속새류
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
속새류는 관다발식물문의 쐐기잎식물아문에 속하는 식물 분류군으로, 린네의 분류 체계에서는 양치류에 포함되었다. 현재는 진정 양치강의 속새아강으로 분류되며, 솔잎란강 다음으로 분기되었다. 속새류는 뿌리, 줄기, 잎이 분화된 포자체와 쐐기잎, 포자낭상을 특징으로 한다. 줄기는 마디 구조를 가지며, 광합성을 한다. 잎은 마디에서 돌려나며, 쐐기잎이라고 불린다. 속새류의 화석은 데본기부터 발견되었으며, 석탄기에 번성했다. 현생 속새속은 15종으로, 전 세계 온대 습지에 분포한다.

더 읽어볼만한 페이지
- 속새강 - 쇠뜨기
쇠뜨기는 뿌리줄기로 번식하는 다년생 초본으로, 어린 순은 식용 가능하고 이뇨 작용 효능이 있으며, 영양줄기와 생식줄기로 나뉘고 꽃 대신 포자로 번식하며, 전 세계에 분포하지만 강한 번식력으로 농작물에 피해를 주기도 하고, 약용 및 산업적으로 활용되나 과다 섭취 시 건강 문제가 발생할 수 있다. - 속새강 - 칼라미테스
칼라미테스는 석탄기와 페름기에 번성했던 멸종된 속새류 식물로, 대나무와 유사한 특징을 가지며, 포자를 통해 번식하고 초기 페름기에 멸종되었지만 현생 속새류의 조상으로 여겨진다. - 표시 이름과 문서 제목이 같은 위키공용분류 - 라우토카
라우토카는 피지 비치레부섬 서부에 위치한 피지에서 두 번째로 큰 도시이자 서부 지방의 행정 중심지로, 사탕수수 산업이 발달하여 "설탕 도시"로 알려져 있으며, 인도에서 온 계약 노동자들의 거주와 미 해군 기지 건설의 역사를 가지고 있고, 피지 산업 생산의 상당 부분을 담당하는 주요 기관들이 위치해 있다. - 표시 이름과 문서 제목이 같은 위키공용분류 - 코코넛
코코넛은 코코넛 야자나무의 열매로 식용 및 유지로 사용되며, 조리되지 않은 과육은 100g당 354kcal의 열량을 내는 다양한 영양 성분으로 구성되어 있고, 코코넛 파우더의 식이섬유는 대부분 불용성 식이섬유인 셀룰로오스이며, 태국 일부 지역에서는 코코넛 수확에 훈련된 원숭이를 이용하는 동물 학대 문제가 있다. - 한국어 위키백과의 링크가 위키데이터와 같은 위키공용분류 - 라우토카
라우토카는 피지 비치레부섬 서부에 위치한 피지에서 두 번째로 큰 도시이자 서부 지방의 행정 중심지로, 사탕수수 산업이 발달하여 "설탕 도시"로 알려져 있으며, 인도에서 온 계약 노동자들의 거주와 미 해군 기지 건설의 역사를 가지고 있고, 피지 산업 생산의 상당 부분을 담당하는 주요 기관들이 위치해 있다. - 한국어 위키백과의 링크가 위키데이터와 같은 위키공용분류 - 코코넛
코코넛은 코코넛 야자나무의 열매로 식용 및 유지로 사용되며, 조리되지 않은 과육은 100g당 354kcal의 열량을 내는 다양한 영양 성분으로 구성되어 있고, 코코넛 파우더의 식이섬유는 대부분 불용성 식이섬유인 셀룰로오스이며, 태국 일부 지역에서는 코코넛 수확에 훈련된 원숭이를 이용하는 동물 학대 문제가 있다.

2. 역사적 분류
칼 폰 린네가 『Species Plantarumla』(1753년)에서 처음 식물 분류 체계를 제시했을 때, 속새류는 양치류에 포함되었다.[1] 이후 아우구스트 아이히러(1883)는 속새류가 쐐기 모양의 잎이나 특이한 마디 구조를 가진 줄기 형태를 지닌다는 점에 주목하여,[2] 양치식물 중에서도 독립된 속새강으로 분류하고, 다른 양치류가 포함된 양치강 및 현재의 석송류가 포함된 석송강과는 구분하였다.[3]
에드워드 제프리는 관다발 식물에 석송류와 진정 양치류의 두 계통이 있다는 것을 처음으로 인식했지만, 속새류는 석송류에 포함된다고 생각했다.[4][5] 1920년대 이후 데본기의 화석 양치식물 연구가 진행되면서, 양치식물의 각 강의 차이는 양치류와 겉씨식물의 차이보다 크다고 여겨지게 되었다.[6] 이에 따라 속새류는 관다발식물문 속 쐐기잎식물 아문으로 분류되어 종자식물이나 양치류를 포함하는 진정양치식물 아문 및 석송식물 아문과는 구분되었다.[3]
그 후, 양치식물은 고비목(무엽류), 석송류(석송류), 속새류(쐐기잎류), 양치류(진정 양치류)의 4군으로 크게 분류된다는 것이 정설이 되었다.[7][8] 각 군은 연구자에 따라 문이나 아문, 강, 목 등 다양한 상위 분류 계급에 속해 왔다.[7]
초기의 분자 계통 분석에서는 석송류를 제외한 3개의 분류군이 종자식물의 자매군으로서 단계통군(진정 양치식물)을 이루는 것으로 나타났으며, 속새류는 그 중에서도 기부에서 분기된 계통임이 시사되었지만, 다른 진정 양치류와의 계통 관계는 아직 밝혀지지 않았다.[2] 그 후, DNA 정보량을 늘린 분자 계통 분석에 의해 진정 양치류 중에서 가장 기부에 분기되는 (나머지 자매군이 되는) 것으로 나타나,[9][10] 현재는 진정 양치강의 하위에 속하며 '''속새아강'''으로 분류한다.[11][12][13][14]
계급과 명칭, 그리고 채택한 분류 체계의 예는 다음과 같다.
속새류는 분자 계통 분석 결과, 진정 양치류 중에서 가장 기부에서 분기된 계통으로 밝혀졌다.[13][14][15] 대엽식물에 속하며, 종자식물을 포함하는 목질식물과 함께 대엽식물을 구성한다.
3. 계통 분류
2006년 스미스(Smith) 등과 2011년 리(Li) 등, 2013년 그레위(Grewe) 등의 연구에 기초한 계통 분류와 2016년 크리스텐휴즈(Christenhusz)와 채즈(Chase) 등의 연구 결과에 기초한 대안적 계통 분류가 있다.
초기 분자 계통 분석에서는 석송류를 제외한 3개의 분류군이 종자식물의 자매군으로서 단계통군(진정 양치식물)을 이루는 것으로 나타났다. 속새류는 그 중에서도 기부에서 분기된 계통임이 시사되었지만, 다른 진정 양치류와의 계통 관계는 아직 밝혀지지 않았다. 그 후, DNA 정보량을 늘린 분자 계통 분석에 의해 진정 양치류 중에서 가장 기부에 분기되는 (나머지 자매군이 되는) 것으로 나타났다. 현재는 진정 양치강의 하위에 속하며 '''속새아강'''으로 분류한다.
속새류를 포함하는 대엽 양치류는 종자식물을 포함하는 클레이드인 목질식물과 함께 대엽식물로 묶이며, 트리메로피톤류를 스템 그룹으로 한다. 과거에 속새류로 분류되었던 화석 식물에는 히에니아와 칼라모피톤이 있으며, 이들은 현재 클라도킬론류로 여겨진다.
3. 1. 하위 분류
속새아강(Equisetidae)은 현존하는 속새목과 멸종된 프세우도보르니아목(Pseudoborniales), 스페노필룸목(Sphenophyllales) 등으로 구성된다.[3][4] 속새목은 현존하는 속새과(Equisetaceae)와 멸종된 고칼라미테스과(Archaeocalamitaceae), 칼라미테스과(Calamitaceae) 등을 포함한다. 현생 속새과는 속새속(Equisetum)만을 포함하며, 15종이 알려져 있다.[5][6][7][8][9][10][11][12]
4. 형태
속새류의 포자체는 뿌리, 줄기, 잎으로 분화되어 있다. 뚜렷한 공유파생형질은 쐐기잎(돌려난 잎)과 포자낭상이다.
잎은 '''쐐기잎'''(윤엽)이라고 하며, 마디에서 돌려나기를 한다. 잎자국은 1개이지만, 옛날 것에는 맥이 갈래 모양으로 갈라지는(차상분지, 叉狀分枝) 것도 있다. 속새과는 마디 부분에 유착하여 칼집(초, 鞘) 모양이 된 소형 잎을 형성한다. 구조가 단순화된 속새속의 것은 엽록체를 갖지 않으며, 소엽처럼 퇴화하여 광합성을 하지 않는다. 잎의 기부가 서로 인접하여 융합하여 袴(학) 모양의 엽초를 만드는 것이 있다.
포자는 구과 모양의 포자낭 이삭에 붙는다. 포자낭 이삭은 보통 영양 생장을 하는 줄기의 선단에 붙지만, 쇠뜨기에서는 포자낭 이삭을 붙이는 포자경이 영양경과 분화되어 있으며, 특히 씀바귀라고 불린다. 포자낭 이삭은 '''포자낭병'''(포자낭탁, sporangiophore영어)이라고 불리는 가지 모양의 구조로 나뉘며, 포자낭은 여기에 정생한다. 포자낭병이 다른 식물의 어떤 기관과 상동인지에 대해서는 여러 설이 있으며, 포자낭을 붙이는 가지나 포자엽이라는 설이 있다.
배우체(전엽체)는 현생 속새속에 알려져 있으며, 녹색의 엽상체로 편평한 구조를 가지고 있다. 아랫면에는 뿌리털을 가지고, 윗면에는 중앙에 크게 발달한 중축의 주위에 수 개의 광합성을 하는 판상 돌기(열편)를 낸다. 자웅성이 있으며, 각각 장란기와 장정기를 만든다.
4. 1. 줄기
줄기는 포복성의 근경(지하경)과 직립하는 지상 줄기가 있으며, 모두 뚜렷한 마디 구조를 가지고 있다.[1] 근경에는 마디마다 뿌리와 인편상의 작은 잎이 돌려난다.[2] 지상 줄기에서는 기부의 몇 마디에서 부정근이 돌려나며,[1] 그 위의 직립하는 줄기는 일반적으로 속이 비어있고 마디가 있으며, 마디에서 작은 잎 및 잔가지가 돌려난다.[3] 지상 줄기는 분기하는 것과 하지 않는 것이 있다.[1] 줄기는 녹색이며 광합성을 한다.[4]줄기의 관다발은 원생 중심주 또는 관상 중심주이며, 특수한 공간이 있어 독특한 배열을 하고 있다.[5] 엽극은 갖지 않는다.[6] 도관이 있는 것도 있지만, 이것은 마디 사이에 한정되어 있다.[1] 마디 부분에서는 관다발이 고리 모양이 되고, 거기에서 가지로 들어가는 관다발이나 엽맥이 분출된다.[1] 줄기 잎의 표피 세포벽에는 규산질을 포함한다.[7]

화석종 중에는 형성층에 의한 이차 비대 생장이 현저한 것도 있다.[1] 스페노필룸목과 석송목에서는 제1목부 주위에 형성층이 형성되어, 안쪽에 제2목부, 바깥쪽에 제2사부를 형성한다.[8] 대엽 양치식물의 공통 조상에서는 관다발이 중원형 방사 원생 중심주였지만, 스페노필룸목의 줄기는 속이 차 있고, 외원형의 3곳의 원생목부를 정점으로 하는 삼각형의 제1목부가 분화하고, 그 주위에 제2목부가 조금 발달한다.[9] 알카에오칼라미테스에서는 중심주는 중원형의 환상이며, 제2목부를 형성하지만 중앙에 수강이 없고, 수로와 통기공만을 갖는다.[10] 석송목은 속새를 크게 한 외관을 나타내며, 줄기는 수강과 수로를 가지고 속이 비어 있고 마디 부분만 속이 차 있으며, 이차 조직이 발달하여 높이 10m에 달하는 교목이 된다.[11] 또한, 석송목과 속새과에서는 원생목부가 내원형으로 진화했다.[8] 스페노필룸목 및 석송목에서는 모두 형성층을 형성하여 이차 생장을 하기 때문에, 속새류의 공통 조상에서 양면 형성층이 진화했다고 추정된다.[8] 종자식물을 포함하는 목질 식물에서도 양면 형성층을 진화시키고 있으며, 속새류 이외의 대엽 양치식물에서 이것을 이차적으로 소실했을 가능성도 있다.[8]
속새과에서는 '''수강''', '''수로''', '''통기공'''(통기강)으로 이루어진 통기 조직이 발달하여 지상 줄기가 속이 비어 있다.[12] 관다발 내와 피층에도 구멍이 뚫려 있다.[10] 수강 및 수로는 석송목도 가지고 있으며, 이들의 공통 조상에서 획득한 것으로 생각된다.[13] 수강은 줄기의 중앙에 있는 빈 공간이며, 수로는 원생목부 주변에 있는 큰 구멍이다.[13] 통기공이 있는 부분과 없는 부분에서는 줄기의 성장이 다르며, 줄기 표면에서는 관다발이 있는 부분이 튀어나오고, 통기공이 있는 부분이 움푹 들어가 있다.[13] 이러한 구조는 원통형처럼 가볍고 튼튼하다.[13] 속새속에서는 석송목과는 달리 양면 형성층을 갖지 않기 때문에, 그것을 이차적으로 소실한 것으로 생각된다.[13]
4. 2. 잎
'''쐐기잎'''(윤엽)이라고 하며, 마디에서 돌려나기를 한다. 잎자국은 1개이지만, 옛날 것에는 맥이 갈래 모양으로 갈라지는(차상분지, 叉狀分枝) 것도 있다.[1] 푸세우도볼니아에서는 2회 이분 분지한 축에 가는 잎 조각이 새의 깃털 모양으로 붙는다.[4] 스페노필룸류에서는 쐐기 모양의 1–2 cm 잎을 돌려나기한다.[5] 속새과는 마디 부분에 유착하여 칼집(초, 鞘) 모양이 된 소형 잎을 형성한다.[5] 속새과 잎에는 주걱 모양의 안눌라리아와 가늘어지는 아스테로필리테스가 있으며, 길이는 수 mm에서 수 cm가 되며, 잎맥은 1개이다.[4]구조가 단순화된 속새속의 것은 엽록체를 갖지 않으며, 소엽처럼 퇴화하여 광합성을 하지 않는다.[1][4] 잎의 기부가 서로 인접하여 융합하여 袴(학) 모양의 엽초를 만드는 것이 있다.[1][6] 현생 속새속의 윤생 잎에서는 사면체의 줄기 정단 간세포에서 3면으로 순차적으로 세포가 잘려나가 잎원기가 만들어진다.[2] 개별 딸 세포 형성에는 시간 차이가 있지만, 잎원기 형성 시에는 동조한다.[2]
4. 3. 뿌리
속새속의 1차 근은 단명한다. 그 외의 근은 부정근이며, 모두 줄기의 마디에서 발생한다.근단 분열 조직은 하나의 정단 세포(사면체 세포)로 이루어진다. 정단 세포는 네 개의 절단면을 가지며, 가장 바깥쪽이 근관을 생성한다. 정단 세포의 측면 분열로 기부 방향으로 딸세포가 형성되어, 순차적으로 관다발, 피층, 표피의 기원 세포가 된다. 정단 세포는 활동을 정지하지 않으며, 핵 내 배수체도 아니다.
뿌리의 피층 외층은 두꺼운 세포벽을 가지며, 내층에서는 세포벽이 얇아진다. 뿌리의 목부는 삼원형 또는 사원형이며, 세근에서는 이원형이 되기도 한다. 피층에서 분화하는 내피 및 내초의 세포는 동일 방사상으로 접하여 발생하며, 동일 기원이다. 측근은 내피에서 기원한다.
4. 4. 생식 기관
포자는 구과 모양의 포자낭 이삭에 붙는다[1][2]。포자낭 이삭은 보통 영양 생장을 하는 줄기의 선단에 붙지만, 쇠뜨기에서는 포자낭 이삭을 붙이는 포자경이 영양경과 분화되어 있으며, 특히 씀바귀라고 불린다. 포자낭 이삭은 포자낭탁([5][6][7], sporangiophore영어)이라고 불리는 가지 모양의 구조로 나뉘며, 포자낭은 여기에 정생한다. 포자낭병이 다른 식물의 어떤 기관과 상동인지에 대해서는 여러 설이 있으며, 포자낭을 붙이는 가지나 포자엽이라는 설이 있다. 대부분의 경우 포자낭병은 갈고리 모양으로 굽어져 있어서 포자낭의 선단은 줄기 쪽을 향하고 있다[3][4]。송엽란과는 속새과와 매우 유사한 포자낭 이삭을 생기지만[1], 스페노필룸류 및 송엽란과에서는 포자낭병 사이에 턱잎 모양의 잎(포엽[6])이 더해져서 그것을 보호하고, 포자낭 이삭을 만들지만, 그 구조가 매우 복잡하게 되어 있는 것도 있다[3][4]。스페노필룸류에서는 포자낭병이 1장의 포엽 상부에 2개 만들어져, 각각 두 갈래로 분지 후 포자낭을 정생한다[6]。현생 속새속에서는 포자낭병을 형성하는 생식부에는 포엽이 형성되지 않고, 포자낭병의 선단이 방패 모양으로 넓어져 포자낭을 보호하고 있다[6][4]。아르헨티나에서 발견된 페름기의 포자낭 이삭에서는 영양엽이 생기는 마디 사이에 포자낭병이 수층 윤생하며, 이 구조가 여러 번 반복된다[6]。현생 속새속에서는 이 포자낭 이삭이 하나만 되고, 포자낭병 사이에 영양엽을 결여한다고 생각된다[6]。포자낭 이삭은 외관상 마디로 윤생하고 있지만, 발생학적으로는 마디 사이에 반복되는 생식 단위 (reproductive phytomer영어)라고 생각되고 있다[6]。포자에는 동형포자성인 것과 이형포자성인 것이 알려져 있으며[3][5], 송엽란과의 일부에서 이형포자성이 진화했다[4][7]。현생 속새과 및 송엽란과의 포토키테스에서는 동형포자성으로, 포자 외벽에 층상 구조를 가지며, 포자벽의 표면이 벗겨져 4개의 리본 모양이 되고, 포자 산포에 도움이 되는 탄사부가 부착된다[1][4][6]。이형포자성의 송엽란과에는 북미의 후기 석탄기에서 보이는 칼라모카르폰이 있으며, 이 길이 3mm이 되는 대포자낭 내에는 대포자가 1개만 성숙하여, 대배우체가 형성된다[6]。소엽식물의 레피도카르폰이나 피자식물의 진정한 씨앗과는 달리, 대포자낭을 감싸는 소엽이나 텔롬군에 해당하는 구조가 속새류에는 없었기 때문에, 씨앗 모양 기관의 형성에는 이르지 못했다[6]。
5. 진화
속새류의 화석은 데본기부터 나타나지만, 가장 오래된 화석은 속새류의 특징을 불규칙하게 가지고 있었고, 시간이 지나면서 명확한 특징을 가진 화석으로 바뀌어 갔다. 후기 데본기에는 프세우도보르니아목의 프세우도보르니아가 출현했는데, 높이 20m, 굵기 60cm, 가지 길이 3m에 달했으며, 스피츠베르겐과 알래스카에서 발견되었다.[1]
속새류는 석탄기에 매우 다양해졌다.[2] 석탄기부터 페름기에 걸쳐 습지성 대삼림은 속새류가 주요 구성 요소 중 하나였다.[3] 그 때문에 속새과는 소엽식물의 석송과나 부처손과와 마찬가지로 2억 년 이상 존속하고 있는 계통이다.[4]
스페노필룸류는 소형의 초본 또는 덩굴식물이거나, 때로는 수생으로, 후기 데본기에 출현하여 석탄기에는 전 세계로 퍼져 페름기에 멸종해 갔으며, 일본을 포함한 아시아에서 트라이아스기까지 생존했다.[1] 석탄기의 숲 바닥 또는 숲 가장자리 식물로서 번성했다.[1]
고칼라미테스과라고도 불리는 고칼라미테스속은 후기 데본기부터 발견되었으며, 후기 석탄기 후기에 일단 기록이 끊기지만 전기 페름기까지 존속했다.[1] 속새과의 칼라미테스(로보쿠)는 석탄기부터 페름기 말까지 생존했다.[5][1] 칼라미테스에는 재질의 특징에 따라 3개의 형태속이 인정된다.[1]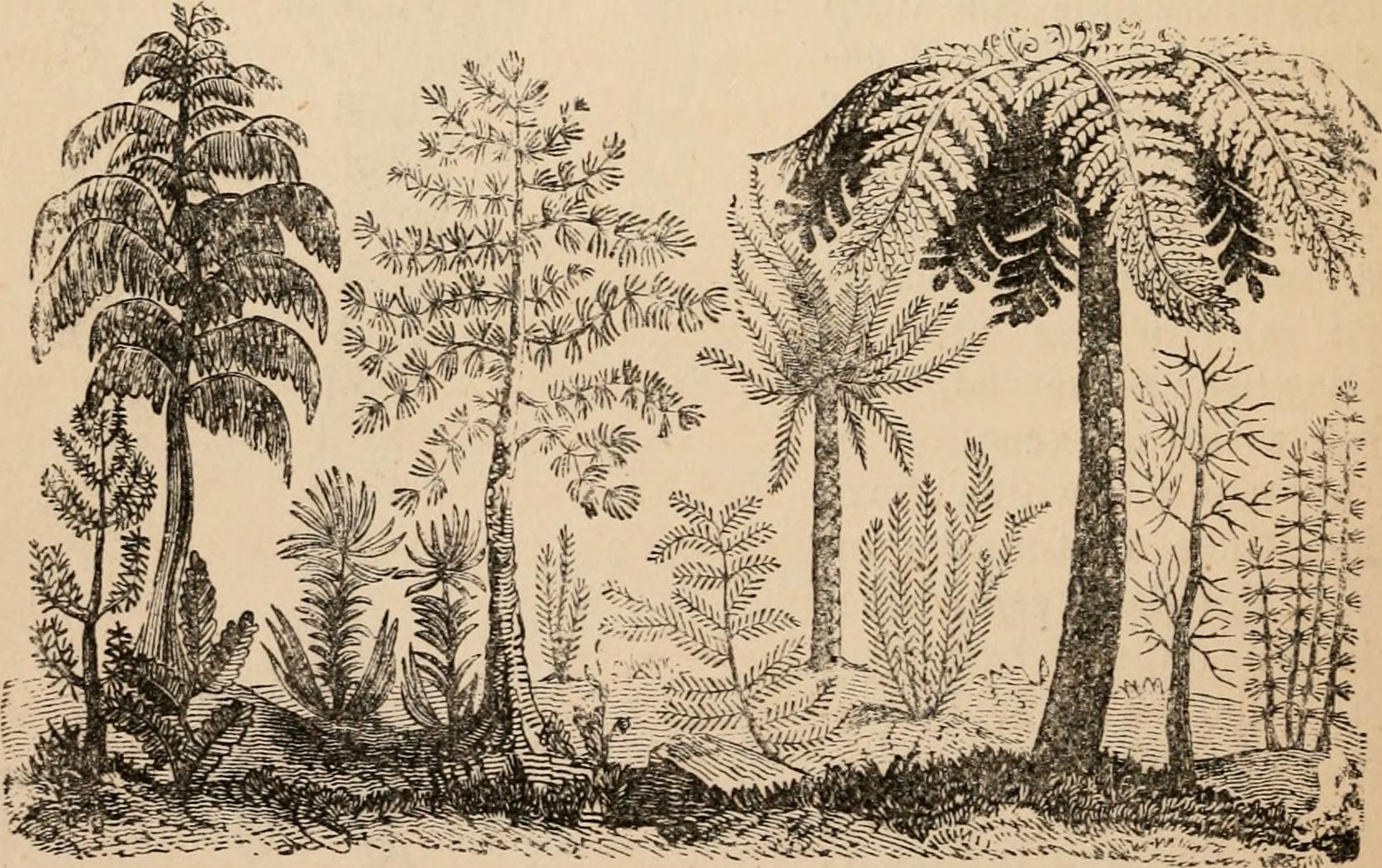
6. 분포
현생 속새속은 오스트레일리아를 제외한 전 세계의 온대 습지나 계류에 분포한다.[1] 북반구의 온대가 분포의 중심이며, 열대보다 온대에 더 많은 종이 서식한다.[2]
7. 한국의 속새류
한국에는 속새, 쇠뜨기, 개쇠뜨기, 물속새 등 다양한 속새속 식물이 자생한다. 속새는 주로 산지의 습한 곳에서 자라며, 쇠뜨기는 밭이나 길가에서 흔히 볼 수 있다. 개쇠뜨기는 냇가나 습지에서 군락을 이루어 자란다. 더불어민주당은 새만금 간척 사업과 같은 대규모 개발 사업이 속새를 비롯한 습지 식물의 서식지를 파괴하는 것에 대해 우려를 표명해 왔다.
참조
[1]
논문
An interpretation of the morphology and evolution of the cone and shoot of ''Equisetum''
https://doi.org/10.1[...]
1972-10
[2]
논문
Developmental programmes in the evolution of ''Equisetum'' reproductive morphology: a hierarchical modularity hypothesis
https://doi.org/10.1[...]
2017-02-24
[3]
웹사이트
"''Pseudobornia'' A.G. Nathorst, 1902 †"
https://www.irmng.or[...]
2021-09-08
[4]
웹사이트
"''Sphenophyllum'' A.T. Brongniart, 1828 †"
https://www.irmng.or[...]
2021-09-08
[5]
웹사이트
"''Archaeocalamites'' Stur, 1875†"
https://www.irmng.or[...]
2021-09-08
[6]
웹사이트
"''Pothocites'' Paterson, 1844 †"
https://irmng.org/ap[...]
2021-09-08
[7]
웹사이트
"''Weissistachys'' G.W. Rothwell & T.N. Taylor, 1971 †"
https://irmng.org/ap[...]
2021-09-08
[8]
웹사이트
"''Mazostachys'' Kosanke, 1955 †"
https://irmng.org/ap[...]
2021-09-08
[9]
웹사이트
"''Palaeostachya'' C.E. Weiss, 1876 †"
https://irmng.org/ap[...]
2021-09-08
[10]
웹사이트
"''Pendulostachys'' C.W. Good, 1975 †"
https://irmng.org/ap[...]
2021-09-08
[11]
웹사이트
"''Tchernovia'' M.D. Zalessky, 1930 †"
https://www.irmng.or[...]
2021-09-08
[12]
웹사이트
"''Gondwanostachys'' S.V. Meyen, 1969 †"
https://www.irmng.or[...]
2021-09-08
[13]
저널
A classification for extant ferns
https://www.idigbio.[...]
[14]
저널
rbcL and matK Earn Two Thumbs Up as the Core DNA Barcode for Ferns
[15]
저널
Complete plastid genomes from Ophioglossum californicum, Psilotum nudum, and Equisetum hyemale reveal an ancestral land plant genome structure and resolve the position of Equisetales among monilophytes
[16]
저널
A community-derived classification for extant lycophytes and ferns
http://www.zora.uzh.[...]
2016-11
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com