키노돈트
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
키노돈트는 수궁류의 한 아목으로, 포유류의 조상으로 여겨지는 멸종된 동물 그룹이다. 2억 6500만 년 전 페름기에 처음 등장하여 트라이아스기를 거쳐 쥐라기까지 번성했다. 초기 키노돈트는 파충류와 유사한 특징을 가졌지만, 턱뼈와 이빨 구조, 2차 구개 발달 등 포유류의 특징을 갖추고 있었다. 페름기-트라이아스기 대멸종 이후 급격히 번성하여 다양한 종으로 분화되었으며, 턱뼈 감소 및 이차 구개 발달 등 포유류로 진화하는 과정에서 중요한 변화를 겪었다. 비포유류 키노돈트는 트라이아스기 말에 대부분 멸종되었으며, 일부는 쥐라기 초까지 생존했다.

더 읽어볼만한 페이지
- 1861년 기재된 화석 분류군 - 콤프소그나투스
콤프소그나투스는 후기 쥐라기에 살았던 작은 육식 공룡으로, 독일과 프랑스에서 발견된 완전한 골격 화석을 통해 알려져 있으며, 깃털 유무, 식성, 조류와의 관계로 논쟁이 있었고 쥬라기 공원에 등장하며 인기를 얻었다. - 1861년 기재된 화석 분류군 - 시조새
시조새는 쥐라기 후기에 살았던 멸종된 속으로, 파충류와 조류의 특징을 가져 조류의 기원과 진화 연구에 중요한 화석이며, 깃털, 날개, 뇌 구조 등을 통해 비행 능력과 생활 방식에 대한 연구가 진행되고 있고 조류의 공룡 기원설을 뒷받침하는 핵심 증거로서 진화론적 논쟁에서 중요한 위치를 차지한다. - 키노돈트 - 심보돈
심보돈은 잠비아의 마두마비사 이암층에서 발견된 페름기 후기의 화석이며, 두개골, 어깨 뼈, 앞다리 뼈 등을 포함하는 여러 개의 뼈로 구성되어 있다. - 키노돈트 - 상키노돈트류
- 리처드 오언이 명명한 분류군 - 극어류
극어류는 멸종된 어류의 한 무리로, 가시 상어라고도 불리며 실루리아기부터 페름기까지 번성했고, 비늘 장식으로 퇴적암 연대 추정에 사용되었으며, 연골어류에 속한다. - 리처드 오언이 명명한 분류군 - 악어
악어는 넓고 납작한 몸, 긴 주둥이, 짧은 목, 납작한 꼬리를 가진 파충류로 열대 및 아열대 지역에 서식하며 강력한 턱과 이빨로 다양한 먹이를 포식하고 인간과 연관이 깊지만 멸종 위기에 처해 보존 노력이 필요하다.

2. 분류
키노돈트의 분류는 학자마다 이견이 있어왔다. 리처드 오웬은 1861년에 키노돈아목(Cynodontia)이라는 학명을 명명하고 키노돈트를 아노모돈아목에 분류했다.[24] 그러나 에버렛 올슨은 키노돈트를 테리오돈트류로, 부루스 루비지와 크리스티안 시더는 키노돈트를 진테리오돈트류로 분류했다.[25][26]
윌리엄 킹 그레고리(1910), 로버트 브룸(1913), 캐럴(1988), 고티에 등(1989), 제임스 홉슨과 키칭(2001), 보타 등(2007)은 모두 키노돈아목을 수궁목 바로 아래에 분류했다. 브링크(2007)는 오웬(1861)의 분류법을 따랐지만 분류 계급을 지정하지 않은 것으로 보인다.[15][16][17]
키노돈트의 어원은 "개 (cyn-)의 이 (odont-)"이다.
2. 1. 주요 분류군
리처드 오웬은 1861년에 '''키노돈아목'''(Cynodontia)이라는 학명을 명명하고 키노돈트를 '''아노모돈아목'''에 분류했다.[24] 그러나 '''에버렛 올슨'''은 키노돈트를 '''테리오돈트류'''로, '''부루스 루비지'''와 '''크리스티안 시더'''는 키노돈트를 '''진테리오돈트류'''로 분류했다.[25][26]키노돈트의 분류에 대한 연구 결과[27]를 참고해서 만든 분지도는 다음과 같다.

2. 2. 분지도
리처드 오웬은 1861년에 '''키노돈아목'''(Cynodontia)이라는 학명을 명명하고 키노돈트를 '''아노모돈아목'''에 분류했었다.[24] 그러나 에버렛 올슨은 키노돈트를 '''테리오돈트류'''(Theriodontia)로, 부루스 루비지와 크리스티안 시더는 키노돈트를 '''진테리오돈트류'''(Eutheriodontia)로 분류했었다. 윌리엄 킹 그레고리(1910), 로버트 브룸(1913), 캐럴(1988), 고티에(1989), 제임스 홉슨과 키칭(2001), 보타(2007) 모두 키노돈아목을 수궁목 바로 아래에 분류했다. 브링크(2007)는 오웬(1861)의 분류법을 따랐지만 분류 계급을 지정하지 않은 것으로 보인다.[25][26]다음은 Ruta, Botha-Brink, Mitchell 및 Benton(2013)의 연구 결과를 참고하여 만든 키노돈트의 분기도이다.[18]
{| class="wikitable"
|-
! 키노돈트
|-
|
{| class="wikitable"
|-
| †카라소그나투스
|-
|
{| class="wikitable"
|-
|
|-
| 에피키노돈트
|-
|
{| class="wikitable"
|-
| †키노사우루스
|-
|
{| class="wikitable"
|-
|
|-
|
{| class="wikitable"
|-
|
|-
|
{| class="wikitable"
|-
| †플라티크라니엘루스
|-
| 유키노돈트
|-
|
|}
|}
|}
|}
|}
|}
|}
{| class="wikitable"
|-
! 키노그나티아
|-
|
{| class="wikitable"
|-
| †키노그나투스
|-
| †곰포돈티아
|-
|
{| class="wikitable"
|-
| †디아데모돈
|-
|
{| class="wikitable"
|-
| †트리라코돈티다에
|-
|
{| class="wikitable"
|-
|
|-
|
{| class="wikitable"
|-
| †트리라코돈
|-
|
|}
|}
|-
| †트라베르소돈티다에
|-
|
{| class="wikitable"
|-
|
|-
|
{| class="wikitable"
|-
| †스칼레노돈
|-
|
{| class="wikitable"
|-
| †루앙과
|-
|
{| class="wikitable"
|-
| †트라베르소돈
|-
|
{| class="wikitable"
|-
|
{| class="wikitable"
|-
| †"스칼레노돈" 아트리지아이
|-
|
{| class="wikitable"
|-
| †만다고포돈
|-
|
{| class="wikitable"
|-
| †나노곰포돈
|-
|
|}
|}
|}
|-
|
{| class="wikitable"
|-
| †마세토그나티나에
|-
|
{| class="wikitable"
|-
| †마세토그나투스
|-
|
|}
|-
| †곰포돈토수키나에
|-
|
{| class="wikitable"
|-
| †메나돈
|-
|
{| class="wikitable"
|-
| †곰포돈토수쿠스
|-
|
{| class="wikitable"
|-
| †프로투베룸
|-
|
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
{| class="wikitable"
|-
! 프로베이노그나티아
|-
|
{| class="wikitable"
|-
| †룸쿠이아
|-
|
{| class="wikitable"
|-
| †에크테니니온
|-
|
{| class="wikitable"
|-
|
|-
|
{| class="wikitable"
|-
| †프로베이노그나투스
|-
|
{| class="wikitable"
|-
| †트루치도키노돈
|-
|
{| class="wikitable"
|-
| †테리오헤르페톤
|-
|
{| class="wikitable"
|-
| †트리티레돈티다에
|-
|
{| class="wikitable"
|-
| †리오그란디아
|-
|
{| class="wikitable"
|-
|
|-
|
|}
|}
|-
|
{| class="wikitable"
|-
| †브라실로돈티다에
|-
|
|-
|
{| class="wikitable"
|-
| †트리티로돈티다에
|-
|
{| class="wikitable"
|-
| †올리고키푸스
|-
|
{| class="wikitable"
|-
| †카옌타테리움
|-
|
|}
|}
|-
| '''포유류형류'''
|-
|
키노돈트는 현재 포유류가 가진 특징 대부분을 이미 가지고 있었다. 이빨은 여러 형태로 분화되었고(이형 치아), 두개골은 뒤쪽이 튀어나와 있었다. 초기 단계에서 이미 털을 획득했을 것으로 보이며, 테로케팔리아를 제외한 다른 수궁류에게는 없었던 골성 이차 구개도 가지고 있었다. 이는 음식을 씹으면서 동시에 숨을 쉴 수 있게 해 주었으며, 많은 양의 산소를 필요로 하는 항온 동물로 진화하고 있었음을 보여준다. 몸을 둥글게 말고 잠든 모습으로 화석화된 키노돈류의 화석은 체온 유지를 위한 행동이었을 것으로 추정된다.
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
|}
3. 특징
키노돈트는 항온 동물로 추정되어 털을 가졌을 것으로 추측되기도 하지만, 털 화석 증거는 찾기 어렵다. 현대 포유류는 털을 덮기 위해 지방질을 분비하는 하르더샘을 가지고 있지만, 이 샘은 원시 포유류인 모르가누코돈과 그 이후의 후손에게서만 발견된다.[28] 그러나 페름기 단궁류의 대변에 대한 최근 연구는 초기 수궁류가 털을 가졌을 수 있다는 사실을 보여주며,[29] 카스토로카우다와 메가코누스 같은 초기 포유류에게 이미 털이 존재했음을 보여준다.
초기 키노돈트는 파충류와 유사하게 코뼈에 많은 작은 구멍을 가지고 있었다. 이는 근육이 없는 입술과 뺨을 가졌음을 시사한다. 근육질의 움직이는 얼굴은 수염 움직임과 손상 방지에 필요하므로, 초기 키노돈트는 수염이 없었을 가능성이 낮다. 프로조스트로돈트 키노돈트(포유류 포함 그룹)에서는 구멍이 단일의 큰 안와하공으로 대체되어, 얼굴 근육과 수염이 존재했음을 나타낸다.[3]
초기 수치류인 고르고놉스류 화석의 콧등에 이미 수염의 흔적이 있는 것으로 보아, 이미 체모도 획득했던 것으로 보인다.
3. 1. 골격
초기 키노돈트는 포유류의 골격적 특징을 많이 가지고 있었다. 치아는 완전히 분화되어 있었고 뇌관은 불룩하게 변했으며, 일부 왕관군 포유류를 제외한 모든 키노돈트는 알을 낳았을 것이다. 특히 측두창은 다른 수궁류보다 컸고, 포유류 두개골의 광대뼈 확대는 비교적 강한 턱 근육을 가지게 했을 것이다. 테로세팔루스류를 제외한 다른 원시 수궁류에게는 없었던 2차 구개를 가지고 있었다.
비포유류형 키노돈트가 진화하는 동안 포유류의 턱 뼈 수는 줄어들고 있었다. 하악골이 안쪽을 향해 이동했고 턱에 있는 다른 뼈들의 관절과 각이 두개골로 이동할 수 있는 길을 열었다. 이렇게 등장한 새로운 형태의 두개골은 포유류의 청각 시스템의 일부라고 할 수 있다.
또 입천장에 2차 구개가 생겼고 이는 콧구멍에서 나오는 공기 흐름이 직접적으로 통하는 대신 입 뒤쪽의 위치로 이동하게 되어 음식을 씹으면서 숨을 쉴 수 있게 되었다. 이는 지구상의 모든 포유류들이 가지고 있는 형질이다.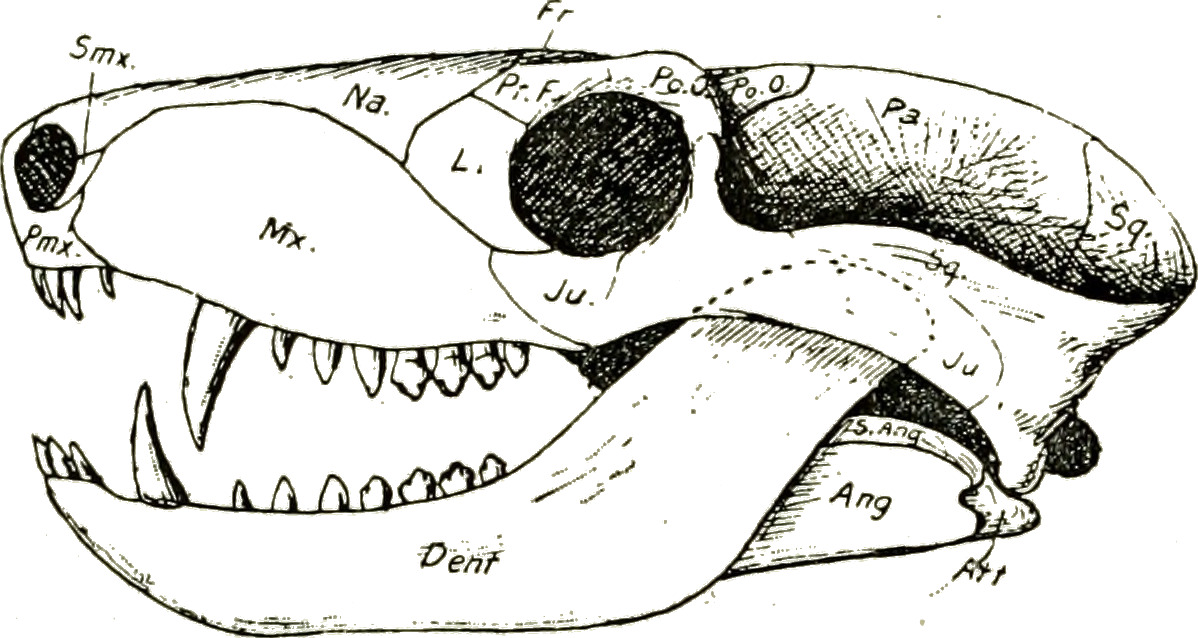
파생된 키노돈트는 서골을 발달시켰다. 이것은 몸통을 강화하고 복부 및 뒷다리 근육을 지지하여 직립 보행의 발달을 도왔지만, 임신 기간이 길어져 현대 유대류 및 단공류처럼 극도로 미숙아를 낳도록 했다. 태반류, 그리고 아마도 ''메가조스트로돈''과 ''에리트로테륨''만이 이것들을 잃었을 것이다.[4][5] ''카옌타테륨''의 표본은 실제로 적어도 트리틸로돈티드가 이미 근본적으로 유대류와 유사한 생식 스타일을 가지고 있었지만, 약 38마리의 주산기 또는 알을 낳는 것과 같은 훨씬 더 많은 새끼를 낳았음을 보여준다.[6]
키노돈트는 하라미이다에서 활공이 알려져 있고,[7] 다양한 포유류 그룹에서 활공이 알려져 있으며, 태반류 포유류가 비행을 발달시킨 유일한 알려진 시냅스류 계통이다.[8]
가장 큰 알려진 비포유류 키노돈트는 traversodontid인 ''스칼레노돈토이데스''이며, 파편 표본을 기반으로 최대 두개골 길이가 약 617mm으로 추정된다.
특별히 언급하지 않는 한, 포유류 이전의 종에 대해 서술한다.
; 두개골
: 앞뒤로 긴 큰 측두창을 가지며, 중간에 깊은 능선이 형성된다(포유류에서는 이차적으로 뇌두개골이 확대). 두정공은 갈레사우루스과의 단계에서 소실. 주둥이는 가늘고, 광대뼈 아치가 튀어나와 있다.
; 후두과
: 아주 초기 종에서는 한 개이지만, 갈라져 있다. 중기 이후의 종은 한 쌍.
; 하악
: 치골이 확대되고, 근돌기가 발달한다. 포유류형류 이후의 종은, 치골만 남는다. 중기 이후의 종에서는, 이생 치성이 된다. 이것은 저작 기능이 발달하여, 위아래의 이가 절구와 절구공이처럼 작동해야 할 필요가 있었기 때문으로 생각된다.
; 턱 관절
: 초기의 종은 방형골과 관절골로 형성되어 있지만, 치니쿼돈과에서는 치골과 인상골이 접촉하여 이중 관절이 된다. 포유류형류 이후는 방형골과 관절골은 턱 관절에서 벗어나, 쥐라기 포유류에서 각각 침골과 추골로 변화하고 있다.
; 골성 이차 구개
: 페름기 종에서는 중앙에 홈이 있는 등 불완전하다. 트라이아스기 이후는 좌우가 완전히 유합.
; 이
: 이치성이 더욱 현저해지고, 협치에 교두가 발달한다.
; 어깨뼈대
: 견갑골, 전까마귀뼈, 까마귀뼈, 상쇄골, 쇄골, 간쇄골의 6종류의 뼈로 구성된다. 이 중, 견갑골과 까마귀뼈가 내골격성 어깨뼈대의 주요 구성 요소가 되어 간다(주룡류의 계통에서는 견갑골과 전까마귀뼈). 이후, 포유류 단계에서 상쇄골, 수아강에 이르는 단계에서 전까마귀뼈, 간쇄골이 소실. 까마귀뼈가 견갑골에 유합되어 있다.
; 척주
: 트리낙소돈 단계에서, 경추 중 환추 극돌기가 축소. 축추와의 간섭이 없어 머리 회전이 가능해졌다. 또한, 흉부 이외의 늑골이 축소. 경추, 흉추, 요추 등으로의 분화 도중에 있다. 허리 부분의 늑골이 축소되면서, 몸통을 비틀 수 있게 되었다.
; 사지
: 초기 종에서는, 파행 또는 중허리라는 형태를 취한다. 골반대에 관해서는, 갈레사우루스류 및 유키노돈류에서는, 불완전하지만 직립 보행을 획득했다. 다만, 어깨뼈대는 아직 상완골이 수직과는 거리가 먼 위치에 있다. 이것이 수직이 되는 것은 포유류 단계에 들어서면서부터이다.
3. 2. 털과 항온성
키노돈트는 항온 동물로 추정되며, 이로 인해 털을 가졌을 것으로 추측되기도 한다. 하지만 털 화석 증거는 찾기 어렵다. 현대 포유류는 털을 덮기 위해 지방질을 분비하는 하르더샘을 가지고 있지만, 하르더샘은 원시 포유류인 모르가누코돈과 그 이후의 후손에게서만 발견된다.[28] 페름기 단궁류의 대변에 대한 최근 연구는 초기 수궁류가 털을 가졌을 수 있다는 사실을 보여주며,[29] 카스토로카우다(Castorocauda)와 메가코누스(Megaconus) 같은 초기 포유류에게 이미 털이 존재했음을 보여준다.
초기 키노돈트는 파충류와 유사하게 코뼈에 많은 작은 구멍을 가지고 있었다. 이는 근육이 없는 입술과 뺨을 가졌음을 시사한다. 근육질의 움직이는 얼굴은 수염 움직임과 손상 방지에 필요하므로, 초기 키노돈트는 수염이 없었을 가능성이 낮다. 프로조스트로돈트 키노돈트(포유류 포함 그룹)에서는 구멍이 단일의 큰 안와하공으로 대체되어, 얼굴 근육과 수염이 존재했음을 나타낸다.[3]
키노돈류는 초기 단계에서 이미 털을 획득했을 것으로 보인다. 또한 테로케팔리아를 제외한 수궁류에는 존재하지 않았던 골성 이차 구개도 초기 단계에서 이미 획득했다. 이는 저작과 동시에 호흡이 가능했음을 나타내며, 많은 양의 산소를 필요로 했던 것으로 보아 항온성을 획득해 가고 있었을 것이다. 이는 몸을 둥글게 말고 잠든 모습으로 화석화된 키노돈류의 화석에서도 엿볼 수 있다. 그 자세는 현재의 소형 포유류와 마찬가지로 체온을 유지하기 위한 것이었을 것이다. 초기 수치류인 고르고놉스류 화석의 콧등에 이미 수염의 흔적이 인정되는 것으로 보아, 이미 체모도 획득했던 것으로 보인다.
3. 3. 굴 생활
적지 않은 키노돈트 종류가 지하에 굴을 파서 생활하며, 그중에는 여러 굴이 묶여서 발견된 사례도 있다.[23]
4. 진화
키노돈류는 멸종된 고르곤놉스아목 및 테로케팔루스아목과 함께 수치류라는 분류군에 속한다. 그 기원은 고생대 페름기 후기로, 수궁류 테로케팔루스아목에 가까운 그룹에서 파생되었다고 여겨진다. 가장 오래된 키노돈류는 2억 4,800만 년 - 2억 4,500만 년 전에 생존했던 ''Charassognathus''와 도비니아이다. 그 다음으로 나타난 프로키노수쿠스는 수달처럼 수중 생활에 적응했을 것으로 여겨진다. 이들은 페름기에 생존했지만, 다른 종들에 밀려 주요 생태적 지위를 차지하지 못하고 물가 등에서 생활했던 것으로 보인다.
페름기 말부터 트라이아스기 초기에 걸쳐 (P-T 경계), 판게아 대륙 완성의 영향으로 발생한 초대형 핫 플룸 "슈퍼 플룸"이 돔 모양으로 솟아올라 지상에 도달하면서 시베리아에 대분화가 일어나 대규모 환경 파괴가 발생했다. 지상은 고온에 노출되었고, 이전까지 30% 가까이 유지되던 산소 농도가 크게 감소했다. 이 대량 멸종으로 지구상 생명체의 90%가 사라졌다. 고온을 피하고 저산소 환경에 견딜 수 있는 능력을 가진 생물(굴성 생활을 한 디키노돈류, 기낭 시스템을 가진 새의 조상인 공룡의 조상 등)만이 살아남았다.
멸종에서 살아남은 키노돈류는 비어버린 생태계를 채우기 위해 빠르게 적응 방산했다. 트리낙소돈 등의 갈레사우루스류는 복부의 늑골을 퇴화시키고, 복식 호흡을 할 수 있는 횡격막을 획득하여 저산소 환경을 극복할 수 있었을 것이다. 이 그룹은 진화하면서 두정공이 소실되고, 후두과가 1쌍이 되며, 늑골이 흉부에만 존재하게 되어 흉강과 복강이 나뉘는 특징을 보인다. 또한, 불완전하나마 직립 보행을 획득했으며, 턱을 구성하는 뼈 중 치골의 확대가 크게 진행되었다.
특히 진화한 종에서는 이 뼈가 관절골과 방형골로 구성된 턱 관절에 접촉하여, 인상골(측두골의 일부)과의 새로운 관절이 형성되었다. 이는 이후 포유류 등을 포함하는 유키노돈류이다. 그중에서 키노그나투스와 같은 강력한 포식자, 디아데모돈, 트라베르소돈과 같은 초식에 적합한 종, 소형 포유류에 가까운 트리티로돈류, 익티도사우루스류 등이 나타났다. 트라이아스기 동안 여러 계통에서 여러 번 대형화되었으며, 육식 종으로는 전기에는 키노그나투스나 갈레사우루스, 중기에는 치니쿠오돈, 후기에는 톨시키노돈이 발견되었다. 초식 종으로는 전기에서 중기에 디아데모돈, 중기 - 후기에 엑사엘레토돈이 발견되었다.
이러한 경향이 더욱 진행되어 관절골과 방형골이 턱에서 분리되어 이소골이 된 것이 포유류이다. 과거에는 이 턱 관절의 특징으로 포유류를 정의했지만, 최근에는 원시적인 그룹이 제외되어 포유류형류로 분류되었다. 가장 오래된 포유류형류는 아데로바실레우스로 2억 2,500만 년 전에 생존했다. 그러나 트라이아스기 후기, 다시 중간 규모의 대량 멸종이 지구를 덮치면서 키노돈류는 트리티로돈류와 포유류 등 극소수 계통을 제외하고 멸종했다.
쥐라기 이후, 키노돈류는 대형 동물의 생태적 지위를 공룡 등 대형 파충류에게 빼앗겨 소형으로 남았다. 다만, 중생대에도 포유류는 수적으로 공룡을 훨씬 능가했다고 여겨진다. 올리고키푸스 등의 트리티로돈과는 점차 수를 줄여갔으며, 백악기 전기, 이시카와현에서 발견된 치아 화석이 그 마지막 기록이다. 캐나다의 6,000만 년 전 지층에서 발견된 키노돈류로 추정되는 턱의 단편은 그 정체가 불명확한 상태로 남아있다.
4. 1. 비포유류형 키노돈트
키노돈트는 페름기 후기에 처음 나타났으며, 초기에는 다른 수궁류의 특징을 많이 가졌지만, 포유류의 특징인 2차 구개 등도 가지고 있었다. 이는 키노돈트와 가까운 테로세팔루스류(Therocephalia)에게도 나타나는 특징이다.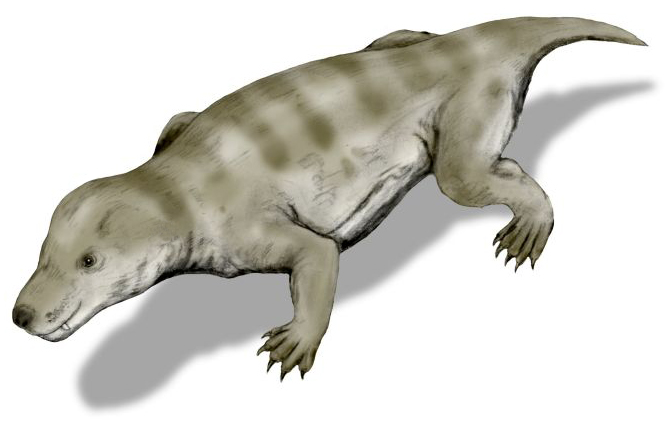
가장 초기의 키노돈트는 차라소그나투스과에 속하며, 주로 남아프리카의 카루 누층군 퇴적물에서 발견된다. 페름기 후기 키노돈트 화석은 남아프리카 이외 지역에서는 드물게 발견되는데, 가장 널리 분포하는 프로키노수쿠스는 남아공, 독일, 탄자니아, 잠비아, 러시아 등에서 발견된다.[30]
키노돈트는 페름기 대멸종 이후 빠르게 다양해졌다. 특히 트라이아스기 초기에는 진키노돈트류(Eucynodontia)가 지배적이었는데, 여기에는 주로 초식성이었던 키노그나투스류(Cynognathia)와 주로 육식성이었던 프로바이노그나투스류(Probainognathia)가 주요 하위 분류군으로 트라이아스기 육상 생태계를 지배했다. 트라이아스기 초-중기에는 키노그나투스류가 다양했지만, 프로바이노그나투스류는 트라이아스기 후기까지 크게 번성하지 못했다.[31] 거의 모든 트라이아스기의 키노돈트는 곤드와나 대륙에 널리 퍼져 있었고, 당시 북반구(로라시아 대륙)에서는 나노곰포돈만이 발견된다. 트라이아스기 중-후기에 번성했던 키노돈트는 당시 곤드와나에 주로 살았던 초식성 키노돈트 트레버소돈과(Traversodontidae)에 속했다.
트라이아스기 말 이후에는 두 형태의 비포유류 키노돈트만이 존재했으며, 둘 다 프로바이노그나투스류에 속했다. 첫 번째는 식충성이었던 트리틸로돈으로 쥐라기 초기에 멸종했다. 두 번째는 중국의 포시오마누스로, 약 1억 2천만년전, 즉 백악기 전기까지 번성했다. 이는 쥐라기 후기에 처음 등장한 초식성 키노돈트이다.
키노돈트의 가장 가까운 친척은 테로케팔리아이며, 이들과 함께 유테리오돈티아라는 분기군을 형성한다.[9]
가장 초기의 키노돈트는 남아프리카 공화국의 카루 초집단의 ''Tropidostoma'' Assemblage Zone의 초기 로핑기아기(초기 우치아핑기안) 퇴적층에서 발견되었으며, 기저 과인 카라소그나투스과에 속한다. 페름기 키노돈트의 화석은 남아프리카 공화국 이외 지역에서는 비교적 드물며, 가장 널리 분포하는 속은 ''프로키노수쿠스''로, 남아프리카 공화국, 독일, 탄자니아, 잠비아, 그리고 아마도 러시아에서도 발견된다.[10]
키노돈트는 페름기-트라이아스기 대멸종 이후 급격하게 다양성을 확장했다. 키노돈트의 최대 분화는 인두안에서 카르니아기, 그리고 중 노리아기에 발생했다.[11] 초기 트라이아스기 이후의 키노돈트는 주로 진화된 분기군인 유키노돈티아의 구성원들이 지배했는데, 여기에는 주로 초식성인 키노그나티아와 주로 육식성인 프로바이나그나티아의 두 가지 주요 하위 분류가 있다. 초기 및 중기 트라이아스기 동안, 키노돈트의 다양성은 키노그나티아의 구성원들이 지배했고, 프로바이나그나티아의 구성원들은 후기 트라이아스기(초기 노리아)까지 두각을 나타내지 못했다.[18] 거의 모든 중기 트라이아스기 키노돈트는 곤드와나에서 발견되었으며, 북반구에서 발견된 속은 단 하나(''나노곰포돈'')뿐이다. 중기 및 후기 트라이아스기 키노돈트의 가장 지배적인 그룹 중 하나는 곤드와나에 주로 서식했던 초식성 트라베르소돈트과이며, 후기 트라이아스기에 최대 다양성에 도달했다. 포유류형류는 후기 트라이아스기 동안 프로바이나그나티안 키노돈트에서 기원했다.[13] 초기 포유류형류는 작은 몸집의 곤충을 먹는 동물이었다.[12] 트라이아스기 말기를 넘어 존재했던 비포유류형류 키노돈트는 단 두 그룹뿐이었으며, 둘 다 프로바이나그나티아에 속했다. 첫 번째는 곤충을 먹는 트리텔레돈트과로, 초기 쥐라기까지 잠시 생존했다. 두 번째는 초식성 트리티로돈트과로, 최후기 트라이아스기에 처음 나타났으며, 쥐라기 동안 북반구에서 번성하고 다양했으며, 중국의 ''Fossiomanus''에 의해 대표되는 것처럼, 적어도 약 1억 2천만 년 전까지 아시아의 초기 백악기(바레미안–압티안)까지 생존했다.[13]
진화 과정에서 키노돈트의 턱뼈 수는 감소했다. 하악골에 단일 뼈로 가는 이러한 움직임은 턱의 다른 뼈인 관절골과 각골이 두개골로 이동하여 포유류 청각 시스템의 일부로 기능할 수 있는 길을 열었다.
키노돈트는 또한 입천장에 이차 구개를 발달시켰다. 이로 인해 콧구멍에서 공기가 입을 직접 통과하는 대신 입 뒤쪽으로 이동하게 되어 키노돈트가 동시에 씹고 숨을 쉴 수 있게 되었다. 이 특징은 모든 포유류에게서 나타난다.

4. 2. 포유류
포유류는 트라이아스기 후기에 프로바이노그나투스류에서 진화한[32] 곤충을 먹는 작은 동물이었다.[33] 비포유류형 키노돈트가 진화하는 동안 포유류의 턱 뼈 수는 줄어들고 있었다. 하악골이 안쪽을 향해 이동했고 턱에 있는 다른 뼈들의 관절과 각이 두개골로 이동할 수 있는 길을 열었다. 이렇게 등장한 새로운 형태의 두개골은 포유류의 청각 시스템의 일부라고 할 수 있다.또한 입천장에 이차 구개가 생겼는데, 이는 콧구멍에서 나오는 공기 흐름이 직접 통과하는 대신 입 뒤쪽으로 이동하게 되어 음식을 씹으면서 숨을 쉴 수 있게 되었다. 이는 지구상의 모든 포유류가 가지고 있는 형질이다.
5. 분포
가장 초기의 키노돈트는 차라소그나투스과에 속하는데, 이들은 주로 남아프리카 카루 누층군의 퇴적물에서 발견된다. 페름기 후기 키노돈트 화석은 남아프리카 이외의 지역에서는 비교적 드물게 발견된다. 이 중 가장 널리 분포하는 프로키노수쿠스는 남아공, 독일, 탄자니아, 잠비아, 러시아 등 다양한 곳에서 발견된다.[30] 비포유류 키노돈트는 남아메리카, 인도, 아프리카, 남극, 아시아, 유럽, 북아메리카에서 발견되었다.
참조
[1]
논문
Selective Factors Associated with the Origin of Fur and Feathers
[2]
논문
Microbiota and food residues including possible evidence of pre-mammalian hair in Upper Permian coprolites from Russia
[3]
논문
Palaeoneurological clues to the evolution of defining mammalian soft tissue traits
http://dx.doi.org/10[...]
2016-05-09
[4]
서적
The Evolution of the Human Placenta
Jay Schulkin
[5]
서적
Mesozoic Mammals: The First Two-Thirds of Mammalian History
University of California Press
1979-12-17
[6]
논문
Jurassic stem-mammal perinates and the origin of mammalian reproduction and growth
[7]
논문
New gliding mammaliaforms from the Jurassic
[8]
웹사이트
Bats
https://basicbiology[...]
2015
[9]
서적
Mammalian Evolution, Diversity and Systematics
https://www.degruyte[...]
De Gruyter
2018-10-22
[10]
논문
A Basal Nonmammaliaform Cynodont from the Permian of Zambia and the Origins of Mammalian Endocranial and Postcranial Anatomy
https://www.tandfonl[...]
2020-12-01
[11]
논문
The radiation of cynodonts and the ground plan of mammalian morphological diversity
2013-10-22
[12]
논문
Untangling the Multiple Ecological Radiations of Early Mammals
2019-10
[13]
간행물
The Late Triassic Record of Cynodonts: Time of Innovations in the Mammalian Lineage
http://link.springer[...]
Springer International Publishing
2021-05-24
[14]
웹사이트
Classification of R. Owen 1861
https://paleodb.org/[...]
[15]
웹사이트
Classification of B. S. Rubidge and C. A. Sidor 2001
https://paleodb.org/[...]
[16]
문서
A revision of the reptiles of the Karroo
1913
[17]
문서
A bibliographical list of Reptilia from the Karroo Beds of South Africa
1954
[18]
논문
The radiation of cynodonts and the ground plan of mammalian morphological diversity
[19]
논문
Distribution and diversity patterns of Triassic cynodonts (Therapsida, Cynodontia) in Gondwana
https://www.scienced[...]
[20]
서적
Chinese Fossil Vertebrates
https://books.google[...]
Columbia University Press
2019-11-30
[21]
서적
In the Shadow of the Dinosaurs: Early Mesozoic Tetrapods
Cambridge University Press
[22]
논문
A Late Triassic Traversodont Cynodont From the Newark Supergroup of North Carolina
https://geodir.ldeo.[...]
[23]
문서
Vertebrate burrow complexes from the Early Triassic Cynognathus Zone (Driekoppen Formation, Beaufort Group) of the Karoo Basin, South Africa
2001
[24]
웹인용
Classification of R. Owen 1861.
http://paleodb.org/c[...]
2023-12-21
[25]
문서
A revision of the reptiles of the Karroo
1913
[26]
문서
A bibliographical list of Reptilia from the Karroo Beds of South Africa
1954
[27]
논문
The radiation of cynodonts and the ground plan of mammalian morphological diversity
https://royalsociety[...]
2013-10-22
[28]
웹인용
https://academic.oup[...]
2023-11-26
[29]
논문
Microbiota and food residues including possible evidence of pre-mammalian hair in Upper Permian coprolites from Russia
https://www.idunn.no[...]
2016-10
[30]
논문
A Basal Nonmammaliaform Cynodont from the Permian of Zambia and the Origins of Mammalian Endocranial and Postcranial Anatomy
https://www.tandfonl[...]
2020-12-01
[31]
논문
The radiation of cynodonts and the ground plan of mammalian morphological diversity
https://royalsociety[...]
2013-10-22
[32]
서적
The Late Triassic Record of Cynodonts: Time of Innovations in the Mammalian Lineage
https://doi.org/10.1[...]
Springer International Publishing
2018
[33]
논문
Untangling the Multiple Ecological Radiations of Early Mammals
https://doi.org/10.1[...]
2019-10
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com