선태식물
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
선태식물은 뿔이끼류, 선류, 태류를 포함하는 육상 식물 분류군이다. 배우체가 우점하며, 관다발 조직이 없고, 세대교번을 한다. 선태식물은 다양한 환경에서 서식하며, 엽상체, 경엽체 등의 형태를 보인다. 생식은 포자와 무성생식을 통해 이루어지며, 삭(capsule)이라고 불리는 포자낭을 형성한다. 일부 종은 토양 보수력 향상, 오염 연구, 조경 등에 이용되며, 이탄의 원료로도 사용된다. 한국에는 약 1665종이 분포하며, 200종 이상이 멸종 위기에 처해 있다.

더 읽어볼만한 페이지
- 민꽃식물 - 양치식물
양치식물은 석송문과 양치식물문으로 구성되며, 고사리류가 다양성의 대부분을 차지하고, 뿌리, 줄기, 잎을 가지며 포자체와 배우체가 세대교번하는 관속식물이다. - 민꽃식물 - 지의류
지의류는 균류와 조류 또는 시아노박테리아의 공생체로, 다양한 형태와 색깔을 가지며 극한 환경에서도 생존하는 복합 유기체로서, 대기오염 지표생물로 활용되는 등 환경 변화에 민감하며 다양한 분야에서 이용된다. - 측계통군 - 맹금류
맹금류는 날카로운 발톱과 부리를 가진 사나운 새를 통칭하며, 생태계 최상위 포식자로서 뛰어난 시력과 비행 능력을 갖추고 있으며, 수리, 매, 독수리, 올빼미 등이 대표적이고, 서식지 파괴 등의 위협에 직면해 있다. - 측계통군 - 돌고래
돌고래는 이빨고래류에 속하는 해양 포유류의 총칭으로, 다양한 종이 존재하며 높은 지능과 사회성을 가지지만, 서식지 파괴, 환경 오염 등으로 멸종 위기에 처해 있다. - 글로벌세계대백과를 인용한 문서/{{{분류3 - 공 (악기)
공은 금속으로 제작된 타악기로, 다양한 문화권에서 의식, 신호, 음악 연주 등에 사용되며, 형태와 용도에 따라 여러 종류로 나뉜다. - 글로벌세계대백과를 인용한 문서/{{{분류3 - 국무회의
국무회의는 대한민국 대통령을 의장으로, 예산, 법률안, 외교, 군사 등 국정 현안을 심의하는 중요한 기관이며, 대통령, 국무총리, 국무위원으로 구성되고, 정례회의는 매주 1회, 임시회의는 필요에 따라 소집된다.

2. 생식 및 생활사
육상식물은 다세포 배우체(단상 세대)와 다세포 포자체(복상 세대)가 세대교번을 하는, 단복상 세대교번형(haplodiplontic영어) 생활환을 가진다.[10] 선태식물의 경우, 핵상이 단상(n)인 배우체가 우점하고, 복상(2n)인 포자체는 배우체에 반기생한다.[11][12]
선태식물은 배우체 우점형 생활사를 가지는데, 이는 더 크고 오래 사는 식물이 반수체 배우체임을 의미한다. 이배체 포자체는 드물게 나타나며, 배우체에 붙어서 영양적으로 의존한다.
포자체의 발달 과정은 선태식물의 각 그룹마다 차이를 보인다. 이끼류와 뿔이끼류는 세포 분열이 일어나는 분열조직 영역을 가지고 있다. 뿔이끼류에서는 분열조직이 발(foot) 끝부분인 기저부에서 시작되어 세포 분열을 통해 포자체 몸체를 위쪽으로 밀어 올린다. 이끼류에서는 분열조직이 삭(capsule)과 줄기(세타, seta) 상단 사이에 위치하며, 아래쪽으로 세포를 생성하여 줄기를 늘리고 삭을 들어 올린다. 반면 우산이끼류에서는 분열조직이 없어 포자체의 신장이 거의 전적으로 세포 확장에 의해 일어난다.[14]
선태류 식물체에서 장정기와 장란기의 배열은 종 내에서 대체로 일정하지만, 일부 종에서는 환경 조건에 따라 달라질 수 있다. 장정기와 장란기가 같은 식물체에 있는 종은 '''자웅동주'''(monoicous), 다른 식물체에 있는 종은 '''자웅이주'''(dioicous)라고 한다.[15] 종자식물에서 사용되는 "자웅동주"(monoecious)와 "자웅이주"(dioecious)는 그리스어로 각각 "한 집"과 "두 집"을 의미하며, 선태류 배우체의 성을 나타낼 때는 종자식물의 포자체 성과 구별하기 위해 "-oicy" 어미를 사용한다.[15]
2. 1. 배우체 세대
우리가 흔히 보는 선태식물의 몸은 배우체로서 유성 세대에 속하며 핵상은 n이다. 즉, 배우자(난세포와 정자)를 만드는 세대로서 양치식물의 전엽체, 종자식물의 배낭과 꽃가루관에 해당된다. 장란기는 긴 목을 가진 플라스크 모양으로, 그 안에 1개의 난세포가 생긴다. 한편, 장정기는 자루가 있는 곤봉 모양이나 공 모양의 부대 모양으로, 그 안에 많은 정자가 생긴다. 2개의 편모를 가진 정자가 헤엄쳐 나와 장란기의 난세포에 닿으면 거기에서 수정이 이루어진다.[10]수정란(접합자)은 장란기 속에서 분열을 되풀이하여 포자체가 되는 배를 형성한다. 그 후 포자체는 배우체로부터 양분을 흡수하면서 성장하지만, 어린 시기에는 포자체 자체도 약간의 광합성을 한다. 따라서, 포자체는 배우체에 반기생하고 있다고 할 수 있다. 성장한 포자체는 성냥개비 모양을 한 하나의 축에 지나지 않으며, 밑부분에 있는 흡수 기관(다리), 자루(꼬투리자루) 및 끝의 포자낭(꼬투리)으로 이루어진다. 단, 뿔이끼에서는 포자체가 이쑤시개처럼 생긴 하나의 축으로 되어 있고, 끝이 뾰족하며 밑부분 외에는 모두 꼬투리로 되어 있다. 포자체는 포자를 만드는 세대로 무성 세대에 속하고 핵상은 2n으로, 우리가 흔히 보는 양치식물이나 종자식물의 몸에 해당한다. 꼬투리 속에 있는 다수의 포자 모세포는 감수 분열하여 포자를 형성한다.[10]
포자는 꼬투리에서 방출되어 바람 등에 날려 살포되는데, 적당한 곳에 떨어지면 싹이 터서 실 모양이나 덩어리진 원사체가 된다. 얼마 후에는 원사체에 싹이 나고, 이것이 발달하여 보통의 식물체(배우체)가 된다. 배우체나 포자체에는 관다발이나 다른 조직들이 분화되어 있지 않다.[10]
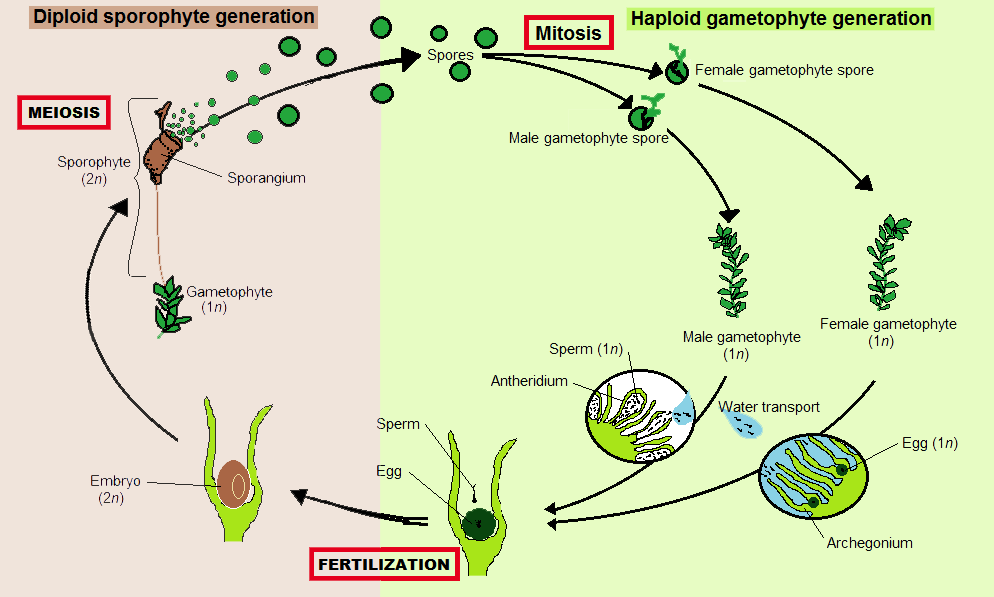
모든 육상식물(유배자식물)과 마찬가지로, 선태식물은 생활환이 세대교번을 한다.[10] 각 세대에서, 세포 하나하나가 일정 수의 짝을 이루지 않은 염색체를 포함하는 반수체 배우체는 세포가 짝을 이룬 두 벌의 염색체를 포함하는 이배체 포자체와 번갈아 나타난다. 배우체는 반수체 정자와 난자를 생성하는데, 이것들이 융합하여 이배체 접합자를 형성하고 이것이 포자체로 자란다. 포자체는 감수분열에 의해 반수체 포자를 생성하고, 이 포자는 배우체로 자란다.[10]
선태식물은 배우체 우점형이다.[11] 즉, 더 크고 오래 사는 식물이 반수체 배우체이다. 이배체 포자체는 드물게 나타나며, 배우체에 부착되어 영양적으로 의존한다.[12] 선태식물에서 포자체는 항상 가지가 없고 단일 포자낭(포자를 생성하는 삭)을 생성하지만, 각 배우체는 동시에 여러 개의 포자체를 만들 수 있다.[10]
우산이끼류, 이끼류, 뿔이끼류는 대부분의 생애를 배우체로 보낸다. 장정기(생식세포를 생성하는 기관)는 배우체에서 생성되며, 때로는 가지 끝, 잎의 겨드랑이 또는 엽상체 아래에 숨겨져 있다. ''마찬치아''와 같은 일부 선태식물은 장정기를 맺는 정교한 구조인 장정기포자체를 만든다. 정자는 편모를 가지고 있어 자신을 생성하는 장정기에서 다른 식물의 장란기로 이동해야 한다. 절지동물이 정자 이동을 돕는 경우도 있다.[13]
수정된 난자는 접합자로 되고, 장란기 내에서 포자체 배로 발달한다. 성숙한 포자체는 배우체에 부착된 상태로 남아 있다. 포자체는 세타라고 하는 줄기와 단일 포자낭 또는 삭으로 구성된다. 포자낭 내부에서 감수분열에 의해 반수체 포자가 생성된다. 이 포자들은 대부분 바람에 의해 흩어지며, 적절한 환경에 떨어지면 새로운 배우체로 발달할 수 있다. 따라서 선태식물은 석송류, 양치류 및 기타 은화식물과 유사한 방식으로 유영하는 정자와 포자의 조합으로 분산된다.[10]
포자체의 발달은 세 그룹에서 다르다. 이끼류와 뿔이끼류 모두 세포 분열이 일어나는 분열조직 영역을 가지고 있다. 뿔이끼류에서는 분열조직이 발이 끝나는 기저부에서 시작되고 세포 분열이 포자체 몸체를 위쪽으로 밀어 올린다. 이끼류에서는 분열조직이 삭과 줄기(세타) 상단 사이에 위치하며, 아래쪽으로 세포를 생성하여 줄기를 신장시키고 삭을 들어 올린다. 우산이끼류에서는 분열조직이 없으며 포자체의 신장은 거의 전적으로 세포 확장에 의해 발생한다.[14]
선태류 식물체에서 장정기와 장란기의 배열은 종 내에서 대체로 일정하지만, 일부 종에서는 환경 조건에 따라 달라질 수 있다. 주요 구분은 장정기와 장란기가 같은 식물체에 있는 종과 다른 식물체에 있는 종으로 나뉜다. 장정기와 장란기가 같은 배우체에 있는 경우 '''자웅동주'''(monoicous)라는 용어를 사용하고, 다른 배우체에 있는 경우 '''자웅이주'''(dioicous)라는 용어를 사용한다.[15]
종자식물에서는 수술(소포자낭)이 있는 꽃과 밑씨(대포자낭)가 있는 꽃이 같은 포자체에 있는 경우 "자웅동주"를 사용하고, 다른 포자체에 있는 경우 "자웅이주"를 사용한다. 이러한 용어는 때때로 선태류 배우체를 설명하기 위해 "자웅동주"와 "자웅이주" 대신 사용될 수 있다. "자웅동주"(monoecious)와 "자웅동주"(monoicous)는 모두 그리스어로 "한 집"에서 유래했으며, "자웅이주"(dioecious)와 "자웅이주"(dioicous)는 그리스어로 "두 집"에서 유래했다. "-oicy" 어미를 사용하는 용어는 종자식물의 포자체 성과 구별되는 선태류 배우체의 성을 나타낸다.[15]
자웅동주 식물은 반드시 자웅동체이며, 즉 같은 식물이 양성의 배우자를 생성한다.[15] 자웅동주 식물에서 장정기와 장란기의 정확한 배열은 다양하다. 다른 가지에 달릴 수도(자웅동주), 같은 가지에 있지만 공통 구조에 함께 있지 않을 수도(이주동체 또는 이주동주) 있으며, 공통된 "화서"(합성자웅동주)에 함께 있을 수도 있다.[15][16] 자웅이주 식물은 '''단성'''이며, 즉 개체 식물은 한 가지 성만 가진다.[15] 자웅동주, 이주동체, 합성자웅동주, 자웅이주의 네 가지 패턴이 모두 이끼 속 ''Bryum'' 종에서 나타난다.[16]
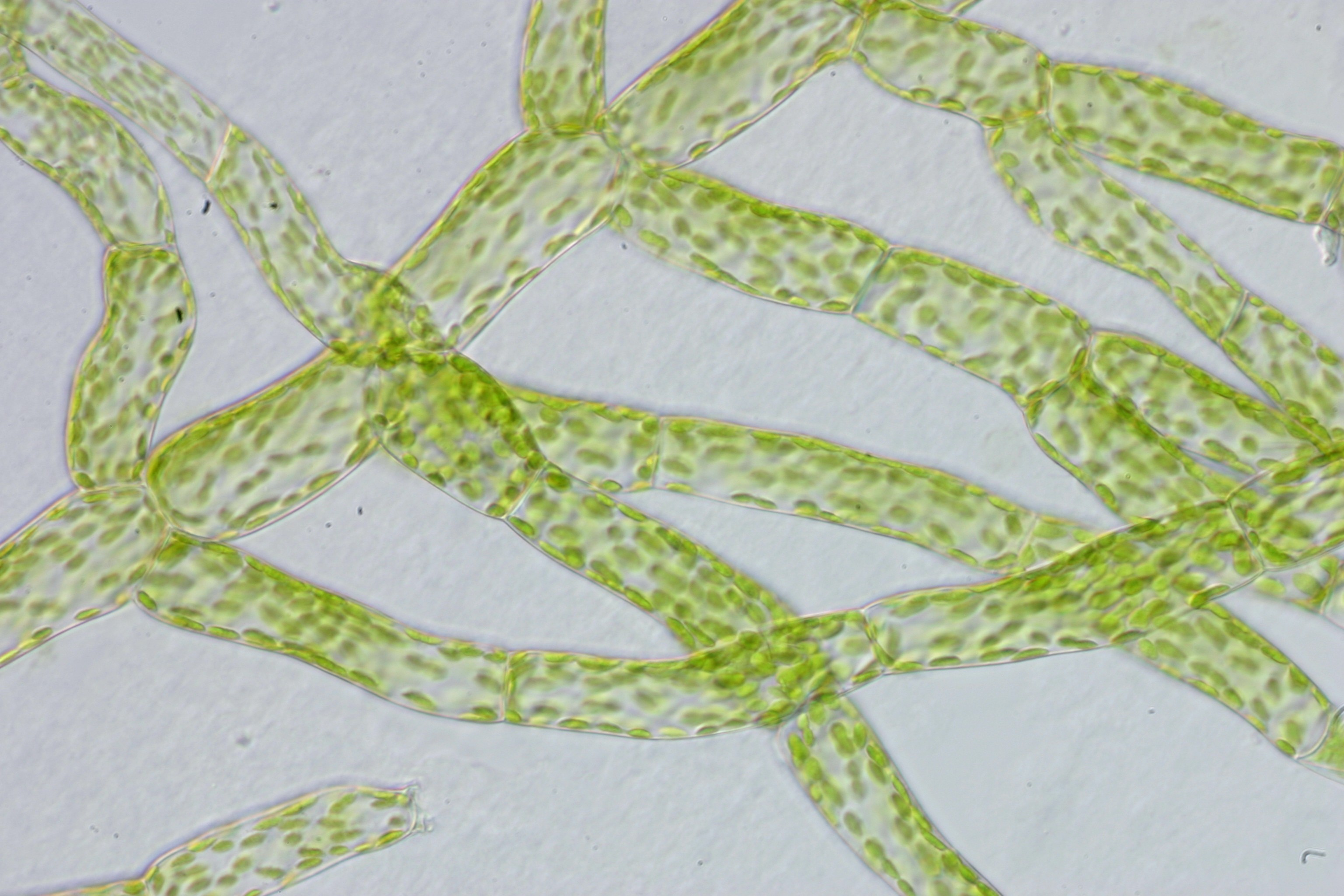
이끼식물의 배우체는 포자가 발아하여 생성된 원사체와, 그것이 분화하여 생성된 배우체의 본체로 구성된다. 배우체의 본체는 종에 따라 줄기와 잎의 분화가 명확한 '''경엽체'''(phyllid gametophore영어) 또는 명확하지 않은 '''엽상체'''(thalloid gametophore영어)인 경우가 있다. 경엽체는 모든 선류와 태류의 일부가 가지고 있으며, 엽상체는 나머지 태류와 모든 뿔이끼류가 가지고 있다. 원사체, 경엽체, 엽상체 어느 체제이든 관다발은 분화하지 않지만, 선류의 경엽체에는 하이드롬이나 렙톰이라고 불리는 통도조직이 분화하는 경우도 있다.[10]
복상의 포자체에서 포자가 방출되어 발아하면, '''원사체'''(protonema영어)라고 불리는 배우체를 형성한다. 선류의 원사체는, 처음에는 섬유상의 엽록체를 가진 클로로네마(chrolonema, feeding filament영어)가 되고, 클로로네마는 카울로네마(caulonema, foraging filament영어)로 분화한다. 카울로네마는 분지하여 배우체의 본체(gametophore영어, 경엽체 또는 엽상체)를 분화한다. 선류의 일부는 영존성의 원사체를 가지는 것이 있다. 태류나 뿔이끼류에서는, 선류보다 발달이 나쁘고, 대부분 세포의 덩어리가 되며, 섬유상의 것이라도 가지가 거의 보이지 않는다. 엽상성 태류의 원사체는 처음에 2~7개의 세포로 이루어진 짧은 섬유상이며, 그 위에 수개에서 수십 개의 세포로 이루어진 발아판을 생성하고, 그 위에서 분화된 정단세포에서 엽상체가 생성된다. 경엽성 태류에서는, 먼저 섬유상 또는 덩어리상의 원사체가 생성되고, 세포 위에서 분화된 정단세포에서 경엽체가 생성된다.[10]
배우체가 어느 정도 성장하면, 그 위에 '''장란기'''와 '''장정기'''가 형성되고, 각각 난세포와 정자를 만든다. 비 등에 의해 물에 닿았을 때, 정자가 헤엄쳐 나와 장란기 속에서 난세포와 수정하여 수정란(접합자)가 만들어진다. 수정란은 그 자리에서 발생을 시작하여, 배우체에 영양을 의존하는 반기생 생활 상태에서 발달하여 포자체를 형성한다.[10]
이끼식물의 암수는 복잡하며, 암수이주(dioicous영어)인 것과 암수동주(monoicous영어)인 것이 있다. 수컷 식물과 암컷 식물이 명확하게 나뉘는 암수이주에서는, 1개체에 장란기만을 붙이는 암컷과 1개체에 장정기만을 붙이는 수컷이 구별된다. 암수이주에서는 보통 암수의 크기는 거의 같은 크기이거나, 암컷 식물이 약간 큰 정도이지만, 일부 종에서는 수컷 개체가 명확하게 작아진다. 선류의 후쿠라고케 등에서는 암컷 식물 위에 왜웅이 착생한다. 암수동주에서는, 장란기와 장정기가 동일 개체 위에 생성되지만, 그 위치에 따라 여러 유형이 구별된다. 암수동포의 암수동주(synoicous영어)에서는, 장란기와 장정기가 동일 포엽 속에 혼생하고, 선류의 많은 종에서 볼 수 있다. 이포의 암수동주(autoicous영어)에서는, 1개체 위의 다른 포엽에 각각 장정기와 장란기만이 싸여 있고, 선류와 태류의 많은 종에서 볼 수 있다. 암수렬립동주(paroicous영어)에서는, 장란기와 장정기가 근접하지만, 장정기가 암포엽 바로 아래에 생성되어 섞이지 않는다. 태류의 많은 과와 선류의 솔이끼과 등의 일부에서 볼 수 있다.[10]
번식은, 포자에 의한 것 외에, 무성생식으로 식물체의 포복지나 탈락한 잎에서 무정아를 내어 증식한다. 무성생식을 위해 분화된 기관인 무성아를 만드는 것도 알려져 있으며, 선류와 태류에서 많은 유형이 있다. 지네이끼류에서는, 엽상체 위에 '''배상체'''(cupule영어)라고 불리는 무성아기를 형성한다.[10]
2. 2. 포자체 세대
우리가 흔히 보는 선태식물의 몸은 배우체로서 유성 세대에 속하며 핵상은 n이다. 배우체는 배우자(난세포와 정자)를 만드는 세대로서 양치식물의 전엽체, 종자식물의 배낭과 꽃가루관에 해당된다. 장란기는 긴 목을 가진 플라스크 모양으로, 그 안에 1개의 난세포가 생긴다. 한편, 장정기는 자루가 있는 곤봉 모양이나 공 모양의 부대 모양으로, 그 안에 많은 정자가 생긴다. 2개의 편모를 가지고 헤엄쳐 나온 정자가 장란기의 난세포에 닿으면 거기에서 수정이 이루어진다.[10]수정란(접합자)은 장란기 속에서 분열을 되풀이하여 포자체가 되는 배를 형성한다. 그 후 포자체는 배우체로부터 양분을 흡수하면서 성장하지만, 어린 시기에는 포자체 자체도 약간의 광합성을 한다. 따라서, 포자체는 배우체에 반기생하고 있다고 할 수 있다. 성장한 포자체는 성냥개비 모양을 한 하나의 축에 지나지 않으며, 밑부분에 있는 흡수 기관(다리), 자루(꼬투리자루) 및 끝의 포자낭(꼬투리)으로 이루어진다. 단, 뿔이끼에서는 포자체가 이쑤시개처럼 생긴 하나의 축으로 되어 있고, 끝이 뾰족하며 밑부분 외에는 모두가 꼬투리로 되어 있다. 포자체는 포자를 만드는 세대로 무성 세대에 속하고 핵상은 2n으로, 우리가 흔히 보는 양치식물이나 종자식물의 몸에 해당한다. 꼬투리 속에 있는 다수의 포자 모세포는 감수 분열하여 포자를 형성한다.[11]
포자는 꼬투리에서 방출되어 바람 등에 날려 살포되는데, 적당한 곳에 떨어지면 싹이 터서 실 모양이나 덩어리진 원사체가 된다. 얼마 후에는 원사체에 싹이 나고, 이것이 발달하여 보통의 식물체(배우체)가 된다. 배우체나 포자체에는 관다발이나 다른 조직들이 분화되어 있지 않다.
모든 육상식물(유배자식물)과 마찬가지로, 선태식물은 생활환이 세대교번을 한다. 각 세대에서, 세포 하나하나가 일정 수의 짝을 이루지 않은 염색체를 포함하는 반수체 배우체는 세포가 짝을 이룬 두 벌의 염색체를 포함하는 이배체 포자체와 번갈아 나타난다. 배우체는 반수체 정자와 난자를 생성하는데, 이것들이 융합하여 이배체 접합자를 형성하고 이것이 포자체로 자란다. 포자체는 감수분열에 의해 반수체 포자를 생성하고, 이 포자는 배우체로 자란다.[12]
선태식물은 배우체 우점형이다. 즉, 더 크고 오래 사는 식물이 반수체 배우체이다. 이배체 포자체는 드물게 나타나며, 배우체에 부착되어 영양적으로 의존한다. 선태식물에서 포자체는 항상 가지가 없고 단일 포자낭(포자를 생성하는 삭)을 생성하지만, 각 배우체는 동시에 여러 개의 포자체를 만들 수 있다.
우산이끼류, 이끼류, 뿔이끼류는 대부분의 생애를 배우체로 보낸다. 장정기(생식세포를 생성하는 기관), 장란기는 배우체에서 생성되며, 때로는 가지 끝, 잎의 겨드랑이 또는 엽상체 아래에 숨겨져 있다. ''마찬티아''와 같은 일부 선태식물은 장정기를 맺는 정교한 구조인 장정기포자체를 만든다. 정자는 편모를 가지고 있어 자신을 생성하는 장정기에서 다른 식물의 장란기로 이동해야 한다. 절지동물이 정자 이동을 돕는 경우도 있다.[13]
수정된 난자는 접합자로 되고, 장란기 내에서 포자체 배로 발달한다. 성숙한 포자체는 배우체에 부착된 상태로 남아 있다. 포자체는 세타라고 하는 줄기와 단일 포자낭 또는 삭으로 구성된다. 포자낭 내부에서 감수분열에 의해 반수체 포자가 생성된다. 이 포자들은 대부분 바람에 의해 흩어지며, 적절한 환경에 떨어지면 새로운 배우체로 발달할 수 있다. 따라서 선태식물은 석송류, 양치류 및 기타 은화식물과 유사한 방식으로 유영하는 정자와 포자의 조합으로 분산된다.
포자체의 발달은 세 그룹에서 다르다. 이끼류와 뿔이끼류 모두 세포 분열이 일어나는 분열조직 영역을 가지고 있다. 뿔이끼류에서는 분열조직이 발이 끝나는 기저부에서 시작되고 세포 분열이 포자체 몸체를 위쪽으로 밀어 올린다. 이끼류에서는 분열조직이 삭과 줄기(세타) 상단 사이에 위치하며, 아래쪽으로 세포를 생성하여 줄기를 신장시키고 삭을 들어 올린다. 우산이끼류에서는 분열조직이 없으며 포자체의 신장은 거의 전적으로 세포 확장에 의해 발생한다.[14]
육상식물은 단상 세대(다세포 배우체)와 복상 세대(다세포 포자체)의 세대교번을 하는, 단복상 세대교번형(haplodiplontic영어)의 생활환을 가지고 있다. 선태식물의 경우, 핵상은 단상(n)의 배우체가 우점하고, 복상(2n)의 포자체는 이에 반기생한다.

포자체는 배우체의 2배의 유전자 세트를 가지고 있지만, 배우체와는 형태가 크게 다르다. 선태류의 포자체는 분지하지 않고, 끝에 단일 포자낭(sporangium영어)을 형성하면 성장을 멈춘다. 특히 선태류의 포자낭은 '''삭'''(capsule영어)이라고 불린다. 삭의 내부에서는 감수분열이 일어나고, 단상의 포자가 형성된다.
히메츠리카네고케의 포자체에서는 클래스 2 KNOX 유전자가 기능하고 있으며, 이를 녹아웃하면 포자체의 발생이 억제되어 배우체가 된다. 또한, 중복되어 있던 유전자의 다른 하나의 클래스 2 KNOX 유전자는 포자체 분열조직의 형성과 유지에 관여하여, 체제 형성을 조절한다.
3. 하위 분류 및 계통
선태식물은 이끼식물이라고도 하며 뿔이끼류, 선류, 태류로 나눌 수 있다.
- 우산이끼문(Marchantiophyta) 또는 태류(苔類) : 6,000 ~ 8,000여 종[56]
- 각태식물문 (Anthocerotophyta) 또는 뿔이끼류(-類)/각태류(角苔類) : 100 ~ 200여 종[57]
- 선태식물문(Bryophyta) 또는 선류(蘚類) : 12,000여 종[58]
- † 호르네오피톤강(Horneophytopsida)
뿔이끼류는 각태류라고도 하는데 원사체는 짧은 사상이거나 없고, 포자 발아 직후 분화한다. 세포 내에는 1개에서 몇 개의 대형 엽록체가 있고 엽록체에는 피레노이드가 있다. 포자체는 선형이며 2줄로 갈라지고 중축이 있다. 불이끼가 대표적이다. 태류는 엽상체 또는 경엽체이고 세포 내의 엽록체는 많이 있으며, 소형이고 피레노이드가 없다. 포자체는 구상의 포자낭이 있으며, 포자낭은 분열하여 포자와 함께 탄사를 가진다. 태류의 대표종은 우산이끼 등이 있다. 선류는 모두 경엽체이고 잎은 나선상으로 줄기에 붙거나 좌우 2열로 붙는다. 포자체는 여러 층의 세포로 이루어지는 삭이 있으며 표면에 기실공, 내부에 중축이 있다. 포자만 만들고 탄사는 없다. 물이끼·검정이끼·솔이끼 등이 있다.







전통적으로, 관다발 조직이 없는 모든 살아있는 육상 식물은 하나의 분류군, 종종 문(또는 계통)으로 분류되었다. "Bryophyta"라는 용어는 1864년 Braun에 의해 처음 제안되었다.[17] 1879년 초, 독일의 선태류학자 Wilhelm Schimper는 세 가지 선태류 분지군(당시 각태류는 우산이끼류의 일부로 간주됨)을 포함하는 그룹을 설명하기 위해 Bryophyta라는 용어를 사용했다.[7] 2005년 연구가 이러한 전통적인 단계통적 관점을 뒷받침했지만,[19] 2010년까지 계통 분류학자들 사이에서 선태류 전체가 자연적인 그룹이 아니라는(즉, 측계통군임) 광범위한 합의가 나타났다.[20][21][22] 그러나 2014년 연구에서는 핵산 서열을 기반으로 한 이전의 계통 발생이 구성 편향의 영향을 받았으며, 아미노산 서열을 기반으로 한 계통 발생은 선태류가 결국 단계통군임을 시사한다는 결론을 내렸다.[23] 그 이후로, 게놈 및 전사체 데이터의 증가 덕분에, 핵 및 엽록체 서열을 기반으로 한 거의 모든 계통 발생 연구는 선태류가 단계통군을 형성한다는 결론을 내렸다.[23][24][25][26][27][28][29][30][31] 그럼에도 불구하고, 미토콘드리아 서열을 기반으로 한 계통 발생은 단계통적 관점을 뒷받침하지 못한다.[32]
세 가지 선태류 분지군은 Marchantiophyta(우산이끼류), Bryophyta(이끼류) 및 Anthocerotophyta(각태류)이다.[33] 그러나 이러한 분지군이 각각 Marchantiopsida, Bryopsida 및 Anthocerotopsida 강으로 계급이 낮춰질 것이 제안되었다.[25] 현재 우산이끼류와 이끼류가 Setaphyta라는 단계통 분지군에 속한다는 강력한 증거가 있다.[24][32][34] 아미노산 계통 발생 분석에 기반한 선호되는 모델은 우산이끼류(bryophytes)를 단계통군(monophyletic group)으로 나타낸다.[23]
다른 살아있는 육상식물과 비교하여, 세 가지 계통 모두 리그닌을 함유하는 관다발 조직과 여러 개의 포자낭을 가진 가지가 있는 포자체가 없다. 생활환에서 배우체의 두드러짐 또한 세 가지 우산이끼류 계통의 공유 특징이다(현존하는 관다발식물은 모두 포자체 우세임). 그러나 이 계통 발생이 정확하다면, 현존하는 관다발식물의 복잡한 포자체는 우산이끼류에 존재하는 단순한 가지가 없는 포자체와는 독립적으로 진화했을 수 있다.[23] 더 나아가, 이 견해는 기공이 식물 진화에서 단 한 번만 진화한 후, 우산이끼류에서 후속적으로 사라졌음을 의미한다.[24][28]
전통적으로 형태적 특징을 기반으로 분류할 때, 선태식물은 관다발 구조가 없다는 점으로 구분되어 왔다. 그러나 이러한 구분은 첫째, 가장 일찍 갈라져 나온 (하지만 현재는 멸종된) 비선태식물 중 일부, 예를 들어 호른식물(Horneophyton)들은 진정한 관다발 조직을 가지고 있지 않았고, 둘째, 많은 이끼류는 잘 발달된 물 전달 세포를 가지고 있기 때문에 문제가 있다.[35][36] 보다 유용한 구분은 포자체의 구조에 있을 수 있다. 선태식물에서 포자체는 단일 포자 형성 기관(포자낭)을 가진 단순한 가지가 없는 구조인 반면, 다른 모든 육상 식물인 다포자낭식물에서는 포자체가 가지를 치고 많은 포자낭을 지닌다.[37][38]
분자계통 분석 결과, 유전자별 계통 추정 결과를 통합하는 최적계통수 추정법과 모든 유전자 서열을 연결하여 분석하는 결합법을 이용한 다양한 계통수에서 다음과 같은 토폴로지를 보이는 것이 확인되었다.
식물의 학명은 국제조류·균류·식물명명규약 (ICN, 2018)에 따라 규제되고 있다. ICN에서는 학명의 공식 발표일자에 대해 그 출발점을 정하고 있지만, 선태식물에서는 분류군에 따라 출발점이 되는 날짜가 다르다. 수태과를 제외한 선류에 대해서는, Hedwig (1801) ''Species muscorum frondosorumla''에 근거하여, 1801년1월 1일을 그 출발점으로 정하고 있다. 또한, 수태과 및 뿔이끼류를 포함한 우산이끼류에 대해서는 린네 (1753) ''Species plantarumla'' ed. 1에 근거하여, 1753년5월 1일을 출발점으로 정하고 있다. 다만, 속보다 상위의 계급의 분류군에 대해서는, 쥐시외 (1789) ''Genera plantarumla''에 근거하여, 1789년8월 4일이 그 출발점으로 여겨진다. 또한, 화석 식물에 관해서는 다른 식물과 마찬가지로 1820년12월 31일이 출발점으로 여겨진다.
최근의 분류 체계에서는 선태식물이 단계통이 아니라고 여겨짐에 따라, 선태식물에 포함되는 선류, 태류, 뿔이끼류를 각각 문의 계급으로 두는 분류가 사용되어 왔다.
''Bryophytala'' 라는 학명이 선태식물 전체와 선류 모두에 사용되어 왔기 때문에, 계급 접미사를 붙인 학명은 다루기 어렵다. 따라서, Brent Mishler (2010) 등은 계급 접미사를 갖지 않는 전통적인 학명을 선호하여, 선류에는 를, 태류에는 를, 그리고 뿔이끼류에는 를 사용했다. Sousa 등 (2018)에서는 이끼식물의 단계통성이 명확해진 현재, 선류, 태류, 뿔이끼류를 이전처럼 강으로 강등해야 한다고 주장했다. 이 경우, 이끼식물의 내부 계통 및 계급은 다음과 같다.
다음은 Goffinet & Shaw (2008)을 기반으로 한 樋口 (2012), 그리고 Crandall-Stotler 외 (2009)(태류), Goffinet 외 (2008)(선류) 및 Renzaglia 외 (2008)(뿔이끼류)를 기반으로 한 海老原 외 (2012)에 따른 내용이다. 3문으로 분류된 이끼식물에 계급을 부여하는 경우, 아계에 ''''를 사용하기도 한다.
- 태류식물문(태류)
- 꼬마이끼강
- 꼬마이끼아강
- 트레우비이끼아강
- 지의이끼강
- 민지의이끼아강
- 지의이끼아강
- 우산이끼강(비늘이끼강)
- 물지의이끼아강
- 두갈래이끼아강
- 우산이끼아강(비늘이끼강)
- 선류식물문(선류)
- subdivision영어 (Superclass I영어 ''sensula'' )
- 나나자몬자이끼강
- subdivision영어 (Superclass II영어 ''sensula'' )
- 물이끼강
- subdivision영어 (Superclass III영어 ''sensula'' )
- 검정이끼강
- subdivision영어 (Superclass IV영어 ''sensula'' )
- 검정솔이끼강
- subdivision영어 (Superclass V영어 ''sensula'' )[51]
- 돌이끼강
- 솔이끼강
- 네잎이끼강
- 이끼강
- 담뱃대이끼아강
- 두줄이끼아강
- 잔디이끼아강
- 병이끼아강
- 꼬마솔이끼아강
- 이끼아강
- 뿔이끼식물문(뿔이끼류)
- 줄뿔이끼강(레이오스포로케로스강)
- 뿔이끼강
- 뿔이끼아강
- 헛뿔이끼아강
- 나무뿔이끼아강
4. 형태
선태식물은 뿔이끼류, 선류, 태류로 나눌 수 있으며, 육상에서 습한 땅, 바위, 썩은 나무 등에 착생한다. 직사광선을 싫어하며, 고산지대, 극지, 열대 우림 등 다양한 환경에서 서식한다.
선태식물은 다음과 같은 특징을 가진다.
형태적 특징으로 분류할 때, 관다발 구조가 없다는 점으로 구분했지만, 일부 이끼류는 잘 발달된 물 전달 세포를 가지고 있어 문제가 있다. 포자체 구조를 보면, 선태식물은 단일 포자낭을 가진 단순한 가지가 없는 구조인 반면, 다른 육상 식물은 가지를 치고 많은 포자낭을 지닌다.
녹조류와 마찬가지로 선태류는 엽록체에 저장된 녹말을 생성하고 세포벽에 셀룰로오스를 함유한다. 또한 큐티클, 기공, 배우자낭 발달, 배 발생 등의 특징은 육상 환경 적응에 도움을 주었으며, 생식과 생존을 위해 물에 의존한다.
세 가지 선태류의 배우체 형태 특징은 다음과 같다.

모든 이끼식물이 가지는 공통점은 포자체가 퇴화하고 배우체에 반기생한다는 것이다.
뿔이끼류와 선류에서는 식물체의 끝 부분 이외에도 개재분열조직이라고 불리는 분열조직이 생기며, 태류에는 없지만, 최소진화적 이끼식물의 공통 조상에서 획득되었다고 생각되는 특징이다.
태류와 뿔이끼류는 '''탄사'''를 가지고 있으며, 포자 형성 세포가 체세포 분열함으로써 포자 모세포와 함께 탄사 세포가 형성된다. 뿔이끼류는 기부에서 분지하며, 태류도 가지고 있으므로, 이끼식물의 공유파생형질이라고 생각되지만, 파생적인 그룹인 키노보리츠노고케속 및 아나나시츠노고케속과 같은 키노보리츠노고케과를 제외한 뿔이끼류는 나선형 비후를 가지지 않는 '''가탄사'''이며, 이끼식물의 공통 조상은 탄사를 가지지 않고, 태류의 공통 조상과 키노보리츠노고케속에서 평행 진화했다고도 추정된다.
선류와 태류는 유사한 독특한 편모 장치를 형성한다.
3그룹의 형태를 비교하면 다음과 같다.
4. 1. 배우체 형태
이끼식물의 배우체는 포자가 발아하여 생성된 원사체와, 그것이 분화하여 생성된 배우체의 본체로 구성된다. 배우체의 본체는 종에 따라 줄기와 잎의 분화가 명확한 '''경엽체'''(phyllid gametophore영어) 또는 명확하지 않은 '''엽상체'''(thalloid gametophore영어)인 경우가 있다. 경엽체는 모든 선류와 태류의 일부가 가지고 있으며, 엽상체는 나머지 태류와 모든 뿔이끼류가 가지고 있다. 원사체, 경엽체, 엽상체 어느 체제이든 관다발은 분화하지 않지만, 선류의 경엽체에는 하이드롬이나 렙톰이라고 불리는 통도조직이 분화하는 경우도 있다.먼저 복상()의 포자체에서 포자가 방출되어 발아하여, '''원사체'''(protonema영어)라고 불리는 배우체()를 형성한다. 선류의 원사체는 처음에는 섬유상의 엽록체를 가진 클로로네마(chrolonema, feeding filament영어)가 되고, 클로로네마는 카울로네마(caulonema, foraging filament영어)로 분화한다. 카울로네마는 분지하여 배우체의 본체(gametophore영어, 경엽체 또는 엽상체)를 분화한다. 선류의 일부는 영존성의 원사체를 가지는 것이 있다. 태류나 뿔이끼류에서는 선류보다 발달이 나쁘고, 대부분 세포의 덩어리가 되며, 섬유상의 것이라도 가지가 거의 보이지 않는다. 엽상성 태류의 원사체는 처음에 2~7개의 세포로 이루어진 짧은 섬유상이며, 그 위에 수개에서 수십 개의 세포로 이루어진 발아판을 생성하고, 그 위에서 분화된 정단세포에서 엽상체가 생성된다. 경엽성 태류에서는 먼저 섬유상 또는 덩어리상의 원사체가 생성되고, 세포 위에서 분화된 정단세포에서 경엽체가 생성된다.
배우체가 어느 정도 성장하면, 그 위에 '''장란기'''와 '''장정기'''가 형성되고, 각각 난세포와 정자를 만든다. 비 등에 의해 물이 닿았을 때, 정자가 헤엄쳐 나와 장란기 속에서 난세포와 수정하여 수정란(접합자)가 만들어진다. 수정란은 그 자리에서 발생을 시작하여, 배우체에 영양을 의존하는 반기생 생활 상태에서 발달하여 포자체를 형성한다.
이끼식물의 암수는 복잡하며, 암수이주(dioicous영어)인 것과 암수동주(monoicous영어)인 것이 있다. 수컷 식물과 암컷 식물이 명확하게 나뉘는 암수이주에서는, 1개체에 장란기만을 붙이는 암컷과 1개체에 장정기만을 붙이는 수컷이 구별된다. 암수이주에서는 보통 암수의 크기는 거의 같은 크기이거나, 암컷 식물이 약간 큰 정도이지만, 일부 종에서는 수컷 개체가 명확하게 작아진다. 선류의 후쿠라고케() 등에서는 암컷 식물 위에 왜웅이 착생한다. 암수동주에서는 장란기와 장정기가 동일 개체 위에 생성되지만, 그 위치에 따라 여러 유형이 구별된다. 암수동포의 암수동주(synoicous영어)에서는 장란기와 장정기가 동일 포엽 속에 혼생하고, 선류의 많은 종에서 볼 수 있다. 이포의 암수동주(autoicous영어)에서는 1개체 위의 다른 포엽에 각각 장정기와 장란기만이 싸여 있고, 선류와 태류의 많은 종에서 볼 수 있다. 암수렬립동주(paroicous영어)에서는 장란기와 장정기가 근접하지만, 장정기가 암포엽 바로 아래에 생성되어 섞이지 않는다. 태류의 많은 과와 선류의 솔이끼과 등에서 볼 수 있다.
번식은 포자에 의한 것 외에, 무성생식으로 식물체의 포복지나 탈락한 잎에서 무정아를 내어 증식한다. 무성생식을 위해 분화된 기관인 무성아를 만드는 것도 알려져 있으며, 선류와 태류에서 많은 유형이 있다. 지네이끼류에서는 엽상체 위에 '''배상체'''(cupule영어)라고 불리는 무성아기를 형성한다.

태류의 공유파생형질은 엽상세포 내에 '''유체'''(oil body영어)라 불리는 막으로 둘러싸인 세포소기관을 갖는 것이며, 다른 육상식물에는 없는 특징이다.
태류의 배우체는 '''경엽체이기도 하고 엽상체이기도 하며''', 전통적으로 경엽성 태류와 엽상성 태류의 두 그룹으로 구분되어 왔다. 또한, 엽상성 태류에는 엽상체 내부에 기실 등의 조직 분화가 보이는 복잡엽상성 태류와 조직 분화가 적은 단순엽상성 태류로 세분되어 왔다. 그러나 분자계통 분석에 의해 이러한 그룹들이 계통을 반영하지 않는다는 것이 밝혀졌고, 현재는 꼬마이끼강, 지네고사리강, 솔이끼강의 세 그룹으로 재편되었다. 경엽성 태류와 단순엽상성 태류는 하나의 클레이드에 모이며, 경엽성 태류에서 여러 차례 단순엽상성으로의 체제 진화가 일어났다는 것을 알 수 있다. 꼬마이끼강은 경엽성을 가지며, 특히 트로이브고케아강(트로이브고케과로 구성된 단형 아강)은 경엽체와 엽상체의 중간적인 형태를 갖는다. 지네고사리강은 엽상체로만 구성된 그룹이다. 특히 복잡엽상성 체제가 전형적이지만, 두 종으로 구성된 얇은지네고사리아강에서는 예외적으로 단순엽상성 체제를 갖는다. 태류의 대부분을 포함하는 솔이끼강은 직립하는 경엽성이나 포복하는 경엽성, 단순엽상성 등 다양한 형태를 갖는다. 그중 솔이끼아강은 경엽성 체제가 대부분이지만, 물지네고사리아강과 두갈래이끼아강은 단순엽상성을 갖는 것이 많다.
가장 기저에서 분지한 꼬마이끼강은 장란기와 장정기를 보호하는 엽상기관을 가지며, 헛뿌리를 형성하지 않고, 잎이 없는 근경으로 기질에 부착한다. 이러한 형질은 원시적인 형태로 간주된다. 태류의 공통 조상이 꼬마이끼강과 같은 경엽성이었다면, 엽상성 태류는 경엽성 태류에서 진화했을 것이다. 엽상성 태류의 복측에 있는 인편은 경엽체의 잎과 같은 발생 과정에 의해 생기므로, 잎이 축소된 것으로 생각된다.
선태류는 모두 '''엽상체''' 형태를 가지며, 대부분 나선형으로 '''잎''' (phyllid영어)을 갖는다. 또한, 가근은 다세포이며 가지를 친다.
선태류의 대부분은 솔이끼강에 포함되며, 나머지 그룹은 선태류 진화의 초기 단계에서 분기된 잔존적인 분류군으로 생각된다. 많은 그룹은 삭의 꼭대기에 '''뚜껑'''(삭뚜껑)이 분화되어 있으며, 삭에서 뚜껑이 분리되면 삭의 개구부 안쪽에 가늘고 긴 치아 모양의 구조물인 '''삭치'''(peristome영어)가 늘어선다. 선태류는 포자체의 삭치 구조에 따라 무관절삭치선태류와 유관절삭치선태류로 크게 나뉜다. 유관절삭치선태류는 단계통군이지만, 무관절삭치선태류는 다계통군이 된다. 솔이끼강은 '''무관절삭치'''(nematodontous peristome영어)를 가지고, 솔이끼강은 '''유관절삭치'''(arthrodontous peristome영어)를 가지며, 이들과 삭치를 갖지 않는 이시즈치고케 (Oedipodium griffithianum) 1종으로 구성되는 이시즈치고케강이 자매군이 된다.
2종으로 구성되는 난자몬자고케강은 잎이 막대 모양이며, 잎이 없는 뿌리줄기 모양의 줄기를 가지고, 가근을 형성하지 않는다. 또한 장란기와 장정기가 노출되고, 포자낭이 비스듬히 갈라지는 것도 다른 선태류와 다른 특징이며, 과거에는 태류로 간주되기도 했다. 물이끼강 및 검은이끼강은 삭병이 없으며, 배우체 조직이 신장한 '''가짜자루'''(가짜발, pseudopodium영어)에 의해 포자체가 들어 올려진다. 검은이끼강은 검은이끼강과 매우 유사하지만, 삭병을 갖는다.
4. 2. 포자체 형태
선태식물 포자체는 배우체와 형태가 크게 다르며, 분지하지 않고 끝에 단일 포자낭(삭)을 형성하면 성장을 멈춘다.[47]뿔이끼류는 엽상체에서 자루와 포자낭의 경계가 뚜렷하지 않은 뿔 모양의 포자체를 가진다. 이는 삽입 분열 조직의 분열 활성이 선태류보다 오래 지속되기 때문이다. 또한, 세포 내에 1~2개의 엽록체만 가지는 단색소체성이며, 엽록체에 피레노이드를 가지는 것이 특징이다. 엽상체 내에는 시아노박테리아가 공생하고 있다.
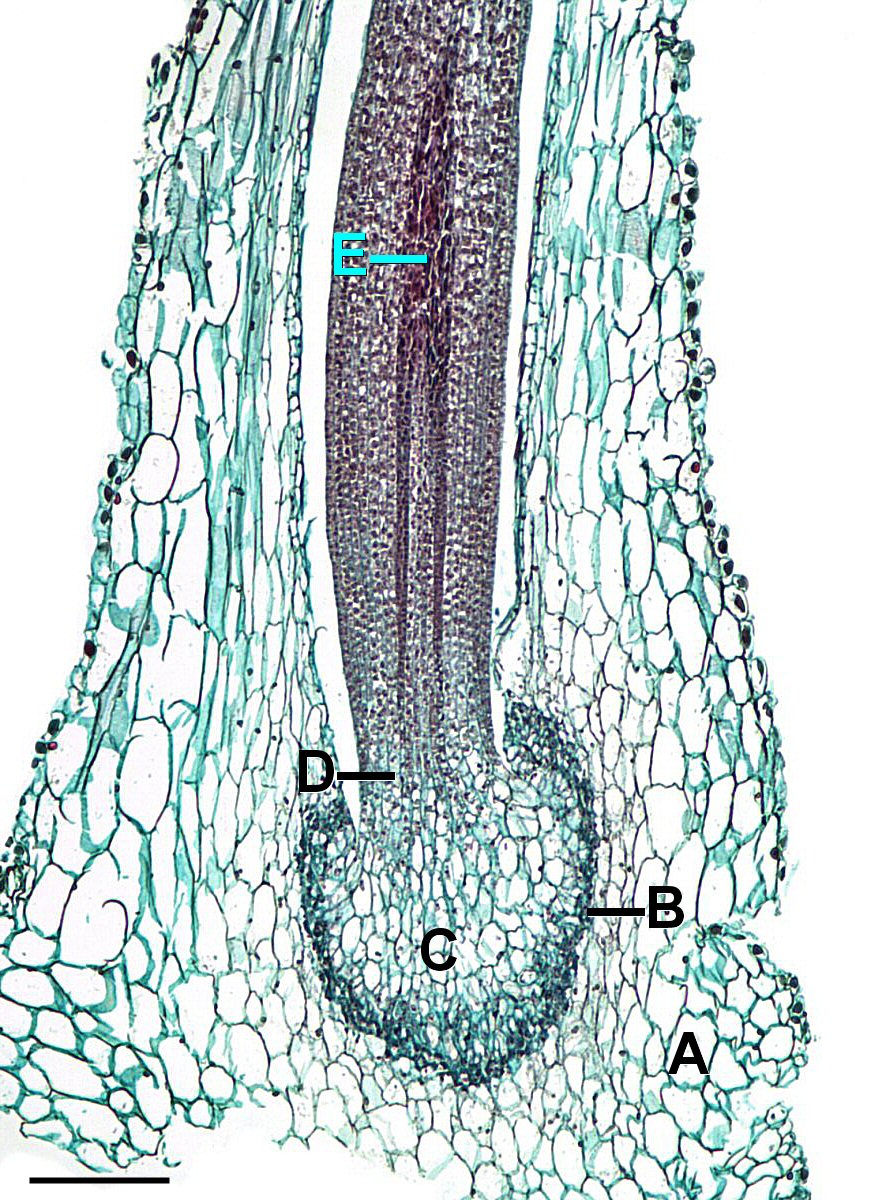
육상 식물 중에서 뿔이끼류의 정자기관 형태는 특이하다. 대부분의 현생 육상 식물에서는 정자기관낭의 최외층 세포가 외부와 접하고, 전관속식물에서도 정자기관은 조직 위로 돌출되어 있지만, 뿔이끼류의 정자기관은 주변 조직 내에서 형성된다. 장란기 또한 다른 육상 식물과 달리, 목의 최선단 세포가 표피 세포 위로 돌출되지 않는다.
세 가지 선태류의 포자체 형태 특징은 다음과 같다.
5. 서식지 및 분포
선태식물은 육상에서 생활하며 습한 땅, 바위 위, 썩은 나무, 나무줄기 등에 붙어서 자란다. 또한 활엽수의 잎에 붙어서 자라는 특수한 종류도 있다. 선태식물은 종류에 따라 생육 장소가 대개 정해져 있다. 예를 들어, 석회암에서만 자라는 히오필라속이나 화강암과 같은 산성암에서만 자라는 종류가 있다.[9]
선태식물은 보통의 고등식물과 달리 직사광선을 매우 싫어하는 경우가 많다. 이는 햇빛의 복사열 때문에 수분이 증발하여 말라 죽기 때문으로 보인다. 선태식물은 높은 산꼭대기부터 극지의 바위 등에서도 자라며, 열대 우림 속에서는 특히 많은 종류를 볼 수 있다. 한국에서도 흔히 볼 수 있다.[9]
선태식물은 추운 북극에서부터 뜨거운 사막, 해수면부터 고산지대, 건조한 사막에서부터 습한 열대우림까지 다양한 환경에서 자란다. 뿌리가 없기 때문에 관다발식물이 자랄 수 없는 바위나 맨땅에서도 생존할 수 있다.[9]
이끼식물은 바닷물과 빙설을 제외한 지구상의 거의 모든 곳에 서식한다. 기본적으로 육상 생활을 하지만, 담수에서 자라는 종도 있다. 토양, 부식토, 암석, 다른 식물체(나무껍질, 잎 표면, 가지) 등 다양한 곳에 붙어 산다.
온대 및 열대 지역 간에 종 다양성의 정도에 큰 차이가 없다는 것이 밝혀졌다. 적어도 이끼식물에서는 열대에 종 다양성이 집중되어 있지 않으며, "모든 것이 어디에나 있다"라고 평가된다. 또한 같은 지역이라도 고산과 저산에서는 종 구성이 크게 다르다. 고산에서 이끼식물의 생육 한계선을 이끼선(moss-line)이라고 한다. 건조에 강하거나 가혹한 환경을 좋아하는 종도 있다.
선태류와 태류는 식물 군락 내 지표면 가까이에 선태층(또는 이끼층)을 만든다. 다소 습한 삼림의 가장 아래층이나 습지에 발달하고, 낙엽이 두껍게 쌓이지 않는 암석이나 쓰러진 나무 위에 형성되어 나무의 어린 싹이 정착하는 장소가 된다. 지표면 근처를 서식지로 하는 곤충 등 작은 동물도 풍부하다.
삼림에 서식하는 종이 많지만, 암벽이나 계곡, 폭포 주변에도 많은 종이 발견된다. 특히 일 년 내내 습도가 높은 운무림에는 숲 바닥뿐만 아니라 나무줄기나 가지에까지 많은 이끼가 착생하는 경우가 있으며, 선태림(또는 이끼림)이라고도 불린다. 밭에는 밭이끼, 논에는 은행잎뜨기이끼 및 뜨기이끼 등 독특한 종이 있으며, 도시 지역에서도 몇몇 종이 자란다. 예를 들어, 히지키이끼는 돌담 등 암석 위에 직접 자란다.
물이끼류 등 이끼식물이 많이 자라는 습한 초원은 이끼습원(moss moor)이라고 불린다. 특히 물이끼류가 많이 자라는 습원을 물이끼습원이라고 하며, 그중에서 빗물에 의해 형성된 것을 Sphagnoplatum이라고 한다. 캐나다의 삼림 내에 있는 물이끼습원은 muskey라고 불린다. 물이끼류는 이탄습지, 고층습원에 많이 서식하며, 많은 양의 수분을 저장하여 건조에도 견딜 수 있다. 고층습원의 토양은 부식산과 불포화 콜로이드 때문에 산성이며, 수산화물 이온을 싫어하는 물이끼류가 중앙부에 잘 자라 이탄화가 진행되어 융기한다. 또한 북극권의 툰드라 지역은 넓은 지역이 이끼식물과 지의류에 덮여 있으며, 다소 습한 곳에 이끼툰드라가 발달한다. 물이끼류는 그 주요 종이다.
매우 특수한 환경에서 자라는 종도 있다. 피자식물이나 양치식물의 잎 위에는 곰팡이이끼나 요우조우이끼와 같은 생엽상태류가 자란다.[52] 말다이이끼 등은 동물의 배설물이나 사체에만 자라는 배설물생 종이다. 가는글씨이끼는 구리에 대한 내성이 강하다. 또한 담수에 자라는 종 중에는, 배나무이끼속처럼 남극의 호수 바닥에 서식하여 이끼 삿갓(이끼보우즈, moss pillars)을 형성하는 것도 있다.[53][54]


6. 진화
"Bryophyta"라는 용어는 1864년 브라운(Braun)에 의해 처음 제안되었다.[17] 1879년 초, 독일의 선태류학자 빌헬름 쉼퍼(Wilhelm Schimper)는 세 가지 선태류 분지군(당시 각태류는 우산이끼류의 일부로 간주됨)을 포함하는 그룹을 설명하기 위해 Bryophyta라는 용어를 사용했다.[7] G.M. Smith는 이 그룹을 조류와 양치식물 사이에 배치했다.[18]
2005년 연구가 이러한 전통적인 단계통적 관점을 뒷받침했지만,[19] 2010년까지 계통 분류학자들 사이에서 선태류 전체가 자연적인 그룹이 아니라는(즉, 측계통군임) 광범위한 합의가 나타났다.[20][21][22] 그러나 2014년 연구에서는 핵산 서열을 기반으로 한 이전의 계통 발생이 구성 편향의 영향을 받았으며, 아미노산 서열을 기반으로 한 계통 발생은 선태류가 결국 단계통군임을 시사한다는 결론을 내렸다.[23] 그 이후로, 게놈 및 전사체 데이터의 증가 덕분에, 핵 및 엽록체 서열을 기반으로 한 거의 모든 계통 발생 연구는 선태류가 단계통군을 형성한다는 결론을 내렸다.[23][24][25][26][27][28][29][30][31] 그럼에도 불구하고, 미토콘드리아 서열을 기반으로 한 계통 발생은 단계통적 관점을 뒷받침하지 못한다.[32]
세 가지 선태류 분지군은 Marchantiophyta(우산이끼류), Bryophyta(이끼류) 및 Anthocerotophyta(각태류)이다.[33] 그러나 이러한 분지군이 각각 Marchantiopsida, Bryopsida 및 Anthocerotopsida 강으로 계급이 낮춰질 것이 제안되었다.[25] 현재 우산이끼류와 이끼류가 Setaphyta라는 단계통 분지군에 속한다는 강력한 증거가 있다.[24][32][34] 아미노산 계통 발생 분석에 기반한 선호되는 모델은 우산이끼류(bryophytes)를 단계통군(monophyletic group)으로 나타낸다.[23]
다른 살아있는 육상식물과 비교하여, 세 가지 계통 모두 리그닌을 함유하는 관다발 조직과 여러 개의 포자낭을 가진 가지가 있는 포자체가 없다. 생활환에서 배우체의 두드러짐 또한 세 가지 우산이끼류 계통의 공유 특징이다(현존하는 관다발식물은 모두 포자체 우세임). 그러나 이 계통 발생이 정확하다면, 현존하는 관다발식물의 복잡한 포자체는 우산이끼류에 존재하는 단순한 가지가 없는 포자체와는 독립적으로 진화했을 수 있다.[23] 더 나아가, 이 견해는 기공이 식물 진화에서 단 한 번만 진화한 후, 우산이끼류에서 후속적으로 사라졌음을 의미한다.[24][28]
5억 1천만 년에서 6억 3천만 년 전 사이에 녹조류(green algae) 내에서 육상 식물이 출현했다.[41] 분자 계통 발생학 연구에 따르면, 선태식물(bryophytes)은 현존하는 육상 식물 중 가장 먼저 갈라져 나온 계통이다.[20][2][42][43]
분자계통 분석 결과, 유전자별 계통 추정 결과를 통합하는 최적계통수 추정법과 모든 유전자 서열을 연결하여 분석하는 결합법을 이용한 다양한 계통수에서 다음과 같은 계통수가 나타났다.
다음은 유배식물의 계통 분류이다.[59][60][61][62][63][64][65]
{| class="wikitable"
|+ 유배식물
|-
! 선태식물
! 다포자낭식물
|-
|