개속(Canis)은 개, 늑대, 코요테, 자칼 등을 포함하는 육식 동물의 속으로, 1758년 칼 폰 린네에 의해 명명되었다. 개속에 속하는 종들은 78개의 염색체를 공유하며 잡종이 가능하다. 2019년 DNA 증거에 따라 줄무늬자칼과 검은등자칼은 털자칼속으로 분류되었다. 개속의 종은 진화 과정을 거쳐 다양한 종으로 분화되었으며, 늑대와 코요테는 북아메리카에서, 아프리카황금늑대는 아프리카에서 발견된다. 개과 동물은 일부일처제 경향을 보이며, 늑대와 자칼은 무리 생활을 한다.
더 읽어볼만한 페이지
개속 - 검은등자칼 검은등자칼은 개과 동물로, 케이프자칼과 동아프리카자칼의 두 아종으로 나뉘며, 가로줄무늬자칼과 함께 계통 분류상 개과 내에서 비교적 기저에 위치한다.
개속 - 에티오피아늑대 에티오피아늑대는 에티오피아 고원에 서식하며 붉은 털과 긴 다리를 가진 개과 동물로, 설치류를 주식으로 하는 주행성 멸종 위기종이다.
측계통군 - 맹금류 맹금류는 날카로운 발톱과 부리를 가진 사나운 새를 통칭하며, 생태계 최상위 포식자로서 뛰어난 시력과 비행 능력을 갖추고 있으며, 수리, 매, 독수리, 올빼미 등이 대표적이고, 서식지 파괴 등의 위협에 직면해 있다.
측계통군 - 돌고래 돌고래는 이빨고래류에 속하는 해양 포유류의 총칭으로, 다양한 종이 존재하며 높은 지능과 사회성을 가지지만, 서식지 파괴, 환경 오염 등으로 멸종 위기에 처해 있다.
칼 폰 린네가 명명한 분류군 - 벵골호랑이 벵골호랑이는 인도아대륙에서 서식하는 호랑이 아종으로, 노란색~주황색 털과 검은 줄무늬를 가지며, 멧돼지, 사슴 등 우제류를 먹이로 하며, 서식지 파괴와 밀렵으로 인해 제한적인 지역에서 서식한다.
칼 폰 린네가 명명한 분류군 - 잉어 잉어는 동아시아에 널리 분포하는 민물고기로, 하천 중류 이하의 진흙 바닥을 선호하며, 길조의 상징으로 여겨지며, 다양한 요리 재료로 활용되며, 기생충 감염 위험에 주의해야 한다.
개속(''Canis'')은 1758년 칼 폰 린네(Carl Linnaeus)가 『자연의 체계 제10판』에서 처음 명명한 분류군으로, 개, 늑대, 코요테, 자칼 등 개와 유사한 개과 동물들을 포함한다. 이 속에 속하는 모든 종들은 계통 발생학적으로 서로 밀접하게 연관되어 있으며, 공통적으로 78개의 염색체를 가지고 있어 서로 잡종을 만들 수 있다. 1926년 국제동물명명규약위원회(ICZN)는 개속(''Canis'')을 공식 동물 명칭 목록에 포함시켰고, 1955년에는 개(''Canis familiaris'')를 개속의 모식종으로 지정했다.
개속 내 종들의 분류는 지속적으로 연구되고 있으며, 특히 DNA 분석 기술의 발달로 기존 분류 체계에 변화가 있었다. 예를 들어, 과거 형태학적 특징에 기반하여 아속 수준의 분류가 제안되기도 했으나,[16] 최근의 유전적 증거는 다른 분류를 지지한다. 2019년 세계자연보전연맹(IUCN)/SSC 개과 전문가 그룹 워크숍에서는 DNA 증거를 바탕으로, 줄무늬자칼(과거 ''Canis adustus'')과 검은등자칼(과거 ''Canis mesomelas'')을 개속(''Canis'')과는 별개의 단계통 분류군을 형성하는 털자칼속(Lupulella)으로 재분류할 것을 권장했다.[18] 또한, 2015년 분자 계통 분석 결과에 따라 기존 금색자칼( ''Canis aureus'')의 아프리카 개체군은 유전적으로 뚜렷이 구분되는 별개의 종인 아프리카황금늑대( ''Canis lupaster'')로 분리되었다.[19]
일반적으로 '늑대'라는 명칭은 회색늑대( ''Canis lupus''), 에티오피아늑대( ''Canis simensis''), 동부늑대( ''Canis lycaon''), 아프리카황금늑대( ''Canis lupaster'') 등 비교적 큰 개속 동물들을 지칭하는 데 사용된다.[9] 이보다 작은 종들은 아메리카 대륙에서는 코요테(coyote)로, 다른 지역에서는 자칼(jackal)로 불리는 경향이 있다.[10] '자칼'이라는 용어는 특히 유라시아 대륙에 분포하는 금색자칼( ''Canis aureus'')에 주로 적용된다.[11]
한편, 개(''Canis familiaris'')와 회색늑대( ''Canis lupus'')의 분류학적 관계에 대한 논의 끝에, 2003년 국제동물명명규약위원회(ICZN)는 개의 학명은 ''C. familiaris''로 유지하고, 야생종인 회색늑대의 학명은 ''C. lupus''를 계속 사용하기로 결정했다.[20]
2. 1. 진화
화석 기록에 따르면 고양이형류와 개형류는 약 4,300만 년 전(시신세)에 식육목 내에서 갈라져 나왔다. 개형류에는 여우와 비슷한 속인 ''렙토키온''이 포함되는데, 이 속의 다양한 종들은 약 2,400만 년 전(점신세)부터 존재했으며, 약 1,190만 년 전(마이오세 후기)에 ''여우속''(여우)과 개족(Canini, 개)으로 분화되었다. 자칼 크기의 ''유키온''은 약 1,000만 년 전부터 북아메리카에 서식했으며, 약 600만~500만 년 전(플리오세 초기) 무렵에는 코요테와 비슷한 ''Eucyon davisi''가 유라시아로 진출했다. 북아메리카에서 유라시아로 이주한 ''유키온'', ''여우속'', 그리고 ''너구리속''은 후기 마이오세와 초기 플리오세 동안 중소형 포식자였지만, 최상위 포식자는 아니었다.
신세계의 ''개속'' 개체군의 경우, 북아메리카의 ''유키온''은 초기 북아메리카 ''개속''으로 이어졌는데, 이는 약 600만 년 전 마이오세 후기에 미국 남서부와 멕시코에서 처음 등장했다. 약 500만 년 전에는 늑대와 코요테의 조상으로 여겨지는 더 큰 ''개속 르포파구스''가 같은 지역에 나타났다. 구세계에서는 약 500만 년 전 ''유키온''의 일부가 최초의 ''개속'' 구성원으로 진화했으며,[3] 이로써 개과 동물은 구북구 전역에서 우점 포식자가 되었다. 늑대 크기의 ''C. chihliensis''는 약 400만~300만 년 전(플리오세 중기) 무렵 중국 북부에 나타났다. 이후 약 180만 년 전(플라이스토세 초기)에 걸쳐 유라시아 전역에서 ''개속''의 진화가 폭발적으로 일어났는데, 이는 "늑대 사건"으로 불리며 맘모스 초원의 형성과 대륙 빙하 작용과 관련이 있다. 이 시기 ''개속''은 ''C. arnensis'', ''C. etruscus'', 그리고 ''C. falconeri'' 등의 형태로 유럽으로 퍼져나갔다.
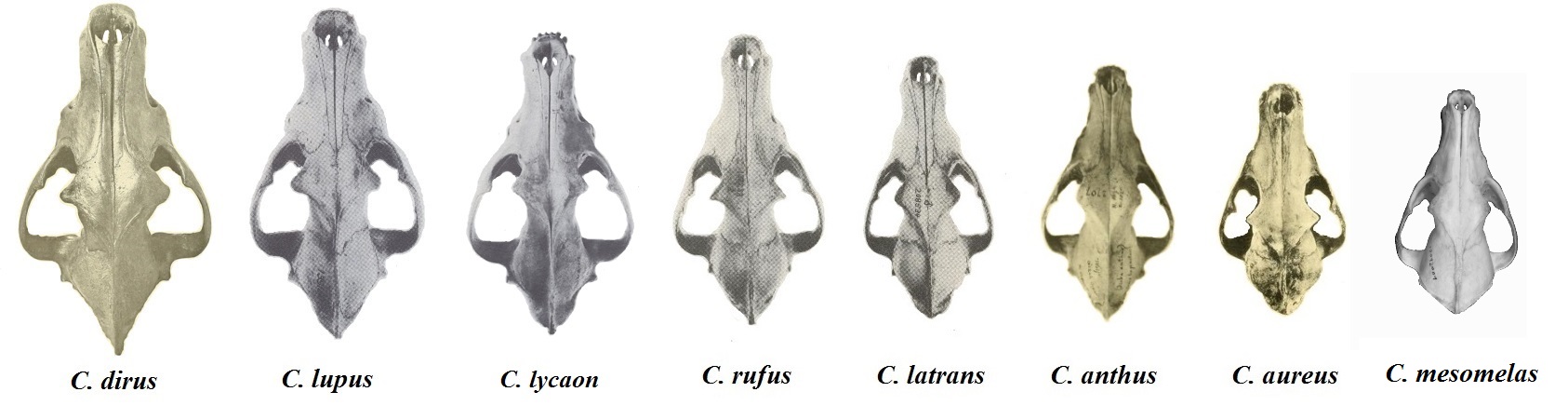
끔찍한 늑대 (''Aenocyon dirus''), 회색 늑대 (''C. lupus''), 동부 늑대 (''C. lycaon''), 붉은 늑대 (''C. rufus''), 코요테 (''C. latrans''), 아프리카 황금 늑대 (''C. lupaster''), 금색 자칼 (''C. aureus'') 및 검은 등 자칼 (''Lupulella mesomelas'')의 두개골
2021년 끔찍한 늑대 (''Aenocyon dirus'')에 대한 유전자 연구 결과는 기존의 분류에 큰 변화를 가져왔다. 이전에는 ''개속''의 일원으로 여겨졌던 끔찍한 늑대가 실제로는 신세계 고유의 고대 개과 동물 계통의 마지막 구성원이며, ''개속''이 등장하기 이전에 갈라져 나와 마이오세 이후 ''개속''과의 유전자 혼입 없이 독립적인 계통을 유지해왔다는 사실이 밝혀졌다. 이 연구는 신세계의 신생대 개과 동물인 ''Canis armbrusteri''와 ''개속 에드워디'' 역시 ''개속''과 유사한 외형으로 수렴 진화한 독립적인 끔찍한 늑대 계통일 가능성을 제시했다. 회색 늑대와 코요테와 같은 진정한 ''개속'' 구성원은 후기 플라이스토세에 신세계에 도착했을 가능성이 높으며, 이들의 유연한 식성과 다른 개과 동물과의 잡종화 능력 덕분에 끔찍한 늑대와 달리 플라이스토세 거대동물군 멸종에서 살아남을 수 있었을 것으로 추정된다.[3]
''크세노키온''(Xenocyon)은 ''개속''의 멸종된 아속으로 간주된다. ''개속'' 무리의 다양성은 초기 플라이스토세 말기에서 중기 플라이스토세까지 감소하여, 유라시아에서는 작은 늑대인 ''C. mosbachensis–C. variabilis'' 무리와 대형의 극단적 육식성 ''개속 (크세노키온) 리카오노이데스''만 남게 되었다. 극단적 육식성 동물인 ''크세노키온''은 현생 들개(Cuon alpinus)와 아프리카들개(Lycaon pictus)의 조상으로 이어진다.
리처드 H. 테드포드(Richard H. Tedford)는 개속(Canis)이 상대적으로 큰 송곳니와 과육식성(hypercarnivory)에 대한 치아 적응(m1–m2의 메타코니드와 엔토코니드가 작거나 없음, M1–M2의 하이포콘이 작음, M1–M2의 설측 치대(lingual cingulum)가 약함, M2와 m2가 작고 단근일 수 있음, m3가 작거나 없음, 넓은 구개)이 부족하기 때문에 붉은늑대속(Cuon), 아프리카들개속(Lycaon), 젠노키온속(Xenocyon)에 비해 원시적인 속이라고 평가했다.
현생 개속 종들의 계통 발생 관계는 DNA 연구를 통해 밝혀지고 있다. Lindblad-Toh 등의 2005년 연구와 후속 연구[2]를 종합한 결과에 따르면, 개속 내 종들의 분화 순서는 다음과 같이 추정된다.
과거에는 줄무늬자칼(Lupulella adusta)과 검은등자칼(Lupulella mesomelas)도 개속으로 분류되었으나, 2019년 세계자연보전연맹(IUCN)/SSC 개과 전문가 그룹 워크숍에서는 이 두 종이 개속(Canis)/붉은늑대속(Cuon)/아프리카들개속(Lycaon) 분기군의 외부에 위치하는 별도의 단계통군을 형성한다는 DNA 증거에 따라, 1906년 Hilzheimer가 명명한 털자칼속(Lupulella)으로 재분류할 것을 권장했다. 따라서 현재는 털자칼속 줄무늬자칼(Lupulella adusta)과 털자칼속 검은등자칼(Lupulella mesomelas)로 분류된다.[18] 또한, 2015년 미토콘드리아 DNA 및 핵 DNA 분석 결과에 따라 기존 금색자칼의 아프리카 개체군은 아프리카황금늑대(Canis lupaster)라는 별도의 종으로 분리되었다.[19]
한때 회색늑대의 학명을 개의 학명과 동일한 ''Canis familiaris''로 통일하려는 논의도 있었으나, 2003년 국제동물명명규약위원회(ICZN)의 결정에 따라 개의 학명은 ''C. familiaris''로 유지하고, 야생종인 회색늑대의 학명은 ''C. lupus''를 계속 사용하는 것으로 정리되었다.[20]
2. 2. 하위 종
개속(''Canis'')은 개, 늑대, 코요테, 자칼 등을 포함하는 개과의 한 속이다. 개속에 속하는 모든 종은 계통 발생학적으로 밀접한 관련이 있으며, 78개의 염색체를 가지고 있어 잡종화가 가능하다. 국제동물명명규약위원회(ICZN)는 1955년 개(''Canis familiaris'')를 개속의 모식종으로 지정했다.
리처드 H. 테드포드는 개속이 붉은늑대속(Cuon), 아프리카들개속(Lycaon), 젠노키온속(Xenocyon)에 비해 상대적으로 큰 송곳니와 과육식성(hypercarnivory)에 대한 치아 적응이 부족하여 원시적인 속이라고 평가했다. 구체적으로는 m1–m2 metaconid와 entoconid가 작거나 없고, M1–M2 hypocone가 작으며, M1–M2 lingual cingulum이 약하고, M2와 m2가 작고 단근일 수 있으며, m3가 작거나 없고, 구개가 넓은 특징을 근거로 들었다.
분자생물학적 연구 결과에 따라 속 분류에 변동이 있어왔다. 다음은 일반적으로 인정되는 개속의 현존하는 주요 종 목록이다.[15][16][19]
DNA 계통 발생 연구[2]에 따르면, 개속 내 종들의 계통 관계는 복잡하며 지속적으로 연구되고 있다. 예를 들어, 코요테는 붉은늑대 및 동부늑대와 가까운 관계를 보이며, 이들은 다시 회색늑대 및 개와 자매군을 형성한다. 아프리카황금늑대, 에티오피아늑대, 황금자칼 등은 이들보다 먼저 분기된 것으로 나타난다.
과거 개속으로 분류되었던 가로줄무늬자칼(과거 학명 ''Canis adustus'')과 검은등자칼(과거 학명 ''Canis mesomelas'')은 2019년 세계자연보전연맹(IUCN)/SSC 개과 전문가 그룹 워크숍에서 DNA 증거를 바탕으로 별도의 속인 털자칼속(Lupulella)으로 재분류할 것이 권장되었다. 이들은 개속(''Canis'')/붉은늑대속(Cuon)/아프리카들개속(Lycaon) 분기군 외부에 위치하는 단계통 분류군을 형성하는 것으로 밝혀졌다.[18]
또한, 2015년 발표된 미토콘드리아 DNA 및 핵 DNA 등의 분자 계통 분석 결과에 따라, 기존에 황금자칼(''Canis aureus'')의 아프리카 대륙 개체군으로 여겨지던 집단이 유전적으로 뚜렷이 구분되는 별개의 종인 아프리카황금늑대(''Canis lupaster'')로 분리되었다.[19]
한편, 회색늑대의 학명을 개와 동일하게 ''Canis familiaris''로 통합하자는 주장이 있었으나, 2003년 국제동물명명규약위원회(ICZN)의 강권에 의해 ''C. familiaris''는 개만을 가리키는 학명으로 하고, 야생종의 학명은 ''C. lupus''를 계속 사용하는 것이 인정되었다.[20]
3. 형태
암컷 회색늑대
수컷과 암컷 개과 동물 간의 외형적 차이는 거의 미미하다. 개과 동물은 일반적으로 일부일처제 쌍으로 생활한다. 늑대, 들개, 코요테, 그리고 자칼은 번식쌍과 그들의 새끼를 포함하는 무리로 생활하며, 늑대는 확장된 가족 무리를 이루기도 한다. 아프리카들개, 들개, 회색늑대는 자신보다 큰 먹이를 사냥할 때 앞다리로 먹이를 붙잡을 수 없기 때문에 강한 턱에 의존한다. 이들은 우두머리 쌍과 현재 및 이전 해의 새끼들로 구성된 무리로 협력하여 사냥한다. 사회성을 가진 포유류 포식자는 포식자 무리의 총 몸무게와 비슷한 체질량의 초식동물을 사냥하는 경향이 있다. 회색늑대는 큰 먹이 중에서도 특히 취약한 개체를 노리는 것을 전문으로 하며, 숲에 사는 늑대 무리는 500kg 무게의 무스를 사냥할 수도 있다.
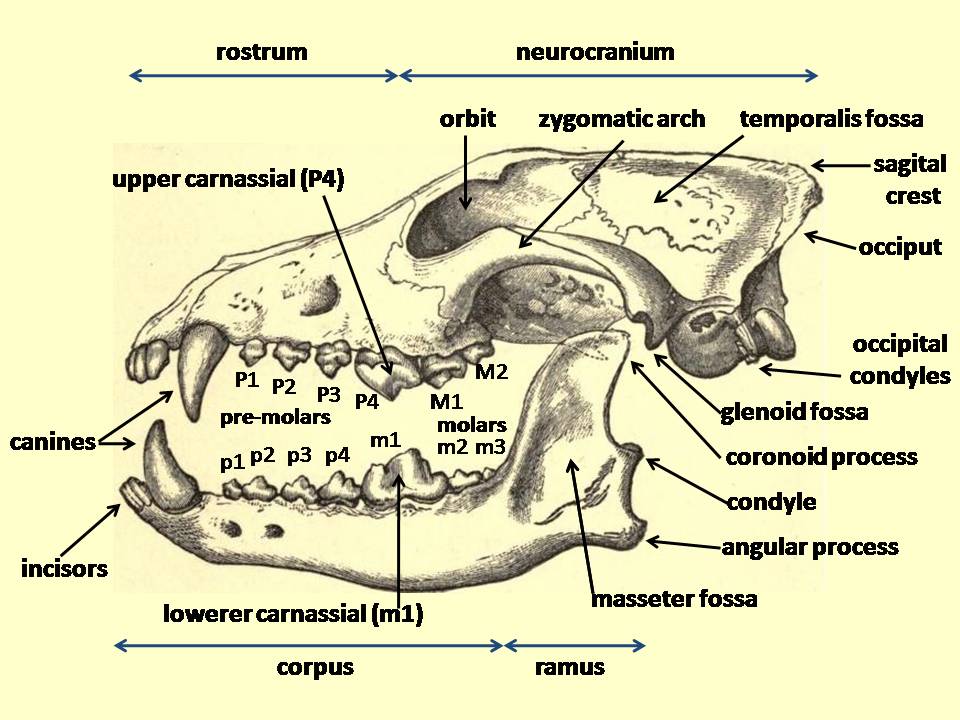
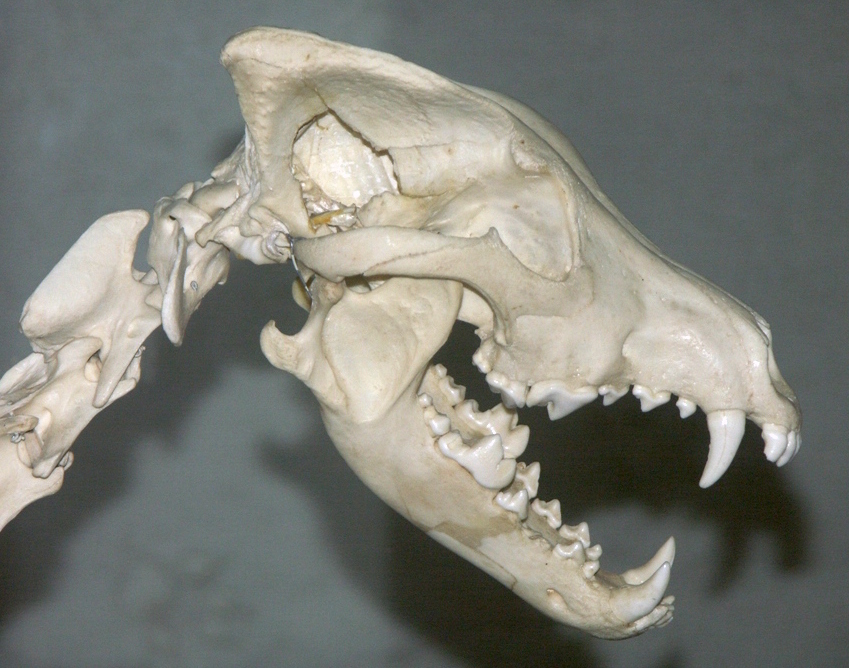
치열은 입 안의 치아 배열을 의미하며, 위턱 치아의 치식은 앞니를 나타내는 대문자 I, 송곳니를 나타내는 C, 작은어금니를 나타내는 P, 큰어금니를 나타내는 M을 사용하고, 아래턱 치아는 소문자 i, c, p, m을 사용한다. 치아는 입의 한쪽을 기준으로 앞에서부터 뒤로 번호가 매겨진다. 육식동물에서 위턱 작은어금니 P4와 아래턱 큰어금니 m1은 가위처럼 작용하여 먹이의 근육과 힘줄을 자르는 육치를 형성한다.
개과 동물은 위턱 네 번째 작은어금니 P4(위턱 육치)를 제외하고는 작은어금니를 잘라서 부수는 데 사용한다. 위턱 네 번째 작은어금니는 자르는 데만 사용된다. 아래턱 첫 번째 큰어금니 m1(아래턱 육치)은 개과 동물의 식단 적응에 따라 자르고 가는 데 사용하도록 진화했다. 아래턱 육치에서 삼각돌기는 자르는 데 사용되고 뒤쪽돌기는 가는 데 사용된다. 삼각돌기와 뒤쪽돌기의 비율은 육식동물의 식단 습관을 나타내며, 삼각돌기가 더 크면 초육식동물을, 뒤쪽돌기가 더 크면 더 잡식성 식단을 나타낸다. 변이가 적기 때문에 아래턱 육치의 길이는 육식동물의 몸집을 추정하는 데 사용된다.
살아있는 포유류 포식자와 화석 포유류 포식자 다수 표본의 송곳니 교합력을 체중으로 조정한 결과, 태반류 포유류의 경우 송곳니 교합력(체중 킬로그램당 뉴턴)은 멸종된 다이어울프(163)가 가장 컸으며, 현대 개과 동물 중에서는 자신보다 큰 동물을 자주 사냥하는 네 종류의 초육식동물, 즉 아프리카들개(142), 회색늑대(136), 붉은개(112), 딩고(108) 순이었다. 육치 교합력도 송곳니와 유사한 경향을 보였다. 포식자가 사냥할 수 있는 먹이의 최대 크기는 생체역학적 한계에 크게 영향을 받는다.
이빨의 기능을 보여주는 늑대의 치열
치아 파손은 육식동물의 먹이 섭취 과정에서 자주 발생한다. 육식동물에는 무리 사냥꾼과 독립 사냥꾼이 모두 포함된다. 독립 사냥꾼은 먹잇감을 제압하기 위해 송곳니로 강하게 무는 것에 의존하므로, 강한 하악골 결합부를 가진다. 반면, 여러 번 얕게 무는 무리 사냥꾼은 비교적 약한 하악골 결합부를 가진다. 따라서 연구자들은 화석 육식동물 표본의 하악골 결합부 강도를 통해 사냥 방식(무리 사냥 또는 독립 사냥)과 먹이 섭취 방식까지 추론할 수 있다. 개과 동물의 아래턱뼈는 육치 뒤쪽에서 지지되어, 육치 뒤의 어금니(M2, M3)로 뼈를 부술 수 있다. 한 연구에 따르면 현대 회색늑대와 붉은늑대 (''C. rufus'')는 다른 모든 현존 개과 동물 및 멸종된 다이어울프보다 아래턱뼈의 지지 구조가 더 크다. 이는 이들이 다른 개과 동물보다 뼈를 부수는 데 더 적합함을 시사한다.
9종의 현대 육식동물을 대상으로 한 연구에 따르면, 성체 4마리 중 1마리꼴로 치아 파손을 경험하며, 이 중 절반은 송곳니에서 발생한다. 파손 빈도가 가장 높은 동물은 뼈를 포함한 먹이 전체를 먹는 것으로 알려진 점박이하이에나였다. 파손 빈도가 가장 낮은 동물은 아프리카들개였다. 회색늑대는 이 두 종의 중간 정도였다. 뼈를 먹는 행위는 상대적으로 높고 예측 불가능한 힘을 치아에 가하므로, 우발적인 골절 위험을 높인다. 가장 흔하게 부러지는 이빨은 송곳니이고, 그 다음으로 작은어금니, 육치, 앞니 순이다. 송곳니는 그 모양과 기능 때문에 예측 불가능한 방향과 크기의 굽힘력을 받아 가장 부러지기 쉽다. 큰 먹잇감을 사냥하고 먹을 때 치아 골절 위험이 더 높다.
현존하는 회색늑대와 비교할 때, 멸종된 베링기아 늑대는 중간 정도에서 심하게 마모된 이빨을 가진 개체가 훨씬 많았고, 부러진 이빨 수도 더 많았다. 치아 골절 빈도는 북부 로키산맥 늑대 (''Canis lupus irremotus'')의 최소 2%에서 베링기아 늑대의 최대 11%까지 다양했다. 치열 전체의 골절 분포도 달라서, 베링기아 늑대는 앞니, 육치, 어금니의 골절 빈도가 훨씬 높았다. 점박이하이에나에서도 유사한 패턴이 관찰되는데, 이는 앞니와 육치의 골절 증가는 습관적인 뼈 섭취를 반영함을 시사한다. 뼈는 앞니로 갉아먹고, 육치와 어금니로 부순다.
4. 생태
개속(Canis)에는 여러 종이 포함되는데, 이 중 회색늑대( *Canis lupus*), 에티오피아늑대( *Canis simensis*), 동부늑대( *Canis lycaon*), 아프리카황금늑대( *Canis lupaster*) 네 종은 흔히 "늑대"라고 불린다.[9] 이들보다 몸집이 작은 종들은 아메리카 대륙에서는 코요테(coyote)로, 다른 지역에서는 자칼(jackal)로 불린다.[10] 코요테와 자칼이 회색늑대와의 관계보다 반드시 서로 더 가까운 것은 아니지만, 같은 개속(Canis)에 속하므로 여우, 갈기늑대(maned wolf) 또는 개속에 속하지 않는 다른 개과 동물보다는 늑대나 개와 유전적으로 더 가깝다. "자칼"이라는 명칭은 아시아 남서부와 중남부, 유럽의 발칸반도 전역에서 발견되는 황금자칼( *Canis aureus*)에도 적용된다.[11]
4. 1. 짝짓기 행동
수컷과 암컷 개과 동물 간의 외형적 차이는 거의 미미하다. 개과 동물은 일반적으로 일부일처제 쌍으로 생활한다. 늑대, 들개, 코요테, 그리고 자칼은 번식쌍과 그들의 새끼를 포함하는 무리를 이루어 생활한다. 늑대는 확장된 가족 무리로 생활할 수도 있다. 아프리카 들개, 들개, 회색늑대는 자신보다 큰 먹이를 사냥할 때 앞다리로 먹이를 붙잡을 수 없기 때문에 턱에 의존한다. 이들은 알파 쌍과 현재 및 이전 해의 새끼들로 구성된 무리로 협력하여 사냥한다. 사회성 포유류 포식자는 포식자 무리의 총 질량과 비슷한 체질량을 가진 초식동물을 사냥하는 경향이 있다. 회색늑대는 큰 먹이 중에서도 취약한 개체를 전문적으로 사냥하며, 숲늑대 무리는 500kg 무게의 무스를 사냥할 수도 있다.
개속(Canis)에는 다양한 종이 있으며, 이들의 짝짓기 행동 양식은 종에 따라 다양하게 나타난다.[4] 2017년 연구에 따르면, 일부 개과 동물 종에서는 암컷이 자신의 발정 상태를 이용하여 먹이 자원을 얻는 것으로 나타났다. 이 연구는 늑대와 개를 대상으로 진행되었는데, 늑대는 일반적으로 일부일처제이며 짝을 이루어 생활하는 반면, 자유롭게 돌아다니는 개는 다양한 개체와 짝짓기하는 다부일처제 경향을 보인다. 연구 결과, 두 종 모두 암컷은 발정기일 때 먹이에 대한 접근을 더 많이 시도했으며, 먹이 자원을 독점하는 데 더 성공적이었다. 번식기가 아닐 때는 이러한 노력이 지속적이지도 성공적이지도 않았다. 이는 '먹이를 위한 섹스 가설'(food-for-sex hypothesis)이 개과 동물의 먹이 공유에 영향을 미치며 암컷에게 직접적인 이점으로 작용할 수 있음을 시사한다.[4]
자유로운 개를 대상으로 한 2014년 연구에서는 사회적 요인이 짝짓기 쌍 결정에 중요한 역할을 한다는 것이 밝혀졌다.[5] 이 연구는 개의 생식에 대한 사회적 조절을 조사했는데, 발정기의 암컷은 우점적인 수컷을 찾아다녔고, 뛰어난 지도자로 보이는 우점적인 수컷과 짝짓기를 할 가능성이 더 높았다. 암컷은 순종적인 수컷을 거부하는 경향이 있었다. 또한, 사회적 지위가 높은 암컷이 있을 경우 수컷 간 경쟁이 더욱 공격적으로 나타났다. 이는 암컷이 우점적인 수컷을 선호하고 수컷은 지위가 높은 암컷을 선호함을 시사하며, 사회적 신호와 지위가 개의 짝짓기 쌍 결정에 큰 영향을 미친다는 것을 의미한다.[5]
개과 동물은 다양한 부모 돌봄 행동을 보인다. 2018년 연구에서는 성적 갈등이 이성 간 부모 투자 결정에 영향을 미친다는 점을 보여주었다.[6] 이 연구는 코요테 짝짓기 쌍을 조사한 결과, 수컷의 투자가 암컷의 투자와 비슷하거나 거의 같아지도록 증가한다는 것을 발견했다. 아비가 제공하는 부모 돌봄의 양 또한 어미가 제공하는 돌봄 수준에 따라 변동하는 것으로 나타났다.
부모 투자에 대한 또 다른 연구는 자유로운 개에서 어미가 새끼의 나이에 따라 에너지와 시간 투자를 조절한다는 것을 보여주었다.[7] 어린 시기에 자유로운 개의 사망률이 높기 때문에 어미의 번식 성공률이 크게 감소할 수 있다. 이 연구는 새끼가 성장함에 따라 어미가 에너지를 많이 소모하는 돌봄에서 적게 소모하는 돌봄으로 전환하여 에너지 요구량을 줄이고 더 오랫동안 새끼를 돌볼 수 있도록 한다는 것을 발견했다. 이를 통해 어미는 새끼가 유아기를 넘어 성체가 될 가능성을 높여 자신의 번식 성공률을 높인다.
2017년에 진행된 연구에 따르면, 회색늑대 수컷과 암컷 사이의 공격성은 나이에 따라 다르게 나타난다.[8] 수컷은 암컷보다 경쟁 무리나 홀로 다니는 개체를 쫓아낼 가능성이 더 높았고, 나이가 들수록 공격성이 증가했다. 반면, 암컷은 공격성이 덜하며 평생 동안 공격성 수준이 비교적 일정하게 유지되는 것으로 나타났다. 이는 추가 연구가 필요하지만, 회색늑대의 성별 간 공격성 수준 차이가 그들의 짝짓기 체계와 관련이 있을 수 있음을 시사한다.
4. 2. 부모 돌봄
개과 동물은 일반적으로 일부일처제 쌍으로 생활하며, 이는 새끼 양육의 기초가 된다. 수컷과 암컷 간의 외형적 차이는 크지 않다.
늑대, 들개, 코요테, 자칼과 같은 많은 개과 동물들은 번식쌍(부모)과 그들의 새끼들로 구성된 무리를 이루어 생활한다. 이는 부모가 함께 새끼를 돌보는 기본적인 형태이다. 회색늑대의 경우, 번식쌍과 여러 세대의 새끼들이 함께 사는 확장된 가족 무리를 이루기도 한다.
이러한 무리는 보통 우두머리인 알파 쌍(부모)을 중심으로 구성되며, 현재와 이전 해의 새끼들이 함께 생활하며 서로 돕는다. 무리 생활은 새끼를 보호하고 양육하는 데 중요한 역할을 할 뿐만 아니라, 생존을 위한 사냥에도 필수적이다. 아프리카 들개, 들개, 회색늑대는 자신보다 큰 먹이를 사냥할 때 앞다리를 효과적으로 사용하기 어렵기 때문에 강력한 턱에 의존하며, 무리 전체가 협력하여 사냥한다. 사회성을 가진 이들 포식자는 무리 전체의 힘을 합쳐 자신들의 총 몸무게와 비슷한 크기의 초식동물을 사냥하기도 한다. 예를 들어, 회색늑대 무리는 큰 먹이 중에서도 약한 개체를 노리는 것을 전문으로 하며, 숲에 사는 늑대 무리는 500kg에 달하는 무스를 사냥할 수도 있다. 이러한 협력적 사냥은 새끼들에게 안정적으로 먹이를 공급하는 데 기여한다.
4. 3. 아프리카로의 이주
아프리카 대륙에서 가장 오래된 개속(Canis)의 기록은 케냐 남투르크웰(South Turkwel)에서 발견된 ''Canis sp. A'' 화석이다. 이 화석은 약 358만 년 전에서 320만 년 전 사이의 것으로 추정된다.[12] 2015년에 발표된 아프리카와 유라시아개과 동물의 미토콘드리아 게놈 및 전체 게놈 핵 서열 연구에 따르면, 현재 살아있는 늑대와 같은 개과 동물들은 플라이스토세와 플라이오세에 걸쳐 최소 다섯 차례 이상 유라시아에서 아프리카로 이주해 온 것으로 보인다. 이러한 결과는 아프리카 개과 동물의 다양성이 유라시아 조상의 이주로부터 비롯되었으며, 건조기와 습윤기가 반복된 플라이오-플라이스토세 시기의 기후 변화와 관련이 있다는 화석 증거와도 일치한다.[13] 또한 2017년에는 튀니지 와디 사라트(Wadi Sarrat)의 70만 년 전 지층에서 새로운 개속 화석인 ''Canis othmanii''가 발견되었다. 이 개과 동물은 형태학적으로 아프리카의 다른 개과 동물보다는 유라시아의 개과 동물들과 더 가까운 관계를 보이는 것으로 분석되었다.[14]
참조
[1]
서적
Mammals of the Soviet Union Vol.II Part 1a, SIRENIA AND CARNIVORA (Sea Cows, Wolves and Bears)
Science Publishers, Inc.
[2]
논문
DNA profiles of the eastern Canadian wolf and the red wolf provide evidence for a common evolutionary history independent of the gray wolf
https://cdnsciencepu[...]
2011-02-15
[3]
논문
Dire wolves were the last of an ancient New World canid lineage
https://www.nature.c[...]
2021-01-13
[4]
논문
Do females use their sexual status to gain resource access? Investigating food-for-sex in wolves and dogs
2017-06-01
[5]
논문
Social Variables Affecting Mate Preferences, Copulation and Reproductive Outcome in a Pack of Free-Ranging Dogs
2014-06-06
[6]
논문
It takes two: Evidence for reduced sexual conflict over parental care in a biparental canid
[7]
논문
Clever mothers balance time and effort in parental care: a study on free-ranging dogs
2017-01-01
[8]
논문
Sexually dimorphic aggression indicates male gray wolves specialize in pack defense against conspecific groups
https://digitalcommo[...] [9]
웹사이트
Wolf - Red, Eastern & Ethiopian Wolves, Extinct Falkland Islands & Dire Wolves Britannica
https://www.britanni[...]
2024-02-24
[10]
웹사이트
Canis latrans
https://animaldivers[...]
2024-02-24
[11]
웹사이트
Canis aureus, Golden jackal
https://www.thainati[...]
2024-02-24
[12]
논문
Plio-Pleistocene Carnivora of eastern Africa: Species richness and turnover patterns
[13]
논문
Genome-wide Evidence Reveals that African and Eurasian Golden Jackals Are Distinct Species
[14]
논문
Canis othmanii sp. nov. (Carnivora, Canidae) from the early Middle Pleistocene site of Wadi Sarrat (Tunisia)
[15]
서적
Canis
http://www.departmen[...]
Johns Hopkins University Press
[16]
서적
イヌ属
東京動物園協会
[17]
논문
Genome sequence, comparative analysis and haplotype structure of the domestic dog
[18]
간행물
Old World Canis spp. with taxonomic ambiguity: Workshop conclusions and recommendations. CIBIO. Vairão, Portugal, 28th – 30th May 2019
(PDF)
IUCN/SSC Canid Specialist Group
2020-03-06
[19]
논문
Genome-wide Evidence Reveals that African and Eurasian Golden Jackals Are Distinct Species
[20]
간행물
International Commission on Zoological Nomenclature, Opinion 2027 (Case 3010). Usage of 17 specific names based on wild species which are predated or contemporary with those based on domestic animals (Lepidoptera, Osteichthyes, Mammalia): conserved
[21]
서적
Systema naturæ per regna tria naturæ, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Tomus I
http://www.biodivers[...]
Laurentius Salvius
2015-11-23
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.