좌우대칭동물
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
좌우대칭동물은 몸의 좌우가 대칭을 이루는 동물을 의미하며, 입과 항문, 내장을 가진 원통형 몸체를 특징으로 한다. 머리와 꼬리가 구분되며, 머리에는 감각 기관이 발달해 있다. 좌우대칭동물은 화석 기록을 통해 에디아카라기에서 처음 발견되었으며, 초기 좌우대칭동물은 벌레 형태를 보였다. 좌우대칭동물은 전통적으로 후구동물과 전구동물로 분류되어 왔으나, 최근에는 무체강동물과 신장동물의 관계를 통해 새로운 계통 분류가 제시되고 있다.
더 읽어볼만한 페이지
- 동물 - 진정후생동물
진정후생동물은 방사대칭동물군과 좌우대칭동물군으로 나뉘며, 좌우대칭동물군은 편형동물상문, 촉수담륜동물상문, 탈피동물상문, 후구동물상문으로 분류된다. - 동물 - 측생동물
측생동물은 비대칭적 특징을 가진 동물군으로, 해면동물 및 판형동물과 함께 연구되고 있으며, 분자생물학적 연구 결과 좌우대칭동물과의 연관성이 제시되고 있다. - 생물 계통도에 관한 - 눈표범
눈표범은 고양이과 동물로 중앙아시아와 남아시아의 고산 지대에 서식하며, 길고 두꺼운 털, 큰 발, 긴 꼬리가 특징이며, 유전자 분석 결과 호랑이와 가장 가까운 종으로 밝혀졌으며, 밀렵과 서식지 파괴로 인해 개체 수가 감소하여 여러 국가에서 보호 노력이 진행 중이다. - 생물 계통도에 관한 - 벵골호랑이
벵골호랑이는 인도아대륙에서 서식하는 호랑이 아종으로, 노란색~주황색 털과 검은 줄무늬를 가지며, 멧돼지, 사슴 등 우제류를 먹이로 하며, 서식지 파괴와 밀렵으로 인해 제한적인 지역에서 서식한다. - 생물 분류에 관한 - 다람쥐
다람쥐는 등 쪽에 줄무늬가 있는 다람쥐속 설치류로, 홀로 생활하며 겨울잠을 자고 씨앗, 견과류, 곤충 등을 먹으며 맹금류 등의 먹이가 되고, 특히 한반도 서식 다람쥐는 새로운 종으로 분류될 가능성이 있으며, 유럽에서는 침입 외래종으로 지정되기도 하고 라임병을 옮길 수 있어 주의가 필요한 동물이다. - 생물 분류에 관한 - 황금랑구르
황금랑구르는 인도 아삼 주와 부탄에 분포하며 크림색에서 황금색 털을 가진 멸종위기종 영장류로, 서식지 파괴로 개체수가 감소하여 보호받고 있다.
| 좌우대칭동물 - [생물]에 관한 문서 | |
|---|---|
| 기본 정보 | |
 | |
| 학명 | Bilateria |
| 명명자 | Hatschek, 1888 |
| 동의어 | 삼배엽성 동물 (Lankester, 1873) |
| 로마자 표기 | Bilateria (바일러티어리아) |
| 로마자 표기 (단수형) | Bilaterian (바일러티어리언) |
| 의미 | 좌우대칭을 가진 동물 |
| 생물 분류 | |
| 계 | 동물계 |
| 계급 없음 | 진정후생동물 |
| 계급 없음 | (미분류) 파라혹소아 |
| 하위 계급 없음 | 좌우대칭동물 |
| 학명 | Bilateria |
| 명명자 | Hatschek, 1888 |
| 일본어 명칭 | 좌우상칭동물 (さゆうそうしょうどうぶつ) |
| 한국어 명칭 | 좌우대칭동물 (좌우대칭동물) |
| 분류 | |
| 하위 분류 | |
| 아문 | 원구동물 (Proarticulata) † 후구동물 (Deuterostomia) 선구동물 (Protostomia) |
| 후구동물 | |
| 하위 분류 | 척삭동물 (Chordata) 암불라크라리아 (Ambulacraria) |
| 선구동물 | |
| 하위 분류 | 탈피동물 (Ecdysozoa) 나선동물 (Spiralia) |
| 분류 불확실 | |
| 하위 분류 | 아그마타 (Agmata)? † 넥토카리다 (Nectocarididae) † 크세나코엘로모르파 (Xenacoelomorpha) '아르밀리막스' (Armilimax)? † '보웬그리푸스' (Bowengriphus) † '다코르하키스' (Dakorhachis) † '이카리아' (Ikaria) † '케레차' (Keretsa)? † '킴베렐라' (Kimberella) † '미크로스케디아' (Microschedia) † '파르반코리나' (Parvancorina) † '플렉서스' (Plexus) † '폴링게리아' (Pollingeria) † '포르탈리아' (Portalia)? † '프로테키우루스' (Protechiurus) † '프로토님파' (Protonympha) † '루고사이비타' (Rugosusivitta) † '솔자' (Solza) † '템녹사' (Temnoxa) † '툴리몬스트룸' (Tullimonstrum) † '이링기아' (Yilingia) † |
| 형태 및 특징 | |
| 대칭 | 좌우대칭 |
| 층 | 삼배엽성 |
| 진화 역사 | |
| 출현 시기 | 에디아카라기 |
| 화석 기록 | 에디아카라기–현세 (5억 6700만 년 전 – 현재) |
| 설명 | 최초의 좌우대칭동물은 크세노투르벨라와 유사한 해양 벌레였을 것으로 추정된다. |
2. 특징
해면동물과 자포동물을 제외한 대부분의 후생동물이 좌우대칭동물에 포함된다. 초기 다세포동물은 해면과 같이 좌우대칭이 아니거나 비대칭적인 형태였으나, 캄브리아기 대폭발 무렵 많은 동물이 좌우대칭 형태를 갖추게 된 것은 생명 진화 역사에서 중요한 단계였다.[40]
좌우대칭동물은 해면동물과 자포동물을 제외한 대부분의 후생동물을 포함한다. 초기 다세포동물은 비대칭적인 해면 형태였으나, 캄브리아기 대폭발 시기에 많은 동물이 좌우대칭 형태를 갖추게 된 것은 생명 진화 역사에서 중요한 전환점이었다.[40]
대부분의 좌우대칭동물은 몸이 외배엽, 중배엽, 내배엽의 세 배엽으로 구성되는 삼배엽성 동물이다.[40] 이름처럼 몸의 좌우가 대칭 또는 거의 대칭적인 형태를 가진다. 몸의 앞뒤(머리와 꼬리 방향)와 등배(등 쪽과 배 쪽)가 뚜렷이 구분되는 것[10][7]이 특징이며, 이는 몸의 앞쪽 끝이 외부 환경과 먼저 접촉하게 만들어 두부화, 즉 감각 기관과 입이 있는 머리가 발달하는 경향으로 이어진다.[8]
초기 좌우대칭동물 중 일부는 벌레와 유사한 형태였을 것으로 추정되며, 이들의 몸은 입에서 항문으로 이어지는 소화관을 가진 원통형 구조로 볼 수 있다. 소화관 주위에는 체강 또는 위체강이라는 내부 공간이 존재한다.[14] 다만, 편형동물이나 복모동물과 같은 일부 무체강동물은 이차적으로 체강이 소실된 것으로 여겨진다. 가장 오래된 좌우대칭동물 화석 중 하나인 베르나니말큘라에서 체강과 유사한 구조가 발견된 것은 체강이 초기부터 존재했을 가능성을 시사한다. 대부분의 좌우대칭동물은 입에서 항문까지 완전한 소화관을 가지지만, 진무체강형동물처럼 입과 항문의 구분이 없는 주머니 형태의 소화관을 가진 경우도 있다.
많은 좌우대칭동물은 몸을 수축시키는 원형 근육과 몸을 늘리는 세로 근육을 함께 가지고 있다.[7] 이러한 근육 구조는 수정골격(hydrostatic skeleton)을 가진 연체 동물이 연동 운동을 통해 이동할 수 있게 한다.[9] 또한, 많은 좌우대칭동물 문(phylum)은 발생 과정에서 섬모를 이용해 헤엄치고 감각 세포를 포함하는 정단 기관(apical organ)을 가진 원발성 유충 단계를 거친다.
그러나 이러한 일반적인 특징에는 예외도 존재한다. 예를 들어, 극피동물은 유생 시기에는 좌우대칭이지만 성체가 되면 방사대칭 형태를 띤다. 또한, 흡충류와 같은 일부 기생충은 서식 환경에 적응하면서 매우 단순화되거나 변형된 몸 구조를 가지기도 한다.[10][7]
좌우대칭동물은 크게 후구동물과 전구동물이라는 두 개의 상위 분류군으로 나뉜다. 이 둘은 배 발생 과정에서 중요한 차이를 보이는데, 특히 발생 초기에 형성되는 원구(blastopore)가 전구동물에서는 입으로 발달하고, 후구동물에서는 항문으로 발달한다. 현재 전구동물은 다시 탈피동물과 관륜동물 등으로 세분되며, 유형동물과 같이 분류학적 위치가 불확실한 그룹도 존재한다.
3. 진화
초기 좌우대칭동물 중 일부는 벌레와 유사한 형태였을 것으로 추정된다. 좌우대칭동물의 기본적인 몸 구조는 입과 항문, 그리고 이 두 구멍을 잇는 내장을 가진 원통 형태로 설명될 수 있다. 내장 주위에는 내부 공간인 체강이나 유사 체강이 존재한다. 이름처럼 좌우 대칭의 신체 구조를 가지며, 앞과 뒤, 등과 배가 명확히 구분된다.
앞뒤 구분이 생기면서 앞쪽(머리 부분)에는 입, 감각 기관 등이 집중되어 발달하는 두부화(cephalisation) 현상이 나타났다. 몸은 머리에서 뒤쪽으로 뻗어 있으며, 많은 좌우대칭동물은 몸을 수축시키는 원형 근육과 이완시키는 근육의 조합을 이용한다. 이는 유체정역학적 골격을 가진 부드러운 몸체의 동물들이 연동 운동을 통해 움직일 수 있게 한다. 대부분의 좌우대칭동물은 입에서 항문까지 이어지는 완전한 소화관을 가지지만, 진무체강형동물처럼 입과 항문의 구분이 없는 주머니 형태의 내장을 가진 경우도 있다.
대부분의 좌우대칭동물은 몸이 세 개의 배엽(내배엽, 중배엽, 외배엽)으로 구성된 삼배엽성 동물이다.[40] 많은 좌우대칭동물은 체내에 체강이라는 공간을 가지고 있다. 과거에는 체강이 없는 무체강동물이 원시적인 그룹으로 여겨졌으나, 현재는 편형동물이나 복모동물과 같은 주요 무체강동물 문에서 체강이 이차적으로 사라진 것으로 보는 견해가 우세하다.
좌우대칭동물은 발생 과정의 차이에 따라 크게 두 그룹, 즉 후구동물과 선구동물로 나뉜다. 가장 큰 차이점 중 하나는 발생 초기 배아의 첫 번째 구멍인 원구가 입으로 발달하는지(선구동물), 항문으로 발달하는지(후구동물)이다. 현재 선구동물은 다시 탈피동물과 나선동물(또는 관륜동물) 등의 상문으로 분류하는 것이 일반적이다. 유형동물과 같이 분류가 어려운 그룹도 있는데, 과거에는 후구동물로 분류되었으나 현재는 선구동물에 포함시키는 견해가 유력하다.
많은 좌우대칭동물은 섬모를 이용해 헤엄치고 감각 세포를 포함하는 정점 기관을 가진 원발성 유충 단계를 거친다. 그러나 이러한 일반적인 특징에는 예외도 존재한다. 예를 들어, 성체 극피동물은 유충 시기에는 좌우대칭이지만 성장하면서 방사대칭 형태를 띤다. 또한 일부 기생충은 매우 특이하고 변형된 신체 구조를 가지기도 한다.
3. 1. 조상

모든 후구동물의 가상적인 최근 공통 조상을 "우르빌라테리아"라고 한다.[12][13] 최초의 후구동물이 어떤 특징을 가졌는지에 대해서는 여러 의견이 있다. 한 가지 견해는 무체강동물이 다른 그룹으로 진화했다는 플라눌로이드-아셀로이드 가설이다. 이는 루트비히 폰 그라프, 일리야 메치니코프, 리비 하이먼, 또는 Luitfried von Salvini-Plawen 등이 주장했다. 반면, 최초의 후구동물이 체강동물이었고, 주요 무체강동물 문(편형동물과 유형동물)은 나중에 체강을 잃었다는 아르키코엘로마타 가설과 그 변형들도 있다. 예를 들어 에른스트 헤켈이나 아담 세드윅의 가스트레아, Gösta Jägersten의 빌라테로가스트레아, 또는 닐센의 트로카에아 등이 이에 해당한다.
한 가설에 따르면, 원시 후구동물은 ''Xenoturbella''처럼 단일 체강을 가진 바닥에 사는 벌레 형태였을 수 있다.[14] 또는 일부 자포동물의 플라눌라 유생과 비슷했을 수도 있는데, 이 유생은 어느 정도 좌우 대칭 형태를 보인다.[15] 그러나 척추동물(후구동물)과 절지동물(전구동물) 사이에 몸의 마디(분절)를 만드는 메커니즘이 공통적으로 발견되기 때문에, 최초의 후구동물이 분절된 몸을 가졌다는 증거도 있다.[16]
우르빌라테리아를 포함한 후구동물은 자포동물이나 무체강동물보다 더 복잡한 신체 발달을 조절하는 호메오박스 유전자를 더 많이 가지고 있다. 여기에는 머리의 형성에 관여하는 유전자도 포함된다.[17]
3. 2. 화석 기록
화석 기록에서 처음 발견된 좌우대칭동물(Bilateria)의 증거는 에디아카라기(Ediacaran) 퇴적물에서 발견된 생흔화석(trace fossils)이다. 최초의 확실한(bona fide) 좌우대칭동물 화석은 킴베렐라(Kimberella)로, 약 5억 5500만 년 전(555 Ma)의 것으로 추정된다.[18] 그 이전의 화석들은 논란의 여지가 있는데, 버니애니멀큘라(Vernanimalcula) 화석은 가장 오래된 좌우대칭동물일 가능성이 있지만, 채워진 기포일 가능성도 있다.[19][20] 버니애니멀큘라가 발견된 시기(약 5억 8000만 년 전, 580 Ma) 무렵의 화석 배아(Fossil embryos)가 알려져 있지만, 이들 중 어떤 것도 좌우대칭동물과의 연관성을 보여주지 않는다.[21] 우루과이 타쿠아리층(Tacuarí Formation)에서 좌우대칭동물 생명체가 만들었다고 여겨지는 굴이 발견되었는데, 적어도 5억 8500만 년 전의 것으로 여겨졌으나,[22] 최근 연구 결과에 따르면 이 화석들은 에디아카라기가 아닌 고생대 후기의 것으로 나타났다.[23]
4. 계통 분류
해면동물과 자포동물을 제외한 대부분의 후생동물은 좌우대칭동물에 속한다. 이들은 생명 진화 역사에서 중요한 단계를 나타내며, 캄브리아기 대폭발 무렵 많은 동물이 좌우대칭 형태를 갖추게 되었다.[40]
대부분의 좌우대칭동물은 몸이 세 개의 배엽(내배엽, 중배엽, 외배엽)으로 구성된 삼배엽성 동물이다.[40] 이름처럼 대부분 좌우대칭 또는 거의 대칭적인 몸 구조를 가지지만, 극피동물처럼 성체는 방사대칭이지만 유생 시기에는 좌우대칭인 예외도 존재한다. 일반적으로 입과 항문이 구분된 완전한 소화관을 가지고 있으며, 많은 경우 체내 공간인 체강을 가진다. 과거 무체강동물로 분류되었던 편형동물 등은 현재 체강이 이차적으로 없어진 것으로 여겨진다.
좌우대칭동물은 전통적으로 배 발생 과정의 차이에 따라 후구동물(Deuterostomia)과 선구동물(Protostomia)로 나뉜다. 최근 분자 계통학적 연구 결과 등을 바탕으로 선구동물을 탈피동물(Ecdysozoa)과 나선동물(Spiralia) 등으로 세분하는 등 분류 체계에 대한 논의가 활발히 이루어지고 있다. 상세한 계통 관계는 하위 섹션에서 다룬다.
다음은 좌우대칭동물의 주요 하위 분류군 목록이다.
| 분류군 | 주요 문(Phylum) |
|---|---|
| 직영동물문 (Orthonectida) | - |
| 능형동물문 (Rhombozoa) | - |
| 진무장동물문 (Acoelomorpha) | (분류 논란 중) |
| 모악동물문 (Chaetognatha) | - |
| 후구동물상문 (Deuterostomia) | 척삭동물문 (Chordata) 반삭동물문 (Hemichordata) 극피동물문 (Echinodermata) 유형동물문 (Xenoturbellida) 고충동물문 (Vetulicolia) † |
| 선구동물 (Protostomia) | 탈피동물상문 (Ecdysozoa) * 유극동물 (Scalidophora): 동문동물문 (Kinorhyncha), 동갑동물문 (Loricifera), 새예동물문 (Priapulida) * 선형동물문 (Nematoda) * 유선형동물문 (Nematomorpha) * 범절지동물 (Panarthropoda): 엽족동물문 (Lobopodia)†, 유조동물문 (Onychophora), 완보동물문 (Tardigrada), 절지동물문 (Arthropoda) 나선동물상문 (Spiralia) (일부 분류 체계에서는 편형동물상문과 관륜동물상문으로 나눔) * 편형동물문 (Platyhelminthes) * 복모동물문 (Gastrotricha) * 윤형동물문 (Rotifera) * 구두동물문 (Acanthocephala) * 악구동물문 (Gnathostomulida) * 미악동물문 (Micrognathozoa) * 유륜동물문 (Cycliophora) * 성구동물문 (Sipuncula) * 히올리테스동물문 (Hyolitha) † * 뉴형동물문 (Nemertea) * 추충동물문 (Phoronida) * 외강동물문 (Bryozoa) * 내강동물문 (Entoprocta) * 완족동물문 (Brachiopoda) * 연체동물문 (Mollusca) * 환형동물문 (Annelida) * 유무시동물문 (Echiura) |
†: 멸종된 분류군
4. 1. 계통도
좌우대칭동물은 전통적으로 두 개의 주요 계통 또는 상문으로 나뉘어 왔다.[24]- 후구동물 (Deuterostomia): 전통적으로 극피동물, 반삭동물, 척삭동물, 그리고 멸종된 베툴리콜리아를 포함한다.
- 선구동물 (Protostomia): 절지동물, 환형동물, 연체동물, 편형동물 등 대부분의 나머지 동물을 포함한다.
이 둘의 주요 차이점 중 하나는 배아 발생 과정에 있다. 배아의 첫 번째 구멍인 원구가 선구동물에서는 입이 되고, 후구동물에서는 항문이 된다.
최근 많은 분류학자들은 선구동물 내에 최소 두 개의 상문, 즉 탈피동물 (Ecdysozoa)[25]과 나선동물 (Spiralia)[25][26][27]을 인정한다. 모악동물 (Chaetognatha, 화살벌레류)은 분류가 어려웠으나, 최근 연구에 따르면 유악동물 (Gnathifera)에 속하는 것으로 보인다.[28][29][30]
후구동물과 선구동물로 나누는 전통적인 분류는 새로운 형태학적 및 분자적 증거에 의해 도전을 받았다. 새로운 증거는 무체강 동물군인 진무장동물 (Acoela)과 뉴메르토데르마티다 (Nemertodermatida) (이 둘을 합쳐 무체강동물상문 (Acoelomorpha)이라 부름)가 나머지 좌우대칭동물과 자매 관계임을 시사한다.[24] 이 나머지 좌우대칭동물 분지군은 Jondelius 등(2002)에 의해 신장동물 (Nephrozoa)로, Baguña와 Riutort(2004)에 의해 진좌우대칭동물 (Eubilateria)로 명명되었다.[24] 과거 진무장동물은 특징이 이차적으로 단순화된 편형동물로 여겨졌으나, 새로운 관계는 이들의 단순한 형태가 좌우대칭동물의 원시적인 몸 구조이며, 체강, 소화관, 배설 기관, 신경삭 등은 신장동물에서 진화했음을 시사한다.[24][31] 이후 진무장동물은 이형체강동물 (Xenacoelomorpha) 문에 키세노투르벨리스 (Xenoturbella)와 함께 배치되었고, 계통유전체 분석 결과 이형체강동물과 신장동물 간의 자매 관계가 확인되었다.[31]
Casey Dunn과 동료들의 2014년 검토에 따른 좌우대칭동물의 현대적 합의 계통발생학적 계통도는 다음과 같은 관계를 보여준다.[32]
- 좌우대칭동물 (Bilateria)
- 이형체강동물 (Xenacoelomorpha)
- 키세노투르벨리스 (Xenoturbellida)
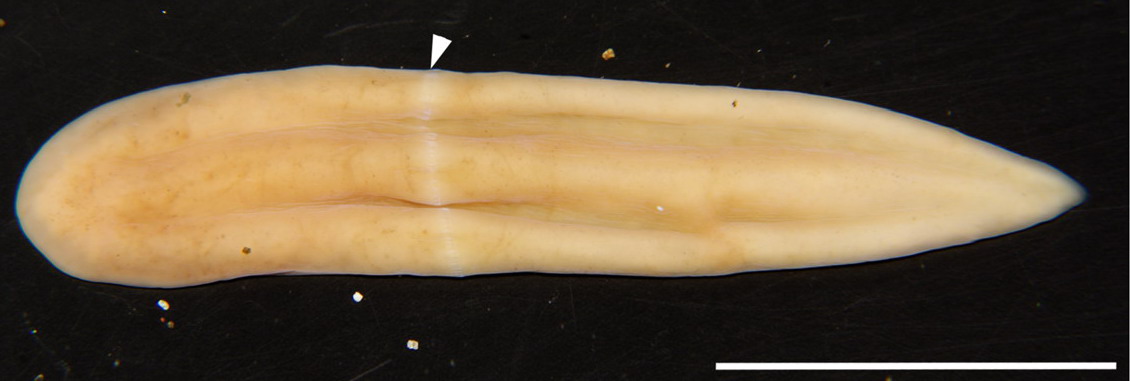
키세노투르벨리스 자포니카 - 무체강동물상문 (Acoelomorpha)
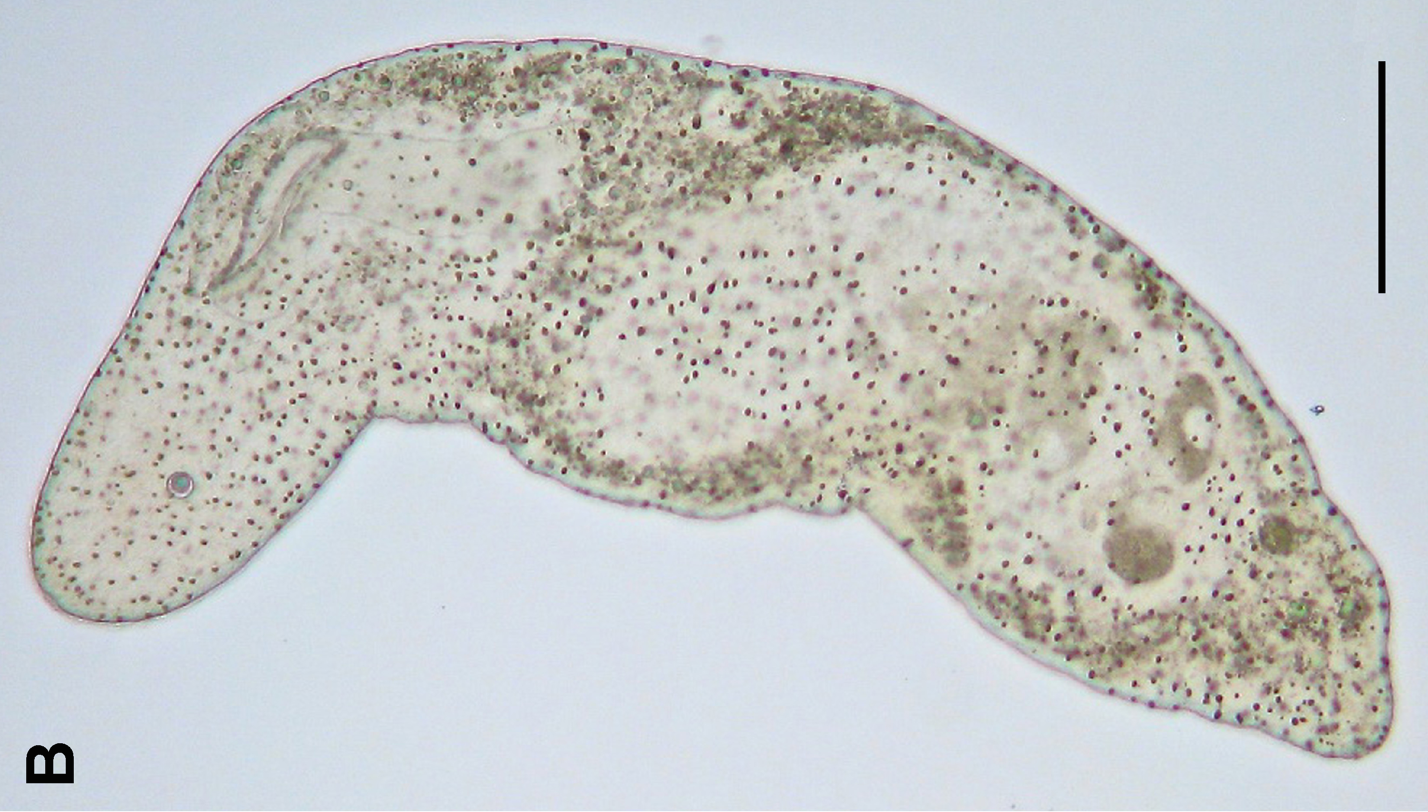
프로포루스 종 - 신장동물 (Nephrozoa) - 약 6억 5천만 년 전 분기 추정
- 후구동물 (Deuterostomia)
- 극피동물 (Echinodermata)

불가사리류 - 척삭동물 (Chordata)

유럽창고기 - 선구동물 (Protostomia) - 약 6억 1천만 년 전 분기 추정
- 탈피동물 (Ecdysozoa)

긴코 바구미 - 나선동물 (Spiralia)

포도원 달팽이
다른 가설도 존재한다. 예를 들어, 극피동물이 이형체강동물과 자매 관계를 이루어 이형극피동물 (Xenambulacraria)을 형성한다는 주장이다. 이 가설에서 이형극피동물은 척삭동물 또는 중추신경동물 (Nephrozoa, 여기서는 척삭동물 + 선구동물에 해당)과 자매 관계일 수 있다.[33] Hervé Philippe와 동료들의 2019년 연구는 이러한 계통 관계를 제시하면서도, "지지 값이 매우 낮아 전통적인 선구동물과 후구동물의 이분법을 반박할 확실한 증거가 부족하다"고 지적했다.[35] 이 연구에서 제시된 계통 관계는 다음과 같다.
- 좌우대칭동물 (Bilateria)
- 이형극피동물 (Xenambulacraria)
- 이형체강동물 (Xenacoelomorpha) --
- 극피동물 (Echinodermata) --
- 중추신경동물 (Nephrozoa) - 약 6억 5천만 년 전 분기 추정
- 척삭동물 (Chordata) --
- 선구동물 (Protostomia) - 약 6억 1천만 년 전 분기 추정
- 탈피동물 (Ecdysozoa) --
- 나선동물 (Spiralia) --
이러한 계통도들은 일부 분지군이 수백만 년 전(Mya)에 새로운 분지군으로 분기된 시점을 대략적으로 나타낸다.[34]
5. 분류
- 직영동물문 (Orthonectida)
- 능형동물문 (Rhombozoa)
- 무장동물문 (Acoelomorpha) (분류 논란 중)
- 모악동물문 (Chaetognatha)
- '''후구동물상문 (Deuterostomia)'''
- * 척삭동물문 (Chordata)
- * 반삭동물문 (Hemichordata)
- * 극피동물문 (Echinodermata)
- * 유형동물문 (Xenoturbellida)
- * 고충동물문 (Vetulicolia) †
- '''전구동물 (Protostomia) (계급 미정)'''
- * '''탈피동물상문 (Ecdysozoa)'''
- ** 유극동물 (Scalidophora) (계급 미정)
동문동물문 (Kinorhyncha)
동갑동물문 (Loricifera)
새예동물문 (Priapulida)
엽족동물문 (Lobopodia)†
유조동물문 (Onychophora)
완보동물문 (Tardigrada)
절지동물문 (Arthropoda)
- * '''편형동물상문 (Platyzoa)'''
- ** 편형동물문 (Platyhelminthes)
- ** 복모동물문 (Gastrotricha)
- ** 윤형동물문 (Rotifera)
- ** 구두동물문 (Acanthocephala)
- ** 악구동물문 (Gnathostomulida)
- ** 미악동물문 (Micrognathozoa)
- ** 유륜동물문 (Cycliophora)
- * '''관륜동물상문 (Lophotrochozoa)'''
- ** 성구동물문 (Sipuncula)
- ** 히올리테스동물문 (Hyolitha) †
- ** 뉴형동물문 (Nemertea)
- ** 추충동물문 (Phoronida)
- ** 외강동물문 (Bryozoa)
- ** 내강동물문 (Entoprocta)
- ** 완족동물문 (Brachiopoda)
- ** 연체동물문 (Mollusca)
- ** 환형동물문 (Annelida)
- ** 유무시동물문 (Echiura)
6. Taxonomic history
좌우대칭동물은 1888년 오스트리아의 발생학자 베르톨트 하체크 Berthold Hatschekde에 의해 처음 명명되었다. 그의 분류 체계에서 이 그룹은 좌우대칭동물, 촉수동물, 그리고 척삭동물을 포함했다.[36][37] 이후 1910년, 오스트리아 동물학자 카를 그로벤 Karl Grobbende은 좌우대칭동물을 원구동물아계 Protostomiala로 개명하고, 촉수동물과 척삭동물을 포함하는 후구동물아계 Deuterostomiala를 새롭게 만들었다.[36][38]
참조
[1]
논문
Patterns of distribution in the Ediacaran biotas: facies versus biogeography and evolution
http://paleobiol.geo[...]
[2]
서적
The Invertebrate Tree of Life
Princeton University Press
2020-03-03
[3]
논문
Careful amendment of morphological data sets improves phylogenetic frameworks: re-evaluating placement of the fossil Amiskwia sagittiformis
2022-12-31
[4]
논문
Death march of a segmented and trilobate bilaterian elucidates early animal evolution
[5]
웹사이트
2024-05-12
[6]
웹사이트
2024-05-12
[7]
서적
Invertebrates
Sinauer Associates
2016
[8]
논문
Did internal transport, rather than directed locomotion, favor the evolution of bilateral symmetry in animals?
http://faculty.weber[...]
2005-11
[9]
논문
Ontogenetic scaling of hydrostatic skeletons: geometric, static stress and dynamic stress scaling of the earthworm lumbricus terrestris
http://jeb.biologist[...]
1998-05
[10]
서적
Perspectives in Animal Phylogeny and Evolution
https://books.google[...]
Oxford University Press
[11]
논문
Discovery of the oldest bilaterian from the Ediacaran of South Australia
2020-04-07
[12]
논문
Early Animal Evolution: Emerging Views from Comparative Biology and Geology
1999-06-25
[13]
논문
The segmented Urbilateria: A testable scenario
2003
[14]
논문
Xenacoelomorpha is the sister group to Nephrozoa
http://urn.kb.se/res[...]
[15]
논문
Back in time: a new systematic proposal for the Bilateria
2008-04
[16]
서적
How the Snake Lost its Legs. Curious Tales from the Frontier of Evo-Devo
Cambridge University Press
2014
[17]
논문
Anterior Hox Genes and the Process of Cephalization
2021-08-05
[18]
논문
The Late Precambrian fossil Kimberella is a mollusc-like bilaterian organism
1997-11
[19]
논문
Comment on 'small bilaterian fossils from 40 to 55 million years before the Cambrian'.
2004-11-19
[20]
논문
A merciful death for the 'earliest bilaterian,' Vernanimalcula
http://urn.kb.se/res[...]
[21]
논문
Cellular and Subcellular Structure of Neoproterozoic Animal Embryos
2006-10-13
[22]
논문
Bilaterian burrows and grazing behavior at >585 million years ago
2012-06-29
[23]
논문
Revisiting the supposed oldest bilaterian trace fossils from Uruguay: Late Paleozoic, not Ediacaran
https://www.scienced[...]
2022-09-15
[24]
논문
Six major steps in animal evolution: are we derived sponge larvae?
[25]
논문
Evidence from 18S ribosomal DNA that the lophophorates are protostome animals
1995-03-17
[26]
논문
Bilaterian phylogeny: a broad sampling of 13 nuclear genes provides a new Lophotrochozoa phylogeny and supports a paraphyletic basal Acoelomorpha
2009-07-14
[27]
논문
Resolving animal phylogeny: A sledgehammer for a tough nut?
2008-04-15
[28]
논문
The mitochondrial genome of ''Paraspadella gotoi'' is highly reduced and reveals that chaetognaths are a sister group to protostomes
2004-07-20
[29]
논문
Identification of chaetognaths as protostomes is supported by the analysis of their mitochondrial genome
2004-11
[30]
논문
Rotiferan Hox genes give new insights into the evolution of metazoan bodyplans
2017-04-04
[31]
논문
Xenacoelomorpha is the sister group to Nephrozoa
http://urn.kb.se/res[...]
[32]
논문
Animal Phylogeny and Its Evolutionary Implications
2014-11-23
[33]
논문
Lack of support for Deuterostomia prompts reinterpretation of the first Bilateria
2021-03-19
[34]
논문
The Ediacaran emergence of bilaterians: congruence between the genetic and the geological fossil records
2008-04-27
[35]
논문
Mitigating Anticipated Effects of Systematic Errors Supports Sister-Group Relationship between Xenacoelomorpha and Ambulacraria
2019
[36]
저널
The 'new phylogeny'. What is new about it?
https://www.palaeodi[...]
[37]
서적
Lehrbuch der Zoologie
Gustav Fischer
[38]
서적
Lehrbuch der Zoologie
Elvert'sche Verlagsbuchhandlung
[39]
저널
Patterns of distribution in the Ediacaran biotas: facies versus biogeography and evolution
http://paleobiol.geo[...]
[40]
웹사이트
動物はいつ左右対称になったのか - 日経サイエンス
https://www.nikkei-s[...]
2005-10-01
[41]
서적
Invertebrates
Sinauer Associates
2016
[42]
저널
The Ediacaran emergence of bilaterians: congruence between the genetic and the geological fossil records
2008-04-27
[43]
저널
Estimating the timing of early eukaryotic diversification with multigene molecular clocks
2011-08-16
[44]
웹인용
Raising the Standard in Fossil Calibration
http://fossilcalibra[...]
[45]
저널
Support for a clade of Placozoa and Cnidaria in genes with minimal compositional bias
[46]
저널
Revisions to the Classification, Nomenclature, and Diversity of Eukaryotes
2018
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com