순각류
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
순각류(지네)는 다족류에 속하는 절지동물로, 각 몸 마디마다 한 쌍의 다리를 가지고 있으며, 턱다리가 변형된 집게발을 특징으로 한다. 전 세계 육상 생태계에 널리 분포하며, 다양한 환경에서 서식한다. 지네는 육식성이며, 턱다리의 독을 사용하여 먹이를 사냥한다. 생식은 정포를 통해 이루어지며, 일부 종은 부모의 보살핌을 제공한다. 지네는 5개의 목으로 분류되며, 화석 기록은 4억 3천만 년 전으로 거슬러 올라간다. 일부 문화권에서는 식용으로 사용되기도 하며, 한의학에서도 약재로 활용된다.

더 읽어볼만한 페이지
- 순각류 - 그리마
그리마는 길고 납작한 몸과 많은 다리를 가진 절지동물로, 습하고 어두운 곳을 좋아하며 작은 곤충을 잡아먹는 육식성 동물이고, 혐오감을 줄 수 있지만 해충을 잡아먹는 익충이기도 하며, 그리마과에는 †''Fulmenocursor''를 포함한 다양한 속이 있다. - 순각류 - 딱정그리마
딱정그리마는 다리가 많고 빠른 절지동물로, 해충을 잡아먹는 포식자이며 야행성으로 활동하고 지중해 지역이 원산지이다. - 유독절지동물 - 왕지네
왕지네는 붉은 머리와 암록색 몸통을 가진 8~15cm의 대형 지네로, 습한 환경에서 작은 동물을 잡아먹는 육식 곤충이며, 독샘을 이용해 먹이를 제압하고, 긴 수명을 가지며, 한약재 오공으로 사용되고, 항균 펩타이드가 아토피 피부염 치료에 효과적인 것으로 밝혀져 화장품 개발에 활용된다. - 다지아문 - 다지류
다지류는 많은 체절과 다리를 가진 절지동물 그룹으로, 지네강, 노래기강, 좀지네강, 에다히게무시강의 4개 강으로 나뉘며, 습한 숲에 서식하며 썩어가는 식물을 분해하는 역할을 한다. - 다지아문 - 결합강
결합강은 작고 눈과 색소가 없는 다지류로, 14마디 몸통을 가지며 짝짓기 없이 정포를 이용해 번식하는 독특한 방식을 가지고 있고 일부 종은 농업 해충으로 작물에 피해를 주기도 한다.

2. 특징
### 낫발 (Forcipules)
집게발(낫발, Forcipules)은 지네에게만 있는 독특한 구조이다.[3] 첫 번째 다리(턱다리)가 변형된 것으로, 머리 바로 뒤에 집게 모양의 부속지를 형성한다.[3] 집게발은 구강 구기는 아니지만, 독을 주입하고 먹잇감을 잡아서 제압하는 데 사용된다.[3] 독샘은 머리 안쪽에서부터 각 집게발 끝까지 튜브를 통해 이어진다.[3]
첫 번째 몸 마디는 턱다리 마디(forcipular segment)로, 그 등쪽은 일반적으로 독립된 짧은 등판(tergite) 한 장을 가지지만, 왕지네류에서는 그것이 없고, 바로 뒤의 제1 유각 몸 마디와 같은 등판으로 덮여 있다[68][71][101]。
독샘을 가지고, 턱과 같은 모양을 한 포식용 '''턱다리''' (forcipule, 또는 maxilliped[81], toxicognath, poison claw[82])는 이 몸 마디의 복면에서 앞으로 튀어나온다. 이것은 종종 "독아" 또는 "턱"으로 소개되어 왔지만, (머리에서 유래한) 턱이 아니라, 몸통에서 유래한 "특화된 다리"이며[72][71][73], 학명 "Chilopoda" 및 "순각류"의 이름에 나타나는 바와 같이, 지네에 특유하고 가장 중요한 공유파생형질로 여겨진다[71][83]。
턱다리의 기절은 제1 몸 마디의 복면을 차지할 정도로 넓고, 중앙의 복판과 함께 기흉판을 형성한다. 왕지네류에서는 기절은 흔적적인 복판에서 분절되어 가동적이며, 앞쪽 가장자리에 긴 강모가 늘어서 있지만[84], 다른 무리에서는 기흉판의 유합이 진행되어 복판은 보이지 않고, 약간만 움직일 수 있거나(돌지네류) 완전히 불동(긴발지네류·왕지네류·줄지네류)으로[129], 앞쪽 가장자리에 견고한 작은 이빨이 늘어서 있다[101][83]。 나머지 부분은 기부에서부터 trochanteroprefemur(전절 trochanter와 전퇴절 prefemur의 복합체)·퇴절(femur)·경절(tibia)·tarsungulum(부절 tarsus와 전부절 ungulum의 복합체)이라는 가동 가능한 4개의 마디로 이루어져 있으며[85][68][80][101],송곳니 모양의 tarsungulum의 말단 바로 전에 독샘의 개구부가 있다[83]。 왕지네류 이외의 무리에서는 tarsungulum이 특히 견고하고, 내연이 칼날처럼 뾰족하다[73]。 왕지네류(속의 일부 제외)와 줄지네류는 중간의 퇴절과 경절은 반환형으로 바깥쪽이 결여되어 있으며, 제1절과 제4절의 바깥쪽이 인접하고, 가동 가능한 4절의 관절이 모두 한 쌍의 관절구(피벗)에 집약된다[73][86]。 다른 무리에서는 환형의 퇴절과 경절을 가지고, 특히 왕지네류에서는 장대한 기둥 모양으로, 턱다리 전체가 걷는 다리 모양에 가깝다[87][101][73][86][83]。 기흉판의 유합 정도에 따라, 턱다리 전체의 운동 방향은 왕지네류에서는 3차원 방향으로 찌르듯이 위아래로, 다른 무리에서는 수평 방향으로 물어뜯듯이 좌우로 움직인다[73][86][83]。
독샘은 일반적으로 턱다리에 격납되지만, 줄지네류 중에서는 독샘이 몸통까지 뻗어 있는 예가 있다[101][83]。 신경독을 가질 것으로 생각되지만, 그 성분과 메커니즘에 관해서는 미해명된 부분이 많다[88]。
2. 1. 감각 기관
많은 지네 종은 눈이 없지만, 일부는 다양한 수의 홑눈을 가지고 있으며, 때로는 함께 뭉쳐서 진정한 겹눈을 형성하기도 한다. 그러나 이 눈은 밝고 어두움을 구별할 수 있을 뿐이며, 실제 시력을 제공하지 못한다.[1] 그물게류의 눈은 1쌍의 진짜 겹눈이지만,[74][75][76][80] 다른 무리는 홑눈이며, 돌거미류에 최대 49쌍, 왕지네에 최대 4쌍, 긴발거미류에 1쌍을 가진다.[101] 줄지네류는 모두 눈이 없으며,[71] 왕지네류나 돌거미류 중에도 눈이 없는 종류가 있다.[101][77]일부 종에서는 첫 번째 다리 한 쌍이 촉각과 유사한 감각 기관으로 기능할 수 있으며, 다른 대부분의 무척추동물의 촉각과는 달리 뒤로 향한다.[1]
토머스바리 기관은 촉각의 기저부에 위치하며, 원반 모양의 구조와 중심 구멍으로 구성되어 있으며, 민감한 세포들이 이를 둘러싸고 있다. 이는 진동을 감지하는 데 사용될 가능성이 높으며, 약한 형태의 청력을 제공할 수 있다.[1] 다족류의 머리 좌우에서 일반적으로 보이는 작은 구멍 모양의 측두 기관 (Tömösváry organ영어)은 그물게류와 돌거미류에서만 발달하고, 긴발거미류에서는 흔적만 남아 있으며, 왕지네류와 줄지네류에서는 보이지 않는다.[94][68][101][73]
2. 2. 낫발 (Forcipules)
집게발(낫발, Forcipules)은 지네에게만 있는 독특한 구조이다.[3] 첫 번째 다리(턱다리)가 변형된 것으로, 머리 바로 뒤에 집게 모양의 부속지를 형성한다.[3] 집게발은 구강 구기는 아니지만, 독을 주입하고 먹잇감을 잡아서 제압하는 데 사용된다.[3] 독샘은 머리 안쪽에서부터 각 집게발 끝까지 튜브를 통해 이어진다.[3]
첫 번째 몸 마디는 턱다리 마디(forcipular segment)로, 그 등쪽은 일반적으로 독립된 짧은 등판(tergite) 한 장을 가지지만, 왕지네류에서는 그것이 없고, 바로 뒤의 제1 유각 몸 마디와 같은 등판으로 덮여 있다[68][71][101]。
독샘을 가지고, 턱과 같은 모양을 한 포식용 '''턱다리''' (forcipule, 또는 maxilliped[81], toxicognath, poison claw[82])는 이 몸 마디의 복면에서 앞으로 튀어나온다. 이것은 종종 "독아" 또는 "턱"으로 소개되어 왔지만, (머리에서 유래한) 턱이 아니라, 몸통에서 유래한 "특화된 다리"이며[72][71][73], 학명 "Chilopoda" 및 "순각류"의 이름에 나타나는 바와 같이, 지네에 특유하고 가장 중요한 공유파생형질로 여겨진다[71][83]。
턱다리의 기절은 제1 몸 마디의 복면을 차지할 정도로 넓고, 중앙의 복판과 함께 기흉판을 형성한다. 왕지네류에서는 기절은 흔적적인 복판에서 분절되어 가동적이며, 앞쪽 가장자리에 긴 강모가 늘어서 있지만[84], 다른 무리에서는 기흉판의 유합이 진행되어 복판은 보이지 않고, 약간만 움직일 수 있거나(돌지네류) 완전히 불동(긴발지네류·왕지네류·줄지네류)으로[129], 앞쪽 가장자리에 견고한 작은 이빨이 늘어서 있다[101][83]。 나머지 부분은 기부에서부터 trochanteroprefemur(전절 trochanter와 전퇴절 prefemur의 복합체)·퇴절(femur)·경절(tibia)·tarsungulum(부절 tarsus와 전부절 ungulum의 복합체)이라는 가동 가능한 4개의 마디로 이루어져 있으며[85][68][80][101],송곳니 모양의 tarsungulum의 말단 바로 전에 독샘의 개구부가 있다[83]。 왕지네류 이외의 무리에서는 tarsungulum이 특히 견고하고, 내연이 칼날처럼 뾰족하다[73]。 왕지네류(속의 일부 제외)와 줄지네류는 중간의 퇴절과 경절은 반환형으로 바깥쪽이 결여되어 있으며, 제1절과 제4절의 바깥쪽이 인접하고, 가동 가능한 4절의 관절이 모두 한 쌍의 관절구(피벗)에 집약된다[73][86]。 다른 무리에서는 환형의 퇴절과 경절을 가지고, 특히 왕지네류에서는 장대한 기둥 모양으로, 턱다리 전체가 걷는 다리 모양에 가깝다[87][101][73][86][83]。 기흉판의 유합 정도에 따라, 턱다리 전체의 운동 방향은 왕지네류에서는 3차원 방향으로 찌르듯이 위아래로, 다른 무리에서는 수평 방향으로 물어뜯듯이 좌우로 움직인다[73][86][83]。
독샘은 일반적으로 턱다리에 격납되지만, 줄지네류 중에서는 독샘이 몸통까지 뻗어 있는 예가 있다[101][83]。 신경독을 가질 것으로 생각되지만, 그 성분과 메커니즘에 관해서는 미해명된 부분이 많다[88]。
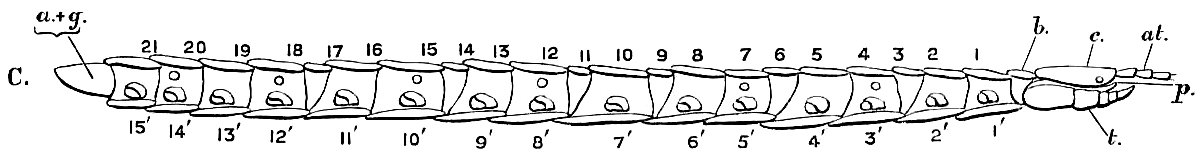
2. 3. 몸통
머리 뒤쪽에는 몸통이 최소 15개의 마디로 이루어져 있다.[1] 대부분의 마디에는 다리가 한 쌍씩 있으며, 턱다리는 첫 번째 몸통 마디에서 앞으로 뻗어 있으며, 마지막 두 마디는 작고 다리가 없다.[1] 각 다리 쌍은 앞선 쌍보다 약간 더 길어 겹치지 않도록 하여, 충돌하여 넘어질 가능성을 줄인다.[1] 마지막 다리 쌍은 첫 번째 쌍의 두 배 길이만큼 길 수 있다. 마지막 마디에는 미절이 있으며, 생식 기관의 구멍이 있다.[1]몸통부(胴部)는 세로로 긴 합체절로, 수많은 가동 가능한 체절(胴節, 몸통 마디)로 이루어져 있다.[1] 앞쪽에는 악지를 가진 악지절, 말단에는 2개의 생식절과 비체절성인 미절, 나머지 대부분은 각각 한 쌍의 다리를 양쪽 배면에 가진 홀수의 유각 胴節(종류에 따라 15~191節)이 늘어서 있다.
다리가 있는 체절, 이른바 유각체절(leg-bearing segment)은 모두 상하로 등판과 복판으로 덮여 있다.[68][101] 좌우는 유연한 절간막으로 덮여 있으며, 작은 측판(pleurite)이나 다리와 관절하는 아기절(subcoxa)이 산재한다.[94][68][80][101]
유각체절의 등판은 분류군에 따라 구조가 다르며, 애벌레지네류 이외에는 특히 변칙적이다(heterotergy).[129][64][144][145][90][101] 지네류는 8개, 돌지네류는 현저한 장단을 반복하고, 긴발돌지네류는 겉보기에는 체절과 일치하지 않는 21개로 세분되며, 왕지네류는 제1개가 턱다리 마디까지 덮는다.[94][71][101] 애벌레지네류는 미지 이외의 등판이 거의 같은 모양이며, 그 중 제1다리 이외의 각 등판은 더 앞뒤로 2개(앞쪽의 짧은 pretergite/intercalary tergite와 뒤쪽의 긴 metatergite)로 세분된다.[94][80][101] 돌지네류와 왕지네류의 제1·3·5다리와 제8 이후의 짝수 번째 유각체절은, 각각의 체절보다 등판이 길다.[91][94][144][145][90][101]
복판은 사변형에서 사다리꼴이며, 좌우가 다리의 기부에 인접하고, 앞뒤로 거의 같은 모양이다. 애벌레지네류의 경우, 다리를 가진 각 체절의 복판은 등판처럼, 앞쪽의 짧은 presternite/intercalary sternite와 뒤쪽의 긴 metasternite로 세분된다.[94][80][101]
미각을 가진 최종 유각 흉절 바로 뒤에는 여러 개의 짧은 흉절(postpedal segment)이 이어지지만, 등판과 복판의 퇴화가 진행되어 겉보기 분절은 불분명하므로 실제 흉절 수는 판단하기 어렵다. 적어도 외성기를 가진 2개의 생식절(genital segment)과 비체절성인 미절 (telson)[69]을 가지고 있는 것이 널리 인정되며, 지네류의 배아와 줄지네류의 초기 유체에서 비교적 뚜렷하게 나타난다.[106][94][107][101] 외성기의 구조는 암수에 따라 다르기 때문에 이 부분에서 성체의 암수를 구별할 수 있다.[101][108] 그러나 지네류의 경우, 이 부분은 퇴화되어 평소에는 미각의 유각 흉절에 격납되어 등쪽에서는 관찰할 수 없다.[101]
제1생식절의 양 복면에 있는 한 쌍의 생식지 (gonopod)는 수컷보다 암컷이 더 발달되어 있으며, 이는 그리마류와 돌지네류에서 특히 현저하다.[106][94] 제2생식절은 생식공 (genital opening, gonopore)을 가지는 단일 외성기(암컷의 산란 구 vulva 혹은 수컷의 음경 penis)를 복면 중앙에 가지며, 그리마류의 수컷에서는 2쌍의 생식지를 양 복면에 가진다.[106][94][68][71][92][80][101] 말단의 미절은 눈에 띄지 않으며, 1개의 눈에 띄지 않는 등판을 배면에, 그리고 항문을 덮어 씌운 한 쌍의 판상 구조(anal valve)를 복면에 가진다.[80][101]
일부 문헌에서는 최종 유각 흉절과 제1생식절 사이에 더 1개의 흉절(intermediate segment)을 가진다고 하여, 이것을 근거로 그리마류·돌지네류·줄지네류의 생식절 바로 앞의 등판을 이 체절 유래(intermediate tergite)라고 주장하지만[68], 회의적이며, 앞서 언급한 지네류의 배아와 줄지네류의 초기 유체에 그러한 체절이 보이지 않는다.[101]
2. 4. 궁극다리 (Ultimate legs)
첫 번째 다리 한 쌍이 낫발로 변형되는 것처럼, 뒤쪽 다리도 "궁극다리"로 변형되며, 이는 꼬리 다리, 항문 다리, 말단 다리라고도 불린다.[4] 종에 따라 사용법이 다르지만, 이동에는 사용되지 않는다.[5][6] 궁극다리는 길고 얇거나, 두껍거나, 또는 집게 모양일 수 있다.[6]
이들은 종종 성적 이형성을 보이며, 짝짓기 의식에 중요한 역할을 할 수 있다.[5][6] "걷는" 다리보다 궁극다리에 더 자주 샘 구멍이 나타나므로, 감각 역할을 할 수 있다.[5][6] 때로는 방어 자세에 사용되며, 일부 종은 이를 사용하여 먹이를 잡거나, 포식자로부터 자신을 방어하거나, 나뭇가지와 같은 물체에 매달릴 때 집게처럼 사용한다.[5] 몇몇 종은 다른 지네를 만났을 때 궁극다리를 사용하여 다른 지네의 몸을 잡으려고 시도한다.[7]
''Alipes'' 속의 구성원들은 잎사귀 모양의 궁극다리로 stridulate하여 포식자를 산만하게 하거나 위협할 수 있다.[7] ''Rhysida immarginata togoensis''는 궁극다리를 흔들 때 희미한 삐걱거리는 소리를 낸다.[7]
2. 5. 노래기와의 차이점
지네와 노래기는 모두 다지류에 속하지만, 여러 가지 차이점이 있다.[8] 두 그룹은 길고, 여러 마디로 나뉜 몸, 많은 다리, 한 쌍의 안테나, 그리고 안테나 후 기관이 존재한다는 공통점을 가지고 있다.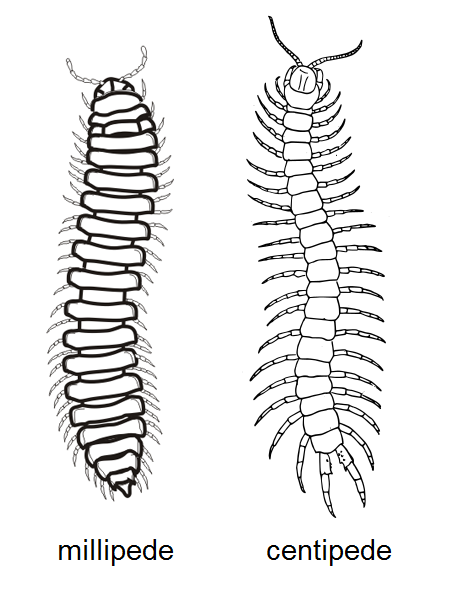
지네는 각 마디마다 한 쌍의 다리가 있는 반면, 노래기는 두 쌍의 다리가 있다. 머리 부분에서도 차이가 있는데, 노래기는 짧고 굽은 안테나, 한 쌍의 튼튼한 턱, 그리고 입술로 융합된 한 쌍의 아래턱을 가지고 있다. 반면 지네는 길고 실 모양의 안테나, 한 쌍의 작은 턱, 두 쌍의 아래턱, 그리고 한 쌍의 큰 독 이빨을 가지고 있다.[9]
지네는 다른 절지동물의 왁스 방수 표피가 없기 때문에 증발을 통한 수분 손실에 더 취약하다.[32] 따라서 지네는 탈수를 피하기 위해 습도가 높은 환경에서 가장 흔하게 발견되며,[33] 대부분 야행성이다.[34]
3. 생태
지네는 토양과 낙엽을 포함한 다양한 서식지에서 살며, 열대 우림,[20] 사막,[35] 그리고 동굴[36]과 같이 다양한 환경에서 발견된다. 일부 지네류는 연안 서식지에 적응하여 따개비를 먹고 산다.[37] 극지를 제외하고, 지네는 전 세계의 육지에 분포하며, 열대 우림에서는 가장 다양하게 분포한다[71]。대부분이 야행성이며 삼림 속 낙엽, 썩은 나무, 돌 밑 등 습도가 높은 장소에 서식하지만, 초원, 사막, 동굴, 해안 등에 서식하는 종류도 몇몇 있다[68][71][110]。
이동할 때, 지네는 앞쪽의 촉각으로 주변을 탐지하며, 양쪽 줄의 보행지를 좌우 상호 간에 물결치듯이 움직여 민첩하게 걷는다[145]。말단의 견인지는 보행에 사용하지 않으며, 분류군에 따라 감각, 위협, 방어, 포식, 배우자 행동, 머리(촉각)에 의태, 이동 중의 균형 조절 등의 기능을 수행한다[111][102][112][109]。촉각이나 다리를 자주 손질하며, 악지로 그것을 한 개씩 잡으면서 제1소악으로 청소한다[68]。
지네의 활동 양식은 지표면을 달리는 배회성에서 흙 속을 파고 들어가는 지하성까지 알려져 있으며, 긴발지네류와 돌지네류는 전자에, 줄지네류는 후자에 해당하며, 긴발돌지네류와 많은 왕지네류는 양자의 중간 정도에 해당한다[145][101]。긴발지네류는 주로 트인 장소를, 다른 무리는 주로 낙엽이나 지하 등 좁은 토양 환경을 선호하여 서식하며, 이는 각자의 신체 구조나 포식 행동(후술)과 크게 관련된다[144][145][73]。긴발지네류의 몸통은 콤팩트한 구조의 등판에 의해 이동 중의 흔들림이 억제되어, 가늘고 긴 다리로 트인 장소를 안정적으로 달리는 데 적합하다[145]。다른 무리에서는 평평한 몸을 이용하여 좁은 환경을 진입하며, 그중에서 돌지네류는 비교적 콤팩트한 체형(등판이 현저하게 장단으로 반복된 짧은 몸통)과 약간 긴 다리로 긴발지네류와 비슷한 안정성을 얻을 수 있지만, 긴발돌지네류, 왕지네류, 줄지네류에서는 몸통이 가늘고 긴 형태와 세분화된 등판과 복판에 의해 폭넓은 가동 범위를 얻을 수 있다[145]。특히 지하성인 줄지네류는 전진과 후퇴 모두 민첩하며, 다리가 짧고 몸마디의 신축성도 뛰어나 흙 속의 좁은 틈새를 진행하는 데 적합하다[145][68]。
지네는 대부분 단독 생활을 하는 육식 동물이며, 매복하거나 우연히 마주친 곤충 등의 작은 동물을 포식한다.[71][73]. 전반적으로 편식성이 없는 제너럴리스트로 여겨지지만, 스페셜리스트의 예도 드물게 알려져 있다(다족류를 전문적으로 포식하는 속의 일종 등). 썩은 고기를 섭취하기도 하며[113], 인공 사육에 의한 관찰에서는 굶주린 경우 식물 조직을 섭취하는 기록도 있다[114]。
턱다리는 모든 지네에게 주요한 포식 기관이며, 먹이를 포획하여 끝에서 독을 주입하여 마비시키는 역할을 한다[71][83][86]。 먹이의 씹기는 주로 큰턱과 작은턱으로 이루어진다[73]。 다른 구체적인 포식 행동은 분류군에 따라 다양한데, 다음과 같다.
천적으로는 조류, 파충류, 포유류 등의 척추동물, 그리고 다른 육식 절지동물 등이 있다. 지네류를 전문적으로 포식하는 육식동물로는 라는 뱀의 일종[114]과 (=) 라는 개미류를 즐겨 포식하는 개미의 일종[121]이 있다.
상대를 물어 자위하는 습성을 가지고 있지만, 어떤 지네류도 자극이나 위험에 처하면 반격보다는 도주하려 한다[88]. 줄지네류 이외의 종류는, 전절의 특수한 갈라진 틈새에서 다리를 자절할 수 있으며, 게지류에서 특히 자주 행해진다[96]。일부 종류, 특히 게지류와 왕지네류는 몸통 말단이 머리와 비슷한 색이나 촉각과 같은 견인지를 가지고 있는데, 이는 머리와의 구별을 어렵게 하는 자기 의태 (automimicry) 효과를 발휘하여, 이를 노리고 포살하는 포식자를 혼란시키는 특징이라고 생각된다[111]。돌지네류는 최종 몇 쌍의 다리를 위아래로 흔들어 점액을 분비하여, 거미나 개미 등의 소형 포식자로부터의 공격을 방어한다[102][112]。왕지네류는 최종 몇 쌍의 다리를 들어 올려 위협하는 경우가 있다[111]。''Edentistoma''속은 형태·행동 모두 노래기를 의태하며, 등판은 경화되어 움직임이 느리고, 자극을 받으면 몸을 둥글게 만다[122]。줄지네류는 배판에 있는 특수한 샘(sternal gland)에서 점성이 있는 화학 물질을 분비하여 몸을 보호한다[123][124]。
지네는 주로 다식성 포식자로, 다양한 먹이를 섭취하도록 적응되어 있다. 흔한 먹이로는 지렁이, 파리 유충, 톡토기 및 다른 지네 등이 있다. 육식성이며, 위 내용물을 연구한 결과 식물성 물질은 지네의 식단에서 중요하지 않은 부분이지만, 실험실 실험에서 굶주린 상태에서는 식물성 물질을 섭취한다.
''큰지네목'' 종, 특히 ''Scolopendra'' 와 ''Ethmostigmus'' 속의 구성원들은 거대한 무척추동물과 지네 자체보다 더 클 수 있는 상당한 크기의 척추동물을 포함한 먹이를 사냥할 수 있다. 예를 들어, ''Scolopendra gigantea'' (아마존 거대 지네)는 타란툴라, 전갈, 도마뱀, 개구리, 새, 생쥐, 뱀 및 심지어 박쥐까지 먹이로 삼아 비행 중에 잡기도 한다. 세 종 (''Scolopendra cataracta'', ''Scolopendra paradoxa'', 그리고 ''Scolopendra alcyona'')는 양서류로, 수생 또는 양서류 무척추동물을 사냥하는 것으로 생각된다.
많은 대형 동물들이 지네를 잡아먹는데, 예를 들어 몽구스, 쥐, 도롱뇽, 딱정벌레, 그리고 일부 특수 뱀 종이 있다. 지네는 많은 종의 중요한 먹이원이며, 아프리카 개미 ''Amblyopone pluto''와 같은 일부 종의 주식이며, 이 개미는 지질류 지네만을 먹고 산다.[26] 남아프리카 케이프 검은머리뱀 ''Aparallactus capensis''도 지네를 주식으로 한다.
일부 지네류(Geophilomorph), 돌지네류(Lithobiomorph), 곱등이 지네류(Scolopendromorph)는 자신을 방어하기 위해 끈적이고 독성 있는 분비물을 생성한다. 다양한 분비물은 포식자를 쫓아내거나 얽히게 한다.[27][28] 곱등이 지네류의 분비물에는 청산가리(hydrogen cyanide)가 포함되어 있다.[29]
지네류 중 ''Geophilus vittatus''의 분비물은 끈적거리고 냄새가 나며 청산가리가 포함되어 있다.[30]
애리조나의 거대 사막 지네인 ''Scolopendra polymorpha''는 머리와 꼬리가 검고 몸통이 주황색이다. 이 눈에 띄는 패턴은 동물의 독성을 나타내는 정직한 신호인 경계색(Aposematism)일 수 있다.[31] 많은 종들이 최종 다리를 들어 올리고 벌리며 다리에 있는 가시를 방어적인 위협 자세로 보여준다.[7]
3. 1. 번식 및 생활사
지네의 생식은 교미를 수반하지 않는다. 수컷은 정포를 낳고, 암컷은 이것을 섭취하여 번식한다.[31] 온대 지역에서는 봄과 여름에 알을 낳으며, 몇몇 종은 처녀생식을 한다.[31] 암컷은 알과 새끼를 몸으로 감싸고, 곰팡이와 박테리아를 제거하기 위해 새끼를 돌보는 등 부모의 보살핌을 제공한다.[31] 
지네는 곤충보다 수명이 길다. 유럽의 ''Lithobius forficatus''는 5~6년,[10] ''Scolopendra subspinipes''는 10년 이상 살 수 있다.[10] 지네는 적은 수의 알을 낳고, 임신 기간과 생식까지의 발달 기간이 길어 K-선택 종으로 분류된다.[11]
지네는 발달 과정에서 여러 단계를 거치며 다리를 성장시킨다. Lithobiomorpha강, Scutigeromorpha강, Craterostigmomorpha강은 비완전 변태를 통해 탈피 과정에서 더 많은 분절과 다리 쌍을 성장시킨다.[12] 예를 들어, 집지네 ''Scutigera coleoptrata''는 다리 4쌍만 가지고 부화하며, 5, 7, 9, 11, 15쌍을 거쳐 성체가 된다.[12] 다리가 15쌍 미만인 생활 단계를 유충기, 다리가 완전히 갖춰진 후를 후유충기라고 한다.[12] Craterostigmomorpha강은 비완전 변태의 한 단계만 거치며, 배아는 12쌍, 성체는 15쌍을 갖는다.[14]
분기도인 Epimorpha는 Geophilomorpha강과 Scolopendromorpha강으로 구성되며, 배아 단계에서 모든 다리 쌍이 발달하고, 자손이 탈피 사이에 더 많은 다리를 발달시키지 않는다.[13] Geophilomorpha강은 흉부 분절의 수가 종 내에서 다르며, 암컷이 수컷보다 더 많은 다리를 가지고 있다.[13] 다리 분절의 수는 Scolopendromorpha강에서는 21개 또는 23개, Geophilomorpha강에서는 27개에서 191개이다.[13] 지네의 분절은 두 단계로 발달하는데, 첫째, 머리는 홀수의 분절을 생성하고, 둘째, 꼬리 끝에 쌍의 분절이 추가된다.[19][18]
배우자의 교환은 교미가 아닌, 정포(spermatophore)의 전달을 통해 이루어진다.[102] 암컷은 말단의 외성기로 수컷의 정포를 받는다. 암수는 원을 그리듯이 서로의 말단을 향해 마주보면서, 더듬이로 상대의 견인지와 접촉한다.[102] 난생이며, 땅지네류, 왕지네류, 긴발돌지네류의 암컷 어미는 육아 습성을 가진다.[125][126] 지네류와 돌지네류의 암컷 어미는 육아를 하지 않지만, 산란 시 발달한 생식지로 알의 위치를 조절한다.[92]
유체의 성장 양식은 분류군에 따라 다르다. 지네류, 돌지네류, 긴발돌지네류, 이른바 개형류 (Anamorpha)의 초령 유체는 성체보다 적은 수의 다리와 체절로 태어나며, 성장할 때마다 탈피를 거쳐 그 수를 뒤에서부터 늘려간다.[71] 지네류와 돌지네류는 여러 번의 탈피를 거쳐 15쌍에 달하지만, 긴발돌지네류는 한 번만으로 15쌍이 된다.[117] 왕지네류와 땅지네류는 이른바 정형류 (Epimorpha)이며, 부화부터 이미 성체와 같은 다리와 체절 수를 갖추고 있다.[71] 결손된 부속지는 다음 탈피에서 재생된다.[68][96]
4. 분류
지네는 크게 5개의 목으로 분류된다. 절지동물 내에서 지네는 현존하는 강 중에서 가장 먼저 최근 공통 조상으로부터 분기된 것으로 여겨진다. 지네의 5개 목은 Craterostigmomorpha, Geophilomorpha, Lithobiomorpha, Scolopendromorpha, Scutigeromorpha이다. 이 목들은 다음과 같은 공통파생형질에 의해 분류군 Chilopoda로 통합된다:[52]
# 첫 번째 두부 후속 부속지는 독액 발톱으로 변형된다.
# 두 번째 턱다리 부속지 배아 큐티클에는 난치가 있다.
# 돌기–전대퇴마디 관절이 고정되어 있다.
# 정자 핵에는 나선형 능선이 발생한다.
Chilopoda는 다시 두 개의 분류군으로 나뉜다: Scutigeromorpha를 포함하는 Notostigmophora와 나머지 네 개의 목을 포함하는 Pleurostigmophora. 다음의 물리적 및 발달적 특징을 사용하여 Pleurostigmomorpha 구성원을 Notostigmomorpha와 구분할 수 있다:[53][54]
- 기문은 지네의 측면에 위치한다(Notostigmomorphs에서는 등쪽에 위치).
- 기문은 깊고 더 복잡하며 항상 쌍으로 존재한다.
- 머리가 약간 더 평평하다.
- 지네는 불완전변태 또는 완전변태를 거쳐 발달할 수 있다.
과거에는 순각강(Chilopoda)이 발달 방식을 기준으로 Anamorpha (Lithobiomorpha와 Scutigeromorpha)와 Epimorpha (Geophilomorpha와 Scolopendromorpha)로 나뉘며, Craterostigmomorpha의 관계는 불확실하다고 여겨졌다.
최근에는 분자적, 형태적 특징을 결합한 계통 발생 분석은 이전의 계통 발생을 뒷받침한다. Epimorpha는 Pleurostigmophora 내에서 여전히 단계통군으로 존재하지만, Anamorpha는 분기도에서 볼 수 있듯이 측계통군이다:[52]




절지동물 중에서 지네(지네강/순각강 킬로포다/Chilopodala)는 노래기(노래기강/다족강/Diplopodala)・얼치기지네(얼치기지네강/합구강/Symphylala)・다족강벌레(다족강벌레강/빈절강/Pauropodala)와 함께 다족류(다족아문 미리아포다/Myriapodala)에 분류되는 강 중 하나이다. 다족류 중에서 지네와 얼치기지네를 근연으로 하는 (트리그나타/Trignathala/아텔로포다/Atelopodala를 이룬다) 설도 있었지만, 지네를 후성류 (오피스토고네아타/Opisthogoneatala, 지네상강이라고도 함), 얼치기지네・다족강벌레・노래기를 전성류 (Progoneata, 노래기상강이라고도 함)로 나누는 설이 많은 형태학과 분자계통학의 견해를 지지한다.[127][95][101] 이름에서 볼 수 있듯이, 후성류는 생식공을 몸통의 후방에, 전성류는 생식공을 몸통의 전방에 배치한다. 한편, 지네와 노래기는 자매군이라는, 지금까지 제창되지 않았던 유연관계를 시사하는 분자계통 분석 결과도 약간 있다.[128]
3,000종 이상의 지네가 알려져 있으며, '''게지'''(게지목 스쿠티게로모르파/Scutigeromorphala), '''돌지네'''(돌지네목 리토비오모르파/Lithobiomorphala), '''긴발지네'''(긴발지네목 크라테로스티그모모르파/Craterostigmomorphala), '''왕지네'''(왕지네목 스콜로펜드로모르파/Scolopendromorphala), '''참지네'''(참지네목 게오필로모르파/Geophilomorphala)라는 5개의 군(목)으로 크게 나뉜다.[63]



21세기 이후의 형태학과 분자 계통 분석에서는 등기문류/측기문류 설과 정형류 설이 널리 인정된다.[131][132][135][133][128]
한국에 서식하는 왕지네목(Scolopendromorpha) 종으로는 베트남왕지네(''Scolopendra subspinipes'')[146][147], 장수지네(''Otostigmus polytus''), 홍지네(''Octocrytops sexspinosus''), 꼬부랑다리장님지네(''Cryptops japonica'')가 있다.[71]
4. 1. 한국에 서식하는 왕지네목(Scolopendromorpha) 종
한국에 서식하는 왕지네목(Scolopendromorpha) 종으로는 베트남왕지네( ''Scolopendra subspinipes'')[146][147], 장수지네(''Otostigmus polytus''), 홍지네(''Octocrytops sexspinosus''), 꼬부랑다리장님지네(''Cryptops japonica'')가 있다.[71] 왕지네목은 800종 이상을 포함하며, 지네로 가장 일반적으로 알려진 목이다. 대부분 10cm 전후의 대형이며, 강한 신경독과 먹이에 대한 높은 공격성을 가진다. 수컷은 정포를 낳고, 암컷은 알과 유체를 기른다. 증절변태를 하지 않으며, 유체는 성체와 동일한 체절과 다리 수로 태어난다.5. 진화
지네의 화석 기록은 4억 3천만년 전 후기 실루리아기 (''Crussolum'')로 거슬러 올라가지만,[42][43] 고생대 전반에 걸쳐 희귀하다.[48] 데본기 팬서 마운틴 층에는 매미류인 ''Crussolum''의 한 종과 멸종된 그룹 Devonobiomorpha에 포함된 ''Devonobius'' 두 종의 지네가 포함되어 있다.[44][45] 라이니 처트에서도 ''Crussolum'' 화석과 매미류 머리 부분의 화석이 발견되었다.[44] 한때 가장 오래된 곤충 화석으로 여겨졌던 ''Rhyniognatha''도 라이니 처트에서 발견되었다.[46] 석탄기인 메이존 크릭 화석 매장지에서는 한 종의 매미류(''Latzelia'')와 두 종의 스콜로펜드라류(''Mazoscolopendra'' 및 ''Palenarthrus'')가 발견되었다.[47] 중생대에는 스콜로펜드라류와 매미류를 포함하여 더 많은 종이 나타난다.[48][49]
지네는 절지동물문 내에서 현존하는 강 중에서 가장 먼저 최근 공통 조상으로부터 분기된 것으로 여겨진다.[50][51]
모든 지네는 독(venom)을 가지고 있다. 지네 분류군(clade)의 진화 역사 초창기 5천만 년 동안, 지네의 독은 약 4가지 성분으로 이루어진 단순한 혼합물이었던 것으로 보이며, 현재 인정되는 5개 목(order)이 발달한 후에야 특정 독 유형으로의 분화가 일어난 것으로 보인다. 독의 진화는 세균, 곰팡이, 난균을 포함하는 수평 유전자 전달을 통해 진화하였다.[55]
6. 인간과의 관계

일부 문화권에서는 지네를 식용으로 사용하기도 한다. 중국에서는 특정 대형 지네를 꼬치에 꿰어 구워 먹거나 튀겨 먹으며, 베이징의 둥화먼과 왕푸징 시장 등 대도시 길거리 노점에서 흔히 볼 수 있다.[56][57] 대형 지네는 지네 보드카를 만들기 위해 술에 담가 사용되기도 한다.[58]
한방에서는 지네를 오공(蜈蚣)이라고 부르며, 평간・지경・해독소종의 효과가 있다고 알려져 있다. 기름에 절이거나 건조시킨 지네는 화상이나 베인 상처에 효과가 있다고 하여 민간 약으로 사용되기도 한다.
산업과의 관련은 적다. 지네류는 지역에 따라 식용으로 사용되며, 오스트레일리아 원주민에게는 전통적인 조미료의 원료로 사용된다[155]. 관상어 등의 먹이로 냉동 지네가 수입되어 시판되고 있다.
일부 지네 종은 물리면 통증과 부기를 유발할 수 있으므로 주의해야 한다. 성인에게 물린 경우 매우 고통스럽고 심한 부기, 오한, 발열, 쇠약을 유발할 수 있지만 치명적일 가능성은 낮다. 하지만 어린 아이나 벌침 알레르기가 있는 사람에게는 위험할 수 있으며, 큰 지네의 독성 물림은 아나필락시스 쇼크를 유발할 수 있다.[59] 작은 지네는 일반적으로 인간의 피부를 뚫을 수 없다.[59]
인간의 생활과 문화에 관련된 지네로서 왕지네류가 특히 대표적이며, 세상 일반에서 지네에 대한 이미지를 형성하고 있다. 유독하고 흉폭한 습성으로 두려움의 대상이 되어, 세계적으로도 몇몇 신화나 전설의 주제가 되기도 한다.[83]
지네는 사람에게 자극을 받을 때마다 몸을 보호하기 위해 턱다리로 사람을 물어 반항하는 경우가 있다. 교상은 대부분 왕지네류에 의해 발생하며, 그 외의 지네류에 의한 경우는 드물다.[88]
대형 왕지네류에 물리면 상당히 아프지만, 인명 피해나 후유증은 거의 없다.[88][154] 그러나 어린이와 아나필락시스 쇼크를 일으키는 체질을 가진 사람에게는 위험성이 높으므로[154], 물린 경우에는 즉시 의사의 진찰을 받는 것이 바람직하다. 주로 여름철, 산림에 가까운 민가에서는 바퀴벌레 등을 포식하기 위해 지네가 종종 가옥 내부로 침입한다. 이 경우, 신발 안이나 침구에 숨어 있다 물리는 경우가 많아 위생 해충으로서 주의가 필요하다. 물린 경우, 환부에 이물질이 남아있으면 이를 제거하고 독을 짜내며, 환부를 수돗물로 세척한다. 항히스타민제를 도포한다.[150][151] 긴발지네는 가옥에 침입하는 경우도 있지만, 독성이 약하고 적극적으로 사람을 물지 않으므로 기본적으로 무해하다고 여겨지며[152], 만약 물린다고 하더라도 증상은 꿀벌의 침에 쏘이는 정도만큼 심하지 않다.[153] 소형 지네는 기본적으로 사람의 피부를 관통할 수 없다.[154]
19세기 티베트 시인은 "만약 당신이 다른 사람들을 두려워하는 것을 즐긴다면, 당신은 지네로 환생할 것입니다."라고 경고했다.[61]
일부 종류는 실내 환경에 침입하기도 한다. 이 경우, 유독생물로 널리 알려진 왕지네류처럼, 높은 방어 기제로 자극받을 때마다 물기도 하고, 또는 지네류처럼 단순히 빠른 움직임과 기이한 모습으로 해충 취급을 받기도 한다.[67] 지네류에 관해서는, 위생적으로 무해하고 실내의 위생 해충을 사냥함으로써 익충으로 여겨지기도 한다.[67]
"매우 흉폭하고 공격성이 강하다"는 이미지, "절대로 뒤로 물러서지 않는다(후퇴하지 않는다)"는 속신, 다리가 많은 것에서 "병사가 많다(대군)"는 연상, 많은 알을 낳아 품어 새끼를 키우는 성질을 "자손 번영"으로 해석하는 등, 무사가 좋아하는 성질을 가지고 있었기 때문에, 전국 시대에는, 투구의 앞장식, 기사물, 갑주나 칼 장식의 장식 등, 지네 디자인을 도입하는 것이 유행했다[156]. 또한 다리가 많은 것으로 인해, 전령을 지네에 비유하기도 했다.
『고요군감』에 따르면 다케다 가문의 금 채취 무리는 터널 전법을 특기로 하는 공병 부대로, 백족중이라고 불렸다고도 한다. 큰 뱀이 하천을 상징하고, 사철의 채집이나 제철의 기술자 집단을 나타내는 것과 비교하여, 지네는 지하 갱도를 파고 들어가, 자연금 등의 광석을 채집하는 기술자 집단을 나타낸다는 설이 있다[157].
소마 나카무라 번에 기원하는 소마 노마오이에서는, "내려가는 지네" 깃발이 군사의 지시물로 지정되어 있다.
아카기 산 등의 신체로서, 또한 『비사문천』의 사자로 여겨져, 신격화되어 있다. 상가에서도, 지네의 많은 다리에서 "손님이 많다", 강한 공격성에서 "다른 가게에 지지 않는다"는 의미로 가문이나 가게 문양, 가게 이름으로 삼는 경우가 있었다[156][158] . 미야기현의 남부에서는 양잠업의 대적인 쥐가 지네를 싫어한다는 전설과 비사문천의 사자라는 이야기가 합쳐져, 양잠업자들의 신앙을 얻은 예도 있다[156] .
남체산의 대사와 닛코의 센조가하라에서 결투한 전설, 후지와라노 히데사토에 의한 미카미 산의 대지네 퇴치 전설 등이 알려져 있다[159] .
곤충이나 거미, 전갈 등과 마찬가지로, 액세서리나 그래픽의 모티프가 되거나, 어린이용 그림책의 캐릭터로 지네가 등장하는 경우도 있다 (무스티 등).
=== 위협 ===
IUCN 적색 목록에 따르면, 지네는 취약종 1종, 위기종 6종, 심각한 위기종 3종이 있다.[38] 예를 들어, 세르펜트 섬 지네(''Scolopendra abnormis'')는 취약종이며, 터크 땅지네(''Nothogeophilus turki'')와 세이셸 긴 다리 지네(''Seychellonema gerlachi'')는 모두 위기종이다.[39][40][41]
=== 한국 문화 속 지네 ===
한국에서는 예로부터 지네가 혐오 동물로 여겨져 왔으며, 다양한 민담과 속담에 등장한다.[156][158][159] 지네는 약재로 사용되기도 했는데, 특히 화상이나 뱀에 물린 상처에 효과가 있다고 알려져 있다. 일부 지역에서는 지네를 퇴치하기 위한 민간요법이 전해져 내려오기도 한다. 무사가 좋아하는 성질을 가지고 있었기 때문에, 전국 시대에는, 투구의 앞장식, 기사물, 갑주나 칼 장식의 장식 등, 지네 디자인을 도입하는 것이 유행했다.[156] 또한 다리가 많은 것으로 인해, 전령을 지네에 비유하기도 했다.
6. 1. 위협
IUCN 적색 목록에 따르면, 지네는 취약종 1종, 위기종 6종, 심각한 위기종 3종이 있다.[38] 예를 들어, 세르펜트 섬 지네(''Scolopendra abnormis'')는 취약종이며, 터크 땅지네(''Nothogeophilus turki'')와 세이셸 긴 다리 지네(''Seychellonema gerlachi'')는 모두 위기종이다.[39][40][41]6. 2. 한국 문화 속 지네
한국에서는 예로부터 지네가 혐오 동물로 여겨져 왔으며, 다양한 민담과 속담에 등장한다.[156][158][159] 지네는 약재로 사용되기도 했는데, 특히 화상이나 뱀에 물린 상처에 효과가 있다고 알려져 있다. 일부 지역에서는 지네를 퇴치하기 위한 민간요법이 전해져 내려오기도 한다. 무사가 좋아하는 성질을 가지고 있었기 때문에, 전국 시대에는, 투구의 앞장식, 기사물, 갑주나 칼 장식의 장식 등, 지네 디자인을 도입하는 것이 유행했다.[156] 또한 다리가 많은 것으로 인해, 전령을 지네에 비유하기도 했다.참조
[1]
서적
Invertebrate Zoology
Holt-Saunders International
[2]
서적
Reference Module in Life Sciences
Elsevier
[3]
논문
Variation and specialisation of the forcipular apparatus of centipedes (Arthropoda: Chilopoda): A comparative morphometric and microscopic investigation of an evolutionary novelty
https://www.scienced[...]
2012-05-01
[4]
논문
A common terminology for the external anatomy of centipedes (Chilopoda)
2010-11-18
[5]
논문
The ultimate legs of Chilopoda (Myriapoda): a review on their morphological disparity and functional variability
2017-11-14
[6]
논문
Comparative morphology of ultimate and walking legs in the centipede Lithobius forficatus (Myriapoda) with functional implications
2019
[7]
논문
On the function of the ultimate legs of some Scolopendridae (Chilopoda, Scolopendromorpha)
2015-06-30
[8]
논문
Centipedes and millipedes with emphasis on North American fauna
http://www.emporia.e[...]
2013-10-14
[9]
서적
Millipedes: Keys and Notes for the Identification of the Species
https://books.google[...]
Brill Archive
[10]
웹사이트
"'Scolopendra subspinipes' (Leach)"
http://www.extento.h[...]
EXTension ENTOmology & UH-CTAHR Integrated Pest Management Program: Knowledge Master
1992-12
[11]
서적
Myriapod biology
Academic Press
[12]
논문
Trunk segment numbers and sequential segmentation in myriapods.
https://onlinelibrar[...]
2005-12
[13]
간행물
Chilopoda – Taxonomic overview
https://brill.com/di[...]
Brill
2024-02-29
[14]
논문
Evolutionary biology of centipedes (Myriapoda: Chilopoda)
https://www.research[...]
[15]
논문
The interaction between developmental bias and natural selection from centipede segmentation to a general hypothesis
[16]
논문
The centipede ''Strigamia maritima'': what it can tell us about development and evolution of segmentation
[17]
서적
Contemporary Debates in Philosophy of Biology
John Wiley and Sons
[18]
논문
Exploring the myriapod body plan: expression patterns of the ten Hox genes in a centipede
https://dev.biologis[...]
2002
[19]
서적
How the Snake Lost its Legs. Curious Tales from the Frontier of Evo-Devo
Cambridge University Press
2014
[20]
논문
Trophic niches, diversity and community composition of invertebrate top predators (Chilopoda) as affected by conversion of tropical lowland rainforest in Sumatra (Indonesia)
2017-08-01
[21]
논문
Arthropod Predation of Vertebrates Structures Trophic Dynamics in Island Ecosystems
2021-08-03
[22]
논문
Predatory behavior of three centipede species of the order Scolopendromorpha (Arthropoda: Myriapoda: Chilopoda)
2016-11-26
[23]
논문
Predation by giant centipedes, ''Scolopendra gigantea'', on three species of bats in a Venezuelan cave
http://www.caribjsci[...]
[24]
뉴스
Giant swimming, venomous centipede discovered by accident in world-first
https://www.theguard[...]
2016-07-01
[25]
웹사이트
A new amphibious species of the genus Scolopendra Linnaeus, 1758 (Scolopendromorpha, Scolopendridae) from the Ryukyu Archipelago and Taiwan
https://www.biotaxa.[...]
2021-04-12
[26]
서적
The Ants
https://archive.org/[...]
Harvard University Press
[27]
논문
Morphology in Chilopoda – a survey
http://soil-organism[...]
2009-12-15
[28]
서적
Secret weapons: defenses of insects, spiders, scorpions and other many-legged creatures
https://archive.org/[...]
The Belknap Press
[29]
논문
Hydrogen cyanide-producing glands in a scolopender, Asanada n.sp. (Chilopoda, scolopendridae)
https://doi.org/10.1[...]
1979-11-01
[30]
논문
Benzoyl cyanide and mandelonitrile in the cyanogenetic secretion of a centipede
[31]
웹사이트
Centipedes & Millipedes
https://www.desertmu[...]
Arizona-Sonora Desert Museum
2023-05-02
[32]
웹사이트
Insects, Spiders, Centipedes, Millipedes
https://www.nps.gov/[...]
2017-10-17
[33]
서적
Invertebrate Medicine
https://www.academia[...]
Blackwell Publishing
[34]
논문
Proteinuria associated with centipede bite
2004-12-15
[35]
웹사이트
Animal Fact Sheet: Desert Centipede
https://www.desertmu[...]
Arizona-Sonora Desert Museum
2023-05-02
[36]
웹사이트
Cave centipede from hell is the deepest-dwelling ever discovered
https://www.nhm.ac.u[...]
Natural History Museum
2015-07-01
[37]
논문
The life history and ecology of the littoral centipede ''Strigamia maritima'' (Leach)
[38]
웹사이트
IUCN 2024
https://www.iucnredl[...]
2024-07-28
[39]
간행물
Scolopendra abnormis
https://www.iucnredl[...]
P. Pearce-Kelly
1996
[40]
간행물
Nothogeophilus turki
https://www.iucnredl[...]
Macadam, C.
2022
[41]
간행물
Seychellonema gerlachi
https://www.iucnredl[...]
Gerlach, J.
2014
[42]
논문
Early life on land
[43]
논문
Land Animals in the Silurian: Arachnids and Myriapods from Shropshire, England
https://horseshoecra[...]
1990-11-02
[44]
논문
Centiped legs (Arthropoda, Chilopoda, Scutigeromorpha) from the Silurian and Devonian of Britain and the Devonian of North America
http://hdl.handle.ne[...]
1998
[45]
논문
Devonobiomorpha, a new order of centipeds (Chilopoda) from the Middle Devonian of Gilboa, New York State, USA, and the phylogeny of centiped orders
https://www.biodiver[...]
1988
[46]
논문
The presumed oldest flying insect: more likely a myriapod?
2017-05-30
[47]
서적
Proceedings of the 7th International Congress of Myriapodology
Brill Publishers
[48]
논문
A new scolopendromorph centipede (Myriapoda: Chilopoda) from the Lower Cretaceous (Aptian) of Brazil
https://onlinelibrar[...]
[49]
논문
First Mesozoic Scutigeromorph Centipede, from the Lower Cretaceous of Brazil
Wiley
[50]
논문
Pancrustacean Evolution Illuminated by Taxon-Rich Genomic-Scale Data Sets with an Expanded Remipede Sampling
2019-07-04
[51]
논문
The Phylogeny and Evolutionary History of Arthropods
[52]
서적
Biodiversidad, Taxonomía y Biogeografia de Artrópodos de México: Hacia una Síntesis de su Conocimiento, Volumen III
Universidad Nacional Autónoma de México
[53]
학위논문
Red List and DNA barcoding of Carinthian and Styrian centipedes (Chilopoda)
https://unipub.uni-g[...]
2019
[54]
논문
Centipedes and Millipedes with Emphasis on North America Fauna
https://sites.google[...]
Emporia State University
1999-03
[55]
논문
Phylogenetic analyses suggest centipede venom arsenals were repeatedly stocked by horizontal gene transfer
[56]
웹사이트
Eating Bugs/Insects in Donghuamen Night Market!
http://www.followmef[...]
2022-11-14
[57]
웹사이트
101 Strangest Foods Around the World
http://whenonearth.n[...]
2015-06-28
[58]
뉴스
Centipede vodka and fried crickets: Is this the future of food?
https://edition.cnn.[...]
2015-07-27
[59]
논문
Centipede envenomation
http://www.wemjourna[...]
[60]
서적
House Centipede
http://ento.psu.edu/[...]
Pennsylvania State University
2009-10-19
[61]
서적
The Life of Shabkar: The Autobiography of a Tibetan Yogin
SUNY Press
1994
[62]
문서
"* 左上:[[オオゲジ]]([[ゲジ目]][[ゲジ科]])* 右上:{{Snamei||Lithobius melanops}}([[イシムカデ目]][[イシムカデ科]])* 左下:{{Snamei||Scolopendra cataracta}}([[オオムカデ目]][[オオムカデ科]])* 右下:{{Snamei||Geophilus}} sp.([[ジムカデ目]][[ジムカデ科]])"
[63]
웹사이트
Chilopoda ムカデ/唇脚綱 - Biological Information System for Marine Life
http://www.godac.jam[...]
2019-02-10
[64]
서적
18 Chilopoda – Fossil history
https://www.research[...]
Brill
2011-01-01
[65]
웹사이트
Chilopoda - Memidex dictionary/thesaurus
http://www.memidex.c[...]
2019-02-10
[66]
논문
A Double Segment Periodicity Underlies Segment Generation in Centipede Development
https://www.cell.com[...]
2004-07-27
[67]
웹사이트
"気になる害虫の知識 {{!}} ムカデ・ヤスデ・ゲジ・ワラジムシ対策 {{!}} 環境機器株式会社"
http://www.semco.net[...]
2019-02-08
[68]
서적
The Biology of Centipedes
https://www.cambridg[...]
Cambridge University Press
1981
[69]
서적
Arthropod Segmentation and Tagmosis
https://doi.org/10.1[...]
Springer
2013
[70]
논문
Neotype designation and a diagnostic account for the centipede, ''Scolopendra gigantea'' L. 1758, with an account of S. galapagoensis Bollman 1889 (Chilopoda Scolopendromorpha Scolopendridae)
https://doi.org/10.1[...]
2000-05-01
[71]
학술지
Evolutionary Biology of Centipedes (Myriapoda: Chilopoda)
https://www.annualre[...]
2006-12-12
[72]
학술지
Exploring the myriapod body plan: expression patterns of the ten Hox genes in a centipede
http://dev.biologist[...]
2002-03-01
[73]
학술지
Variation and specialisation of the forcipular apparatus of centipedes (Arthropoda: Chilopoda): A comparative morphometric and microscopic investigation of an evolutionary novelty
https://www.scienced[...]
2012-05-01
[74]
문서
ゲジ類の複眼は側単眼から二次的に形成したものと解釈され、「偽複眼」(pseudofacetted eye)とも呼ばれてきたが、21世紀以降ではこの解釈は否定的で、節足動物の祖先形質としての真の複眼であることが確定的になっている。詳細はゲジ#眼を参照。
[75]
학술지
The compound eye of ''Scutigera coleoptrata'' (Linnaeus, 1758) (Chilopoda: Notostigmophora): an ultrastructural reinvestigation that adds support to the Mandibulata concept
https://doi.org/10.1[...]
2003-11-01
[76]
학술지
Mechanisms of eye development and evolution of the arthropod visual system: The lateral eyes of myriapoda are not modified insect ommatidia
http://www.sciencedi[...]
2007-04-12
[77]
학술지
Evolution of blindness in scolopendromorph centipedes (Chilopoda: Scolopendromorpha): insight from an expanded sampling of molecular data
https://onlinelibrar[...]
2012
[78]
문서
解剖学的には5節の肢節のみ含め、そのうち基部の短い柄部は癒合した2節、残りの数多くの環状節に分れた細長い部分は3節由来で、途中2つの関節によりそれぞれの真の境目を示される。
[79]
문서
イシムカデ類:15-111、ナガズイシムカデ類:17-18、オオムカデ類:17-34、ジムカデ類:14
[80]
학술지
A common terminology for the external anatomy of centipedes (Chilopoda)
https://zookeys.pens[...]
2010-11-18
[81]
문서
「maxilliped」は通常では甲殻類の顎脚を専門に指す用語である。
[82]
문서
ただしムカデの毒は正確には「venom」(注入経由で侵入する生物毒)であり、「poison」(接触経由で侵入する生物毒)ではない。
[83]
간행물
Evolution, Morphology and Development of the Centipede Venom System
https://www.research[...]
Springer Netherlands
2022-08-19
[84]
학술지
Detecting taxonomic signal in an under-utilised character system: geometric morphometrics of the forcipular coxae of Scutigeromorpha (Chilopoda)
https://zookeys.pens[...]
2011-12-20
[85]
문서
顎肢は脚と同じ肢節名を用いられるが、両者の肢節相同性は未解明である。順に femuroid・脛節 tibia・跗節 tarsus・apical claw とする体系もある。
[86]
학술지
The evolution of centipede venom claws – Open questions and possible answers
http://www.sciencedi[...]
2014-01-01
[87]
문서
イシムカデ類における Paitobius zinus の雄もこれに類する形の顎肢をもつ。
[88]
학술지
Centipede Venom: Recent Discoveries and Current State of Knowledge
https://www.research[...]
2015-02-25
[89]
문서
An: 肛門、Cx, Cxpd: 基節、Dct: 精管、Gp: 生殖肢、gS: 生殖体節の腹板、gSeg: 生殖節、gT: 生殖節の背板、Pen: 陰茎、Scx, 側板/亜基節、Stn: 有脚胴節の腹板、T: 有脚胴節の背板、Tel: 尾節、Tr: 転節、Vul: 産卵口
[90]
학술지
CLASS CHILOPODA: EVOLUTION AND ENVIRONMENT ADAPTATION
https://www.semantic[...]
2010
[91]
문서
ナガズイシムカデ類の2枚ずつ第1・3・5・7・8・10・12・14有脚胴節の背板は元々該当胴節の1背板由来とされており、これを2枚ずつ1背板と扱えば、残りの脚をもつ胴節の背板がそれより短いと言える。ゲジ類の見かけ上前7枚の背板を順に第1・3・5・7+8・10・12・14有脚胴節のみに由来と考える場合も同様である。
[92]
학술지
A century later – a total evidence re-evaluation of the phylogeny of scutigeromorph centipedes (Myriapoda : Chilopoda)
https://www.research[...]
2006-10-12
[93]
문서
転節を「第1転節」、前腿節を「第2転節」とする体系もある。
[94]
서적
VII. The Chilopoda
https://www.degruyte[...]
Cornell University Press
1952
[95]
학술지
The geological record and phylogeny of the Myriapoda
https://www.research[...]
2010-03-01
[96]
문서
A. Matsui; K. Yahata. (2012). [http://aesj.co-site.jp/vol47/2012_Vol.47_11.pdf Comparative study of autotomic structures in centipedes Arthropoda Chilopoda]. Proc. Arthropod. Embryol. Soc. Jpn., Vol.47, pp. 11-19.
[97]
학술지
Variability in trunk segmentation in the centipede order Scolopendromorpha: a remarkable new species of Scolopendropsis Brandt (Chilopoda: Scolopendridae) from Brazil
https://www.research[...]
2008-12-31
[98]
학술지
Saltational evolution of trunk segment number in centipedes
https://www.research[...]
2009
[99]
학술지
Discovery of a second geophilomorph species (Myriapoda: Chilopoda) having twenty-seven leg-bearing segments, the lowest number recorded up to the present in the centipede order Geophilomorpha
http://www.scielo.br[...]
2013
[100]
학술지
Developmental abnormalities in ''Himantarium gabrielis'' (LINNAEUS, 1767) (Chilopoda: Geophilomorpha: Himantariidae)
https://www.research[...]
2007-08
[101]
서적
Treatise on Zoology - Anatomy, Taxonomy, Biology. The Myriapoda
https://books.google[...]
BRILL
2011-03-21
[102]
학술지
The ultimate legs of Chilopoda (Myriapoda): a review on their morphological disparity and functional variability
https://peerj.com/ar[...]
2017-11-14
[103]
학술지
Interaction of the tracheal tubules of ''Scutigera coleoptrata'' (Chilopoda, Notostigmophora) with glandular structures of the pericardial septum
https://zookeys.pens[...]
2015-06-30
[104]
문서
'* 스リット状:イシムカデ類 * クレーター状:ナガズイシムカデ類 * 耳状:オオムカデ類アオムカデ亜科 * 三角形:オオムカデ類オオムカデ亜科 * 楕円形:他のオオムカデ類・ジムカデ類
[105]
문서
An: 肛門、Cx, Cxpd: 基節、Dct: 精管、Gp: 生殖肢、gS: 生殖体節の腹板、gSeg: 生殖節、gT: 生殖節の背板、Pen: 陰茎、Scx, 側板/亜基節、Stn: 有脚胴節の腹板、T: 有脚胴節の背板、Tel: 尾節、Tr: 転節、Vul: 産卵口
[106]
서적
Morphology of the Insect Abdomen: The male genitalia (including arthropods other than insects)
https://archive.org/[...]
Smithsonian Institution
1936
[107]
서적
Desert Arthropods: Life History Variations
https://books.google[...]
Springer Science & Business Media
1999-12-14
[108]
논문
The embryoid development of ''Strigamia maritima'' and its bearing on post-embryonic segmentation of geophilomorph centipedes
https://doi.org/10.1[...]
2014-08-13
[109]
논문
When SEM becomes a deceptive tool of analysis: the unexpected discovery of epidermal glands with stalked ducts on the ultimate legs of geophilomorph centipedes
https://doi.org/10.1[...]
2021-04-20
[110]
웹사이트
Order Geophilomorpha
https://keys.lucidce[...]
2019-02-06
[111]
논문
On the function of the ultimate legs of some Scolopendridae (Chilopoda, Scolopendromorpha)
https://zookeys.pens[...]
2015-06-30
[112]
논문
Comparative morphology of ultimate and walking legs in the centipede ''Lithobius forficatus'' (Myriapoda) with functional implications
https://doi.org/10.1[...]
2019-01-14
[113]
웹사이트
Order Scolopendromorpha
https://keys.lucidce[...]
2019-02-09
[114]
서적
The Biology of Centipedes
https://books.google[...]
Cambridge University Press
2006
[115]
서적
Multicellular Animals: Volume II: The Phylogenetic System of the Metazoa
https://books.google[...]
Springer Science & Business Media
2013-03-09
[116]
웹인용
Order Lithobiomorpha
https://keys.lucidce[...]
2022-08-23
[117]
웹사이트
Order Craterostigmomorpha
https://keys.lucidce[...]
2019-02-10
[118]
웹인용
Predation by Giant Centipedes , ''Scolopendra gigantea'' , on Three Species of Bats in a Venezuelan Cave
https://www.research[...]
2005
[119]
논문
A taxonomic review of the centipede genus ''Scolopendra'' Linnaeus, 1758 (Scolopendromorpha, Scolopendridae) in mainland Southeast Asia, with description of a new species from Laos
https://zookeys.pens[...]
2016-05-17
[120]
논문
Arthropod Predation of Vertebrates Structures Trophic Dynamics in Island Ecosystems
https://www.journals[...]
2021-10-01
[121]
서적
The Ants
https://books.google[...]
Harvard University Press
1990
[122]
논문
First Molecular Data and the Phylogenetic Position of the Millipede-Like Centipede ''Edentistoma octosulcatum'' Tömösváry, 1882 (Chilopoda: Scolopendromorpha: Scolopendridae)
https://www.research[...]
2014-11-12
[123]
논문
The sternal pore areas of geophilomorph centipedes (Chilopoda: Geophilomorpha)
https://academic.oup[...]
1995-10
[124]
논문
Chemistry of the sternal gland secretion of the Mediterranean centipede Himantarium gabrielis (Linnaeus, 1767) (Chilopoda: Geophilomorpha: Himantariidae)
https://pubmed.ncbi.[...]
2013-09
[125]
서적
Parental care in centipedes (Myriapoda: Chilopoda): A phylogenetic perspective
https://www.research[...]
Conference: 16th International Congress of Myriapodology At: Olomouc
2014-07
[126]
논문
Brooding behaviour of the centipede ''Otostigmus spinosus'' Porat, 1876 (Chilopoda: Scolopendromorpha: Scolopendridae) and its morphological variability in Thailand
https://www.research[...]
2014-05-30
[127]
논문
Arthropod relationships revealed by phylogenomic analysis of nuclear protein-coding sequences
https://www.nature.c[...]
2010-02
[128]
논문
Four myriapod relatives – but who are sisters? No end to debates on relationships among the four major myriapod subgroups
https://doi.org/10.1[...]
2020-11-04
[129]
논문
Devonobiomorpha, a new order of centipeds (Chilopoda) from the Middle Devonian of Gilboa, New York State, USA, and the phylogeny of centiped orders. American Museum novitates ; no. 2927
https://hdl.handle.n[...]
1988
[130]
논문
Filogenia de chilipoda: combinando secuencias de los genes ribosómicos 18S y 28S y morfología
https://www.semantic[...]
1999
[131]
논문
Further use of nearly complete 28S and 18S rRNA genes to classify Ecdysozoa: 37 more arthropods and a kinorhynch
https://www.scienced[...]
2006-09-01
[132]
논문
Including secondary structure, fossils and molecular dating in the centipede tree of life
https://www.scienced[...]
2010-10-01
[133]
논문
Exploring Phylogenetic Relationships within Myriapoda and the Effects of Matrix Composition and Occupancy on Phylogenomic Reconstruction
https://academic.oup[...]
2016-09-01
[134]
논문
The tracheal system of scutigeromorph centipedes and the evolution of respiratory systems of myriapods
https://www.scienced[...]
2021-01-01
[135]
논문
Evaluating Topological Conflict in Centipede Phylogeny Using Transcriptomic Data Sets
https://academic.oup[...]
2014-06-01
[136]
논문
Centipedal Hemocyanin: Its Structure and Its Implications for Arthropod Phylogeny
https://www.jstor.or[...]
1985
[137]
웹사이트
Order Lithobiomorpha - Stone Centipedes - BugGuide.Net
https://bugguide.net[...]
2019-02-07
[138]
논문
First records of ''Lithobius'' (''Lithobius'') ''forficatus'' (Linnaeus 1758) (Chilopoda: Lithobiomorpha: Lithobiidae) from mainland Hokkaido, Japan
https://www.jstage.j[...]
2021
[139]
서적
Treatise on Zoology - Anatomy, Taxonomy, Biology. The Myriapoda
https://books.google[...]
BRILL
2011-03-21
[140]
웹사이트
帯広畜産大学、日本本土新記録のムカデを帯広で発見
https://univ-journal[...]
2022-08-16
[141]
문서
'1:前方背面、2:[[顎肢]]と基胸板腹面、3:前端側面、後端側面、後端背面、6:後端腹面、後端側面、8:頭部腹面、9:[[上唇 (節足動物)|上唇]]、10-12:[[大顎]]、13:第1[[小顎]]、14:第2小顎、15:[[気門]]と[[側板]]、16-17:第14[[歩肢]]、18-19:[[曳航肢]]'
[142]
논문
A New Zealand species of the trans-Tasman centipede order Craterostigmomorpha (Arthropoda : Chilopoda) corroborated by molecular evidence
https://www.research[...]
2008-03-18
[143]
논문
A new and annectant type of chilopod (Craterostigmus)
https://www.biodiver[...]
1902
[144]
논문
HABITS OF LIFE AND EVOLUTION OF BODY DESIGN IN ARTHROPODA
https://doi.org/10.1[...]
1958-07
[145]
논문
The evolution of arthropodan locomotory mechanisms. Part 8. Functional requirements and body design in Chilopoda, together with a comparative account of their skeleto-muscular systems and an Appendix on a comparison between burrowing forces of annelids an
https://doi.org/10.1[...]
1965-10
[146]
웹사이트
Tropical Centipedes (Order Scolopendromorpha)
https://www.inatural[...]
2019-02-07
[147]
웹사이트
Order Scolopendromorpha - Bark Centipedes - BugGuide.Net
https://bugguide.net[...]
2019-02-09
[148]
논문
An overview of the extant genera and subgenera of the order Scolopendromorpha (Chilopoda): a new identification key and updated diagnoses
https://www.biotaxa.[...]
2020-08-10
[149]
웹사이트
Geophilidae ジムカデ科 - Biological Information System for Marine Life
http://www.godac.jam[...]
2019-02-06
[150]
문서
危険な生物
https://www.rinya.ma[...]
東北森林管理局
[151]
문서
保健管理センター
http://www1.ous.ac.j[...]
岡山理科大学
[152]
서적
Kaufman Field Guide to Insects of North America
https://books.google[...]
Houghton Mifflin Harcourt
2007
[153]
웹사이트
'House centipede {{!}} Arthropod Museum'
https://arthropod.ua[...]
2022-08-19
[154]
논문
Centipede envenomation
http://www.wemjourna[...]
[155]
문서
Aboriginal names and utilization of the fauna in the Eyrean region
Transactions of the Royal Society of South Australia
1943
[156]
웹사이트
宮城・角田のムカデ絵馬:時事ドットコム
https://www.jiji.com[...]
2021-09-30
[157]
문서
赤城山と俵藤太
https://web.archive.[...]
藤原秀郷
[158]
웹사이트
川越の文化財建造物「百足屋」を再生 日本文化を体験する場にも
https://kawagoe.keiz[...]
2021-09-30
[159]
웹사이트
立皇嗣の礼を前に秋篠宮さまが小6の時見た絵「再発見」:朝日新聞デジタル
https://www.asahi.co[...]
2021-09-30
[160]
논문
First venomous crustacean discovered
http://www.nature.co[...]
[161]
논문
How many Arachnida and Myriapoda are there worldwide and in Amazonia?
[162]
웹인용
Chilopoda Latreille 1817
http://www.insecta.b[...]
''
2012-06-24
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com