창고기목
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
창고기목은 해저 모래 속에 서식하며 여과 섭식을 하는 자웅이체 동물이다. 척추동물과 유사한 해부학적 특징을 가지며, 척추동물의 조상 모델로 연구된다. 전 세계적으로 다양한 종이 존재하며, 일부 지역에서는 식용으로 사용되기도 한다.
더 읽어볼만한 페이지
2. 생태
창고기목 동물은 주로 특정 조건을 갖춘 해저의 기질에 서식하며 생활한다. 이들은 몸의 일부를 밖으로 내놓은 채 물속의 미세한 플랑크톤 등을 걸러 먹는 독특한 섭식 방식을 가지고 있다. 번식은 암수가 구분되는 자웅이체이며, 몸 밖에서 수정이 이루어지는 외부 수정을 통해 이루어진다. 종에 따라 차이가 있지만, 주로 특정 계절에 산란하는 경향을 보인다.
2. 1. 서식지
성체 창고기목은 주로 해저에 서식하며, 부드럽고 유기물 함량이 적으며 통풍이 잘 되는 기질에 굴을 파고 산다. 다양한 종이 가는 모래, 굵은 모래, 조개껍질 퇴적물 등 여러 종류의 기질에서 발견되지만, 대부분은 미세 입자 함량이 낮은 굵은 모래를 뚜렷하게 선호한다.예를 들어, 아프리카 서해안의 ''Branchiostoma nigeriense'', 미시시피강 하구(Mississippi Sound)와 사우스캐롤라이나에서 조지아에 이르는 해안의 ''Branchiostoma caribaeum'', 북서 아프리카 연안 대륙붕 지역의 ''B. senegalense'', 프랑스 남부 지중해 연안의 ''B. lanceolatum'' 등이 이러한 선호도를 보인다.
그러나 플로리다주 탬파 만에 서식하는 ''Branchiostoma floridae''는 가는 모래 바닥을 선호하는 경향을 보여 예외적인 경우로 여겨진다.
2. 2. 먹이 섭취
창고기목의 서식지 선호도는 먹이를 섭취하는 방식과 관련이 있다. 이들은 몸의 앞부분만 물 밖으로 내놓고 플랑크톤을 걸러 먹는다. 이는 아가미에 있는 섬모가 점액질 막을 통해 물을 통과시키면서 먹이를 걸러내는 방식으로 이루어진다. ''Branchiostoma floridae''는 미생물부터 작은 식물성 플랑크톤 크기까지 다양한 입자를 포획할 수 있으며,[16] ''B. lanceolatum''은 상대적으로 더 큰 입자(>4 μm)를 우선적으로 포획하는 경향이 있다.[17]2. 3. 번식
창고기류는 암수가 다른 자웅이체 동물이며, 몸 밖에서 수정이 이루어지는 외부 수정을 통해 번식한다. 종에 따라 약간의 차이는 있지만, 일반적으로 봄과 여름에 해당하는 산란기에만 번식한다.[18] 모든 창고기류 종은 해가 진 직후에 알을 낳으며, 산란기 동안 약 2주마다 한 번씩 동시에 산란하는 종(예: ''Branchiostoma floridae'')과 점진적으로 나누어 산란하는 종(예: ''Branchiostoma lanceolatum'')이 있다.[19][20]니콜라스 홀랜드(Nicholas Holland)와 린다 홀랜드(Linda Holland)는 사육 환경에서 인위적으로 산란을 유도하고 체외 수정을 통해 창고기류 배아를 얻는 방법을 처음으로 기술한 연구자들이다.[21] 실험실에서는 전기 충격이나 열 충격을 이용하여 인공적으로 산란을 유도할 수 있다.[22]
3. 해부학적 구조
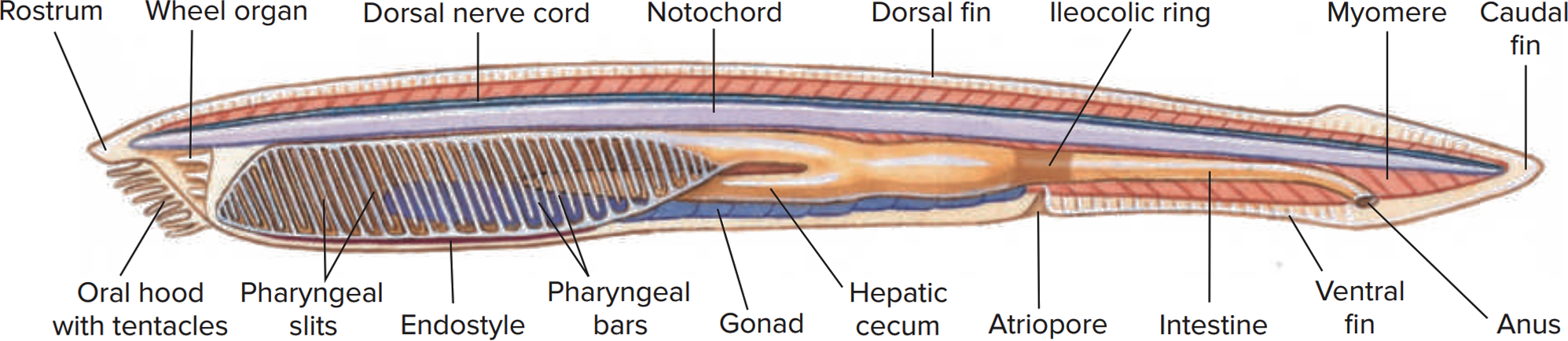
유생 단계에서는 몸의 구조가 매우 비대칭적이다. 입과 항문은 몸의 왼쪽에 위치하고, 아가미 구멍은 오른쪽에 있다.[41][42] 인두와 관련된 기관들도 몸의 왼쪽 또는 오른쪽에만 자리 잡고 있으며, 분절된 근육 블록과 신경계 일부 역시 비대칭적인 구조를 보인다.[43]
성체로 변태하면서 해부학적 구조는 점차 대칭적으로 변하지만, 성체에서도 신경계와 생식선의 위치 등 일부 비대칭적인 특징이 남아있다. 생식선의 경우, 아시메트론속(Asymmetron)과 에피고니크티스속(Epigonichthys) 속에서는 오른쪽에만 위치하지만, 창고기속(Branchiostoma) 속에서는 몸 양쪽에 모두 발달한다.[44][45]
창고기의 최대 길이는 종에 따라 다르지만, 일반적으로 2.5cm에서 8cm 사이이다.[54][46] 벨체르창고기(Branchiostoma belcheri)와 창고기(B. lanceolatum)가 가장 큰 종에 속한다.[54] 크기를 제외하면 대부분의 종은 외형이 매우 비슷하며, 주로 근육분절의 수나 유생 시기 몸의 색소 침착 정도에서 차이를 보인다.[54]
몸은 반투명하며 물고기와 비슷하게 생겼지만, 짝을 이루는 지느러미나 다른 팔다리는 없다. 꼬리지느러미는 비교적 덜 발달하여 수영 능력이 뛰어나지는 않다. 아가미 구멍, 입, 꼬리 부분에는 연골 물질이 있어 형태를 유지하지만, 복잡한 골격 구조는 없다.[47]
3. 1. 신경계와 척삭
19세기 중반부터 창고기목의 해부학적 관찰이 시작되었으며, 처음에는 성체, 이후에는 배아의 해부학이 기술되었다.[34]러시아의 동물학자 알렉산더 코발레프스키는 성체 창고기목의 주요 해부학적 특징인 속이 빈 등쪽 신경관, 내주피, 분절된 몸, 항문 뒤 꼬리 등을 처음으로 기술했다.[34] 이는 척삭동물의 중요한 특징들을 밝힌 연구였다. 프랑스의 드 콰트르파주는 창고기목의 신경계를 최초로 완벽하게 기술했다.[35] 독일의 하인리히 라트케[36]와 스코틀랜드의 존 굿시어[37] 또한 창고기목 성체의 해부학 연구에 중요한 기여를 했다.
코발레프스키는 창고기목 배아의 첫 번째 완전한 묘사를 발표했으며,[34] 슐츠와 로이카르트는 유충을 처음으로 묘사했다.[38] 이후 오스트리아의 하체크, 미국의 콘클린[39], 그리고 중국의 퉁(실험 발생학) 등 여러 학자들이 창고기목 배아 해부학 연구에 중요한 기여를 했다.[40]
3. 2. 시각 시스템
창고기목은 네 종류의 빛 감지 구조를 가지고 있는 것으로 알려져 있다. 이는 요셉 세포, 헤세 기관, 짝을 이루지 않는 전방 눈, 그리고 판상체이며, 이 구조들은 모두 빛 수용체로 옵신을 이용한다. 이러한 모든 기관과 구조는 신경관 내부에 위치하며, 가장 앞쪽에 전방 눈이 있고 그 뒤로 판상체, 요셉 세포, 헤세 기관 순서로 배열되어 있다.3. 3. 형광 단백질
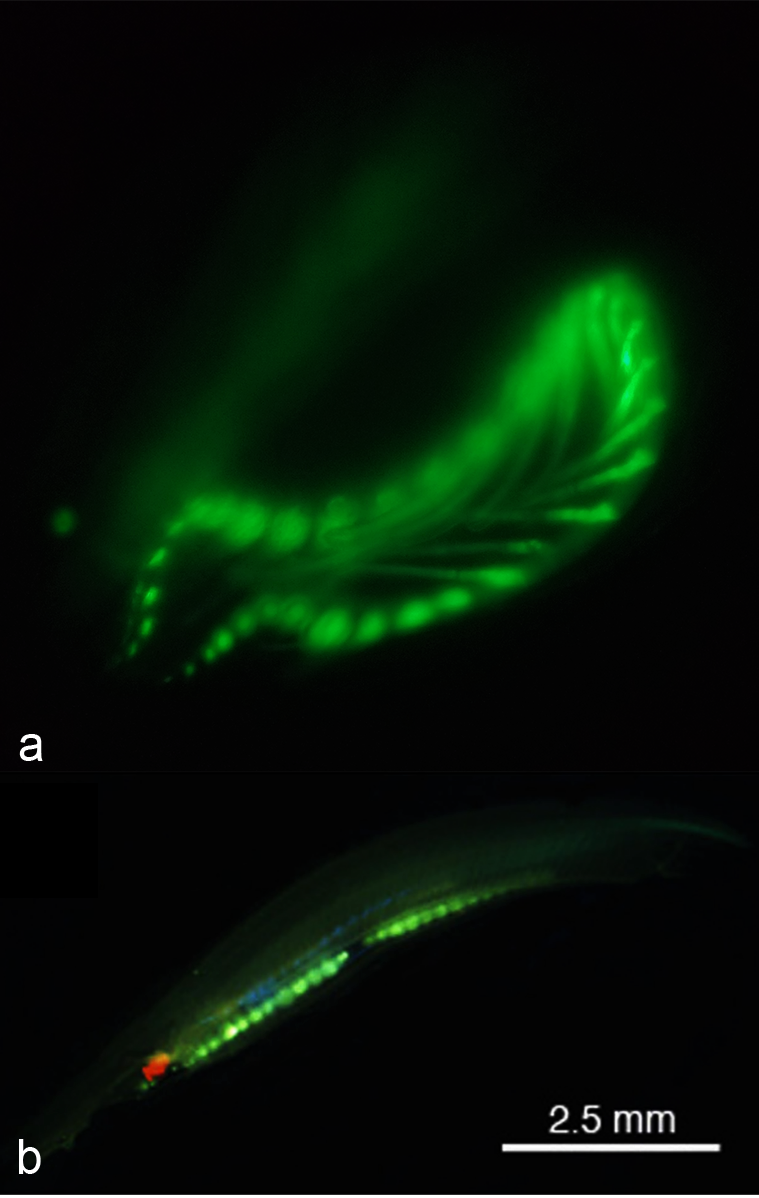
창고기목은 구강 촉수 내부와 눈점 근처에서 자연적으로 녹색 형광 단백질 (GFP)을 발현한다.[62] 종에 따라 꼬리와 생식선에서도 발현될 수 있지만, 이는 ''Asymmetron'' 속에서만 보고되었다.[63] 전 세계의 창고기목 종에서 여러 형광 단백질 유전자가 기록되었으며, ''Branchiostoma floridae'' 종만 해도 16개의 GFP-코딩 유전자를 가지고 있다. 그러나 창고기목에서 생성된 GFP는 해파리 (''Aequorea victoria'')의 것보다 요각류에서 생성된 GFP와 더 유사하다.
GFP는 창고기목에서 플랑크톤을 입으로 유인하는 등 여러 역할을 하는 것으로 추정된다. 창고기목은 여과 섭식 동물이므로 자연적인 해류가 근처의 플랑크톤을 소화관으로 끌어들이는데, 이때 GFP가 플랑크톤 유인에 관여할 수 있다. 또한 GFP는 유충 단계에서도 발현되는데, 이는 높은 에너지의 청색광을 덜 해로운 녹색광으로 변환하여 광보호 기능을 수행할 수 있음을 시사한다.
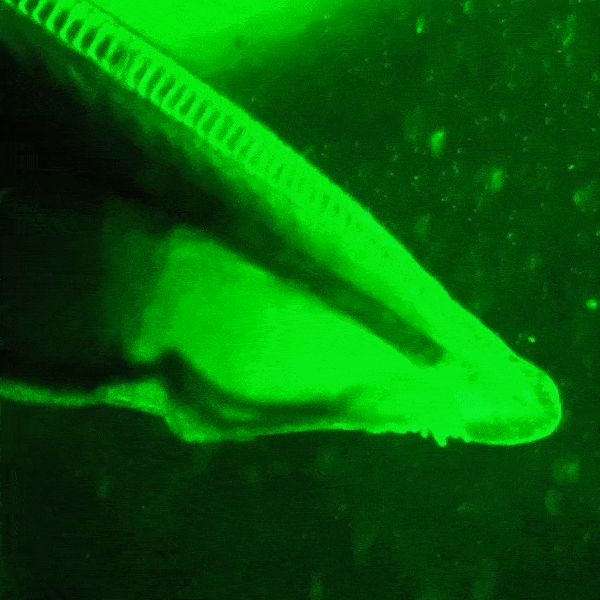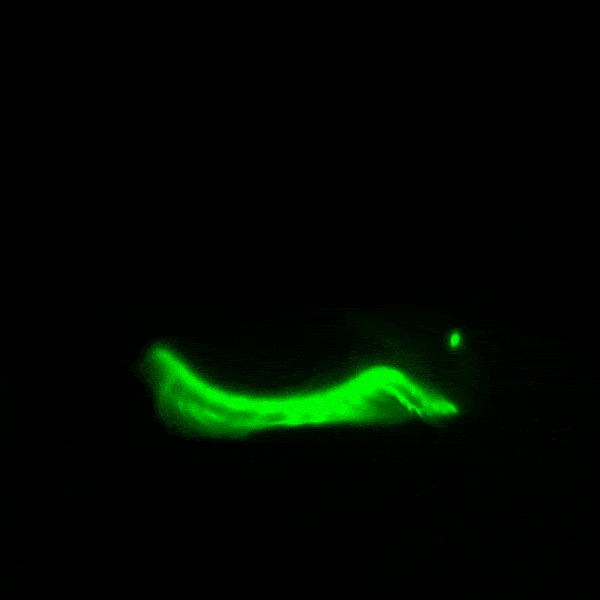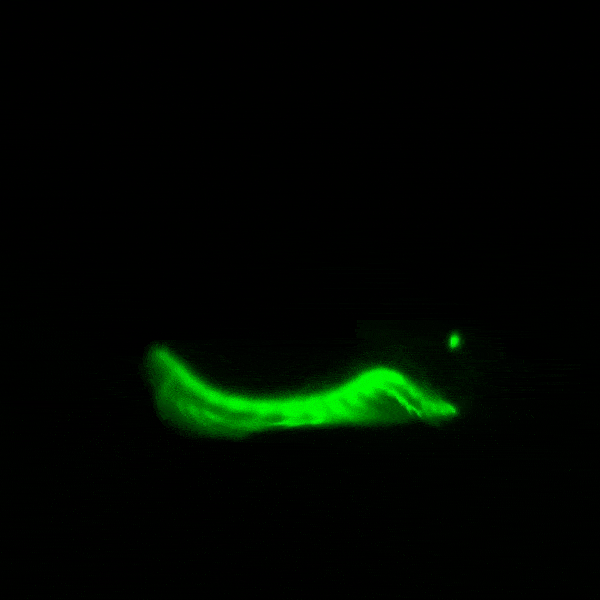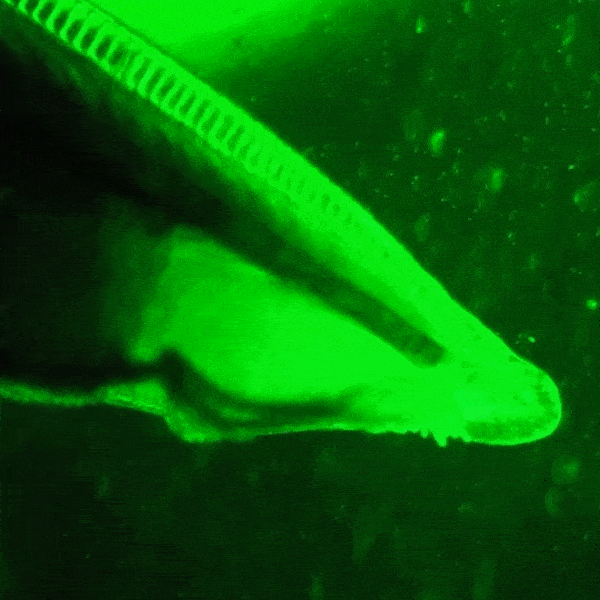
창고기목의 형광 단백질은 분자생물학 및 현미경 검사에 활용하기 위해 개량되었다. ''Branchiostoma lanceolatum''에서 유래한 [https://www.ncbi.nlm.nih.gov/nuccore/EU482389 노란색 형광 단백질]은 매우 높은 양자 수율 (~0.95)을 나타낸다.[64] 이 단백질은 [https://www.fpbase.org/protein/mneongreen/ mNeonGreen]이라는 이름의 단량체 녹색 형광 단백질로 유전자 조작되었으며, 현재까지 알려진 단량체 녹색 또는 노란색 형광 단백질 중 가장 밝은 것으로 알려져 있다.
3. 4. 소화계
창고기목은 몸의 앞부분만 물에 노출시킨 채 플랑크톤을 걸러 먹는 방식으로 먹이를 섭취하며, 이는 서식지 선호도와 관련이 있다. 이 과정은 점액질 막을 통해 물을 통과시키는 아가미 섬모에 의해 이루어진다. ''Branchiostoma floridae''는 미생물부터 작은 식물성 플랑크톤 크기의 입자까지 포획할 수 있으며,[16] ''B. lanceolatum''은 상대적으로 더 큰 입자(>4 μm)를 우선적으로 포획한다.[17]창고기목은 수동적인 여과 섭식 생물로,[65] 대부분의 시간을 모래 속에 반쯤 몸을 묻고 앞부분만 내놓은 채 보낸다.[66] 이들은 박테리아, 균류, 규조류, 동물성 플랑크톤과 같은 다양한 작은 플랑크톤 생물을 먹이로 삼으며, 유기물 부스러기도 섭취한다.[67] 야생 상태의 창고기목 유생이 무엇을 먹는지는 잘 알려져 있지 않지만, 사육 환경에서는 여러 종의 유생이 식물성 플랑크톤을 먹고 생존할 수 있다. 다만, ''Asymmetron lucayanum'' 종에게는 이것이 최적의 먹이 조건이 아닌 것으로 보인다.[67]
입 앞에는 구강 섬모라는 얇은 촉수 같은 구조물이 매달려 있는데, 이는 감각 기능을 수행하고 몸 안으로 들어가는 물을 걸러주는 역할을 한다. 물은 입을 통해 많은 아가미 틈새가 있는 큰 인두로 들어간다. 인두의 배 쪽 표면에는 내주피(endostyle)라는 홈이 있으며, 이는 Hatschek's pit이라는 구조와 연결되어 점액 막을 생성한다. 섬모의 움직임은 이 점액을 아가미 틈새 표면 위로 밀어 올리면서 물에 떠다니는 음식 입자를 붙잡는다. 이 점액은 다시 등 쪽에 있는 상인두 홈(epipharyngeal groove)에 모여 소화관의 나머지 부분으로 전달된다. 아가미 틈새를 통과한 물은 인두를 둘러싼 심방으로 모였다가, 심방구멍을 통해 몸 밖으로 배출된다.[47]
성체와 유생 모두 삼키기 어려운 큰 입자나 입, 목에 걸린 이물질을 제거하기 위해 "기침"과 유사한 반사 행동을 보인다. 유생의 경우 이 반사는 인두 근육에 의해 조절되지만, 성체에서는 심방의 수축을 통해 이루어진다.[68][69]
소화계의 나머지 부분은 인두에서 항문까지 이어지는 단순한 관 형태로 이루어져 있다. 간 맹장(hepatic caecum)은 창자 아래쪽에서 갈라져 나오는 하나의 막힌 주머니(맹장)로, 그 내벽 세포가 음식 입자를 식세포 작용으로 직접 소화할 수 있다는 특징이 있다. 이는 척추동물에서는 볼 수 없는 방식이다. 간 맹장은 간의 여러 기능을 수행하지만, 진정한 간으로 간주되지는 않으며 척추동물 간의 상동 기관으로 여겨진다.[70][71][72]
3. 5. 기타 시스템
Joseph 세포는 미세융모 띠로 둘러싸인 노출된 광수용체이다. 이 세포는 옵신 멜라놉신을 가지고 있다. 헤세 기관(등쪽 눈이라고도 함)은 미세융모 띠로 둘러싸인 광수용체 세포로 구성되어 있으며 멜라놉신을 가지고 있지만, 반은 컵 모양의 색소 세포에 싸여 있다. 두 세포 모두의 최대 감도는 약 470nm[56](파란색)이다.Joseph 세포와 헤세 기관은 모두 신경관에 있으며, Joseph 세포는 등쪽 기둥을 형성하고 헤세 기관은 관의 길이를 따라 복부 부분에 있다. Joseph 세포는 전두 소포(또는 대뇌 소포)의 꼬리쪽 끝에서 근절 3과 4 사이의 경계까지 뻗어 있으며, 헤세 기관은 그곳에서 시작하여 거의 꼬리까지 이어진다.[57][58]
4. 분류
창고기목에서 처음으로 기록된 대표적인 생물은 ''창고기''(''Branchiostoma lanceolatum'')이다. 이 종은 1774년 페터 시몬 팔라스에 의해 연체동물 민달팽이의 일종인 ''Limax'' 속으로 처음 묘사되었다.[23] 이후 1834년, 가브리엘 코스타는 이 생물이 무악류 척추동물(먹장어, 칠성장어 등)과 더 가깝다고 판단하여, 새로운 속인 ''Branchiostoma''(그리스어로 branchio = 아가미, stoma = 입)를 만들어 분류했다.[24][25] 1836년에는 야렐이 이 속의 이름을 ''Amphioxus''(그리스어로 '양쪽이 뾰족함')로 바꾸었으나,[26] 현재 ''Amphioxus''는 ''Branchiostoma'' 속의 동의어로 취급된다. 그럼에도 'amphioxus'라는 이름은 오늘날에도 영어권 등지에서 창고기류를 가리키는 일반명으로 'lancelet'과 함께 널리 사용되고 있다.
창고기류는 Cephalochordata 아문, Leptocardii 강에 속한다. Cephalochordata 아문명은 1866년 에른스트 헤켈이 제안했으며,[1] Leptocardii 강 이름은 1845년 요하네스 페터 뮐러가 아강(subclass) 수준에서 처음 사용했다.[29]
창고기목은 전통적으로 아심메트론과(Asymmetronidae)와 창고기과(Branchiostomidae)의 두 과로 분류되어 왔다.[27] Branchiostomidae라는 과 이름은 1846년 샤를 루시앙 보나파르트가 처음 사용했지만, 당시에는 철자가 "Branchiostomidae"로 잘못 표기되었다.[28] (보다 자세한 하위 분류는 #하위 과 섹션 참조)
창고기류가 속하는 목(order) 수준의 분류명으로는 Amphioxi Bonaparte, 1846,[30] Amphioxiformes Berg, 1937,[31][32] 또는 Branchiostomiformes Fowler, 1947[33] 등이 사용되기도 한다. 또한, 창고기류를 포함하는 더 상위 분류군으로 Acrania Haeckel, 1866[32]이라는 이름이 쓰이기도 한다.
4. 1. 하위 과

창고기는 전통적으로 척추동물의 자매 그룹으로 간주되어 왔으며, 이 두 그룹은 멍게와 같은 피낭동물과는 구분되는 최소 10가지의 형태학적 특징을 공유하는 것으로 여겨졌다.[82] 그러나 최근 광범위한 분자계통학 연구 결과, 피낭동물이 실제로는 척추동물의 더 가까운 자매 그룹이며, 창고기아문이 척삭동물 내에서 가장 먼저 분기한 기저 분류군임이 밝혀졌다.[83][84] 이는 멍게류가 이전에 척추동물과 창고기만이 공유하는 파생 형질로 여겨졌던 일부 특징들을 이차적으로 잃어버렸음을 시사한다. 또한 창고기는 엑손 셔플링 및 도메인 조합과 같은 유전자 변형이 활발하여, 염기서열이 분석된 동물 중 유전적으로 매우 다양한 편에 속한다.[13]
현존하는 창고기목은 하나의 과(Branchiostomatidae) 아래 3개의 속으로 분류된다. 세계 해양 종 등록부는 아래 목록의 종들을 인정하고 있으며, 다른 출처에서는 약 30종을 인정하기도 한다.[74][65][85] 아직 발견되지 않은 은닉종이 더 존재할 가능성도 제기된다.[67]
분자 시계 연구에 따르면, 세 속 중 Asymmetron이 가장 먼저 분기한 것으로 보이며, 그 시점은 연구에 따라 약 1억 년 전[74] 또는 약 4,600만 년 전[65]으로 추정된다. 후자의 추정에 따르면, Branchiostoma와 Epigonichthys는 약 3,830만 년 전에 서로 분기한 것으로 보인다.[65] 이렇게 오래전에 분기했음에도 불구하고, ''Asymmetron lucayanum''과 ''Branchiostoma floridae'' 사이의 잡종은 생존 가능한 것으로 알려져 있다.[67]
다음은 세계 해양 종 등록부 기준에 따른 창고기과 하위 속 및 종 목록이다.[74][65][85]
5. 계통 발생
전통적으로 창고기는 척추동물과 자매 분류군을 이루며, 이 두 그룹(때때로 척삭동물(Notochordata)이라 불림)은 피낭동물(Urochordata)과 자매 그룹 관계에 있다고 여겨졌다. 이 관점에 따르면, 창고기와 척추동물은 최소 10가지의 공통된 형태학적 특징을 공유하지만, 피낭동물은 그렇지 않다.[82] 그러나 최근의 광범위한 분자 계통발생 분석 결과는 이러한 전통적인 관계 설정이 잘못되었음을 강력하게 시사한다. 새로운 연구들은 두미삭동물(창고기 포함)이 척삭동물의 가장 기저 분기군이며, 오히려 피낭동물이 척추동물과 가장 가까운 자매 그룹임을 설득력 있게 제시한다.[83][84] 이러한 수정된 계통 발생 관계는, 과거 척추동물과 창고기만의 공유파생형질로 간주되었던 일부 형태학적 특징들을 피낭동물이 진화 과정에서 이차적으로 잃어버렸을 가능성을 제기한다.
또한 창고기는 엑손 셔플링(exon shuffling) 및 도메인 조합(domain combination)과 같은 유전자 변형이 활발하여, 현재까지 염기서열이 분석된 동물 중 유전적으로 가장 다양한 그룹 중 하나로 밝혀졌다.[13]
현존하는 세 개의 속(''Asymmetron'', ''Branchiostoma'', ''Epigonichthys'') 중에서는 ''Asymmetron'' 속이 가장 먼저 분기한 기저 그룹이다. 이들 속의 분기 시점에 대한 분자 시계 연구 결과는 다소 차이를 보인다. 일부 연구는 ''Asymmetron'' 속이 다른 창고기 그룹으로부터 약 1억 년 전에 분기했다고 추정하는 반면[74], 다른 연구에서는 약 4600만 년 전에 분기가 일어났다고 제안한다.[65] 후자의 추정에 따르면, ''Branchiostoma'' 속과 ''Epigonichthys'' 속은 약 3830만 년 전에 서로 갈라진 것으로 보인다.[65] 이렇게 오랜 시간 동안 분리되어 있었음에도 불구하고, ''Asymmetron lucayanum''과 ''Branchiostoma floridae'' 사이의 잡종은 생존이 가능한 것으로 확인되었다. 이는 현재까지 알려진 잡종 형성 사례 중 가장 깊은 분기 시간을 가진 종들 간의 교배 사례 중 하나이다.[67]
다음 표는 WoRMS에서 인정하는 창고기목의 종 목록이다. 다른 자료에서는 약 30종을 인정하기도 하며[74][65][85], 아직 발견되지 않은 은닉종이 더 존재할 가능성이 높다.[67]
6. 모델 생물
창고기목은 1860년대 에른스트 헤켈이 모든 척추동물의 조상 모델로 제시하면서 유명해졌다. 1900년경 창고기목은 모델 생물이 되었다. 그러나 20세기 중반까지 비교 해부학과 발생학 연구의 쇠퇴, 그리고 창고기목이 겉보기보다 더 진화했다는 인식(예를 들어 유생 단계의 심한 비대칭성) 등 여러 이유로 인해 인기가 떨어졌다.[73][74] 최근에는 척추동물의 기본적인 대칭적 발달과 꼬인 발달을 설명하는 축 비틀림 이론이 제기되었는데, 이 이론에 따르면 척추동물과 두삭동물, 나아가 모든 척삭동물 사이에는 깊은 일치가 있다.[75][76]
분자 유전학의 등장으로 창고기목은 다시 척추동물의 조상 모델로 중요하게 여겨지며 모델 생물로 사용되고 있다.[77][25]
과학 연구에 사용되면서 여러 종에 대한 사육 및 번식 방법이 개발되었다. 대표적인 종으로는 유럽의 Branchiostoma lanceolatumlat, 서태평양의 Branchiostoma belcherilat와 Branchiostoma japonicumlat, 멕시코 만과 서대서양의 Branchiostoma floridaelat, 그리고 열대 지역에 분포하는 Asymmetron lucayanumlat 등이 있다. 다만 Asymmetron lucayanumlat의 경우, 유전적 증거에 따르면 대서양과 인도-태평양 개체군은 별도로 인식해야 한다는 주장이 있다.[65] 창고기목은 최대 7~8년까지 살 수 있다.[78]
7. 인간과의 관계
1860년대 에른스트 헤켈이 모든 척추동물의 조상 모델로 창고기목을 소개하면서 널리 알려졌다. 1900년경에는 모델 생물로 자리 잡았다. 그러나 20세기 중반에는 비교 해부학과 발생학 연구가 줄어들고, 창고기 유생 단계의 심한 비대칭성 등 외형과 달리 실제로는 더 진화했다는 인식이 퍼지면서 연구에서의 중요성이 다소 줄어들었다.[73][74] 최근에는 축 비틀림 이론을 통해 척추동물의 기본적인 대칭적 발달과 꼬인 발달 과정을 설명하며, 척추동물과 두삭동물, 나아가 모든 척삭동물 사이에 깊은 연관성이 있다는 주장이 제기되었다.[75][76]
분자 유전학이 발전하면서 창고기목은 다시 척추동물의 조상 모델로서 주목받으며 모델 생물로 활발히 연구되고 있다.[77][25]
과학 연구를 위해 여러 종의 사육 및 번식 방법이 개발되었다. 대표적인 종으로는 유럽의 Branchiostoma lanceolatum|브란키오스토마 란케올라툼lat, 서태평양의 Branchiostoma belcheri|브란키오스토마 벨케리lat와 Branchiostoma japonicum|브란키오스토마 야포니쿰lat, 멕시코 만과 서대서양의 Branchiostoma floridae|브란키오스토마 플로리다이lat, 그리고 열대 지역에 분포하는 Asymmetron lucayanum|아심메트론 루카야눔lat 등이 있다. 다만, Asymmetron lucayanumlat의 경우 유전적 증거에 따르면 대서양과 인도-태평양 개체군은 별개의 종으로 봐야 한다는 의견도 있다.[65][67][78] 창고기목은 최대 7~8년까지 살 수 있다.[78]
창고기목은 식용으로도 이용되며, 세계 일부 지역에서는 직접 수확하기도 한다. 신선한 상태에서는 청어와 비슷한 맛이 나며, 기름에 구워 건조시킨 형태로 식품 첨가물로 사용되기도 한다. 봄에 생식소가 성숙하기 시작하면 맛에 영향을 주어 번식기에는 맛이 떨어진다고 알려져 있다.[79]
참조
[1]
논문
The authorship of higher chordate taxa
2012-07
[2]
서적
Synopsis of the Contents of the British Museum. Forty-fourth Edition
https://www.biodiver[...]
G. Woodfall and son
[3]
논문
The lancelets and lampreys of Australia
http://biostor.org/r[...]
[4]
논문
A list of the lancelets of the world with diagnoses of five new species of ''Branchiostoma''
[5]
논문
Lancelets (cephalochordata: Branchiostomattdae): How Many Species Are Valid?
https://www.tandfonl[...]
1996-01-01
[6]
서적
The fishes of New Zealand
https://www.wikidata[...]
Te Papa Press
2015-01-01
[7]
논문
Decay of Branchiostoma: implications for soft-tissue preservation in conodonts and other primitive chordates
https://onlinelibrar[...]
1993
[8]
논문
Worms and gills, plates and spines: the evolutionary origins and incredible disparity of deuterostomes revealed by fossils, genes, and development
https://onlinelibrar[...]
2023-02
[9]
논문
A Phylogenomic Framework and Divergence History of Cephalochordata Amphioxus
2018-12-18
[10]
뉴스
Worm-like Marine Animal Providing Fresh Clues About Human Evolution
http://newswise.com/[...]
Newswise
2008-07-08
[11]
논문
An amphioxus homeobox gene: sequence conservation, spatial expression during development and insights into vertebrate evolution
http://dx.doi.org/10[...]
1992
[12]
논문
The amphioxus genome and the evolution of the chordate karyotype
[13]
논문
Decelerated genome evolution in modern vertebrates revealed by analysis of multiple lancelet genomes
2014-12-19
[14]
논문
Amphioxus functional genomics and the origins of vertebrate gene regulation
null
2018-12
[15]
논문
A New Population of the Amphioxus (Branchiostoma belcheri) in the Enshu-Nada Sea in Japan
1998
[16]
논문
The size range of suspended particles trapped and ingested by the filter-feeding lancelet Branchiostoma floridae (Cephalochordata: Acrania)
https://www.cambridg[...]
2000
[17]
논문
Filter Feeding in Lancelets (Amphioxus), vertebrate Biology
1999
[18]
논문
My Favorite Animal, Amphioxus: Unparalleled for Studying Early Vertebrate Evolution
https://hal.sorbonne[...]
2018
[19]
논문
Reproduction of the Florida Lancelet (Branchiostoma floridae): Spawning Patterns and Fluctuations in Gonad Indexes and Nutritional Reserves
1996
[20]
논문
Insights into spawning behavior and development of the european amphioxus (Branchiostoma lanceolatum)
2007
[21]
논문
Fine Structural Study of the Cortical Reaction and Formation of the Egg Coats in a Lancelet
https://www.biodiver[...]
[22]
논문
Year-Round Reproduction and Induced Spawning of Chinese Amphioxus, Branchiostoma belcheri, in Laboratory
[23]
서적
Spicilegia Zoologica. Fasciculus decimus
Gottlieb August Lange
[24]
서적
Cenni zoologici ossia descrizione sommaria delle specie nuove di animali discoperti in diverse contrade del regno nell' anno 1834
https://books.google[...]
Tipografia di Azzolino e Comp.
[25]
논문
It's a long way from amphioxus: descendants of the earliest chordate
2008-06
[26]
서적
A History of British Fishes. Vol. II
https://www.biodiver[...]
John van Voorst
[27]
웹사이트
Branchiostomatidae Bonaparte, 1846
2024-02-11
[28]
서적
Catalogo metodico dei pesci europei
https://www.biodiver[...]
Stamperia e Cartiere del Fibreno
[29]
논문
Ueber den Bau und die Grenzen der Ganoiden, und über das natürliche System der Fische
https://www.zobodat.[...]
[30]
논문
Occurrence of planktonic lancelets from Louisiana's continental shelf, with a review of pelagic ''Branchiostoma'' (order Amphioxi)
[31]
논문
A catalog of World Fishes. Part II
[32]
논문
Lancelets (Cephalochordata: Branchiostomatidae): how many species are valid?
[33]
논문
New taxonomic names of fish-like vertebrates
https://books.google[...]
[34]
서적
Entwickelungsgeschichte des Amphioxus lanceolatus
Mém Acad Sci St Petersburg.
[35]
서적
Annales des sciences naturelles
Libraires-editeurs
[36]
서적
Bemerkungen uber den Bau des Amphioxus lanceolatus eines Fisches aus der Ordnung der Cyclostomen
https://archive.org/[...]
Gebrüder Bornträger
[37]
서적
Proc. R. Soc. Edinb
[38]
서적
Z. Wiss. Zool.
[39]
간행물
The Amphioxus and Its Development
https://archive.org/[...]
[40]
서적
Dev. Growth. Differ
"1999"
[41]
간행물
Amphioxus as a model to study the evolution of development in chordates
2023-09-18
[42]
간행물
Insights from amphioxus into the evolution of vertebrate cartilage
2007-08-29
[43]
간행물
Left-right asymmetry specification in amphioxus: review and prospects
https://pubmed.ncbi.[...]
2017
[44]
간행물
Establishment of left-right asymmetric innervation in the lancelet oral region
https://onlinelibrar[...]
2001-07-09
[45]
간행물
Evolutionary history of the extant amphioxus lineage with shallow-branching diversification
2017-04-25
[46]
웹사이트
Lancelet (Branchiostoma lanceolatum)
http://www.marlin.ac[...]
Marine Life Information Network: Biology and Sensitivity Key Information Reviews
2018-01-07
[47]
서적
The Vertebrate Body
Holt-Saunders International
[48]
간행물
Muscular system
https://www.accesssc[...]
2021
[49]
간행물
Serial blockface SEM suggests that stem cells may participate in adult notochord growth in an invertebrate chordate, the Bahamas lancelet
[50]
간행물
A neurochemical map of the developing amphioxus nervous system
[51]
간행물
The origin and evolution of chordate nervous systems
[52]
간행물
Molecular regionalization of the developing amphioxus neural tube challenges major partitions of the vertebrate brain
[53]
서적
The Central Nervous System of Vertebrates
https://books.google[...]
Springer
2015-11-25
[54]
서적
Evolutionary Developmental Biology of Invertebrates 6: Deuterostomia
https://books.google[...]
Springer
2015-11-21
[55]
간행물
Evolution of phototransduction, vertebrate photoreceptors and retina
[56]
간행물
Light-transduction in melanopsin-expressing photoreceptors of Amphioxus
[57]
서적
Neural Crest Cells: Evolution, development and disease
https://books.google[...]
Academic Press
2015-11-25
[58]
간행물
The nervous system of amphioxus: Structure, development, and evolutionary significance
[59]
간행물
Molecular analysis of the amphioxus frontal eye unravels the evolutionary origin of the retina and pigment cells of the vertebrate eye
2012-09-18
[60]
서적
The Evo-Devo Origin of the Nose, Anterior Skull Base and Midface
https://books.google[...]
Springer Science & Business Media
2015-12-07
[61]
간행물
Frontal Eye Circuitry, Rostral Sensory Pathways and Brain Organization in Amphioxus Larvae: Evidence from 3D Reconstructions
http://royalsocietyp[...]
2015-12-14
[62]
간행물
Endogenous Green Fluorescent Protein (GFP) in Amphioxus
https://www.journals[...]
2007-10-01
[63]
간행물
The evolution of genes encoding for green fluorescent proteins: insights from cephalochordates (amphioxus)
null
2016-06-17
[64]
간행물
A bright monomeric green fluorescent protein derived from Branchiostoma lanceolatum
null
2013-05
[65]
간행물
Evolutionary history of the extant amphioxus lineage with shallow-branching diversification
[66]
서적
Modern Text Book of Zoology: Vertebrates
Rastogi Publications
[67]
간행물
Keeping amphioxus in the laboratory: an update on available husbandry methods
[68]
서적
Comparative Vertebrate Lateralization
https://books.google[...]
Cambridge University Press
2002-03-25
[69]
간행물
Evolutionary diversification of secondary mechanoreceptor cells in tunicata
[70]
간행물
Amphioxus as a model for investigating evolution of the vertebrate immune system
http://mosas.sysu.ed[...]
2015-12-16
[71]
논문
Identification, Evolution and Expression of an Insulin-Like Peptide in the Cephalochordate Branchiostoma lanceolatum
[72]
논문
A Novel Serpin with Antithrombin-Like Activity in Branchiostoma japonicum: Implications for the Presence of a Primitive Coagulation System
[73]
논문
The cult of amphioxus in German Darwinism; or, Our gelatinous ancestors in Naples' blue and balmy bay
2015-01
[74]
서적
The Variety of Life
Oxford University Press
[75]
논문
An ancestral axial twist explains the contralateral forebain and the optic chiasm in vertebrates
[76]
논문
Somatic twist: a model for the evolution of decussation
[77]
논문
The chordate amphioxus: an emerging model organism for developmental biology
https://www.research[...]
2004-09
[78]
웹사이트
Amphioxus Branchiostoms lanceolatum
http://www.embrc-fra[...]
EMBRC France
2018-01-07
[79]
논문
The Lancelet
https://www.research[...]
1998
[80]
논문
Functional genomics thickens the biological plot
[81]
뉴스
Lancelet (amphioxus) genome and the origin of vertebrates
https://arstechnica.[...]
Ars Technica
2008-06-19
[82]
서적
Vertebrate Palaeontology, Third Edition
Blackwell Publishing
[83]
논문
Tunicates and not cephalochordates are the closest living relatives of vertebrates.
http://worldcat.org/[...]
[84]
논문
The amphioxus genome and the evolution of the chordate karyotype
2008-06
[85]
웹사이트
World Register of Marine Species- Cephalochordates species list
http://www.marinespe[...]
2013-10-22
[86]
웹사이트
UNESCO-IOC Register of Marine Organisms (URMO) - Branchiostoma mortonense Kelly, 1966
http://www.marinespe[...]
[87]
웹사이트
WoRMS - World Register of Marine Species - Branchiostoma mortonense Kelly, 1966
http://www.marinespe[...]
[88]
웹사이트
WoRMS - World Register of Marine Species - Epigonichthys Peters, 1876
http://www.marinespe[...]
[89]
웹인용
Classification of Class: Leptocardii - Ocean Biogeographic Information System: Canadian Museum of Nature (OBIS Canada)
http://data.gbif.org[...]
2010-11-14
[90]
웹인용
활유어 (amphioxus)의 게놈 해독에 성공
http://www.gstep.re.[...]
경기과학기술원
2010-11-14
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com