탈라토사우루스
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
탈라토사우루스는 중생대 트라이아스기 전반에 걸쳐 존재했던 멸종된 해양 파충류로, 긴 몸체와 노와 같은 꼬리를 특징으로 한다. 탈라토사우루스류는 짧고 넓은 사지, 높은 신경 가시, 그리고 해양 생활에 적응된 두개골을 가지고 있으며, 아스켑토사우루스상과와 탈라토사우루스상과로 분류된다. 이들은 주로 해안 근처의 따뜻한 물에서 서식하며, 다양한 해양 동물을 섭취했을 것으로 추정된다. 초기에는 두판류와 유사하다고 여겨졌으나, 현재는 이궁류로 분류되며, 분류학적 위치는 불확실하다.
더 읽어볼만한 페이지
- 1904년 기재된 화석 분류군 - 켄트로사우루스 (각룡)
켄트로사우루스는 백악기 후기 북아메리카에 서식한 6m 크기의 켄트로사우루스아과 각룡류 공룡으로, 코 위의 뿔과 프릴 가장자리의 작은 뿔이 특징이며 무리 생활을 했을 것으로 추정되지만, 종과 표본 간 변이와 좁은 지리적 분포로 인해 분류학적 논쟁이 있다. - 1904년 기재된 화석 분류군 - 다테오사우루스
다테오사우루스는 페름기에 살았던 멸종된 파충류 속으로, 앞다리가 뒷다리보다 강하고 꼬리가 긴 특징을 가지며 잡식성이었고, 폴란드에서 화석이 발견되어 기저 케이스과에 속하는 것으로 밝혀졌다. - 기룡 - 노토사우루스
노토사우루스는 반 해양성 파충류로, 물개와 유사한 생활 방식을 가졌으며 물고기를 잡아먹고 플레시오사우루스류로 진화했을 것으로 여겨진다. - 기룡 - 란타놀라니아
란타놀라니아는 페름기부터 트라이아스기 초기에 생존했던 디아피드 파충류의 멸종된 속으로, 러시아에서 화석이 발견되었으며, 긴 목과 노와 같은 다리를 가진 기룡으로 바다 생활에 적응했다. - 아시아의 고대 파충류 - 스켈로코무스
스켈로코무스는 트라이아스기 후기에 동남아시아 서부 태평양에 서식했던 멸종된 해양 파충류로, 좁고 긴 몸통, 작은 머리, 뾰족한 이빨을 지녔으며 흡입 섭식 방식으로 먹이를 섭취했을 것으로 추정되는 코부리류 어룡이다. - 아시아의 고대 파충류 - 아토포덴타투스
아토포덴타투스는 특이한 이빨을 가진 2.75~3m 크기의 중생대 초식성 해양 파충류로, 망치 모양 머리에 촘촘한 끌 모양 이빨을 사용하여 해저 조류를 긁어모아 먹었으며 육지 활동 가능성도 시사하는 독특한 골격 구조를 지녔다.
2. 특징
탈라토사우루스류는 해양 생활에 적응한 다양한 신체적 특징을 가지고 있다. 이들은 긴 노와 같은 꼬리와 가느다란 몸을 가지고 있으며, 수영에 적합한 형태를 갖추고 있다.
탈라토사우루스는 이궁류 파충류로, 머리 뒤쪽에 두 개의 측두창이 있는 것이 특징이다. 그러나 일부 탈라토사우루스는 흔적만 남은 위쪽 측두창을 가지거나, 아예 닫혀 있는 경우도 있다. 또한, 아래쪽 측두창은 아래에서 열려 있으며, 사각관골이 없다. 두개골의 뒤쪽 가장자리에는 큰 삼각형 모양의 만입부가 있다.[1]
탈라토사우루스류는 육상 이동에 적합하지 않은 짧고 넓은 사지 뼈를 가지고 있다. 상완골은 어깨 근처에서 가장 넓고, 넙다리뼈는 무릎 근처에서 가장 넓으며, 요골은 콩팥 모양이다. 손가락뼈는 길고 판 모양이다.[1]
탈라토사우루스는 해양 퇴적물에서만 발견되기 때문에, 이들이 주로 수생 파충류였음을 알 수 있다. 움푹 들어간 콧구멍과 길고 노 모양의 꼬리는 수생 생활에 대한 추가적인 증거이다. 또한, 짧고 넓은 사지, 발목과 발목뼈의 발달 부진, 측면 파동을 통한 수영에 적응된 키가 큰 척추뼈를 가지고 있어, 물 속에서 모든 시간을 보냈을 것으로 보인다.[3][32][21][26][4]
탈라토사우루스는 다양한 먹이를 먹었을 것으로 추정되는데, 아마도 모두 어떤 식으로든 해양 동물을 섭취했을 것이다. 엔데나사우루스는 이빨이 없어 어린 물고기나 작은 갑각류와 같은 작은 동물을 잡아먹었을 것으로 보인다.[21] 일부 탈라토사우루스류는 입 앞에 큰 송곳니와 입 뒤쪽에 두껍고 단추 모양의 이빨을 가지고 있어, 갑옷을 입은 물고기, 큰 갑각류, 얇은 껍질의 암모나이트 등을 먹었을 것으로 추정된다.[5][10][6] 구나카데이트의 가느다란 이빨은 부드럽고 빠르게 움직이는 물고기와 오징어를 먹었을 가능성이 있음을 시사하며, 큰 설골을 가지고 있어 흡입 섭식에 역할을 했을 수도 있다.[26]
2. 1. 골격
탈라토사우루스는 긴 노 모양의 꼬리와 20개 이상의 등뼈를 가진 가느다란 몸을 가지고 있어 해양 생활에 적합하다. 모든 탈라토사우루스들이 공유하는 두개골 이후 골격의 독특한 특징은 거의 없지만, 이 골격은 아스켑토사우루스류와 탈라토사우루스류를 구별하는 데 유용하다. 아스켑토사우루스류는 짧은 신경 가시를 가진 긴 목과 최소 11개의 척추뼈를 특징으로 하는 반면, 탈라토사우루스류는 때로는 4개의 척추뼈만을 가지는 짧은 목을 가지고 있다.탈라토사우루스류는 목, 등, 특히 꼬리 척추에 높은 신경 가시를 가지고 있어 횡방향 굴절을 통해 수영할 수 있는 표면적이 증가한다. 또한, 탈라토사우루스류는 짧고 넓은 사지뼈를 가지고 있어 육상 이동에는 적합하지 않다. 상완골은 어깨 부근이 가장 넓고, 넙다리뼈는 무릎 부근이 가장 넓으며, 요골은 콩팥 모양이다. 손가락뼈는 길고 판 모양이다. 아스켑토사우루스류는 육상 파충류와 같은 모래시계 모양의 사지뼈를 유지하지만, 짧은 경골과 종아리뼈와 같은 탈라토사우루스류의 특수성을 공유하며, 종아리뼈는 발목 근처까지 확장되어 있다.[1]
2. 2. 두개골
탈라토사우루스는 이궁류 파충류로, 머리 뒤쪽의 안와(눈 구멍)에 두 개의 측두창이 있다는 것을 의미한다. 그러나 많은 탈라토사우루스는 흔적적인 위쪽 측두창을 가지고 있는데, 이는 틈새와 같으며, 어떤 것들은 주변 뼈에 의해 완전히 닫혀 있다.[19] 탈라토사우루스는 사각관골이 없어 아래쪽에서 아래 측두창이 열려 있다. 또한 후두정골과 탁상골이 없으며, 린상골은 작고, 상측두골은 넓으며, 사골은 크다. 위에서 보면, 두개골의 뒤쪽 가장자리는 사골보다 더 앞으로 뻗어 있는 큰 삼각형의 만입부를 가지고 있다.[1]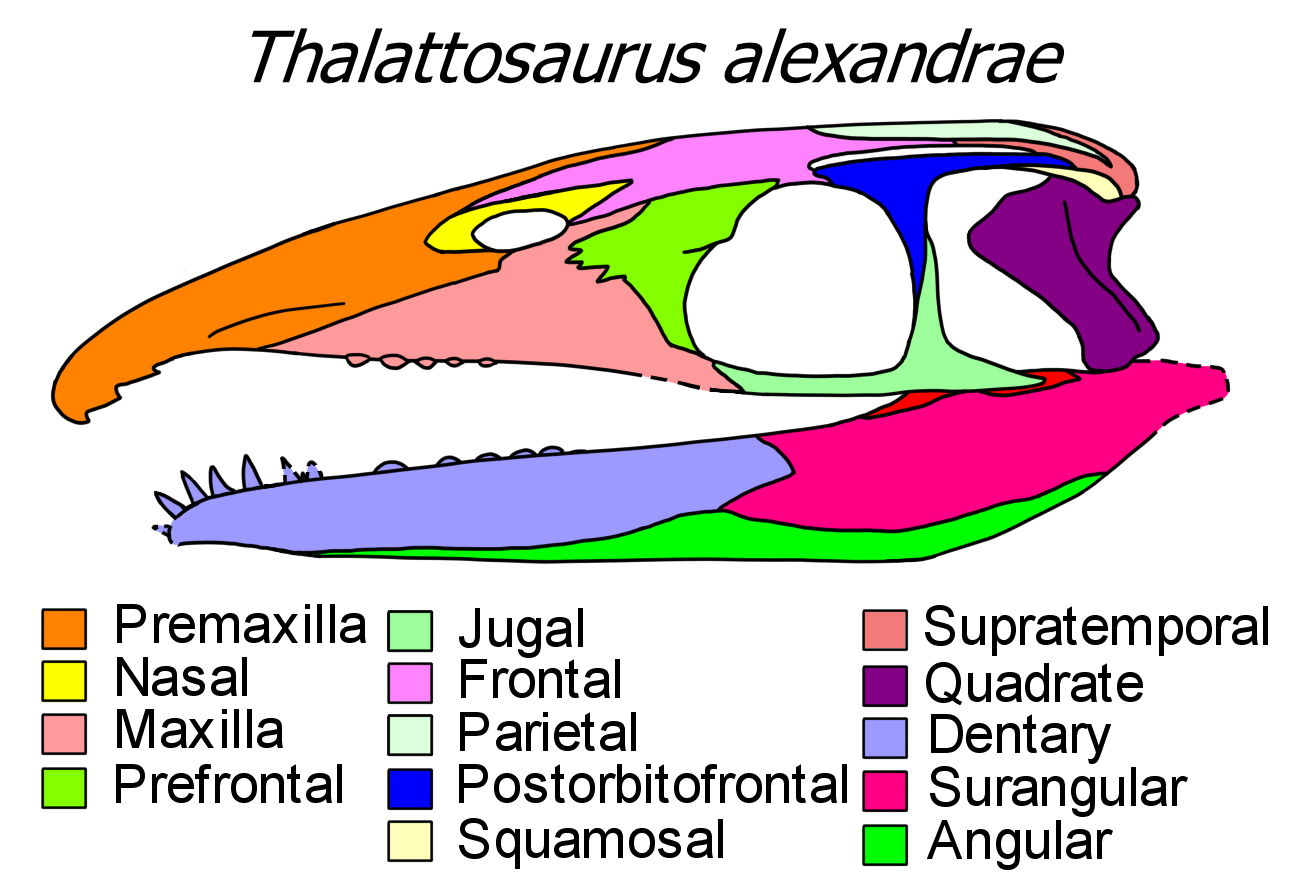
탈라토사우루스는 눈 뒤쪽 두개골 부분보다 현저하게 더 긴 로스트럼(주둥이)을 가지고 있다. 이 길이의 대부분은 전상악골에 의해 형성되며, 콧구멍은 눈 가까이 뒤로 이동한다. 전상악골은 매우 멀리 뻗어 있으며 전두골에 절개되어 있다. 이것은 탈라토사우루스의 특징인 특이한 특징으로 이어지며, 좌우 비골은 서로 분리되어 콧구멍 근처의 작은 부분으로 제한된다. 누골은 일반적으로 손실되거나 안와 앞쪽의 큰 초승달 모양 전전두골에 융합되며, 안와 뒤쪽의 세 갈래 후안와골에 일반적으로 융합된 후전두골을 반영한다.[19][2][1]
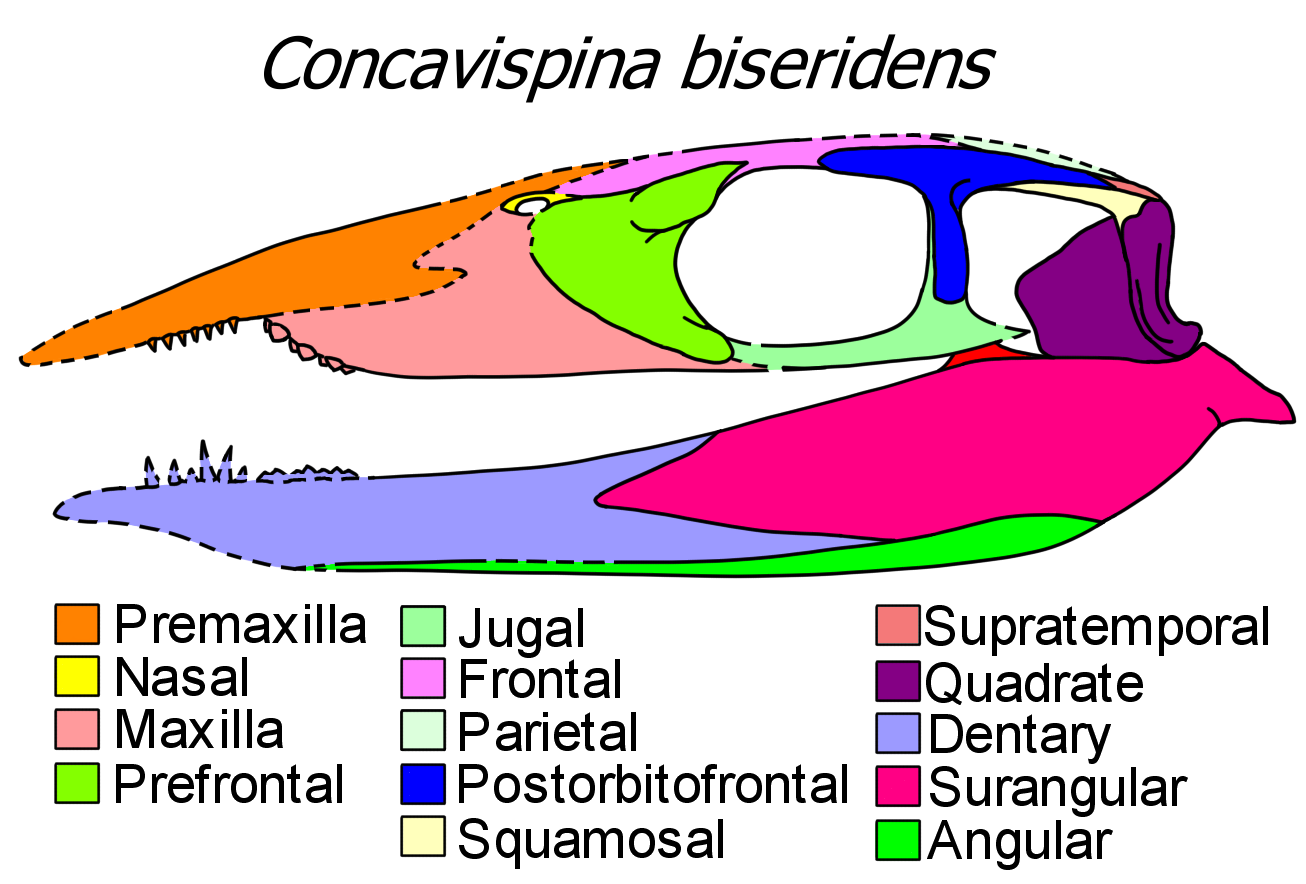
아스케토사우로이데아는 좁고, 가장자리가 직선인 주둥이를 가지고 있으며, 종종 길쭉하고 원뿔형 이빨로 채워져 있다. 한 아스케토사우로이드인 ''엔데나사우루스''(Endennasaurus)는 이빨이 전혀 없고[21], 또 다른 종인 ''미오덴토사우루스''(Miodentosaurus)는 짧고 뭉툭한 주둥이를 가지고 있다.[24] 두 번째 탈라토사우루스 그룹인 탈라토사우로이데아의 대부분 구성원은 더 뚜렷하게 아래로 굽은 주둥이를 가지고 있다. ''클라라지아''(Clarazia)와 ''탈라토사우루스''(Thalattosaurus)는 모두 좁은 끝으로 뾰족해지는 주둥이를 가지고 있다. 주둥이의 대부분은 직선이지만, 팁의 전상악골은 아래로 굽어 있다. ''신푸사우루스''(Xinpusaurus)와 ''콘카비스피나''(Concavispina)도 아래로 굽은 전상악골을 가지고 있지만, 상악골의 끝은 날카롭게 위로 구부러져 두개골에 노치를 형성한다. ''헤셸레리아''(Hescheleria) (그리고 잠재적으로 ''넥토사우루스''(Nectosaurus)와 ''파라로넥테스''(Paralonectes))에서 전상악골은 주둥이 끝에서 갑자기 아래로 굽어 턱의 나머지 부분과 거의 직각을 이룬다. 이러한 형태에서 주둥이 끝은 턱의 나머지 부분과 이개라고 하는 공간으로 분리된 이빨이 있는 갈고리이다. 탈라토사우로이드는 또한 이질치를 가지고 있어 주둥이 앞쪽에 뾰족한 관통 이빨과 뒤쪽에 낮은 으깨는 이빨이 있다.[2] 이 규칙의 예외는 곧은 주둥이와 많은 가느다란 이빨을 가진 ''구나카데이트''(Gunakadeit)이다.[26] 탈라토사우루스는 종종 하악골 뒤쪽에 뚜렷한 후관절 돌기를 가지고 있다. 탈라토사우로이드는 턱 해부학에서 아스케토사우로이드보다 더 전문화되어 있는데, 큰 봉우리 모양의 관상골과 턱의 아래쪽 가장자리를 따라 멀리 뻗어 있는 각골을 진화시켰기 때문이다. 구개 치아는 탈라토사우로이드에서 광범위하지만 아스케토사우로이드에서는 존재하지 않는다.[10][26]
3. 고생물학
탈라토사우루스는 긴 노 모양의 꼬리와 20개 이상의 등뼈를 가진 가느다란 몸 등 해양 생활에 적합한 신체 구조를 가지고 있었다. 모든 탈라토사우루스류가 공유하는 두개골 이후 골격의 특징은 거의 없지만, 이 골격은 아스켑토사우루스류와 탈라토사우루스류를 구별하는 데 유용하다. 아스켑토사우루스류는 짧은 신경 가시와 최소 11개의 척추뼈를 가진 긴 목을 특징으로 하는 반면, 탈라토사우루스류는 때로는 4개의 척추뼈만으로 구성된 짧은 목을 가지고 있다.
탈라토사우루스류는 목, 등, 특히 꼬리 척추에 높은 신경 가시가 있어 측면 파동을 통해 수영할 수 있는 표면적이 증가했다. 또한, 탈라토사우루스류는 짧고 넓은 사지 뼈를 가지고 있어 육상 이동에는 적합하지 않았다. 상완골은 어깨 근처에서 가장 넓고, 넙다리뼈는 무릎 근처에서 가장 넓으며, 요골은 콩팥 모양이고, 손가락뼈는 길고 판 모양이다. 반면 아스켑토사우루스류는 육상 파충류와 유사한 모래시계 모양의 사지 뼈를 유지했지만, 짧은 경골과 종아리뼈 등 탈라토사우루스류와 특수성을 공유하며, 종아리뼈는 발목 근처까지 확장되어 있다.[1]
1980년대 후반 분류학의 인기가 높아지면서 탈라토사우루스 분류에도 일부 영향이 있었다. 지속적인 연구를 통해 사우리아 (살아있는 모든 파충류를 포함하는 주요한 분지군인 이궁류)가 페름기에 레피도사우로모르파(도마뱀, 뱀, 투아타라로 이어짐)와 궁형류 (악어와 조류를 포함한 공룡으로 이어짐)의 두 분기로 나뉘는 등 파충류 분류의 일부 측면이 확고해지는 데 기여했다. 많은 고생물학자들이 여전히 탈라토사우루스를 레피도사우로모르파로 보고 있지만, 일부 연구(예: 에반스의 계통발생 분석, 1988)는 대신 탈라토사우루스가 사우리아의 궁형류 계통에 속할 수 있음을 시사했다.[1] 리에펠(1998)은 탈라토사우루스류인 파키플레로사우루스 ''하노사우루스''를 재평가하여, 탈라토사우루스가 수생 파충류 목인 사우롭테리기아와 친연성이 있으며, 이 자체는 레피도사우로모르파의 광범위한 해석 내에서 거북이와 관련이 있다고 주장했다.[11]
뮐러(2004)의 분석은 탈라토사우루스가 사우리아 바로 바깥에 속한다고 보기까지 했다. 특이하게도 탈라토사우루스는 특정 기저 사우리아 또는 근사우리아가 데이터 세트에서 제외될 경우 어룡류(어룡형류 그룹)와 친연성을 보인다.[12] 뮐러(2004)에서 파생된 일부 분석에서는 탈라토사우루스를 어룡형류 및 사우롭테리기아류와 함께 "해양 상위 분지군"으로 묶으며, 때로는 거북이, 궁형류 또는 레피도사우로모르파와 함께 묶기도 한다. 그러나 이러한 분석에서 생성된 분지도는 방법론을 변경(예: 수생 적응 여부 포함 또는 제외, 최대 절약법과 베이즈 추론 간의 전환)함에 따라 예측할 수 없는 방식으로 바뀌며, 일부에서는 "해양 상위 분지군"의 타당성에 대한 우려를 제기하고 있다.[13][14][15][16][17] 탈라토사우루스가 거의 확실히 이궁류인 반면, 외부군 관계에 대한 불확실성이 매우 커서 대부분의 현대 고생물학자들은 이들을 이궁류 ''incertae sedis''(불확실한 위치)로 분류하고 있다.
3. 1. 수생 생활 방식
움푹 들어간 콧구멍과 길고 노와 같은 모양의 꼬리는 탈라토사우루스가 수생 생활을 했다는 것을 보여주는 증거이다. 탈라토사우루스류는 짧고 넓은 사지, 발목과 발목뼈의 발달 부진, 측면 파동을 통해 헤엄치는 데 적합한 높은 척추뼈를 가지고 있어, 물 속에서 모든 시간을 보냈을 것으로 보인다. 그럼에도 불구하고, 이들은 어룡과 사우로프테리지아와는 다르게 지느러미로 변형되지 않은 강한 발톱과 기능적인 손가락을 유지했다.[3][32][21][26][4]다른 해양 파충류와는 달리, 탈라토사우루스가 대양에서 완전히 원양 생활에 적응했다는 증거는 없으며, 대신 모두 해안 가까운 따뜻한 물에서 살았을 것이다. 아스켑토사우루스류는 육상 파충류의 전형적인 더 강한 사지를 가지고 있어, 어느 정도 육상에서 움직일 수 있었음을 보여준다. 아스켑토사우루스류는 헤엄칠 때 주로 꼬리를 사용했을 가능성이 있으며, 탈라토사우루스류는 몸과 꼬리를 함께 사용했을 수 있다.[3][32][21][26][4]
3. 2. 식성
탈라토사우루스는 다양한 먹이를 먹었지만, 아마도 모두 어떤 식으로든 해양 동물을 먹이로 삼았을 것이다. 엔데나사우루스는 이빨이 없어서 어린 물고기나 작은 갑각류와 같은 작은 동물을 잡아먹었을 것이다.[21]탈라토사우루스, 신푸사우루스, 콘카비스피나와 같은 다양한 탈라토사우루스류는 입 앞에는 큰 송곳니가 있었고, 입 뒤쪽에는 두껍고 단추 모양의 이빨이 있었다. 매사레(1987년)[5]가 제시한 이빨 모양과 먹이 종류 간의 관계를 설명하는 기술을 보면, 더 키가 큰 이빨은 갑옷을 두른 물고기, 큰 갑각류, 얇은 껍데기를 가진 암모나이트를 포함하는 "으깨는" 식단에 적합했다. 낮고 튼튼한 이빨은 큰 연체동물이나 기타 두꺼운 껍데기를 가진 먹이를 전문적으로 먹는 "부수는" 식단에 유용했을 것이다.[10][6]
구나카데이트의 가느다란 이빨은 매사레(1987년)의 "피어스 II" 길드와 관련이 있는데, 이는 부드럽고 빠르게 움직이는 물고기와 오징어를 먹었을 가능성이 있음을 시사한다. 구나카데이트는 또한 큰 설골을 가지고 있었는데, 이것은 흡입 섭식에 역할을 했을 수도 있다.[26]
탈라토사우루스는 다른 해양 파충류의 먹이가 되기도 했다. 약 4미터(13피트) 길이의 신푸사우루스 싱이엔시스의 몸통이 포식성 어룡인 구이저우익티오사우루스의 5미터(16피트) 길이 골격의 체강 내에서 발견되었다. 이것은 해양 파충류 사이에서 가장 오래된 포식 관계를 보여주는 것이며, 신푸사우루스는 다른 해양 파충류 내부에 보존된 가장 큰 먹이일 수도 있다.[7]
4. 분포
탈라토사우루스는 트라이아스기 동안 전 세계적으로 분포했다. 이들은 판탈라사와 테티스 해의 해안을 따라 살았으며, 화석은 중국, 유럽, 북아메리카 등지에서 발견되었다.[10] 특히, 아스켑토사우루스와 탈라토사우루스는 테티스 해에서 발견되었고, 반수생의 물고기를 먹는 동물로, 상당한 지상 능력을 가졌을 가능성이 있다. 반면 탈라토사우루스는 수생 생활에 더 적합했으며, 아래로 쳐진 주둥이와 촘촘한 이빨을 가지고 있었다.[10]
4. 1. 기원
트라이아스기 동안 지구에는 판게아라는 하나의 거대한 초대륙이 있었고, 그 주변에는 판탈라사라는 초대양이 있었다. 판게아의 동쪽 부분에는 중국에서 서유럽까지 뻗어 있는 거대한 열대 내해인 테티스 해가 있었다. 탈라토사우루스류는 전 세계의 트라이아스기 해양 퇴적물에서 발견되지만, 아스켑토사우루스류는 테티스 해 퇴적물에서만 발견된다.[10] 아스켑토사우루스과는 현재 유럽의 알프스 지역인 서테티스 해에서 기원했을 가능성이 있다. 탈라토사우리아의 자매군은 여전히 논쟁의 대상이지만, 가능한 가능성 중 하나인 어룡형류는 초기 트라이아스기 또는 그 이전에 동테티스(중국)에서 진화한 것으로 보인다.[10]4. 2. 확산
가장 오래된 탈라토사우루스류는 현재 북아메리카 서해안을 따라 동쪽 판탈라사에 살았다. 탈라토사우루스류의 적어도 한 갈래가 진화 초기에 전 세계로 확산되었을 가능성이 있다.[26] 탈라토사우루스류의 전 세계적인 분포는 탈라토사우루스가 열린 바다를 횡단하는 데 적응력이 부족한 것으로 여겨진다는 점을 고려할 때 흥미롭다. 열도와 가이요트와 같은 해안 "피난처"는 탈라토사우루스가 해양 분지 사이로 확산되는 것을 용이하게 했을 수 있다.[26] 탈라토사우루스류는 멸종하기 전에 여러 번 주요 해양 지역으로 흩어질 수 있었으며, 탈라토사우루스류는 아스켑토사우루스류보다 수생 적응력이 더 뛰어나기 때문에 확산에 더 능숙했을 것이다.[26]5. 분류
탈라토사우루스류는 긴 노들 모양의 꼬리와 20개가 넘는 등뼈를 가진 가느다란 몸 등 해양 생활에 적합한 특징을 가지고 있다. 모든 탈라토사우루스류가 공유하는 후두골의 독특한 특징은 거의 없지만, 골격은 아스켑토사우루스과와 탈라토사우루스과를 구별하는 데 유용하다. 아스켑토사우루스과는 짧은 신경 가시와 최소 11개의 척추뼈를 가진 긴 목을 특징으로 하는 반면, 탈라토사우루스과는 때로는 4개의 척추뼈를 수반하는 짧은 목을 가지고 있다. 탈라토사우루스류는 목, 등, 특히 꼬리 척추에 키 큰 신경 가시가 있어 횡방향 굴절을 통해 수영할 수 있는 표면적이 증가한다. 또한 짧고 넓은 사지뼈는 육지 이동에 적합하지 않다.
탈라토사우루스류는 이궁류 파충류로, 눈 소켓 뒤쪽 머리에 두 개의 구멍인 시간적 페네스트라(fenestrae)가 있다는 것을 의미한다. 그러나 많은 탈라토사우루스류는 슬릿처럼 생긴 상측두측두측두엽을 가지고 있으며, 일부는 주변 뼈에 의해 완전히 닫혔다. 하악 측두골은 아래에서 열려 있고, 후두골과 표골뼈가 부족하며, 편평골은 작고 초석골은 넓으며, 사분골은 크다. 위에서 보았을 때 두개골 뒤쪽 가장자리에는 사분면보다 더 앞쪽으로 뻗은 삼각형 모양의 큰 전각이 있다.[3]
고생물학자들은 탈라토사우루스과의 분류에 대해 혼란스러워했다. 탈라토사우루스과는 트라이아스기 중기에서 후기까지 살았던 해양 파충류의 멸종된 목(目)이다. 탈라토사우루스류는 크기와 형태가 다양했으며, 두 상악골로 나뉜다. 아스켑토사우루스와 탈라토사우루스는 테티스 해 고유 화석이 유럽과 중국에서 발견되었고, 이들은 반수생의 물고기를 먹는 동물로, 주둥이가 곧고 상당한 지상 능력을 가졌을 가능성이 있다. 탈라토사우루스류는 수생 생활에 더 가까웠으며, 대부분 특이하게 아래로 꺾인 주둥이는 곧았고, 이빨은 고르고 전문화되어 있었다. 탈라토사우루스류는 판탈라사와 테티스 해의 해안을 따라 살았으며, 중국과 북아메리카 서부에서 가장 다양했다. 큰 종은 4미터(13피트)까지 자랐고, 길고 평평한 꼬리는 물속에서 추진하는 데 사용되었다.
탈라토사우루스류와 도마뱀은 외견상 유사하지만, 그들의 정확한 관계는 미해결 상태이다. 그들은 어룡에 가까운 파충류지만, 많은 종들이 뱀목(도마뱀, 뱀)과 두판목(투아타라)과 그들의 친척들을 가리킨다.) 또는 지배파충류(악어, 공룡, 조류)와 그들의 친척들) 전문가들 여러가지로 인룡 하강 사이에서 파충류 가계도에 그들을 장착했다.
탈라토사우루스가 어디에서 유래했는지는 확실하지 않다. 트라이아스기 동안 지구에는 판게아라는 하나의 거대한 초대륙이 있었는데, 판게아는 판탈라사라는 거대한 바다로 둘러싸여 있었다. 판게아의 동부는 거대한 열대 내륙 바다인 테티스 해에 의해 침식되어 중국에서 서유럽까지 확장되었다. 탈라토사우루스류는 전 세계 트라이아스기 해양 퇴적물에서 알려진 반면, 아스켑토사우루스류는 테티스 해 퇴적물에서만 알려져 있다. 엔데나사우루스와 아스켑토사우루스가 가장 기초적인 아스켑토사우루스라고 가정하면, 아스켑토사우루스과는 현재 유럽의 알프스 지역인 서부 테티스 해에서 기원했을 것이다. 그러나 미오덴토사우루스가 더 기초적이라면 서부 테티스(유럽) 기원은 현저히 덜할 것이다. 탈라토사우르시아의 자매 집단은 여전히 논의되고 있지만, 트라이아스기 초기나 그 이전 시기에 동부 테티스(중국)에서 진화한 것으로 보이는 어룡이 한 가지 가능성이다.
가장 오래된 탈라토사우루스류는 현재 북아메리카 서부 해안인 판탈라사 동부에 살았다. 뮐러(2005년, 2007년)는 적어도 탈라토사우루스류의 한 가지가 진화 초기에 전 세계로 퍼져나갈 수 있었다고 주장했다. 그러나 이는 넥토사우루스(캘리포니아 출신), 신푸사우루스(중국 출신), 오스트리아 출신 이름 없는 종족 등이 다른 탈라토사우루스류에 기저를 형성했다는 가설에 근거한 것으로, 다른 많은 연구와 대비되는 분류 체계이다.
탈라토사우루스상과의 전 세계적인 분포는 탈라토사우루스상과가 판탈라사 동부 해안과 테티스 해 사이에 퍼지기 위해 필요했을 탁 트인 대양을 횡단하는 데 부적응한 것으로 간주되고 있다는 점을 고려할 때 흥미롭다. 화산섬 호와 가이오트와 같은 해안 "개척"은 탈라토사우루스류의 해양 분지들 사이에 퍼지는 능력을 촉진시켰을 것이다. 헤스켈레리아와 유사한 형태는 이전에는 북아메리카와 유럽에서만 보고되었으나, 2021년에는 중국에서 헤스켈레리아와 같은 코딱지 파편이 보고되어 그들도 널리 분포하고 있음을 알 수 있었다. 익룡과 암모나이트와 같은 다른 트라이아스기 해양 생물에서도 트랜스-판탈라사 연결이 관찰된다. 분명히 탈라토사우루스류는 그 집단이 멸종되기 전에 여러 번 주요 해양 지역에 분산될 수 있었고, 탈라토사우루스류는 더 큰 수생 적응으로 인해 아스켑토사우루스류보다 더 번식할 가능성이 있었다.
일부 고생물학자들은 탈라토사우루스류를 과(family)로 나누려고 시도했다. 한 과인 아스켑토사우루스과는 전형적으로 아스켑토사우루스와 안순사우루스를 포함하는 것으로 간주되며, 몇몇 연구에서는 미오덴토사우루스나 엔데나사우루스를 그 안에 배치하기도 한다. 또 다른 과인 탈라토사우루스과는 원래 탈라토사우루스와 넥토사우루스를 그룹화하는 데 사용되었고, 나중에 넥토사우루스를 제외하도록 재정의되었으며, 이후에는 사실상 모든 탈라토사우루스상과를 포함시켰다. 많은 탈라토사우루스 중심 고생물학자들은 일관성 없는 사용법과 의심스러운 타당성 때문에 과 이름을 사용하는 것을 기피한다.[19]
다른 탈라토사우루스류로는 오스트리아의 쾨센 층과 브리티시 컬럼비아의 황산과 사면 층, 네바다의 나체즈 고개 층을 기초로 하여 오리건 주의 베스터 층이 있다. 독일 무셀칼크의 단편적인 해양 파충류인 ''Blezingeria''도 일부 작가들에 의해 탈라토사우루스류로 여겨졌지만, 이러한 과제는 기껏해야 불확실하다. 네오시노사우루스와 와야오사우루스는 중국 샤오와 층의 파충류로 잘 알려져 있지 않은 한 쌍의 파충류로, 또한 탈라토사우루스류로 여겨져 왔다. 탈라토사우루스류 파편들은 스페인 무셸칼크로부터 알려져 있다. 이전에 알래스카에서 온 이름 없는 표본은 2020년에 구나카데이트로 묘사되었다.
먹이로는 당대에 서식했던 물고기, 갑각류, 무척추동물을 주로 먹고 살았을 육식성 포식자로 추정된다.
5. 1. 초기 가설
1904년 메리엄이 처음 명명했을 때, 탈라토사우루스류는 ''Thalattosaurus alexandrae'' 종만 알려져 있었다. 주로 두개골 형태를 바탕으로, 살아있는 투아타라인 ''Sphenodon''을 포함하는 파충류 목인 두판류와 가까운 관계라고 추정되었다.
메리엄(1905)은 비슷한 생태 때문에 어룡과의 관계를 고려했지만, 만약 그들이 가까운 공통 조상을 가지고 있다면 두개골과 척추 해부학이 왜 그렇게 넓게 달라지는지에 대한 의문을 제기했다. 그는 잠재적인 유사성은 수렴 진화로 가장 잘 설명된다고 제안했다. 탈라토사우루스가 도마뱀과 가까운 파충류(예: ''Paliguana'')에서 갈라졌을 가능성이 더 자세히 설명되었으며, 탈라토사우루스는 거의 도마뱀이 바다로 돌아가려는 단명한 초기 시도로 작용했으며, 나중에 모사사우루스가 진정한 도마뱀에서 진화했을 때 더 성공적으로 반복된 진화 과정이었다. 그럼에도 불구하고, 메리엄은 이전에 알려진 어떤 파충류 그룹이 탈라토사우루스의 직접적인 조상이거나 그 반대라는 명확한 증거를 찾지 못했다. 그들은 아마도 육지에 사는 페름기 파충류에서 유래했으며, 트라이아스기에 처음 진화한 다른 해양 파충류 그룹과 밀접한 관련이 없었다.[3] 20세기 후반의 연구자들은 일반적으로 탈라토사우루스를 현재 인룡류로 알려진 그룹의 일부로 두판류 또는 뱀류와 가깝게 배치했다.[10]
5. 2. 현대 분류 및 외부 관계
1980년대 후반, 분류학의 인기가 높아지면서 탈라토사우루스 분류에도 영향을 주었다. 지속적인 연구를 통해 파충류 분류의 일부 측면이 확고해졌다. 예를 들어, 사우리아(살아있는 모든 파충류를 포함하는 주요 분지군인 이궁류)는 페름기에 레피도사우로모르파(도마뱀, 뱀, 투아타라로 이어짐)와 궁형류(악어와 조류를 포함한 공룡으로 이어짐)의 두 분기로 나뉘는 것이 밝혀졌다.[3]많은 고생물학자들은 여전히 탈라토사우루스를 레피도사우로모르파로 보고 있지만, 일부 연구에서는 탈라토사우루스가 사우리아의 궁형류 계통에 속할 수 있다고 주장했다. 리펠(1998)은 탈라토사우루스류인 하노사우루스를 재평가하여, 탈라토사우루스가 수생 파충류 목인 사우롭테리기아와 친연성이 있으며, 이 자체는 레피도사우로모르파의 광범위한 해석 내에서 거북과 관련이 있다고 주장했다. 뮐러(2004)의 분석은 탈라토사우루스가 사우리아 바로 바깥에 속한다고 보았다. 특이하게도 탈라토사우루스는 특정 기저 사우리아 또는 근사우리아가 데이터 세트에서 제외될 경우 어룡(어룡형류 그룹)과 친연성을 보인다. 뮐러(2004)에서 파생된 일부 분석에서는 탈라토사우루스를 어룡형류 및 사우롭테리기아류와 함께 "해양 상위 분지군"으로 묶으며, 때로는 거북, 궁형류 또는 레피도사우로모르파와 함께 묶기도 한다. 그러나 이러한 분석에서 생성된 분지도는 방법론을 변경함에 따라 예측할 수 없는 방식으로 바뀌며, 일부에서는 "해양 상위 분지군"의 타당성에 대한 우려를 제기하고 있다.[3][7][8][24][25]
탈라토사우루스가 거의 확실히 이궁류인 반면, 외부군 관계에 대한 불확실성이 매우 커서 대부분의 현대 고생물학자들은 이들을 이궁류 ''incertae sedis''(불확실한 위치)로 분류하고 있다.[3]
5. 3. 내부 관계
Nicholls|니콜스영어(1999)가 탈라토사우루스와 넥토사우루스를 재평가하면서 탈라토사우루스에 초점을 맞춘 최초의 계통 발생학적 분석 중 하나가 이루어졌다. 그녀는 탈라토사우리아에 대한 제한적인 정의를 사용했는데, 이는 엔데나사우루스나 아스켑토사우루스보다 넥토사우루스와 헤스켈레리아에 더 가까운 모든 파충류를 포함하는 분지군을 지칭했다. 아스켑토사우루스, 엔데나사우루스, 그리고 전통적인 탈라토사우루스를 포함하는 더 포괄적인 그룹에는 탈라토사우루스형류(Talattosauriformes)라는 이름이 부여되었다. 그러나 이 그룹에 초점을 맞춘 대부분의 연구에서는 니콜스의 탈라토사우루스형류 분지군과 동등한, 아스켑토사우루스와 탈라토사우루스 모두에 가까운 파충류를 포함하는 더 넓은 정의의 탈라토사우리아를 유지하는 것을 선호했다.[19]이러한 연구에서 탈라토사우리아는 두 개의 분기로 나뉘는데, 하나는 아스켑토사우루스의 친척으로 이어지고 다른 하나는 탈라토사우루스의 친척으로 이어진다. 아스켑토사우루스과보다 탈라토사우루스에 더 가까운 파충류를 포함하는 분지군은 탈라토사우로이데아(Talattosauroidea, 때로는 탈라토사우리데아(Talattosauridea)라고도 함)라고 명명된다.[9][19] 한편, 아스켑토사우루스과에 더 가까운 파충류를 포함하는 분지군은 아스켑토사우로이데아(Askeptosauroidea) 또는 아스케프토사우리데아(Askeptosauridea)라고 불린다.[19]
니콜스(1999) 이후의 후속 연구에서는 안순사우루스, 신푸사우루스와 같은 새로 기술된 중국 분류군을 포함하여 더 많은 분류군을 포함하기 시작했다. 그러나 엔데나사우루스의 탈라토사우루스 계통에 대한 불확실성으로 인해 이러한 분석에서 제외되었다. 뮐러 외(2005)가 엔데나사우루스가 아스켑토사우루스와 밀접한 관련이 있음을 재확인한 후,[12] 당시 알려진 모든 탈라토사우루스는 마침내 계통 발생학적 분석에 통합되었다.
리펠, 류, 청, 우 등의 연구에서는 미오덴토사우루스와 다양한 종의 안순사우루스와 신푸사우루스와 같은 새로운 중국 분류군을 계속 식별했지만, 이러한 새로운 분류군의 동형성으로 인해 탈라토사우리아의 두 주요 분기 구조에 대한 해상도가 거의 이루어지지 않았다. 이러한 문제를 해결하기 위해, 콘카비스피나를 기술하는 동안 류 외(2013)에서 새로운 계통 발생학적 분석이 개발되었고, 구나카데이트를 기술하는 동안 드루켄밀러 외(2020)에서 개발되었다.
탈라토사우루스의 내부 관계는 아직 잠정적이고 결정적이지 않은 것으로 간주되지만, 그룹의 기본 구조(아스켑토사우루스과와 탈라토사우루스과로 나뉜 단일 계통의 탈라토사우리아 분지군)는 매우 안정적이다. 일부 고생물학자들은 탈라토사우루스를 과로 나누려고 시도했다. 한 과인 아스켑토사우루스과는 일반적으로 아스켑토사우루스와 안순사우루스를 포함하는 것으로 간주되며, 일부 연구에서는 미오덴토사우루스[13] 또는 엔데나사우루스[12]를 그 안에 포함하기도 한다. 또 다른 과인 탈라토사우루스과는 원래 탈라토사우루스와 넥토사우루스를 그룹화하는 데 사용되었으며, 나중에 넥토사우루스를 제외하도록 재정의되었고, 이후에는 사실상 모든 탈라토사우로이드를 포괄했다. 탈라토사우루스에 초점을 맞춘 많은 고생물학자들은 일관성 없는 사용법과 의심스러운 유효성 때문에 과 이름을 사용하는 것을 피한다.[19]
6. 속 목록




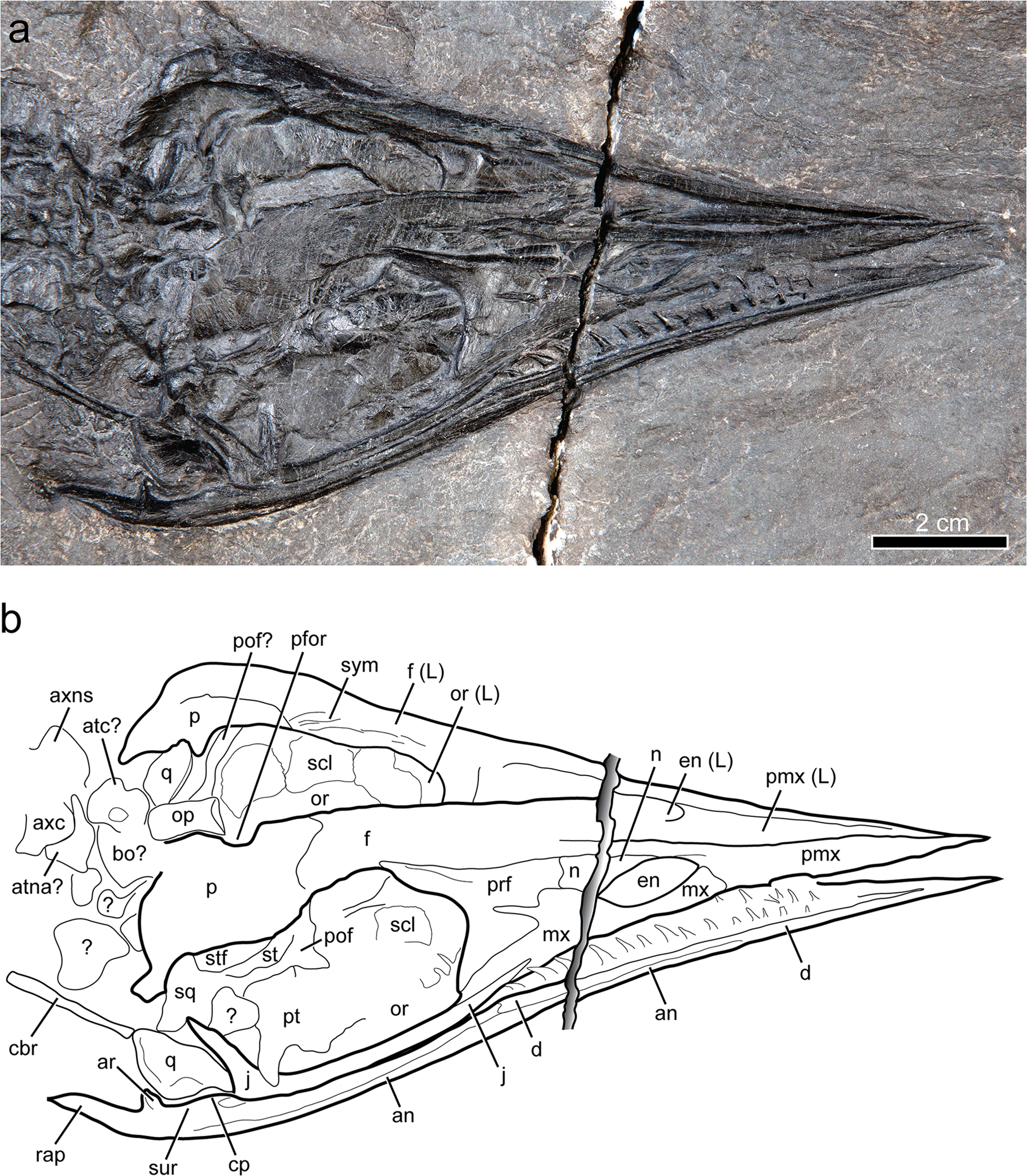




참조
[1]
서적
The Phylogeny and Classification of the Tetrapods, Volume 1: Amphibians, Reptiles, Birds
Clarendon Press
[2]
논문
Rostral structure in Thalattosauria (Reptilia, Diapsida)
[3]
논문
The Thalattosauria: a group of marine reptiles from the Triassic of California
https://archive.org/[...]
[4]
논문
Footprints of marine reptiles from the Middle Triassic (Anisian-Ladinian) Guanling Formation of Guizhou Province, southwestern China: The earliest evidence of synchronous style of swimming
https://www.scienced[...]
2020-11-15
[5]
논문
Tooth morphology and prey preference of Mesozoic marine reptiles
1987-06-18
[6]
논문
A new species of Xinpusaurus (Reptilia, Thalattosauria) from the Ladinian (Middle Triassic) of Xingyi, Guizhou, southwestern China
https://figshare.com[...]
2016-11-01
[7]
논문
Evidence Supporting Predation of 4-m Marine Reptile by Triassic Megapredator
2020-09-25
[8]
논문
A New Specimen of Thalattosauroidea (Reptilia, Thalattosauriformes) from the Middle Triassic (Ladinian) of Xingyi, Southernwestern China
https://bioone.org/j[...]
2021-06-04
[9]
논문
A new marine reptile from the Triassic of California
https://archive.org/[...]
1904
[10]
논문
A reexamination of ''Thalattosaurus'' and ''Nectosaurus'' and the relationships of the Thalattosauria (Reptilia: Diapsida)
https://docubase.ber[...]
1999-04-15
[11]
논문
The systematic status of Hanosaurus hupehensis (Reptilia, Sauropterygia) from the Triassic of China
1998-09-15
[12]
서적
Recent Advances in the Origin and Early Radiation of Vertebrates
Verlag Dr. Friedrich Pfeil
[13]
논문
The Enigmatic Marine Reptile Nanchangosaurus from the Lower Triassic of Hubei, China and the Phylogenetic Affinities of Hupehsuchia
2014-07-11
[14]
논문
A basal ichthyosauriform with a short snout from the Lower Triassic of China
2014-11-05
[15]
논문
A large aberrant stem ichthyosauriform indicating early rise and demise of ichthyosauromorphs in the wake of the end-Permian extinction
2016-05-23
[16]
논문
A new, exceptionally preserved juvenile specimen of Eusaurosphargis dalsassoi (Diapsida) and implications for Mesozoic marine diapsid phylogeny
2017-06-30
[17]
논문
Successive climate crises in the deep past drove the early evolution and radiation of reptiles
2022-08-19
[18]
논문
Osteology of ''Concavispina biseridens'' (Reptilia, Thalattosauria) from the Xiaowa Formation (Carnian), Guanling, Guizhou, China
https://www.academia[...]
[19]
논문
The first record of a thalattosaur reptile from the Late Triassic of southern China (Guizhou Province, PR China)
https://www.research[...]
2000-09-25
[20]
논문
A new species of Xinpusaurus (Thalattosauria) from the Upper Triassic of China
https://www.research[...]
2004-03-25
[21]
논문
The marine diapsid reptile ''Endennasaurus'' from the Upper Triassic of Italy
2005
[22]
논문
Restudy of ''Anshunsaurus huangguoshuensis'' (Reptilia: Thalattosauria) from the Middle Triassic of Guizhou, China
http://digitallibrar[...]
[23]
논문
First record of a thalattosaur from the Upper Triassic of Austria
https://www.research[...]
2007-03-12
[24]
논문
''Miodentosaurus brevis'' Cheng et al. 2007 (Diapsida: Thalattosauria): Its postcranial skeleton and phylogenetic relationships
https://www.research[...]
2009
[25]
논문
New Study of Anshunsaurus huangnihensis Cheng, 2007 (Reptilia: Thalattosauria): Revealing its Transitional Position in Askeptosauridae
2011
[26]
논문
An articulated Late Triassic (Norian) thalattosauroid from Alaska and ecomorphology and extinction of Thalattosauria
2020-02-04
[27]
논문
New thalattosaurs (Reptilia: Diapsida) from the Triassic Sulphur Mountain Formation of Wapiti Lake, British Columbia
https://www.research[...]
1993-03-01
[28]
논문
Note on a second occurrence of thalattosaur remains (Reptilia: Neodiapsida) in British Columbia
https://www.research[...]
1991-12-01
[29]
논문
Thalattosaurs from the Late Triassic (Carnian) of Nevada and their paleobiogeographic significance
http://www.miketaylo[...]
2005
[30]
논문
A new thalattosaur from the Vester Formation (Carnian) of Central Oregon, USA
http://vertpaleo.org[...]
2015
[31]
웹사이트
Triassic Reptile Skewered Clams with Teeth on Roof of Its Mouth
https://www.livescie[...]
2015-11-13
[32]
논문
The anatomy of ''Askeptosaurus italicus'' from the Middle Triassic of Monte San Giorgio, and the interrelationships of thalattosaurs (Reptilia, Diapsida)
https://www.research[...]
[33]
논문
Fossil reptiles from the Spanish Muschelkalk (mont-ral and alcover, province Tarragona)
https://www.research[...]
[34]
뉴스
Recent fossil discovery near Keku Strait a first for Alaska
http://www.capitalci[...]
2011-07-13
[35]
논문
The relationships and paleoecology of ''Pachystropheus rhaeticus'', an enigmatic latest Triassic marine reptile
2024-06-04
[36]
논문
To glide or to swim? A reinvestigation of the enigmatic Wapitisaurus problematicus (Reptilia) from the Early Triassic of British Columbia, Canada
2023-11
[37]
논문
Reidentification of Wayaosaurus bellus and the conservative trunk and tail shape of Thalattosauria
https://www.tandfonl[...]
2023-10-23
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com