스-5-인산으로부터 생성되며, 트랜스케톨레이스에 의해 세도헵툴로스-7-인산, 글리세르알데히드-3-인산, 과당-6-인산으로 전환된다. 글리세르알데히드-3-인산과 과당-6-인산은 해당 과정의 중간 생성물이기도 하다. 또한, 리보스 인산 디포스포키나아제에 의해 포스포리보실 피로인산으로 전환된다.[5]
4. 1. 뉴클레오타이드 생합성
뉴클레오타이드는 핵산(DNA와 RNA)의 단위체로 역할을 한다.[23] 뉴클레오타이드는 핵염기, 5탄당, 적어도 하나 이상의 인산기로 구성된다. 뉴클레오타이드는 퓨린 계열의 염기 또는 피리미딘 계열의 염기를 가지고 있다. 퓨린 뉴클레오타이드 합성에서 모든 대사 중간생성물들은 리보스 5-인산을 포함하고 있으며,[24] 리보스 5-인산은 피리미딘 뉴클레오타이드 합성에서 중요한 전구체로도 작용한다.뉴클레오타이드 생합성 과정 동안, 리보스 5-인산(R5P)은 리보스-인산 다이포스포키네이스(PRPS1)에 의해 포스포리보실 피로인산(PRPP)으로 전환된다. 포스포리보실 피로인산의 생성은 퓨린의 신생합성 경로 및 퓨린의 회수 경로에 필수적이다.[25] 신생합성 경로는 포스포리보실 피로인산의 리보스 5-인산 부분의 활성화로부터 시작되며, 이어서 뉴클레오타이드의 전구체인 포스포리보실아민으로 전환된다. 퓨린의 회수 경로에서[26] 포스포리보실트랜스퍼레이스는 포스포리보실 피로인산을 염기에 첨가한다.[27]
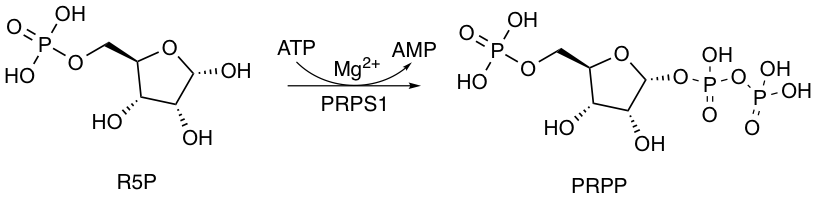
포스포리보실 피로인산은 또한 피리미딘 뉴클레오타이드 합성에서 중요한 역할을 한다. 피리미딘 뉴클레오타이드 합성의 5번째 단계에서 포스포리보실 피로인산(PRPP)의 리보스 부분의 1번 탄소는 오로트산과 공유 결합으로 연결된다. 이 반응은 오로트산 포스포리보실트랜스퍼레이스(PRPP 트랜스퍼레이스)에 의해 촉매되어, 오로티딘 일인산(OMP)을 생성한다.[25]
4. 2. 히스티딘 생합성
히스티딘은 사람에서 새롭게 합성되지 않는 필수 아미노산이다. 뉴클레오타이드처럼 히스티딘 생합성은 리보스 5-인산이 포스포리보실 피로인산(PRPP)으로 전환되면서 시작된다. 히스티딘 생합성의 속도 결정 단계는 ATP 포스포리보실트랜스퍼레이스에 의한 포스포리보실 피로인산과 ATP의 축합 반응이다. 히스티딘 생합성은 피드백 억제에 의해 조절된다.[28][11]4. 3. 기타 기능
리보스 5-인산은 리보스 5-인산 아데닐릴트랜스퍼레이스에 의해 아데노신 이인산 리보스로 전환될 수 있으며,[29] 아데노신 이인산 리보스는 TRPM2 이온 통로에 결합하여 이를 활성화시킨다.[12]리보스 5-인산(R5P)와 그 유도체는 DNA, RNA, ATP, 코엔자임 A, FAD (플라빈 아데닌 디뉴클레오타이드), 그리고 히스티딘을 포함한 많은 생체 분자의 전구체로 작용한다.[5]
또한, 리보스 5-인산은 리보스 인산 디포스포키나아제에 의해 포스포리보실 피로인산으로 전환된다.
5. 질병 관련성
세포 내 리보스 5-인산(R5P)의 불균형은 여러 질병과 관련이 있다. 암과 종양은 DNA와 RNA 합성 증가와 관련되어 리보스 5-인산 생성이 조절되는 경향을 보인다.[19]
리보스 5-인산 이성질화효소 결핍증은 리보스 5-인산 불균형과 관련된 질환이다.[30][31]
통풍은 포도당 6-인산 양이 증가하여 해당과정 대사 중간생성물들이 축적되고, 이것이 리보스 5-인산 생성으로 전환되면서 발생한다. 리보스 5-인산은 포스포리보실 피로인산(PRPP)으로 전환되어 퓨린의 과잉 생산을 유발하고, 이는 요산 축적을 야기한다.[25] 리보스-인산 다이포스포키네이스(PRPS1)의 과다활성은 통풍뿐만 아니라 신경발달 장애 및 감각신경성 난청과도 관련이 있다.[34]
레쉬-니한 증후군은 포스포리보실 피로인산(PRPP) 축적으로 인해 발생한다.[33] 이는 하이포잔틴-구아닌 포스포리보실트랜스퍼레이스(HGPRT) 결핍으로 발생하며, 뉴클레오타이드 합성을 감소시키고 요산 생성을 증가시킨다.
5. 1. 암
암과 종양은 RNA 및 DNA 합성이 증가함에 따라 R5P 생성이 증가하는 경향을 보인다.[2] 세계에서 가장 희귀한 질병인 리보스 5-인산 이성질체 결핍증[13][14] 또한 R5P 불균형과 관련이 있다. 이 질병의 분자 병리학은 잘 알려져 있지 않지만, RNA 합성이 감소하는 것이 가설로 제기되었다.R5P와 관련된 또 다른 질병은 통풍이다.[15] G6P의 높은 수치는 해당 과정 중간체의 축적으로 이어지고, 이는 R5P 생성으로 전환된다. R5P는 PRPP로 전환되고, 이는 퓨린의 과잉 생산을 유발하여 요산이 축적된다.[8]
PRPP 축적은 레쉬-니한 증후군에서 발견된다.[16] 이러한 축적은 효소 HGPRT의 결핍으로 인해 발생하며, 이는 뉴클레오타이드 합성을 감소시키고 요산 생성을 증가시킨다.
R5P를 PRPP로 촉매하는 효소인 PRPS1의 과잉 활성 또한 통풍뿐만 아니라 신경 발달 장애 및 감각 신경성 난청과 관련이 있다.[17]
5. 2. 리보스 5-인산 이성질화효소 결핍증
리보스 5-인산 이성질화효소 결핍증은 세포의 리보스 5-인산 불균형과 관련된, 세계에서 가장 드문 질환이다.[30][31] 이 질병의 분자병리학은 아직 잘 알려져 있지 않지만, RNA 합성 감소가 원인으로 제시되었다.5. 3. 통풍
포도당 6-인산의 양이 증가하면 해당과정의 대사 중간생성물들이 축적되어 리보스 5-인산의 생성으로 전환된다. 리보스 5-인산은 포스포리보실 피로인산(PRPP)으로 전환되어 퓨린의 과잉 생산을 야기하고 이로 인해 요산이 축적된다.[25] 리보스-인산 다이포스포키네이스(PRPS1)의 과다활성은 신경발달 장애 및 감각신경성 난청 뿐만 아니라 통풍과 관련이 있다.[34]5. 4. 레쉬-니한 증후군
레쉬-니한 증후군에서는 포스포리보실 피로인산(PRPP)이 축적된다.[33] 이러한 축적은 하이포잔틴-구아닌 포스포리보실트랜스퍼레이스(HGPRT) 효소의 결핍으로 인해 발생하며, 이는 뉴클레오타이드 합성을 감소시키고 요산 생성을 증가시킨다.5. 5. 기타 질환
세포의 리보스 5-인산 불균형은 여러 질병과 관련이 있다. 암과 종양은 DNA와 RNA의 합성 증가와 관련하여 리보스 5-인산의 조절된 생성을 보여준다.[19] 세계에서 가장 드문 질환인 리보스 5-인산 이성질화효소 결핍증은 리보스 5-인산의 불균형과 관련이 있다.[30][31] 리보스 5-인산 이성질화효소 결핍증의 분자병리학은 아직 이해되지 않고 있지만, RNA 합성 감소가 가설로 제시되었다. 리보스 5-인산과 관련된 또 다른 질병은 통풍이다.[32] 포도당 6-인산의 양이 증가하면 해당과정의 대사 중간생성물들이 축적되어 리보스 5-인산의 생성으로 전환된다. 리보스 5-인산은 포스포리보실 피로인산(PRPP)으로 전환되어 퓨린의 과잉 생산을 야기하고 이로 인해 요산이 축적된다.[25]포스포리보실 피로인산의 축적은 레쉬-니한 증후군에서 발견된다.[33] 이는 하이포잔틴-구아닌 포스포리보실트랜스퍼레이스(HGPRT)의 결핍으로 인해 발생하며, 뉴클레오타이드 합성을 감소시키고 요산 생성을 증가시킨다.
리보스 5-인산(R5P)을 포스포리보실 피로인산(PRPP)으로 전환시키는 반응을 촉매하는 효소인 리보스-인산 다이포스포키네이스(PRPS1)의 과다활성은 신경발달 장애 및 감각신경성 난청 뿐만 아니라 통풍과도 관련이 있다.[34]
참조
[1]
논문
The Synthesis of Ribose-5-Phosphoric Acid
http://www.jbc.org/c[...]
1934-02
[2]
서적
Biochemistry
W.H. Freeman
[3]
논문
The oxidative pentose phosphate pathway: structure and organisation
2003-06
[4]
논문
Structure of Escherichia coli ribose-5-phosphate isomerase: a ubiquitous enzyme of the pentose phosphate pathway and the Calvin cycle
2003-01
[5]
서적
X ''Pharm'': The Comprehensive Pharmacology Reference
[6]
서적
IUPAC Compendium of Chemical Terminology
International Union of Pure and Applied Chemistry
[7]
서적
Textbook of Veterinary Physiological Chemistry
2015
[8]
서적
Elsevier's Integrated Review Biochemistry
2011
[9]
서적
Textbook of Veterinary Physiological Chemistry
2015
[10]
서적
Phosphoribosyltransferase Mechanisms and Roles in Nucleic Acid Metabolism
[11]
논문
Histidine biosynthesis
2011-01
[12]
논문
Phosphorolysis of adenosine diphosphoribose
1966-01
[13]
논문
The difference between rare and exceptionally rare: molecular characterization of ribose 5-phosphate isomerase deficiency
https://research.vum[...]
2010-09
[14]
논문
Ribose-5-phosphate isomerase deficiency: new inborn error in the pentose phosphate pathway associated with a slowly progressive leukoencephalopathy
2004-04
[15]
서적
Gout & Other Crystal Arthropathies
2012
[16]
서적
Genetic Diseases of the Kidney
2009
[17]
서적
Movement Disorders in Childhood
2010
[18]
저널
The Synthesis of Ribose-5-Phosphoric Acid
http://www.jbc.org/c[...]
1934-02
[19]
서적
Biochemistry
W.H. Freeman
[20]
저널
The oxidative pentose phosphate pathway: structure and organisation
2003-06
[21]
저널
Structure of Escherichia coli ribose-5-phosphate isomerase: a ubiquitous enzyme of the pentose phosphate pathway and the Calvin cycle
2003-01
[22]
서적
X ''Pharm'': The Comprehensive Pharmacology Reference
[23]
서적
IUPAC Compendium of Chemical Terminology
International Union of Pure and Applied Chemistry
[24]
서적
Textbook of Veterinary Physiological Chemistry
2015
[25]
서적
Elsevier's Integrated Review Biochemistry
2011
[26]
서적
Textbook of Veterinary Physiological Chemistry
2015
[27]
서적
Phosphoribosyltransferase Mechanisms and Roles in Nucleic Acid Metabolism
[28]
저널
Histidine biosynthesis
2011-01
[29]
저널
Phosphorolysis of adenosine diphosphoribose
1966-01
[30]
저널
The difference between rare and exceptionally rare: molecular characterization of ribose 5-phosphate isomerase deficiency
2010-09
[31]
저널
Ribose-5-phosphate isomerase deficiency: new inborn error in the pentose phosphate pathway associated with a slowly progressive leukoencephalopathy
https://archive.org/[...]
2004-04
[32]
서적
Gout & Other Crystal Arthropathies
https://archive.org/[...]
2012
[33]
서적
Genetic Diseases of the Kidney
2009
[34]
서적
Movement Disorders in Childhood
https://archive.org/[...]
2010
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com