스피노사우루스과
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
스피노사우루스과는 백악기 전기부터 번성하여 전 세계적으로 널리 분포했던 수각류 공룡 과이다. 스피노사우루스과는 악어와 유사한 길고 좁은 두개골, 톱니가 없거나 작은 이빨, 코 끝의 로제트, 이차 구개 등의 특징을 공유하며, 반수생 생활을 했던 것으로 추정된다. 이들은 어식성을 기반으로 기회주의적 포식자로서 작은 공룡, 익룡, 악어 등을 섭취했고, 튼튼한 앞다리와 발톱을 이용하여 먹이를 잡았을 것으로 보인다. 스피노사우루스과는 스피노사우루스아과와 바리오닉스아과로 나뉘며, 백악기 후기 세노마니안 시대에 멸종했다.
더 읽어볼만한 페이지
- 스피노사우루스과 - 수코미무스
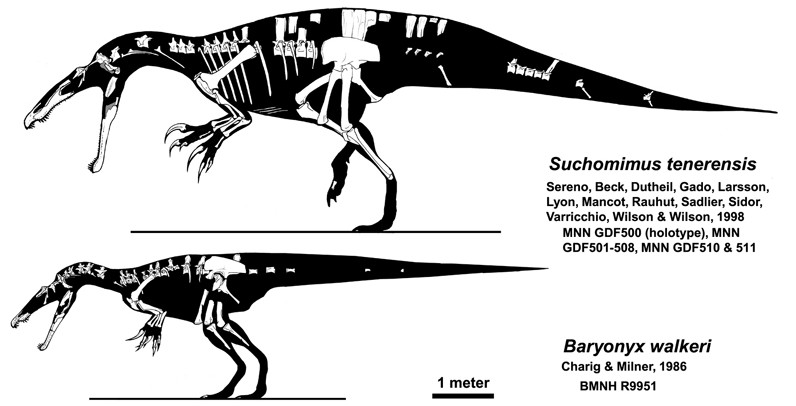
수코미무스는 1997년 니제르에서 발견된 스피노사우루스과의 수각류 공룡으로, 악어를 닮은 머리 모양에서 유래되었으며, 긴 주둥이와 튼튼한 앞다리를 가지고 어식성을 가졌을 것으로 추정된다. - 스피노사우루스과 - 이리타토르
이리타토르는 백악기 전기 브라질에 살았던 스피노사우루스과 수각류 공룡으로, 인위적으로 수정된 화석 발견에 연구자들이 느낀 짜증에서 속명이 유래되었으며, 길고 좁은 두개골과 톱니 없는 이빨을 가진 어식성 동물로 추정된다. - 1915년 기재된 화석 분류군 - 기간토피테쿠스
기간토피테쿠스는 거대한 어금니 화석을 토대로 명명된 멸종된 유인원 속으로, 큰 덩치와 초식 또는 잡식성 식습관을 가진 것으로 추정되지만, 오랑우탄과 더 가까운 관계이며 기후 변화 등으로 멸종했을 가능성이 있고, 미확인 동물의 정체로도 거론된다. - 1915년 기재된 화석 분류군 - 켄트로사우루스 (검룡)
켄트로사우루스는 쥐라기 후기 탄자니아에서 서식한 몸길이 4~5m의 비교적 작은 검룡류 공룡으로, 스테고사우루스와 유사하지만 크기, 갑옷 형태, 유연성에서 차이를 보이며 꼬리 가시로 방어했을 것으로 추정된다.

2. 역사
스피노사우루스과의 역사는 1820년경 영국 고생물학자 기드온 맨텔이 와드허스트 점토층에서 발견한 원뿔형 이빨 화석에서 시작된다.[11] 1841년 리처드 오웬은 이 이빨을 '악어 도마뱀'을 뜻하는 ''수코사우루스''로 명명하며 악어의 일종으로 잘못 분류했다.[12][13] 1897년 ''S. girardi''라는 두 번째 종이 명명되었으나,[14] ''수코사우루스''가 스피노사우루스과라는 사실은 1998년 ''바리오닉스''가 재기술될 때까지 알려지지 않았다.[15]
1912년 이집트 바하리야 지층에서 발견된 화석은 1915년 에른스트 슈트로머에 의해 ''Spinosaurus aegyptiacus''(스피노사우루스)로 명명되었다.[16][17] 1934년 슈트로머는 같은 지층에서 발견된 부분 골격을 새로운 ''Spinosaurus'' 종으로 언급했지만,[18] 이 표본은 이후 ''시길마사사우루스''로 재분류되었다.[19]
이후 1983년 잉글랜드 서리에서 ''바리오닉스''가, 1997년 니제르에서 ''수코미무스''가 발견되었고, 2004년 알칸타라 지층에서 발견된 턱뼈는 2011년 ''옥살라이아''로 분류되었다.[17] 2021년 와이트 섬 벡티스 지층에서 새로운 스피노사우루스과 화석이 발견되었고,[20] 2024년에는 라리오하에서 ''리오하베나트릭스 락스트리스''가 발표되었다.[21]
2. 1. 발견사
최초로 발견된 스피노사우루스과 화석은 1820년경 영국 고생물학자 기드온 맨텔이 발견한 원뿔형 이빨 하나이다.[11] 1841년 리처드 오웬 경은 이 이빨을 '악어 도마뱀'을 뜻하는 ''수코사우루스''라는 악어의 일종으로 잘못 분류했다.[12][13] 그러나 ''수코사우루스''가 스피노사우루스과라는 사실은 1998년 ''바리오닉스''가 재기술될 때까지 알려지지 않았다.[15]
1912년 이집트 바하리야 지층에서 척추, 두개골 조각, 이빨 등으로 구성된 스피노사우루스과 화석이 발견되었다. 이 화석은 1915년 독일 고생물학자 에른스트 슈트로머에 의해 '이집트 가시 도마뱀'을 의미하는 ''Spinosaurus aegyptiacus''(스피노사우루스)라는 새로운 속 및 종의 모식표본으로 기술되었다. 이 공룡의 이름은 다른 수각류에서는 이전에 보이지 않았던 유난히 긴 신경 가시에서 유래했다.[16][17] 그러나 이 모식표본은 제2차 세계 대전 중인 1944년 4월 연합군의 폭격으로 파괴되었다.[16][17]
1983년 잉글랜드 서리의 스모크잭스 채석장에서 비교적 완전한 골격이 발굴되었다. 이 골격은 1986년 영국 고생물학자 앨런 J. 캐릭과 앤젤라 C. 밀너에 의해 ''Baryonyx walkeri''(바리오닉스)라는 새로운 종의 모식표본으로 기술되었다.[17] 1998년 폴 세레노와 동료들은 1997년 니제르에서 발견된 부분 골격을 토대로 수코미무스를 기술했다. 2011년 알렉산더 켈너는 2004년 알칸타라 지층에서 발견된 부분적인 턱뼈를 옥살라이아라는 새로운 스피노사우루스아과 속으로 분류했다.[17]
2021년 잉글랜드 남부 해안 와이트 섬의 벡티스 지층에서 새로운 스피노사우루스과 공룡 화석이 발견되었다. 이 화석은 새로운 종으로 추정되며, 몸길이 약 10미터, 무게는 수 톤에 달하는 것으로 알려졌다. 이는 벡티스 지층에서 확인된 최초의 수각류 화석이다.[20]
2024년 2월에는 2005년 라리오하에서 처음 발견된 ''리오하베나트릭스 락스트리스''가 새로운 스피노사우루스과 종으로 발표되었다. 이 공룡은 이베리아 반도에서 발견된 다섯 번째 스피노사우루스과 종이며, 1억 2천만 년 전에 살았고 몸길이는 약 7~8미터, 체중은 1.5톤으로 추정된다.[21]
2. 2. 과의 정의 및 분류
스피노사우루스과는 1915년 에른스트 슈트뢰머가 스피노사우루스를 처음 명명하면서 만들어졌다.[112] 1998년 폴 세레노는 스피노사우루스과를 "토르보사우루스보다 스피노사우루스에 더 가까운 모든 스피노사우루스류"로 분지학적으로 정의했다.[89]스피노사우루스과는 전통적으로 두 개의 아과로 나뉜다.[112]
- 스피노사우루스아과: 톱니가 없고 곧은 이빨, 두개골에서 바리오닉스아과보다 뒤쪽에 있는 외부 콧구멍이 특징이다. ''이크티오베나토르'', ''이리타토르'', ''옥살라이아'', ''시기르마사사우루스'', ''스피노사우루스'' 등이 속한다.[89][45]
- 바리오닉스아과: 톱니가 있고 약간 구부러진 이빨, 작은 크기, 스피노사우루스아과보다 끝 부분 로제트 뒤쪽에 더 많은 수의 이빨을 가졌다. ''바리오닉스'', ''크리스타투사우루스'', ''수코사우루스'', ''수코미무스'', ''세라토수콥스'', ''리파로베나토르'' 등이 속한다.[92][89][45]
- 세라토수코프시니족: 2021년 바커 등이 ''수코미무스'', ''리파로베나토르'', ''세라토수콥스''를 포함하여 바리오닉스아과 내에 새로 명명했다.[46]
2018년 연구에서는 ''스피노사우루스''와 ''시기르마사사우루스''를 포함하는 스피노사우루스족을 명명했다.[28]
3. 특징
스피노사우루스과는 모두 거대한 몸집을 가진 것이 특징이다. 정확한 크기와 무게 추정은 자료 부족으로 어렵지만, 알려진 속 중에서는 이리테이터가 몸길이 6~8m, 몸무게 약 1t으로 가장 작았다.[22][23] 익티오베나토르, 바리오닉스, 수코미무스는 몸길이 7.5~11m, 몸무게 1~5.2t 사이였다.[24][23][25] 옥살라이아는 몸길이 12~14m, 몸무게 5~7t에 달했을 것으로 추정된다.[26] 스피노사우루스는 몸길이 14m, 몸무게 약 7.4t으로 가장 큰 속이자, 가장 긴 수각류 공룡이자 육상 포식 동물이었다.[27] 시길마사사우루스는 스피노사우루스와 비슷하거나 더 큰 크기였을 수 있지만, 분류학적 관계는 불확실하다.[28]
스피노사우루스과의 큰 몸집은 반수생 생활 방식의 부산물로 진화했을 가능성이 있으며, 다른 대형 수각류와의 먹이 경쟁을 피하기 위해 거대하게 성장했을 수 있다.[29]
3. 1. 두개골
스피노사우루스과의 두개골은 악어류의 두개골과 여러 면에서 유사하며, 길고 낮으며 좁았다.[17] 다른 수각류처럼 두개골의 다양한 창(구멍)은 무게를 줄이는 데 도움이 되었다. 그러나 스피노사우루스류에서는 전안와창이 악어류와 유사하게 크게 감소했다.[35] 전상악골(가장 앞쪽의 주둥이 뼈)의 끝부분은 숟가락 모양으로 확장되어 확대된 이빨의 "말단 로제트"라고 불리는 것을 형성했다. 이러한 확장 뒤에는 위턱에 노치가 있어 상당히 작은 이빨을 가지고 있었고, 이 노치에 치골(하악의 이빨을 포함하는 뼈)의 또한 확장된 끝부분이 맞물렸다. 치골의 확장에 노치가 있었다.[17] 상악골(주요 위턱뼈)은 길었고 콧구멍 아래에 낮고 가지가 뻗어 전상악골의 뒤쪽에 연결되었다. 상악골의 가장 앞부분에 있는 이빨은 작았고, 곧 상당히 커졌으며 턱 뒤쪽으로 갈수록 점차 크기가 줄어들었다.[34] 스피노사우루스류의 이빨 분석과 티라노사우루스류의 이빨과의 비교는 스피노사우루스류의 깊은 뿌리가 이러한 동물의 이빨을 더 잘 고정하고, 포식 및 먹이 섭취 시나리오에서 물어뜯는 과정에서 발생하는 측면 힘에 대한 스트레스를 분산시키는 데 도움이 되었음을 시사한다.[30]
두개골이 고도로 변형되었음에도 불구하고, ''바리오닉스 워커리''와 ''세라토수코프스 인페로디오스''의 내형 분석은 스피노사우루스류의 뇌가 다른 비마니랩토르형 수각류의 뇌와 높은 수준의 유사성을 공유한다는 것을 보여준다.[31]
두개골 위쪽으로는 머리 앞쪽으로 갈수록 짧아지거나 완전히 사라지는 얇고 얕은 시상 능선이 있었으며, 보통 눈 근처나 위에 가장 높았다.[17][32][62] ''스피노사우루스''의 머리 볏은 빗 모양이었고 뚜렷한 수직 홈이 있었고,[32] 반면 ''바리오닉스''와 ''수코미무스''의 볏은 작은 삼각형 돌기처럼 보였다.[33][89] ''이리테이터''의 중간 볏은 눈 위와 뒤에서 둥근 모양으로 멈췄다. 그러나 이 속에 대해 완전히 보존된 두개골이 알려지지 않았기 때문에, ''이리테이터''의 볏의 완전한 모양은 알려지지 않았다.[34] 크리스타투사우루스와 ''수코미무스''(전자의 가능한 동의어)는 모두 좁은 전상악골 볏을 가지고 있었다.[93] ''안가투라마''(''이리테이터''의 가능한 동의어)는 주둥이 끝을 거의 가리고 작은 앞으로 돌출된 특이하게 높은 전상악골 볏을 가지고 있었다.[62]
스피노사우루스류의 콧구멍은 대부분의 수각류에서처럼 주둥이 앞쪽에 있지 않고, 두개골 뒤쪽에, 최소한 전상악골의 이빨 뒤에 위치했다.[17] ''바리오닉스''와 ''수코미무스''의 콧구멍은 컸으며 첫 번째와 네 번째 상악골 이빨 사이에서 시작되었고, 반면 ''스피노사우루스''의 콧구멍은 훨씬 작고 더 뒤로 물러나 있었다. ''이리테이터''의 콧구멍은 ''바리오닉스''와 ''수코미무스''의 콧구멍과 유사하게 위치해 있었고, 크기 면에서 ''스피노사우루스''와 ''수코미무스''의 콧구멍 사이에 있었다.[62] 스피노사우루스류는 길고 보조 구개를 가지고 있었는데, 이는 현존하는 악어류에서도 발견되지만 대부분의 수각류 공룡에서는 발견되지 않는 입천장의 뼈와 거친 구조였다.[35] ''옥살라이아''는 특히 정교한 보조 구개를 가지고 있었고, 대부분의 스피노사우루스는 더 매끄러운 보조 구개를 가지고 있었다.[26] 스피노사우루스류의 이빨은 원뿔형이었고, 타원형에서 원형의 단면을 가지고 있었으며, 톱니가 없거나 매우 미세했다. 이빨은 ''바리오닉스''와 ''수코미무스''의 이빨처럼 약간 휘어진 것부터 ''스피노사우루스''와 ''시아모사우루스''의 이빨처럼 직선적인 것까지 다양했으며, 치관은 종종 세로 홈이나 능선으로 장식되었다.[62][36]
3. 2. 두개골 이후 골격
스피노사우루스과의 어깨뼈에 있는 오훼골은 튼튼하고 갈고리 모양이다. 앞다리는 비교적 크고 잘 발달되어 있으며, 첫 번째 손가락에는 큰 발톱이 있다.[17] 뒷다리는 다소 짧은데, 특히 스피노사우루스에서 두드러진다. 발은 4개의 발가락으로 이루어져 있으며, 발톱은 얕고 길며 바닥이 평평하다(스피노사우루스).[81][82] 척추의 신경돌기는 매우 길어서 수코미무스, 익티오베나토르, 스피노사우루스 등에서 볼 수 있듯이 등 돛을 형성한다.[77][78]
스피노사우루스과의 튼튼한 앞다리와 거대한 갈고리 발톱의 사용에 대해서는 여전히 논쟁 중이다. 1986년, Charig와 Milner는 바리오닉스가 강둑에 웅크리고 앉아 작살질처럼 발톱을 사용하여 물고기를 잡았을 것이라고 추측했는데, 이는 회색곰과 유사하다.[69] 1987년, 영국 생물학자 Andrew Kitchener는 악어와 같은 주둥이와 커다란 발톱을 모두 갖춘 ''바리오닉스''는 하나만으로 충분했을 때 과도하게 많은 물고기 사냥 적응을 한 것처럼 보인다고 주장하며, ''이구아노돈''과 같은 큰 공룡의 시체를 뜯어먹는 데 팔을 사용했을 가능성이 더 높다고 추정했다.[70] 그러나 Charig와 Milner는 1997년 논문에서 대부분의 경우 시체가 이미 초기 포식자들에 의해 상당 부분 비워졌을 것이라고 지적하며 이 가설을 거부했다.[33] 이후 연구에서도 이러한 종류의 특화된 시체 청소는 배제되었다.[17]
1986년, Charig와 Milner는 튼튼한 앞다리와 거대한 엄지 발톱이 ''바리오닉스''가 큰 먹이를 잡고, 죽이고, 찢는 주요 방법이었을 것이라고 제안했다. 반면 긴 주둥이는 주로 물고기를 잡는 데 사용되었을 것이다.[33] 캐나다 고생물학자 François Therrien과 그의 동료들의 2005년 연구는 스피노사우루스의 앞다리가 주둥이가 굽힘 응력을 견딜 수 없다는 점을 감안할 때, 더 큰 먹이를 사냥하는 데 사용되었을 것이라고 동의했다.[71] 2017년, David Hone과 Holtz는 이 과에 대한 검토에서 물 공급원을 파거나 도달하기 어려운 먹이를 파내는 기능, 그리고 둥지를 짓기 위해 땅을 파는 기능 등을 고려했다.[17]
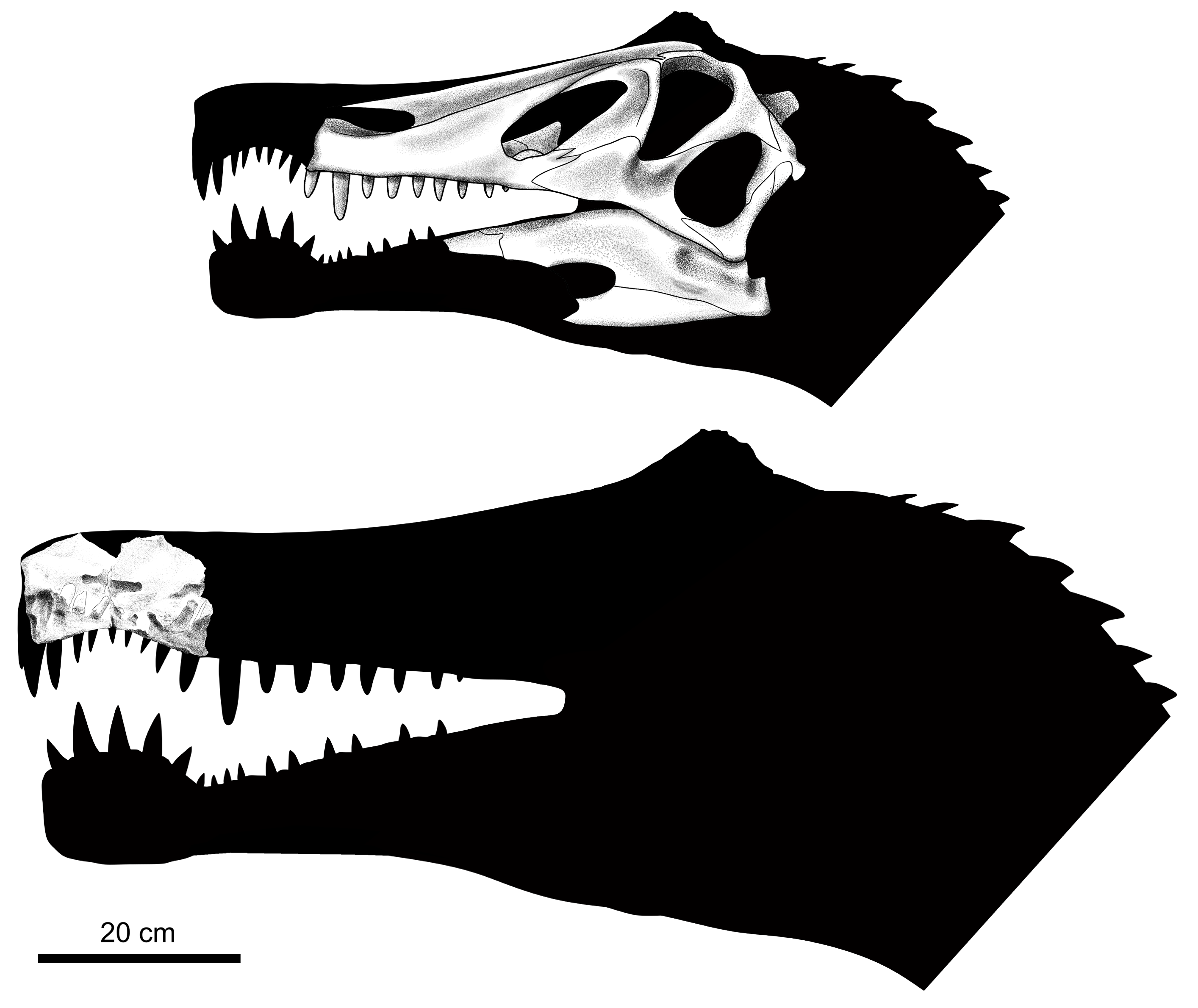
수각류의 머리는 종종 어떤 형태의 볏, 뿔 또는 거친 구조로 장식되는데, 살아있을 때는 케라틴으로 확장되었을 것이다.[72] 스피노사우루스의 머리 볏은 잠재적인 짝에게 성적 과시를 하거나 라이벌 및 다른 포식자를 위협하는 수단이었을 것으로 생각된다.[17] 이전에 수각류 두개골 구조에 대해서도 이와 같은 주장이 제기되었으며, 이는 더욱 시각적인 단서를 제공하기 위해 특이하거나 밝은 색상이 보조했을 수 있다.[72]
스피노사우루스과의 등 돛의 용도에 대해 오랫동안 많은 이론이 제안되어 왔다. 체온 조절,[73] 수영 보조,[74] 에너지 저장 및 단열, 또는 과시 목적으로 사용되었을 수 있다.[75][76] ''Spinosaurus''의 돛이 공작의 꼬리와 유사한 방식으로 구애에 사용되었을 가능성이 있다.[76] 2012년 프랑스 고생물학자 Ronan Allain과 동료들은 ''Ichthyovenator''의 사인파 돛이 구애 과시 또는 종 인식에 사용되었을 가능성이 있다고 제안했다.[77] 2013년 블로그 게시물에서 Darren Naish는 후자의 기능을 가능성이 낮다고 보고, ''Ichthyovenator''의 돛에 대한 성 선택 가설을 지지했다.[78]
2015년 독일 생물물리학자 Jan Gimsa와 동료들은 등돛이 잠수 시 기동성을 향상시켜 수중 운동을 돕고, 돛새치 또는 환도상어와 유사하게 목과 꼬리의 강력한 움직임에 지렛대 역할을 했을 수 있다고 제안했다.[79][80]
어린 스피노사우루스과의 화석은 다소 희귀하다. 하지만 매우 어린 ''Spinosaurus''에 속하는 발톱뼈는 ''Spinosaurus''와 아마도 다른 스피노사우루스과 역시 반수생 적응을 태어날 때 또는 아주 어린 나이에 발달시켜 평생 동안 그 적응을 유지했을 수 있음을 시사한다. 1999년에 발견되어 2018년 시모네 마가누코(Simone Maganuco), 크리스티아노 달 사소(Cristiano Dal Sasso)와 동료들에 의해 기술된 이 표본은 매우 작은 어린 개체에서 나온 것으로 여겨지며, 현재 기술된 스피노사우루스과 중 가장 작은 표본이다.[81][82]
4. 고생태
2010년 로메인 아미오트(Romain Amiot) 등은 스피노사우루스과 뼈의 산소 동위원소 비율을 분석하여 이들이 반수생 생활을 했다는 것을 밝혀냈다. 바리오닉스, 이리타토르, 시아모사우루스, 스피노사우루스의 이빨에서 얻은 동위체를 거북, 악어 등과 비교한 결과, 스피노사우루스과는 거북 및 악어와 가장 유사했다. 시아모사우루스는 다른 스피노사우루스과와 큰 차이를 보여 수중 적응이 덜 된 것으로 나타났으며, 스피노사우루스는 다른 수각류와 가장 큰 차이를 보였다. 아미오트 등은 스피노사우루스과가 하마나 악어처럼 생활의 많은 부분을 물속에서 보냈으며, 이러한 반수생 및 어식성 습성이 다른 대형 수각류와의 공존을 가능하게 했다고 결론지었다. 즉, 서로 다른 먹이와 서식지를 통해 생태적 지위 경쟁을 피했다는 것이다.
4. 1. 식성
스피노사우루스과는 주로 어식성이었지만, 기회주의적인 포식자로서 작은 공룡, 익룡, 악어류 등도 먹이로 삼았다.[45][59][33][63][64]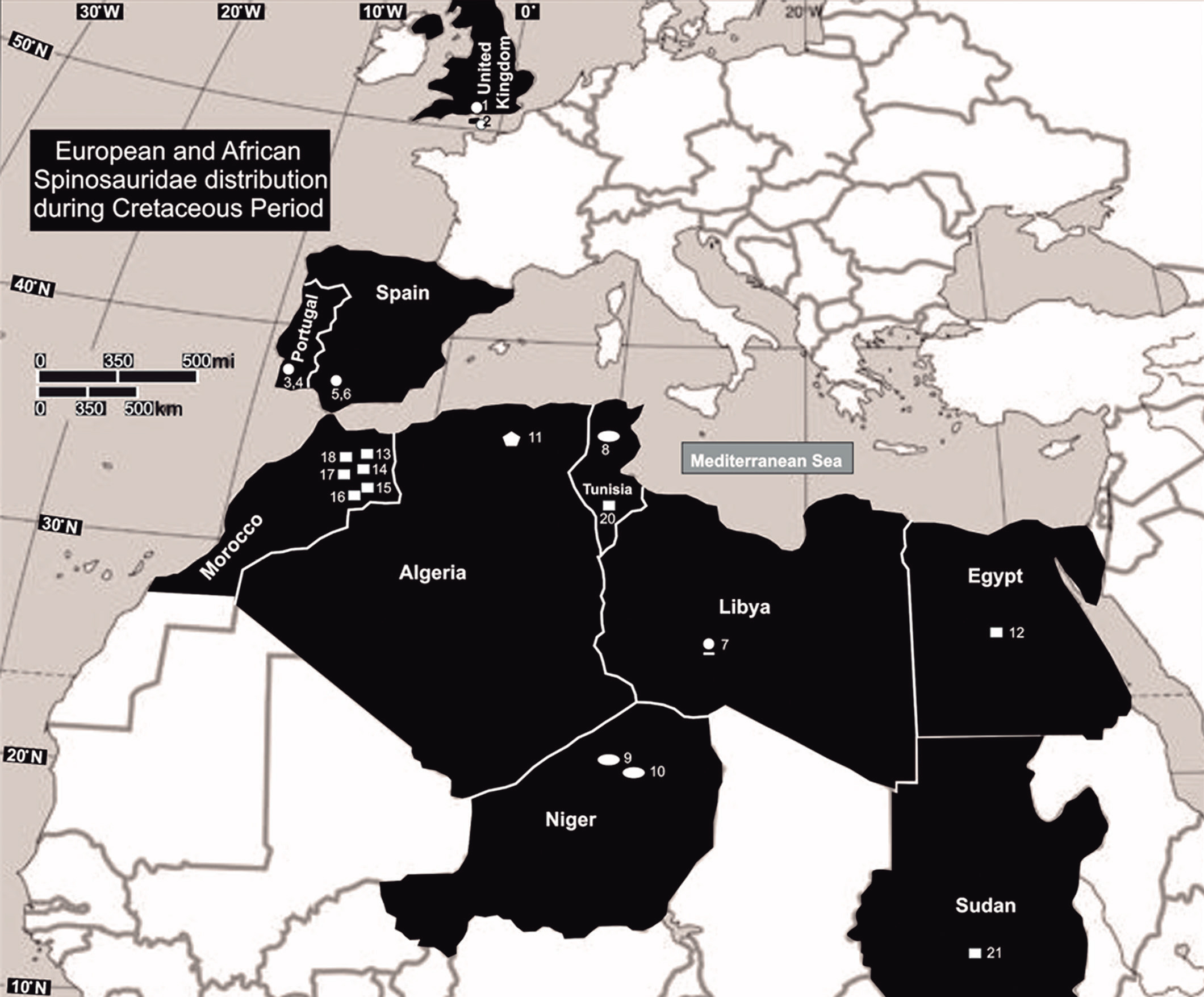
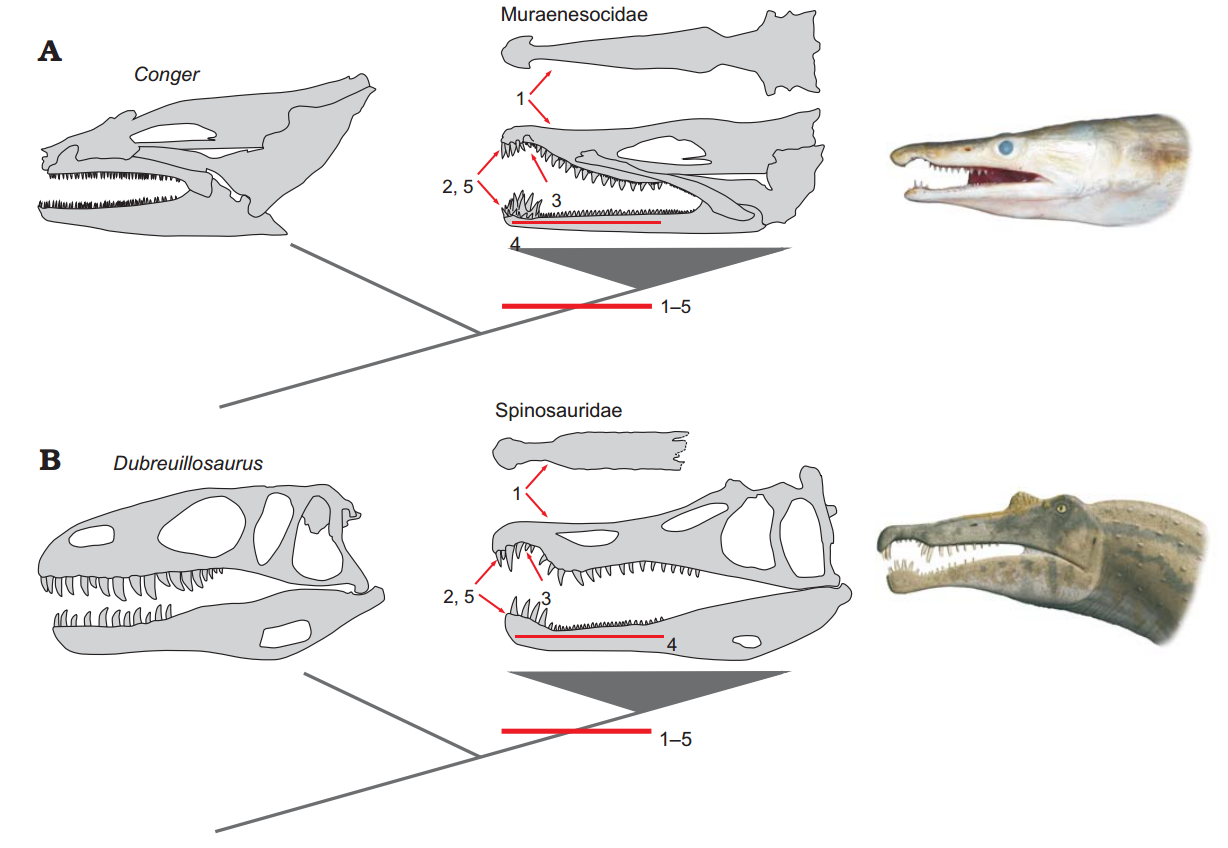
스피노사우루스과의 이빨은 먹이를 찌르고 잡는 데 사용되는 악어의 이빨과 유사하다. 톱니가 작거나 없는 이빨은 살점을 자르거나 찢는 데는 적합하지 않지만, 몸부림치는 먹이를 꽉 잡는 데는 유리했다.[59] 로맹 빌로와 그의 동료들은 스피노사우루스과 턱이 뱀장어의 턱과 유사하며, 수생 먹이를 위한 수렴 진화를 거쳤다고 보았다. 두 종류 모두 위턱과 아래턱 끝 부분에 다른 이빨보다 큰 이빨이 있고, 위턱에는 작은 이빨이 있는 부위가 있어 아래턱의 커다란 이빨이 들어갈 수 있는 틈을 만들어, 이 전체 구조를 말단 로제트라고 한다.[60]

과거에는 턱의 비교를 통해 스피노사우루스과가 현대 악어류와 유사하여 주로 어식성으로 여겨졌다.[45] 1907년, 영국의 고생물학자 에밀리 J. 레이필드와 동료들은 ''바리오닉스''의 두개골에 대한 생체역학 연구를 통해 바리오니키네 턱의 구조가 가비알의 구조와 유사하다는 것을 발견했다. 가비알은 어류 전문이지만, 호주 민물 악어처럼 기회주의적인 포식자이기도 하다. 따라서 스피노사우루스과 주둥이는 어식성과 상관관계가 있지만, 오직 어식성이었다는 것을 나타내지는 않는다.[35]

2013년 앤드루 R. 커프와 레이필드의 연구에서는 ''바리오닉스''와 가비알 두개골의 유사성을 발견하지 못했다. ''바리오닉스''는 ''스피노사우루스''와 가비알보다 비틀림과 굽힘에 대한 저항력이 더 컸다. 두 스피노사우루스과는 가비알, 앨리게이터, 가늘고 긴 코 악어보다 비틀림과 굴곡에 대한 저항력이 열등했다. 스피노사우루스과는 두개골 구조가 아닌 크기 우위를 통해 먹이를 제압했을 수 있다.[61] 2002년, 한스-디터 수에스와 동료들은 스피노사우루스과가 턱을 사용하여 빠르고 강력한 공격으로 작은 먹이를 잡고, 강력한 목 근육을 사용하여 머리를 위아래로 움직였다고 결론지었다. 좁은 주둥이 때문에 먹이를 잡는 동안 두개골이 좌우로 움직이는 것은 불가능하다.[59] 2017년 마르코스 살레스와 세자르 슐츠는 콧구멍의 크기와 위치를 바탕으로 ''스피노사우루스''가 후각에 더 의존하고 ''이리타토르'' 및 바리오니키네보다 더 어식성 생활 방식을 가졌다고 제안했다.[62]

직접적인 화석 증거는 스피노사우루스과가 물고기뿐만 아니라 다양한 종류의 작고 중간 크기의 동물들을 먹었다는 것을 보여준다. ''바리오닉스''는 몸속에서 선사 시대 물고기 ''셰엔스티아''의 비늘과 어린 ''이구아노돈''의 뼈를 발견했다.[45][59][33] ''이리타토르''의 이빨이 익룡의 척추뼈에 박혀 있는 것도 발견되었다.[63][64] ''스피노사우루스'' 주둥이에서는 스클레로린쿠스과의 ''온코프리스티스'' 척추뼈가 박혀 있는 것이 발견되었다.[32] 태국의 사오 쿠아 지층에서는 ''시아모사우루스''의 치관이 용각류 유해와 함께 발견되었다.[65] 포르투갈의 이베로스피누스 화석도 분리된 이구아노돈 이빨과 함께 발견되었다.[66]
2018년 오거스트 하슬러와 동료들의 연구에 따르면, 스피노사우루스과는 물고기와 초식 공룡을 섞어 먹는 식단을 가지고 있었고, 다른 수각류는 주로 초식 공룡을 먹었다. 이는 이 수각류들 사이의 생태적 분할을 나타낼 수 있다.[67] 같은 해, 티토 아우렐리아노와 동료들은 스피노사우루스아과의 식단이 익룡 외에 육상 및 수생 악어류, 자신의 종의 유체, 거북이, 작고 중간 크기의 공룡을 포함했을 수 있다고 제안했다. 이는 스피노사우루스과가 이 특정 생태계 내에서 최상위 포식자가 되었을 것임을 시사한다.[29]
2024년 다모레 등의 연구는 스피노사우루스과가 일반적인 악어류와 유사한 틈새 시장을 가졌다는 이론을 입증했다. 이 연구는 또한 그들의 턱과 이빨이 빠른 공격과 깊고 찌르는 물기에 적합했지만, 살점을 베거나 뼈를 부수는 데는 적합하지 않았다고 제안한다. 바리오니키네 스피노사우루스과는 먹이를 먹을 때 먹이에 대한 구강 처리를 거의 하지 않았지만, 스피노사우루스과는 비교적 큰 척추동물 먹이의 살점을 처리할 수 있는 것으로 밝혀졌다. 이러한 발견은 두 아과에서 어떤 스피노사우루스과도 물고기와 작은 수생 척추동물에만 제한되었음을 시사하지 않는다.[68]
4. 2. 서식지 및 수생 적응
스피노사우루스과는 약 1억 3천만 년에서 9천 5백만 년 전, 바레미안에서 세노마니안 지질 시대까지 백악기에 걸쳐 널리 분포했던 것으로 보인다.[1] 스피노사우루스과의 초기 유해는 니제르의 중생대 쥐라기와 인도에서 알려져 있다.[1][48]2010년 로메인 아미오트(Romain Amiot) 등은 스피노사우루스과 뼈의 산소 동위원소 비율을 분석하여 이들이 반수생 생활을 했음을 밝혀냈다. 바리오닉스, 이리타토르, 시아모사우루스, 스피노사우루스의 이빨에서 얻은 동위체를 거북, 악어 등과 비교한 결과, 스피노사우루스과는 거북 및 악어와 가장 유사했다. 시아모사우루스는 다른 스피노사우루스과와 큰 차이를 보여 수중 적응이 덜 된 것으로 나타났으며, 스피노사우루스는 다른 수각류와 가장 큰 차이를 보였다. 아미오트 등은 스피노사우루스과가 하마나 악어처럼 생활의 많은 부분을 물속에서 보냈으며, 이러한 반수생 및 어식성 습성이 다른 대형 수각류와의 공존을 가능하게 했다고 결론지었다. 즉, 서로 다른 먹이와 서식지를 통해 생태적 지위 경쟁을 피했다는 것이다.
4. 3. 앞다리와 등 돛의 기능
스피노사우루스과의 튼튼한 앞다리와 거대한 갈고리 발톱은 먹이를 잡는 데 사용되었을 것으로 보인다. 1986년, Charig와 Milner는 바리오닉스가 회색곰처럼 강가에 웅크리고 앉아 발톱으로 물고기를 잡았을 것이라고 추측했다.[69] 그러나 1987년, 영국 생물학자 Andrew Kitchener는 바리오닉스가 큰 발톱으로 사체를 찢고 뼈를 부순 뒤, 긴 주둥이로 내장을 파먹었을 것이라고 주장했다.[70] 1997년 Charig와 Milner는 이 가설을 반박했는데, 대부분의 사체는 이미 다른 포식자들에 의해 비워졌을 것이라고 지적했다.[33]1986년, Charig와 Milner는 튼튼한 앞다리와 거대한 엄지발톱이 바리오닉스가 큰 먹이를 잡고, 죽이고, 찢는 데 사용되었을 것이라고 제안했다.[33] 2005년, 캐나다 고생물학자 François Therrien과 동료들은 스피노사우루스의 앞다리가 큰 먹이를 사냥하는 데 사용되었을 것이라고 주장했다.[71] 2017년, David Hone과 Holtz는 물을 파거나 먹이를 파내는 기능, 둥지를 짓기 위해 땅을 파는 기능 등도 고려했다.[17]
스피노사우루스과의 등 돛은 체온 조절,[73] 수영 보조,[74] 에너지 저장, 과시[75][76] 등 여러 기능을 했을 것으로 추정된다. 1915년 Stromer는 신경 가시의 크기가 수컷과 암컷 사이에 다를 수 있다고 추측했다.[76] 2012년 프랑스 고생물학자 Ronan Allain과 동료들은 이크티오베나토르의 돛이 구애 과시나 종 인식에 사용되었을 가능성이 있다고 제안했다.[77] 2013년 Darren Naish는 성 선택 가설을 지지했다.[78]
2015년 독일 생물물리학자 Jan Gimsa와 동료들은 돛이 잠수 시 기동성을 향상시켜 수중 운동을 돕고, 돛새치나 환도상어처럼 목과 꼬리의 강력한 움직임에 지렛대 역할을 했을 수 있다고 제안했다.[79][80]
5. 분포
확인된 스피노사우루스과는 북아메리카, 호주, 남극 대륙을 제외한 모든 대륙에서 발견되었으며, 그중 첫 번째는 이집트의 바하리야 지층에서 발견된 ''스피노사우루스 아에깁티아쿠스''이다.[76] 바리오니키네는 흔했으며, 예를 들어 영국의 바레미안과 스페인에서 살았던 ''바리오닉스''가 있다. ''바리오닉스''와 유사한 이빨은 스페인의 오트리비안 초기와 압티안 후기 퇴적물, 영국의 오트리비안에서도 발견된다.[17][88] 바리오니키네는 아프리카에서 ''수코미무스 테네렌시스'', ''크리스타투사우루스 라파렌티'' 뿐만 아니라 니제르 압티안에서 발견된 ''바리오닉스'' 유사 이빨로 대표되었다.[92][89][90] 유럽에서도 영국의 ''수코사우루스 쿨트리덴스''와 ''수코사우루스 기라르디''가 발견되었다. ''바리오닉스'' 유사 이빨은 영국 서식스의 애시다운 샌즈와 스페인 부르고스 주에서도 보고되었다.[88] 다른 유럽 스피노사우루스과인 ''카마릴라사우루스 키루게다에''와 ''이베로스피누스 나타리오이''는 각각 스페인과 포르투갈의 바레미안에서 알려져 있다.[91][47]
스피노사우루스아과의 가장 초기의 기록은 스페인의 바레미안 종인 ''발리보나베나트릭스 카니''에서 발견되었다.[41] 스피노사우루스아과는 튀니지와 알제리의 알비안 퇴적물, 이집트와 모로코의 세노마니안 퇴적물에서도 존재한다. 아프리카에서 바리오니키네는 압티안에서 흔했고, 이후 알비안과 세노마니안 시대에 스피노사우루스아과로 대체되었다.[92] 예를 들어, 모로코의 켐 켐 지층에서는 많은 대형 포식자가 공존하는 생태계가 존재했다.[93][83] 초기 백악기의 스피노사우루스아과 하악골 파편도 튀니지에서 보고되었으며, ''스피노사우루스''로 분류되었다.[92] 스피노사우루스아과의 분포는 브라질의 ''이리타토르 챌린저리, 안가투라마 리마이,'' ''옥살라이아 퀼롬벤시스'' 발견으로 남아메리카로 확장되었다.[64][26] 레오나르도 살가도와 그의 동료들은 아르헨티나에서 스피노사우루스과로 분류되는 화석 이빨도 발견했다.[94] 겐고 타나카 등은 이 분류에 대해 회의적이며, 이 이빨의 기원으로 가장 유력한 동물로 악어인 ''하다마수쿠스''를 제시했다.[95]
부분 골격과 수많은 화석 이빨은 스피노사우루스과가 아시아에 널리 분포했음을 나타낸다. 세 개의 분류군(모두 스피노사우루스아과)이 명명되었다. 태국의 ''시아모사우루스 수티토르니'', 중국의 "''시노플리오사우루스''" ''푸수이엔시스'', 라오스의 ''이크티오베나토르 라오엔시스''가 있다.[40][92][96] 스피노사우루스과 이빨은 말레이시아에서 발견되었으며, 이는 이 나라에서 발견된 최초의 공룡 유해였다.[97] 일부 중간 단계 표본은 명명된 분류군의 가장 최근 시점을 지나 스피노사우루스과의 알려진 범위를 확장한다. 하나의 바리오니키네 이빨이 중국 허난의 마지아춘 지층에서 중간 산토니안에서 발견되었다.[96] 그러나 이 이빨에는 스피노사우루스과 고유의 형질공유가 부족하다.[98] 스페인 트루엘의 초기 바레미안 블레사 지층에 있는 라 칸탈레라-1 유적지에서 두 종류의 스피노사우루스과 이빨이 발견되었으며, 이는 잠정적으로 불확실한 스피노사우루스아과 및 바리오니키네 분류군으로 지정되었다.[99] 호주 초기 백악기의 에우메랄라 지층에서 불확실한 스피노사우루스과가 발견되었다.[100] 이는 NMV P221081로 지정된 4cm 길이의 부분적인 경추 1개로 알려져 있다. 신경궁의 대부분이 없다. 이 표본은 길이가 약 2m~3m로 추정되는 어린 개체에서 유래했다. 모든 스피노사우루스과 중, ''바리오닉스''와 가장 유사하다.[101] 2019년에는 이 척추뼈가 스피노사우루스가 아닌 메가랍토르과 수각류에 속한다는 주장이 제기되었다.[102]
6. 진화
스피노사우루스과는 쥐라기 중기에 메갈로사우루스상과 공룡으로부터 진화했을 것으로 추정된다. 스피노사우루스과 두개골의 독특한 적응은 악어와의 수렴 진화 결과일 수 있다. 이들은 길고 좁으며 악어와 같은 두개골, 섬세하거나 톱니가 없는 둥근 이빨, 코의 끝부분 로제트, 비틀림에 더 강하게 만드는 이차 구개와 같은 특징을 공유했다. 반대로, 수각류의 원시적이고 전형적인 상태는 날카로운 톱니 모양의 칼날 모양(ziphodont) 이빨을 가진 높고 좁은 주둥이였다.[49] 스피노사우루스과의 두개골 적응은 악어의 두개골과 수렴 진화했으며, 후기 그룹의 초기 구성원은 전형적인 수각류와 유사한 두개골을 가지고 있었지만, 나중에는 길어진 주둥이, 원뿔형 이빨, 이차 구개를 발달시켰다. 이러한 적응은 육상 먹이에서 물고기로의 식단 변화의 결과일 수 있다. 악어와 달리, 바리오니키네 스피노사우루스과의 사지 골격은 수생 적응을 보이지 않는 것으로 보인다.[5][49]
세레노와 동료들은 1998년에 스피노사우루스과의 큰 엄지 발톱과 강력한 앞다리가 쥐라기 중기에 진화했다고 제안했는데, 그 이유는 전자의 특징이 그들의 메갈로사우루스과 친척과 공유되기 때문이다. 또한 스피노사우루스아과와 바리오니키네가 초기 백악기의 바레미안 시대 이전에 분기되었다고 제안했다.[50]
초기에는 초대륙 판게아에 분포했으나, 테티스 해가 열리면서 스피노사우루스아과와 바리오닉스아과로 분리되었다. 스피노사우루스아과는 남쪽(아프리카와 남아메리카: 곤드와나)에서, 바리오니키네는 북쪽(유럽: 로라시아)에서 진화했으며, 수코미무스는 한 번의 북쪽에서 남쪽으로의 분산 사건의 결과였다.[50]
스피노사우루스과의 생물지리학적 분포와 진화 경로는 복잡하며, 추가 연구가 필요하다.
7. 계통 발생
스피노사우루스과는 1915년 에른스트 슈트뢰머가 스피노사우루스 한 속만을 포함하는 것으로 명명했다. 이 분류군은 스피노사우루스와 가까운 친척들이 더 많이 발견되면서 확장되었다. 스피노사우루스과에 대한 최초의 분지학적 정의는 1998년 폴 세레노에 의해 제공되었는데, "토르보사우루스보다 스피노사우루스에 더 가까운 모든 스피노사우루스류"였다.[89]
전통적으로 스피노사우루스과는 두 개의 아과로 나뉜다.
- 스피노사우루스아과: 톱니가 없고 곧은 이빨, 그리고 두개골에서 바리오닉스아과보다 뒤쪽에 있는 외부 콧구멍을 특징으로 한다. ''이크티오베나토르'', ''이리타토르'', ''옥살라이아'', ''시기르마사우루스'', ''스피노사우루스'' 속을 포함한다.[89][45]
- 바리오닉스아과: 톱니가 있고 약간 구부러진 이빨, 작은 크기, 그리고 스피노사우루스아과보다 끝 부분에 있는 로제트 뒤쪽에 더 많은 수의 이빨을 가진다. ''바리오닉스'', ''크리스타투사우루스'', ''수코사우루스'', ''수코미무스'', ''세라토수코프스'', ''리파로베나토르'' 속을 포함한다.[92][89][45]
''시아모사우루스''와 같은 다른 종들은 바리오닉스아과 또는 스피노사우루스아과에 속할 수 있지만, 너무 불완전하게 알려져 있어 확신을 가지고 할당할 수 없다.[92] ''시아모사우루스''는 2018년에 스피노사우루스아과로 분류되었지만, 그 결과는 잠정적이며 완전히 결정적이지 않다.[28]
스피노사우루스아과는 1998년 세레노에 의해 명명되었고, 2004년 토마스 홀츠와 그의 동료들에 의해 ''바리오닉스 왈케리''보다 ''스피노사우루스 아에깁티아쿠스''에 더 가까운 모든 분류군으로 정의되었다. 바리오닉스아과는 1986년 찰리그와 밀너에 의해 명명되었다. 그들은 새로 발견된 ''바리오닉스''에 대해 아과와 바리오닉스과를 모두 세웠고, 그 후 스피노사우루스과에 포함되었다. 그들의 아과는 2004년 홀츠와 그의 동료들에 의해 ''스피노사우루스 아에깁티아쿠스''보다 ''바리오닉스 왈케리''에 더 가까운 모든 분류군의 보완적인 분류군으로 정의되었다.
2017년 마르코스 살레스와 세자르 슐츠의 연구에 따르면, 두개골-치열 특징과 분지학적 분석을 기반으로 볼 때 남아메리카 스피노사우루스류인 ''앙가투라마''와 ''이리타토르''는 바리오닉스아과와 스피노사우루스아과의 중간에 위치했다. 2018년 아덴 등의 연구는 ''스피노사우루스''와 ''시기르마사우루스''를 포함하는 스피노사우루스족을 명명했는데,[28] 후자는 스피노사우루스과로서의 타당성에 대해 논쟁의 여지가 있다. 2021년 바커 등은 ''수코미무스'', ''리파로베나토르'', 그리고 ''세라토수코프스''를 포함하는 바리오닉스아과 내에 새로운 세라토수코프시니족을 명명했다.[46]
2017년 연구는 바리오닉스아과가 사실 단계통군이 아닐 수 있음을 시사한다. 그들의 분기도는 아래와 같다.[62]
다음 분기도는 2012년 알랭과 그의 동료들이 스피노사우루스과만을 보여주도록 단순화한 테타누라의 분석을 보여준다:[40]
2018년 아덴과 그의 동료들의 계통 발생 분석은 많은 명명되지 않은 분류군을 포함하여 바리오닉스아과를 단계통군으로 해결했으며, 또한 ''시기르마사우루스''와 ''스피노사우루스''의 분류군에 대해 새로운 용어인 스피노사우루스족을 만들었다.[28]
2021년, 크리스 바커, 혼, 대런 내쉬, 안드레아 코, 록우드, 포스터, 클라킨, 슈나이더, 그리고 고슬링은 두 개의 새로운 스피노사우루스과 종인 ''세라토수코프스 인페로디오스''와 ''리파로베나토르 밀네라이''를 기술했다. 이 논문에서 그들은 테로포드의 일반적인 범위를 통합했지만, 주로 스피노사우루스과에 초점을 맞춘 계통 발생 분석을 수행했다. [46]
참조
[1]
논문
First Jurassic Evidence of a Possible Spinosaurid Pedal Ungual, from the Jaisalmer Basin, India
[2]
서적
The Dinosauria
University of California Press
2004
[3]
논문
A new theropod dinosaur from the Callovian Balabansai Formation of Kyrgyzstan
2024-08-01
[4]
논문
A Unified Framework for Predatory Dinosaur Macroevolution
https://www.research[...]
[5]
논문
Semiaquatic adaptations in a giant predatory dinosaur
https://researchport[...]
2014
[6]
논문
Sigilmassasaurus is Spinosaurus: A reappraisal of African spinosaurines
2020-10
[7]
논문
A description of Megalosaurus bucklandii (Dinosauria: Theropoda) from the Bathonian of the UK and the relationships of Middle Jurassic theropods
2010
[8]
논문
A NEW PHYLOGENY OF THE CARNIVOROUS DINOSAURS
https://www.geol.umd[...]
[9]
논문
Probable basal allosauroid from the early Middle Jurassic Cañadón Asfalto Formation of Argentina highlights phylogenetic uncertainty in tetanuran theropod dinosaurs
2019
[10]
논문
Exceptionally preserved juvenile megalosauroid theropod dinosaur with filamentous integument from the Late Jurassic of Germany
2012
[11]
서적
The fossils of the South Downs or, Illustrations of the geology of Sussex.
https://www.biodiver[...]
L. Relfe
1822
[12]
문서
Odontography
London: Hippolyte Baillière
1840–1845
[13]
문서
Report on British fossil reptiles. Part II
[14]
문서
Vertébrés fossiles du Portugal. Contribution à l’étude des poissons et des reptiles du Jurassique et du Crétacique.
Lisbonne: Direction des Travaux géologiques du Portugal
1897–1898
[15]
문서
Fish-eating theropods: A short review of the systematics, biology and palaeobiogeography of spinosaurs
[16]
논문
New Information Regarding the Holotype of Spinosaurus Aegyptiacus Stromer, 1915
2006-03
[17]
논문
A Century of Spinosaurs - A Review and Revision of the Spinosauridae with Comments on Their Ecology
http://qmro.qmul.ac.[...]
2017-06
[18]
논문
Ergebnisse der Forschungsreisen Prof. E. Stromers in den Wüsten Ägyptens. II. Wirbeltier-Reste der Baharije-Stufe (unterstes Cenoman). 13. Dinosauria
[19]
논문
A reappraisal of the morphology and systematic position of the theropod dinosaur Sigilmassasaurus from the 'middle' Cretaceous of Morocco
2015-10-20
[20]
웹사이트
Scientists unearth remains of one of Europe's biggest predatory dinosaurs
https://www.cnn.com/[...]
2022-06-11
[21]
웹사이트
Researchers discover a new species of carnivorous dinosaur in La Rioja, Spain
https://phys.org/new[...]
2024-05-25
[22]
서적
The Ultimate Guide to Dinosaurs
https://archive.org/[...]
Ticktock Books
2009
[23]
문서
'Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages'
http://www.geol.umd.[...]
[24]
서적
The Princeton field guide to dinosaurs
2016-10-25
[25]
논문
My theropod is bigger than yours … or not: estimating body size from skull length in theropods
2007-03-12
[26]
논문
A new dinosaur (Theropoda, Spinosauridae) from the Cretaceous (Cenomanian) Alcântara Formation, Cajual Island, Brazil
2011-03
[27]
논문
Spinosaurus is not an aquatic dinosaur
[28]
논문
Aquatic adaptation in the skull of carnivorous dinosaurs (Theropoda: Spinosauridae) and the evolution of aquatic habits in ''Spinosaurus''
[29]
논문
Semi-aquatic adaptations in a spinosaur from the Lower Cretaceous of Brazil
http://paleorxiv.org[...]
2018-10
[30]
서적
Tyrannosaurus Rex, the Tyrant King
Indiana University Press
[31]
논문
Modified skulls but conservative brains? The palaeoneurology and endocranial anatomy of baryonychine dinosaurs (Theropoda: Spinosauridae)
[32]
논문
New information on the skull of the enigmatic theropod Spinosaurus , with remarks on its size and affinities
2005-12-30
[33]
논문
Baryonyx walkeri, a fish-eating dinosaur from the Wealden of Surrey
http://biostor.org/r[...]
[34]
논문
Irritator challengeri, a spinosaurid (Dinosauria: Theropoda) from the Lower Cretaceous of Brazil
[35]
논문
Functional morphology of spinosaur 'crocodile-mimic' dinosaurs
2007-12-12
[36]
논문
A Catalogue of Material and Review of the Spinosauridae
https://archives.pal[...]
2010
[37]
논문
Semiaquatic adaptations in a giant predatory dinosaur
https://researchport[...]
[38]
논문
New Information on the Cranial Anatomy of ''Acrocanthosaurus atokensis'' and Its Implications for the Phylogeny of Allosauroidea (Dinosauria: Theropoda)
2011
[39]
뉴스
Fish Swam in Fear
https://www.newscien[...]
New Scientist
1998-11-21
[40]
논문
The first definitive Asian spinosaurid (Dinosauria: Theropoda) from the early cretaceous of Laos
[41]
논문
A new spinosaurid theropod (Dinosauria: Megalosauroidea) from the late Barremian of Vallibona, Spain: Implications for spinosaurid diversity in the Early Cretaceous of the Iberian Peninsula
2019
[42]
웹사이트
http://www.estadao.c[...]
2009-05-14
[43]
논문
Preliminary information on a dinosaur (Theropoda, Spinosauridae) pelvis from the Cretaceous Santana Formation (Romualdo Member) Brazil
2005
[44]
논문
Tail-propelled aquatic locomotion in a theropod dinosaur
2020-05-07
[45]
논문
Structural performance of tetanuran theropod skulls, with emphasis on the Megalosauridae, Spinosauridae and Carcharodontosauridae
2011
[46]
논문
New spinosaurids from the Wessex Formation (Early Cretaceous, UK) and the European origins of Spinosauridae
[47]
논문
A new theropod dinosaur from the early cretaceous (Barremian) of Cabo Espichel, Portugal: Implications for spinosaurid evolution
2022-02-16
[48]
논문
Isolated theropod teeth from the Middle Jurassic of Niger and the early dental evolution of Spinosauridae
2015
[49]
논문
Spinosaurs as crocodile mimics
[50]
논문
A long-snouted predatory dinosaur from Africa and the evolution of spinosaurids
[51]
논문
A new specimen of ''Spinosaurus'' (Dinosauria, Theropoda) from the Lower Cretaceous of Tunisia, with remarks on the evolutionary history of the Spinosauridae
http://doc.rero.ch/r[...]
[52]
논문
Fish-eating theropods: A short review of the systematics, biology and palaeobiogeography of spinosaurs
[53]
논문
The spinosaurid dinosaur ''Baryonyx'' (Saurischia, Theropoda) in the Early Cretaceous of Portugal
http://doc.rero.ch/r[...]
[54]
논문
The distribution of dental features in non-avian theropod dinosaurs: Taxonomic potential, degree of homoplasy, and major evolutionary trends
https://palaeo-elect[...]
2019
[55]
논문
''Ceratosaurus'' (Theropoda, Ceratosauria) teeth from the Tacuarembó Formation (Late Jurassic, Uruguay)
http://adsabs.harvar[...]
2020
[56]
논문
Spinosaurid Dinosaurs from the Early Cretaceous of North Africa and Europe: Fossil Record, Biogeography and Extinction
2017
[57]
논문
An update of the spinosaurid (Dinosauria: Theropoda) fossil record from the Lower Cretaceous of the Iberian Peninsula: distribution, diversity, and evolutionary history
2020
[58]
논문
New spinosaurids from the Wessex Formation (Early Cretaceous, UK) and the European origins of Spinosauridae
2021
[59]
논문
Irritator challengeri , a spinosaurid (Dinosauria: Theropoda) from the Lower Cretaceous of Brazil
2002-09-19
[60]
논문
Convergent evolution of jaws between spinosaurid dinosaurs and pike conger eels
2016
[61]
논문
Feeding Mechanics in Spinosaurid Theropods and Extant Crocodilians
2013-05-28
[62]
논문
Spinosaur taxonomy and evolution of craniodental features: Evidence from Brazil
[63]
논문
Pterosaurs in Mesozoic food webs: a review of fossil evidence
2018
[64]
논문
Pterosaurs as part of a spinosaur diet
2004-07
[65]
논문
The dinosaur fauna of the Sao Khua Formation of Thailand and the beginning of the Cretaceous radiation of dinosaurs in Asia
http://doc.rero.ch/r[...]
1999-06
[66]
논문
Morphofunctional Analysis of the Quadrate of Spinosauridae (Dinosauria: Theropoda) and the Presence of Spinosaurus and a Second Spinosaurine Taxon in the Cenomanian of North Africa
2016
[67]
논문
Calcium isotopes offer clues on resource partitioning among Cretaceous predatory dinosaurs
2018-04-11
[68]
논문
Prey size and ecological separation in spinosaurid theropods based on heterodonty and rostrum shape
https://anatomypubs.[...]
2024-08-28
[69]
논문
''Baryonyx'', a remarkable new theropod dinosaur
[70]
논문
Function of Claws' claws
1987-01
[71]
서적
The Carnivorous Dinosaurs
Indiana University Press
[72]
서적
The Carnivorous Dinosaurs
Indiana University Press
[73]
서적
The Evolution and Ecology of the Dinosaurs
Eurobook Limited
[74]
논문
The riddle of Spinosaurus aegyptiacus' dorsal sail
https://repository.p[...]
2016-05
[75]
논문
Neural spine elongation in dinosaurs: sailbacks or buffalo-backs?
[76]
논문
Ergebnisse der Forschungsreisen Prof. E. Stromers in den Wüsten Ägyptens. II. Wirbeltier-Reste der Baharije-Stufe (unterstes Cenoman). 3. Das Original des Theropoden ''Spinosaurus aegyptiacus'' nov. gen., nov. spec
http://www.megauploa[...]
[77]
논문
The first definitive Asian spinosaurid (Dinosauria: Theropoda) from the early cretaceous of Laos
[78]
웹사이트
Dinosaurs and their exaggerated structures : species recognition aids, or sexual display devices?
https://blogs.scient[...]
2013
[79]
논문
The riddle of ''Spinosaurus aegyptiacus'' dorsal sail
2015
[80]
논문
Neural spine elongation in dinosaurs: sailbacks or buffalo-backs?
2015
[81]
간행물
The smallest biggest theropod dinosaur: A new fossil from Africa represents a small juvenile individual of the huge sail-backed Spinosaurus
https://www.scienced[...]
PeerJ
2018-05-30
[82]
뉴스
Paleontologists Find Fossil of Smallest Spinosaurus
http://www.sci-news.[...]
2018-05-31
[83]
논문
Oxygen isotope evidence for semi-aquatic habits among spinosaurid theropods
[84]
논문
Taphonomic evidence supports an aquatic lifestyle for Spinosaurus
https://researchport[...]
2021-01
[85]
웹사이트
Diving dinosaurs? Caveats on the use of bone compactness and pFDA for inferring lifestyle
https://www.biorxiv.[...]
[86]
논문
Diving dinosaurs? Caveats on the use of bone compactness and pFDA for inferring lifestyle
2024
[87]
논문
A buoyancy, balance and stability challenge to the hypothesis of a semi-aquatic ''Spinosaurus'' Stromer, 1915 (Dinosauria: Theropoda)
2018
[88]
논문
A new specimen of the theropod dinosaur ''Baryonyx'' from the early Cretaceous of Portugal and taxonomic validity of ''Suchosaurus''
http://docentes.fct.[...]
[89]
논문
A Long-Snouted Predatory Dinosaur from Africa and the Evolution of Spinosaurids
1998-11-13
[90]
논문
New data on spinosaurid dinosaurs from the early cretaceous of the Sahara
1998-09
[91]
논문
A spinosaurid from Thailand (Sao Khua Formation, Early Cretaceous) and a reassessment of Camarillasaurus cirugedae from the Early Cretaceous of Spain
2021-12-02
[92]
논문
A new specimen of Spinosaurus (Dinosauria, Theropoda) from the Lower Cretaceous of Tunisia, with remarks on the evolutionary history of the Spinosauridae
http://doc.rero.ch/r[...]
2002-09-01
[93]
논문
Morphofunctional Analysis of the Quadrate of Spinosauridae (Dinosauria: Theropoda) and the Presence of Spinosaurus and a Second Spinosaurine Taxon in the Cenomanian of North Africa.
2016-01-06
[94]
논문
Upper Cretaceous vertebrates from El Anfiteatro area, Río Negro, Patagonia, Argentina
2009-06
[95]
서적
Bulletin of Gunma Museum of Natural History
http://www.gmnh.pref[...]
[96]
논문
A PROBABLE BARYONYCHINE (THEROPODA: SPINOSAURIDAE) TOOTH FROM THE UPPER CRETACEOUS OF HENAN PROVINCE, CHINA
http://www.vertpala.[...]
2010-03-15
[97]
웹사이트
First discovery of dinosaur fossils in Malaysia
https://www.scienced[...]
2014
[98]
논문
Second discovery of a spinosaurid tooth from the Sebayashi Formation (Lower Cretaceous), Kanna Town, Gunma Prefecture, Japan
http://www.gmnh.pref[...]
2017
[99]
논문
On the spinosaurid theropod teeth from the early Barremian (Early Cretaceous) Blesa Formation (Spain)
2016-08-17
[100]
뉴스
Australian 'Spinosaur' unearthed
http://www.australia[...]
2018-04-15
[101]
논문
First spinosaurid dinosaur from Australia and the cosmopolitanism of Cretaceous dinosaur faunas
2011-12-23
[102]
논문
New megaraptorid (Dinosauria: Theropoda) remains from the Lower Cretaceous Eumeralla Formation of Cape Otway, Victoria, Australia
https://figshare.com[...]
[103]
간행물
"Spinosaurid teeth from the Late Jurassic of Tengaduru, Tanzania, with remarks on the evolutionary and biogeographical history of the Spinosauridae." In J.-M. Mazin, J. Pouech, P. Hantzpergue, V. Lacombe., ''Mid-Mesozoic Life and Environments''. Cognac (France), June 24th-28th 2008, pp. 26-28.
[104]
간행물
"Upper Cretaceous vertebrates from El Anfiteatro area, Río Negro, Patagonia, Argentina." ''Cretaceous Research'' in press.
[105]
논문
Fine sculptures on a tooth of ''Spinosaurus'' (Dinosauria, Theropoda) from Morocco
[106]
논문
A probable baryonychine (Theropoda: Spinosauridae) tooth from the Upper Cretaceous of Henan Province, China
[107]
논문
"''Irritator challengeri'', a spinosaurid (Dinosauria: Theropoda) from the Lower Cretaceous of Brazil"
[108]
논문
Functional morphology of spinosaur 'crocodile-mimic' dinosaurs
[109]
논문
Feeding mechanics in spinosaurid theropods and extant crocodilians
[110]
논문
Pterosaurs as part of a spinosaur diet
[111]
논문
Oxygen isotope evidence for semi-aquatic habits among spinosaurid theropods
[112]
논문
New spinosaurids from the Wessex Formation (Early Cretaceous, UK) and the European origins of Spinosauridae
https://doi.org/10.1[...]
[113]
논문
An Early Cretaceous spinosaurid theropod from southern China
[114]
논문
Australian 'Spinosaur' unearthed
http://www.australia[...]
[115]
논문
The first definitive Asian spinosaurid (Dinosauria: Theropoda) from the early Cretaceous of Laos
http://link.springer[...]
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com