파란트로푸스 로부스투스
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
파란트로푸스 로부스투스는 플라이스토세에 살았던 멸종된 호미닌의 일종으로, 1938년 로버트 브룸에 의해 처음 발견되었다. 남아프리카에서 화석이 주로 발견되며, 튼튼한 턱과 큰 어금니를 특징으로 한다. 파란트로푸스 로부스투스는 오스트랄로피테쿠스 아프리카누스와 동시대에 살았으며, 뇌 용량은 고릴라와 비슷했다. 이들의 식성은 잡식성이었으며, 뼈 도구를 사용했을 가능성이 제기된다. 멸종 원인에 대해서는 기후 변화와 호모 속과의 경쟁 등이 거론된다.

더 읽어볼만한 페이지
- 파란트로푸스 - 파란트로푸스 아에티오피쿠스
파란트로푸스 아에티오피쿠스는 약 260만~330만 년 전 아프리카 동부에 살았던 파란트로푸스속의 초기 종으로, '강건한 오스트랄로피테쿠스'라고도 불리며 두개골 KNM WT 17000("블랙 스컬")을 통해 알려져 있고, 파란트로푸스 보이세이의 조상으로 여겨지나 분류학적 위치와 유효성에 대한 논쟁이 있으며 오스트랄로피테쿠스 아파렌시스와 유사한 특징을 가진다. - 파란트로푸스 - 파란트로푸스 보이세이
파란트로푸스 보이세이는 약 230만 년 전부터 134만 년 전까지 아프리카에 살았던 초기 인류의 한 종으로, 거대한 어금니와 튼튼한 두개골로 "호두까기 인간"이라는 별명을 가졌으며, 도구 사용 능력과 고생물학적 특징에 대한 연구가 진행 중이다. - 1938년 기재된 화석 분류군 - 파란트로푸스
파란트로푸스는 플라이오세 후기부터 홍적세 중기 아프리카에 살았던 멸종된 호미닌 속으로, 질긴 식물성 음식을 씹기에 적합한 튼튼한 두개골 구조를 지녔으며, 오스트랄로피테쿠스 및 초기 호모 속과 공존했고, 파란트로푸스 보이세이, 파란트로푸스 로부스투스, 파란트로푸스 에티오피쿠스 등의 종이 속한다. - 1938년 기재된 화석 분류군 - 레포파구스
레포파구스(Canis lepophagus)는 북아메리카에서 유래하여 유라시아로 퍼져나간 멸종된 개과 동물로, 늑대, 코요테, 황금 자칼의 조상으로 추정되며, 텍사스 등 여러 지역에서 화석이 발견된다. - 플라이스토세 아프리카의 포유류 - 호모 에렉투스
호모 에렉투스는 약 200만 년 전부터 10만 년 전까지 생존하며 직립 보행을 하고 아슐리안 석기를 제작했으며 불을 사용한 최초의 인류로 추정되는 종이다. - 플라이스토세 아프리카의 포유류 - 오스트랄로피테쿠스
오스트랄로피테쿠스는 420만 년 전부터 200만 년 전까지 아프리카에 살았던 초기 인류 속으로, 작은 뇌와 이족보행 능력을 지녔으며, 다양한 종과 화석 증거를 통해 호모 속의 조상일 가능성이 제기되는 등 인류 진화 연구에 중요한 자료이다.

2. 연구사
레이먼드 다트가 오스트랄로피테쿠스 아프리카누스를 발견한 이후, 스코틀랜드 출신 의사였던 로버트 브룸은 다트의 주장에 동의하며 남아프리카에서 발굴을 시작했다. 1938년, 브룸은 스콰르트크란스에서 다트가 발견한 오스트랄로피테쿠스 아프리카누스와 유사하지만 훨씬 더 튼튼한 형태의 두개골 조각과 치아 화석을 발견했다[107]. 이는 오스트랄로피테쿠스 아프리카누스에 이어 두 번째로 발견된 화석 유인원이자, 처음으로 발견된 '튼튼한(robust)' 형태의 화석 인류였다. 브룸의 발견은 호모 사피엔스로 이어지는 인류 진화의 과정이 하나의 직선적인 경로가 아니라 여러 갈래로 나뉘어 진행되었음을 보여주는 중요한 계기가 되었다. 이후 파란트로푸스 로부스투스 화석은 크롬드라이, 스콰르트크란스, 드리모렌, 곤돌린, 쿠퍼스 등 인류의 요람 지역 내 여러 동굴 유적에서 추가로 발견되었다. 스콰르트크란스 동굴에서는 130개체 분량으로 추정되는 뼈가 발견되었으며, 치아 형태 연구를 통해 이들 중 17세까지 생존한 개체는 드물었다는 것이 밝혀졌다.
2. 1. 발견

최초의 파란트로푸스 로부스투스 유해는 1938년 6월 남아프리카 공화국 크롬드라이 동굴 유적에서 발견되었다. 당시 지역 학생이었던 게르트 터블랑셰가 턱뼈 일부를 포함한 두개골 조각(TM 1517)을 찾아낸 것이다. 그는 이 유해를 남아프리카 보존 운동가 찰스 시드니 바로우에게 넘겼고, 바로우는 다시 고생물학자 로버트 브룸에게 전달했다.[3] 브룸은 유적 조사를 시작하여 몇 주 후 오른쪽 상완골 아래쪽 부분, 오른쪽 척골 위쪽 부분, 엄지발가락의 끝마디뼈(지골)를 추가로 발견했으며, 이들을 모두 TM 1517 개체에 속하는 것으로 보았다. 처음에는 개코원숭이의 것으로 여겨졌던 발가락뼈 하나도 나중에 TM 1517과 관련 있는 것으로 밝혀졌다.[2] 브룸은 크롬드라이에서 발견된 이 유해가 다른 호미닌들에 비해 유난히 튼튼하다는 점에 주목했다.[3]
1938년 8월, 브룸은 이 튼튼한 유해를 새로운 속으로 분류하고 '파란트로푸스 로부스투스(''Paranthropus robustus'')'라는 학명을 부여했다.[3] '파란트로푸스(''Paranthropus'')'라는 속명은 고대 그리스어로 '곁에' 또는 '나란히'를 뜻하는 παρα|paragrc와 '사람'을 뜻하는 άνθρωπος|anthroposgrc를 합쳐 만든 이름이다.[4]
당시 호주 출신 인류학자 레이먼드 다트는 1924년 타웅 아이 화석을 근거로 오스트랄로피테쿠스 아프리카누스(''Australopithecus africanus'')가 인류의 조상이라고 주장했지만, 이는 학계에서 큰 논란거리였다.[5] 브룸 역시 1936년 크롬드라이에서 서쪽으로 약 2km 떨어진 스터크폰테인 동굴에서 '플레시안트로푸스 트란스발렌시스(''Plesianthropus transvaalensis'')'를 발견했는데, 이 종은 현재 ''A. africanus''와 같은 종(동의어)으로 여겨진다. 이들 종은 모두 플라이스토세 시기의 화석이며, 같은 지역(현재 '인류의 요람'으로 불림)에서 발견되었다. 브룸은 이러한 발견들을 플리오세 시대 인류 조상의 다양성을 보여주는 증거로 해석했으며, 여러 호미닌 분류군이 초기 인류와 공존했을 가능성을 시사한다고 보았다.[3]
파란트로푸스 로부스투스로 분류된 화석들은 1948년 근처의 스와르트크란스 동굴에서도 발견되기 시작했다. 20세기 초까지는 크롬드라이와 스와르트크란스에서만 확실하게 확인되었으나, 이후 인류의 요람 내 다른 지역인 스터크폰테인, 곤돌린 동굴, 쿠퍼스 동굴, 드리몰렌 동굴에서도 이 종의 화석이 보고되었다. 현재까지 파란트로푸스 로부스투스 화석은 이 제한된 지역 밖에서는 발견되지 않았다.[10] 스와르트크란스 동굴에서는 130개체 이상에 해당하는 뼈가 발견되었으며[96], 치아 형태 연구 결과 17세까지 생존한 개체는 드물었던 것으로 추정된다.
파란트로푸스 로부스투스는 오스트랄로피테쿠스 아프리카누스에 이어 두 번째로 발견된 고인류 유인원이자, 처음으로 발견된 '튼튼한(robust)' 형태의 화석 인류였다. 브룸의 발견은 호모 사피엔스로 이어지는 인류 진화의 과정이 하나의 직선적인 경로가 아니라 여러 갈래로 나뉘어 진행되었음을 보여주는 중요한 계기가 되었다.
2. 2. ''P. crassidens'' 논쟁
1948년, 로버트 브룸은 스와르트크란스 동굴에서 발견된 어린 개체의 하악골(SK 6)을 근거로 ''Paranthropus crassidens''(''P. crassidens'')라는 새로운 종을 제안했다.[10] 그는 스와르트크란스 동굴과 인근의 크롬드라이 동굴에서 발견된 동물군 화석들의 차이점을 바탕으로 두 동굴이 서로 다른 시기에 형성되었다고 판단하여 ''P. robustus''와는 다른 종으로 본 것이다.[6]당시 인류와 그 조상들은 사람과(Hominidae)로, 인간이 아닌 유인원들은 "오랑우탄과(Pongidae)"로 분류되었다. 1950년, 브룸은 초기 인류를 오스트랄로피테쿠스아과(''Au. africanus''와 "''Pl. transvaalensis''"), 파란트로푸스아과(''Pa. robustus''와 "''Pa. crassidens''"), 아르칸트로푸스아과("''Au. prometheus''")의 세 아과로 나누자고 제안했다.[7] 그러나 이 분류 체계는 종을 너무 세분화한다는 이유로 널리 비판받았다.[11] 또한, 발견된 화석들의 정확한 연대가 확정되지 않았고, 인류 진화 과정이 여러 계통으로 나뉘는지 아니면 단일 계통인지에 대한 논쟁이 있었다. 이러한 상황 속에서 브룸과 남아프리카의 고생물학자 존 탤벗 로빈슨은 ''파란트로푸스''(''Paranthropus'') 속이 타당하다고 계속 주장했다.[8]
1951년, 인류학자 셔우드 워시번과 브루스 D. 패터슨은 ''파란트로푸스''를 ''오스트랄로피테쿠스'' 속과 같은 것으로 보고, 인류의 속을 ''오스트랄로피테쿠스''와 ''호모''만으로 제한하자고 처음 제안했다.[9] 이후 ''파란트로푸스''가 ''오스트랄로피테쿠스''의 주니어 동의어인지에 대한 논쟁이 계속되었다.[10] 인류 분류 기준을 좀 더 엄격하게 적용하기 위해, 1954년 로빈슨은 "''P. crassidens''"를 아종 수준인 "''P. robustus crassidens''"(''P. r. crassidens'')로 강등시켰다.[11]
한편, 1949년 브룸과 로빈슨은 스와르트크란스 동굴에서 또 다른 하악골을 발견하고 이를 "원숭이 인간과 진정한 인간 사이의 중간 단계"로 잠정 해석하며 새로운 속과 종인 "''Telanthropus capensis''"로 명명했다. 당시 대부분의 학자들은 이 화석 역시 스와르트크란스에서 많이 발견된 "''P. crassidens''"와 같은 종으로 보았다.[13] 그러나 1957년, 로빈슨은 (구체적인 이유는 밝히지 않았지만) 이를 ''호모 에렉투스''(오늘날 아프리카의 ''H. erectus''는 때때로 ''H. ergaster''로 불림)와 같은 종으로 분류했다. 1965년, 남아프리카의 고생물학자 필립 V. 토비아스는 이러한 분류의 타당성에 의문을 제기하기도 했다.[14]
21세기에 이르러, "''P. crassidens''"라는 명칭은 대부분 사용되지 않고 ''P. robustus''로 통합되어 불리는 경향이 강해졌다. 하지만 미국의 고생물학자 프레데릭 E. 그린과 같은 일부 학자들은 여전히 ''P. crassidens''를 ''P. robustus''와는 구별되는 독립적인 종으로 보아야 한다고 주장하고 있다.[10]
2. 3. ''기가토피테쿠스''와의 관계
1939년, 브룸은 ''파란트로푸스 로부스투스''가 아시아에서 발견된, 유사하게 큰 이빨을 가진 멸종 유인원인 ''기가토피테쿠스''와 밀접한 관련이 있다고 가설을 세웠다. 당시 그는 ''기가토피테쿠스''를 호미닌으로 생각했다.[15] 이후 20세기 중반까지 ''기가토피테쿠스''가 호미닌인지, 아니면 인간이 아닌 유인원인지에 대한 많은 논쟁이 있었다. 특히 유대계 독일 인류학자 프란츠 바이덴라이히와 독일-네덜란드 고생물학자 랄프 폰 쾨니히스발트는 ''기가토피테쿠스''가 아시아의 ''H. erectus''의 직접적인 조상이거나 밀접한 관련이 있다고 주장했다.[16]1972년, 로빈슨은 미오세 시대 파키스탄에서 발견된 ''기가토피테쿠스 빌라스푸렌시스''("''G. bilaspurensis''", 현재는 ''인도피테쿠스''로 분류)를 ''파란트로푸스''와 중국의 ''기가토피테쿠스 블랙아이''(''G. blacki'')의 조상으로 보고, ''기가토피테쿠스''를 "파란트로피나이(Paranthropinae)" 아과에 포함시키자고 제안했다. 그는 이들 모두 거대한 체격을 가졌다고 믿었다. 반면, 그는 ''오스트랄로피테쿠스 아프리카누스''(그는 "''호모'' ''아프리카누스''"라고 불렀다)는 매우 작은 체격을 가졌으며, 인간 계통의 일원으로서 문화적 능력과 사냥 능력이 있었지만 "파란트로피네"는 그렇지 않았다고 추측했다.[49]
그러나 1970년대 후반에서 1980년대에 걸쳐 분지학이 널리 사용되고 미오세 유인원과 후기 유인원 간의 관계가 더 명확해지면서, ''기가토피테쿠스''는 호미니나에에서 완전히 제외되었다. 현재 ''기가토피테쿠스''는 오랑우탄과 함께 퐁기나에 아과에 속하는 것으로 분류된다.[16]
2. 4. ''P. boisei''의 발견과 분류 논쟁
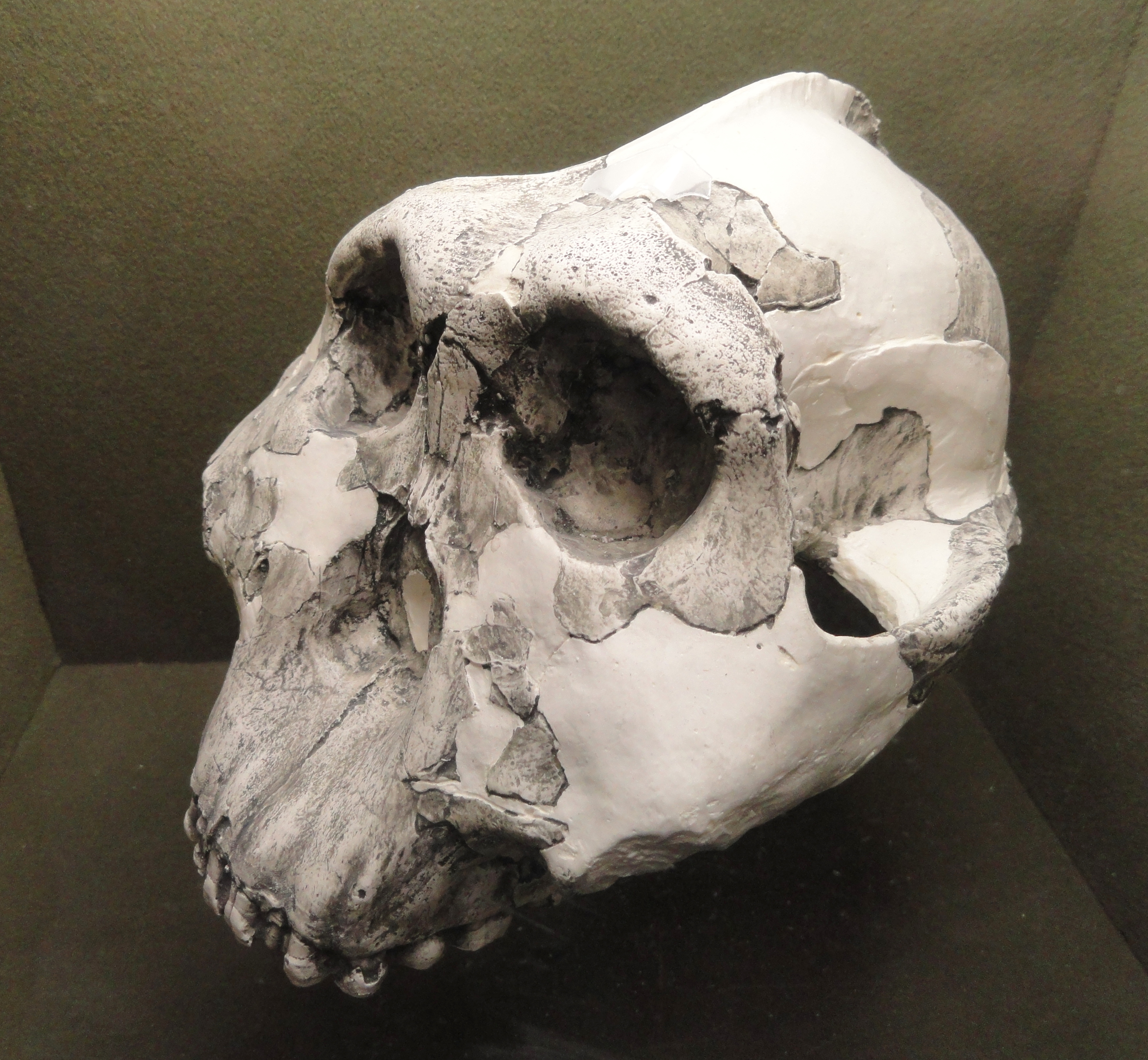
1959년 동아프리카에서는 ''파란트로푸스 로부스투스''보다 훨씬 더 강건한 특징을 가진 새로운 오스트랄로피테쿠스류인 ''P. boisei''가 발견되었다. 이후 1975년에는 ''P. boisei''의 두개골 KNM-ER 406이 인류의 조상으로 여겨지는 ''호모 에르가스테르'' 또는 ''호모 에렉투스''의 두개골 KNM ER 3733과 같은 시기에 존재했다는 사실이 밝혀졌다. 이는 일반적으로 ''Paranthropus'' 속이 ''Australopithecus'' 속의 특정 종(당시에는 ''A. africanus''만 알려져 있었음)에서 진화하여 ''Homo'' 속의 자매 분류군이라는 견해를 뒷받침하는 증거로 여겨졌다.[8]
그러나 파란트로푸스 속의 분류와 계통 발생에 대해서는 학자들 사이에 여러 의견이 존재한다. 1979년, ''A. afarensis''를 발표한 지 1년 후, 인류학자 도널드 조한슨과 팀 D. 화이트는 ''A. afarensis''가 ''Homo'' 속과 ''Paranthropus'' 속의 마지막 공통 조상이며, ''A. africanus''는 ''Paranthropus'' 계통의 가장 초기 구성원이거나 최소한 ''P. robustus''의 조상일 것이라고 주장했다. 이는 ''A. africanus''가 ''P. robustus''보다 먼저 남아프리카에 살았고, 당시 ''A. afarensis''가 약 350만 년 전으로 가장 오래된 호미닌 종으로 알려졌기 때문이었다.[8] 하지만 현재 가장 오래된 남아프리카 오스트랄로피테쿠스류("리틀 풋")는 367만 년 전으로 거슬러 올라가 ''A. afarensis''와 동시대에 존재했다는 것이 밝혀지면서[17], 이 문제는 여전히 활발히 논의되고 있다.[18]
오랫동안 ''Paranthropus'' 속이 유효하다면 ''P. robustus''가 ''P. boisei''의 조상일 것이라고 추정되었다. 하지만 1985년, 인류학자 앨런 워커와 리처드 리키는 약 250만 년 전의 동아프리카 두개골 KNM WT 17000을 발견하고, 이를 새로운 종 ''A. aethiopicus''로 명명했다. 그들은 이 종이 ''P. boisei''의 조상이라고 주장하며(이들은 ''Paranthropus''를 ''Australopithecus''와 동일한 속으로 간주했다), ''P. boisei'' 계통이 ''P. robustus''가 등장하기 훨씬 이전에 시작되었음을 보여주었다.[19] 이러한 발견들은 파란트로푸스 속 내의 종들 간의 관계와 전체적인 인류 진화 과정에 대한 이해를 더욱 복잡하게 만들었으며, 관련 연구와 논쟁은 계속되고 있다.
3. 형태
''파란트로푸스 로부스투스''는 파란트로푸스 속의 전형적인 외형을 가지고 있으며, 두개골 형태는 고릴라와 유사하지만 더 튼튼한 턱과 인간에 가까운 교합면의 치아를 가졌다. 수컷의 경우 머리뼈 가운데에 시상 능선이 두드러지게 발달했는데, 이는 강력한 씹기 근육을 지탱하기 위한 구조로 고릴라에게서도 나타나는 특징이다.[108]
수컷과 암컷의 크기 차이가 뚜렷한 성적 이형성을 보인다. 치아 크기는 동아프리카의 ''P. boisei''와 비슷했다.[108] 뇌 용량은 현생 고릴라와 비슷한 수준이었으며,[108] 튼튼한 턱과 큰 어금니는 견과류나 식물 뿌리처럼 단단한 음식을 먹는 데 적합했을 것으로 추정된다.
3. 1. 두개골
''파란트로푸스 로부스투스''(''Paranthropus robustus'')는 전형적으로 거대한 뺨니를 가진 송곳니 이후 거대치아증을 보이지만, 사람 크기의 앞니와 송곳니를 가지고 있다. 작은 어금니는 어금니와 같은 모양이다.[23] 뺨니의 에나멜 두께는 현대 인간과 거의 같지만, 오스트랄로피테쿠스 뺨니 에나멜은 특히 첨두(cusp)의 끝 부분에서 두꺼워지는 반면, 인간은 첨두의 기저부에서 두꺼워진다.[22]
''P. robustus''는 약간의 전돌증 (턱이 약간 돌출됨)을 가진 키가 큰 얼굴을 가지고 있다. 수컷의 두개골은 두개골 덮개의 정중선에 잘 발달된 시상 능선과 부풀어 오른 뺨 뼈를 가지고 있으며, 이는 물기에 중요한 거대한 측두근을 지탱했을 가능성이 있다.[23] 시상 능선은 고릴라와 공통적인 형질이다.[108] 뺨이 얼굴에서 너무 멀리 튀어나와서, 위에서 보면 코가 오목한 부분의 바닥에 있는 것처럼 보인다 (접시 모양의 얼굴). 이것은 눈 구멍을 약간 앞으로 이동시켜 약한 눈썹 능선과 후퇴하는 이마를 유발했다. 부풀어 오른 뺨은 또한 교근 (아래로 물 때 중요)을 앞으로 밀어내고 치열을 뒤로 밀어내어 작은 어금니에 더 높은 물기 힘을 만들었을 것이다. 아래턱을 위턱에 연결하는 턱뼈의 지점인 턱뼈가지가 높아서 지레 팔 (및 토크)을 증가시켰을 것이고, 이는 교근과 내측익돌근 (둘 다 아래로 물 때 중요)의 토크를 증가시켜 물기 힘을 더욱 증가시켰을 것이다.[23]
잘 발달된 시상 능선과 부풀어 오른 뺨은 암컷으로 추정되는 두개골 DNH-7에는 없으므로, Keyser는 수컷 ''P. robustus''가 암컷보다 더 튼튼했을 수 있다고 제안했다. (''P. robustus''는 성적 이형성을 보였다).[102] Eurydice라고 명명된 암컷 두개골 화석(DNH-7)은 1994년 남아프리카 공화국의 드리모렌 동굴에서 앙드레 카이저 등에 의해 발견되었으며, 230만 년 전의 것으로 추정된다.[108]
드라이몰렌(Drimolen)에서 발견된 ''P. robustus'' 재료는 더 기초적인 특징을 보여 비교적 가늘고, 따라서 더 후대의 스와르트크란스(Swartkrans)와 크롬드라이(Kromdraii) ''P. robustus''보다 물기 힘이 더 작았을 것으로 추정된다. 드라이몰렌 표본의 눈썹 또한 각진 것이 아니라 둥글고, 수컷으로 추정되는 DNH 155의 시상 능선은 더 뒤쪽 (머리 뒤쪽)에 위치해 있다.[24]
SK 46과 SK 47의 후반고리관 형태는 유인원 같은 ''오스트랄로피테쿠스''(Australopithecus)나 ''호모''(Homo)와 다르다. 내이(inner ear) 해부학은 전정 시스템 (균형 감각)에 영향을 미치므로, 이는 ''P. robustus''가 다른 운동 패턴과 머리 움직임 패턴을 가졌음을 시사한다. 현대 인간의 후반고리관은 달리는 동안 안정화에 도움이 되는 것으로 생각되는데, 이는 ''P. robustus''가 지구력 주자가 아니었을 수 있음을 의미할 수 있다.[25]

3. 2. 뇌
파란트로푸스 로부스투스를 처음 기술할 때, 브룸은 TM 1517 표본의 조각난 뇌 케이스를 바탕으로 뇌 용량을 600cc로 추정했다.[3] 1946년에는 남아프리카 인류학자 게릿 빌렘 헨드릭 셰퍼스와 함께 이 추정치를 575–680cc로 수정했다.[27] 이는 당시 알려진 호모 속의 뇌 용량 500-900cc와 비교되는 수치였다.[26] 그러나 1년 후, 영국의 영장류학자 윌프리드 르 그로 클라크는 해당 표본이 한쪽 측두골 일부만 남아있어 정확한 뇌 용량 측정이 불가능하다고 지적했다.[27] 2001년, 폴란드 인류학자 카타르지나 카지츠카는 브룸이 초기 호미닌의 뇌 크기를 실제보다 상당히 부풀렸을 가능성을 제기했다.[28]1972년, 미국의 생물 인류학자 랄프 할로웨이는 전두엽 일부가 없는 두개골 덮개 SK 1585를 연구하여 약 530cc의 뇌 용량을 보고했다. 그는 또한 다른 오스트랄로피테쿠스와 비교했을 때 파란트로푸스가 호모 속과 유사하게 확장된 소뇌를 가졌을 수 있다고 언급했는데, 이는 1967년 토비아스가 ''P. 보이세이'' 두개골 연구에서 처음 제기한 내용이었다.[29] 2000년, 미국의 신경인류학자 딘 팔크와 동료들은 ''P. 보이세이'' 표본들을 이용해 SK 1585의 누락된 전두엽 부분을 복원하고 뇌 용량을 약 476cc로 재계산했다. 그들은 ''P. 로부스투스''의 전반적인 뇌 구조가 현생인류보다는 다른 유인원에 더 가깝다고 결론지었다.[30] 2020년에 발견된 거의 완전한 두개골 DNH 155의 뇌 용량은 450cc로 측정되었다.[24]
현재까지의 연구 결과들을 종합하면, ''P. 로부스투스''의 평균적인 뇌 크기는 410~530cc 정도로 추정되며, 이는 현생 고릴라와 비슷하거나 약간 작은 수준이다.
뇌 혈관 구조에 대한 연구도 진행되었다. 1983년, 프랑스 인류학자 로저 사반은 SK 1585와 KNM-ER 407(''P. 보이세이'') 표본을 연구하면서, 중간 뇌막 동맥의 두정 분지가 초기 호미닌과 달리 앞쪽 가지가 아닌 뒤쪽 가지에서 시작된다는 점을 발견했다. 그는 이것이 뇌 용량 증가와 관련된 파생 형질일 수 있다고 보았다.[31] 그러나 이후 연구에서 적어도 ''P. 보이세이''의 경우, 두정 분지가 앞쪽 또는 뒤쪽 가지 모두에서 시작될 수 있으며, 심지어 한 개체 내에서도 양쪽이 다르게 나타날 수 있다는 사실이 밝혀졌다.[32]
경막 정맥동과 관련해서는, 1983년 팔크와 인류학자 글렌 콘로이가 ''A. 아프리카누스''나 현생 인류와 달리 모든 ''파란트로푸스'' 속(및 ''A. 아파렌시스'')은 확장된 후두동과 가장자리 정맥동(대후두공 주변)을 가지며, 이것이 횡정맥동과 S상 정맥동을 완전히 대체한다고 주장했다. 그들은 이러한 구조가 이족 보행에 대한 초기 적응 과정에서 머리로 가는 혈류를 조절하는 방식의 변화와 관련이 있을 것으로 추정했다.[33] 하지만 1988년 팔크와 토비아스는 초기 호미닌들이 후두/가장자리 시스템과 횡/S상 시스템을 동시에 가지거나, 혹은 두개골의 양쪽에서 다르게 가질 수도 있음을 보여주었다.[34]
3. 3. 신체
''파란트로푸스 로부스투스''는 뚜렷한 성적 이형성을 보인다. 수컷은 신장 약 120cm, 체중 약 54kg으로 추정되는 반면, 암컷은 신장 약 100cm, 체중 약 40kg으로 추정된다.[108] 이는 과거 여러 학자들의 추정치와 차이가 있는데, 예를 들어 1972년 로빈슨은 키 140cm~150cm, 몸무게 68kg~91kg으로 매우 크게 추정했으나[49], 이후 연구들에서 점차 작은 값으로 수정되었다. 1991년 헨리 맥헨리는 수컷 평균 키 132cm, 암컷 평균 키 110cm를 보고했으며[54], 2001년 서스먼 등은 수컷 평균 42kg, 암컷 평균 30kg을 추정했다.[55] 2015년 그라보스키 등의 연구에서는 수컷 평균 32.3kg, 암컷 평균 24kg으로 제시하기도 했다.[57] 이러한 다양한 추정치는 사용된 화석 표본과 분석 방법에 따라 달라질 수 있음을 보여준다.두개골 형태는 고릴라와 유사하지만, 더 튼튼한 턱과 인간에 가까운 교합면의 치아를 가지고 있다.[108] 특히 수컷의 경우, 강력한 저작근(씹는 근육)을 지탱하기 위해 머리 꼭대기를 따라 시상능이 발달해 있었다.[108] 뇌 용량은 평균 410~530ml 정도로, 고릴라와 비슷한 수준이었다.[108] 치아 크기는 동아프리카의 ''P. boisei''와 유사했다.[108]
척추 화석(SKW 14002, SK 3981 등) 분석 결과, 제12흉추는 다른 오스트랄로피테쿠스보다 높이가 더 압축되어 있었다.[36] 요추는 다른 오스트랄로피테쿠스보다 ''H. 에르가스터''/''H. 에렉투스'' 및 현생 인류와 더 유사한 형태를 보이며, 튼튼한 척주와 횡돌기는 강력한 장요인대의 존재를 시사하고 이는 직립 보행 자세와 관련이 있을 수 있다.[36]
골반은 ''A. 아프리카누스''나 ''A. 아파렌시스''와 유사하지만, 더 넓은 엉덩뼈 날개와 더 작은 비구(고관절 소켓) 및 엉덩이 관절을 가진다.[44]
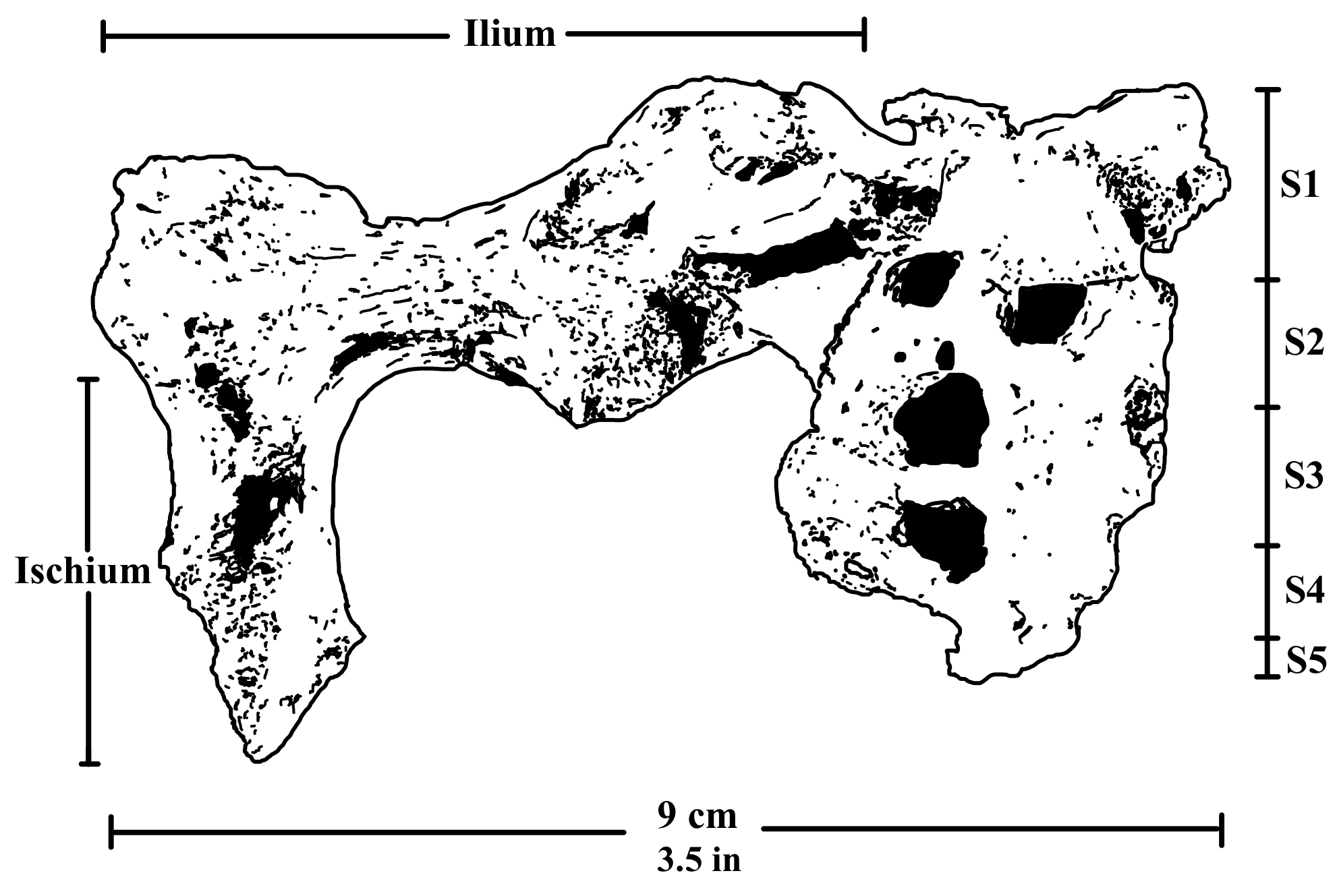
팔의 상완골(윗팔뼈)은 현생 인류 및 침팬지와 유사한 형태를 보인다.[39] 요골(아래팔뼈)은 다른 ''오스트랄로피테쿠스'' 종들과 비슷하다. 손목 관절은 현생 인류와 비슷한 가동성을 가졌지만, 팔꿈치 부위(요골 머리)는 팔을 굽혔을 때 안정성을 유지하는 데 능숙했던 것으로 보여, 부분적인 수목 생활(나무 위 생활)을 했을 가능성을 시사한다.[40] 손에서는 강력한 요골 경상 돌기가 확인되며, 이는 강한 팔 근육과 관련된다. 손가락 뼈는 현생 인류처럼 덜 휘어 있고 근육 부착 부위가 약해, 비인간 유인원이나 ''P. boisei''에 비해 나무를 타는 능력이 감소했을 수 있다.[41][42] 반면, 손의 구조는 현생 인류와 유사한 정밀 파지(정교하게 쥐기)가 가능했음을 시사하며, 이는 도구 제작이나 사용과 관련될 수 있다.[43]
대퇴골(넙다리뼈)은 앞뒤로 납작한 형태를 보이는데, 이는 현생 인류보다 덜 효율적인 방식의 보행을 했을 가능성을 나타낸다.[44] 그러나 대퇴골의 미세 구조 분석 결과, 고관절의 이동성은 비인간 유인원보다 감소했으며 이는 인간과 유사한 이족 보행과 일치한다.[45] StW 311 대퇴골두 화석의 마모 패턴은 습관적으로 고관절을 매우 구부린 자세를 취했음을 시사하는데, 이는 빈번한 등반 활동이나 쪼그리고 앉는 자세와 관련될 수 있다.[46] 슬개골(무릎뼈) SKX 1084의 질감 복잡성 분석 결과는 무릎 관절 사용 및 이족 보행 수준이 현생 인류와 침팬지의 중간 정도였음을 시사한다.[47] 발의 엄지발가락 뼈는 물건을 잡는 데 능숙하지 않았으며, 이는 인간과 유사한 발 자세와 운동 범위를 가졌음을 나타낸다. 그러나 발목 관절 구조는 현생 인류의 특징적인 발뒤꿈치를 들고 발가락으로 미는(toe-off) 보행 방식을 제한했을 수 있다. 전반적으로 ''P. robustus''는 ''H. habilis''와 비슷한 수준의 이족 보행 능력을 가졌을 것으로 추정된다.[48]
4. 생태
''파란트로푸스 로부스투스''는 플라이스토세 시기 남아프리카의 사바나와 숲이 혼합된 환경에서 서식했던 것으로 보인다. 이들의 생태는 발견된 화석과 유적을 통해 추정되며, 특히 식성, 사회 구조, 도구 사용, 그리고 포식자와의 관계 등이 주요 연구 주제이다.
튼튼한 턱과 큰 어금니 때문에 초기에는 견과류와 같은 단단한 음식을 주로 먹는 전문적인 섭식자로 여겨졌으나, 최근 연구들은 이들이 잡식성에 가까운 일반적인 섭식자였으며, 특정 시기에만 단단한 음식을 먹었을 가능성을 제시한다. 식단에는 계절에 따라 C4 사바나 식물과 C3 숲 식물이 모두 포함되었을 것으로 추정된다. (자세한 내용은 식성 문단 참고)
사회 구조에 대해서는 뚜렷한 성적 이형성(수컷이 암컷보다 훨씬 컸던 특징)을 근거로 여러 가설이 제기되었다. 수컷 한 마리가 여러 암컷을 거느리는 하렘과 유사한 사회였을 가능성과, 스트론튬 동위원소 분석 결과를 바탕으로 암컷이 태어난 집단을 떠나는 부계 거주 사회였을 가능성 등이 논의되고 있다. (자세한 내용은 사회 구조 문단 참고)
인류의 요람과 같은 유적지에서는 이들의 화석과 함께 뼈 도구가 발견되기도 한다. 이 뼈 도구는 ''P. robustus''가 식물 가공이나 땅 파기 등에 사용했을 가능성이 있으며, 이는 이들이 단순하지만 도구를 활용하는 능력이 있었음을 시사한다. 불을 사용했는지에 대해서는 논쟁이 있다. (자세한 내용은 기술 문단 참고)
또한, ''P. robustus''는 표범, 검치호의 일종인 ''메간테레온'', 점박이 하이에나 등 당대의 대형 육식동물에게 주요 사냥 대상이었을 것으로 보인다. 동굴에서 발견되는 많은 ''P. robustus''의 뼈는 이들 포식자가 사냥 후 남긴 흔적으로 해석되기도 한다. (자세한 내용은 포식자 문단 참고)
4. 1. 식성

1954년 로빈슨은 ''파란트로푸스 로부스투스''의 튼튼한 두개골과 강력한 씹는 힘이 견과류처럼 단단한 음식을 자주 먹는 데 특화된 전문적인 식단을 나타낸다고 주장했다. 이 때문에 20세기 후반에는 ''파란트로푸스''가 플라이스토세의 변덕스러운 기후에 적응하지 못해 멸종했다는 설명이 지배적이었는데, 이는 적응력이 더 뛰어났던 ''호모'' 속과는 대조적이었다. 이후 연구자들은 얼굴 근육, 치아 마모 패턴, 영장류 생태학 연구를 통해 이 모델을 뒷받침했다.[59]
그러나 1981년 영국의 인류학자 앨런 워커는 ''P. boisei'' 두개골 연구를 통해, 씹는 힘은 가해지는 총 압력뿐 아니라 압력이 분산되는 치아 표면적과도 관련 있음을 지적했다. ''파란트로푸스''의 치아는 현대 인류보다 4~5배 크기 때문에, 워커는 튼튼한 구조가 단단한 음식이 아니라 한 번에 많은 양의 음식을 씹기 위한 적응일 수 있다고 주장했다. 또한 ''P. boisei'' 어금니의 미세 마모 패턴이 맨드릴, 침팬지, 오랑우탄과 구별되지 않는다는 점도 발견했다.[58] 그럼에도 불구하고, ''파란트로푸스''가 전문적인 섭식자가 아니라는 주장이 제기되었음에도 로빈슨의 초기 모델은 20세기 동안 지배적인 견해로 남았다.[59]
2004년 인류학자 버나드 우드와 데이비드 스트레이트는 ''파란트로푸스'' 식단 관련 연구들을 검토한 결과, ''파란트로푸스''가 실제로는 일반적인 섭식자였으며, 특히 ''P. robustus''는 잡식성이었을 가능성이 높다고 결론지었다. 그들은 ''P. robustus''의 치아 미세 마모 패턴이 단단한 음식을 자주 섭취하지 않았음을 시사하며, 따라서 튼튼한 두개골 구조는 구하기 어려운 시기에 덜 선호하는 대체 음식을 먹을 때만 중요했을 것이라고 보았다.[59] 이러한 전략은 현대 고릴라가 사용하는 것과 유사하다. 고릴라는 가볍게 지어진 침팬지(그리고 아마도 가늘어진 오스트랄로피테쿠스)와 달리, 연중 내내 저품질의 대체 음식만으로도 생존할 수 있다. 반면 침팬지는 고품질 음식을 꾸준히 섭취해야 한다.[60]
한편, 1980년 인류학자 톰 해틀리와 존 캐펠만은 초기 호미닌이 뿌리나 덩이줄기 같은 거칠고 칼로리가 높은 지하 저장 기관 (USO)을 먹는 데 적응했을 수 있다고 제안했다.[61] 이후 호미닌의 USO 활용 가설은 더 많은 지지를 받았다. 2005년 생물학적 인류학자 그레그 레이든과 리처드 랭엄은 ''파란트로푸스''가 USO를 대체 식량원 또는 주요 식량원으로 의존했을 수 있으며, USO가 풍부한 지역과 호미닌 서식지 사이에 연관성이 있을 수 있다고 주장했다.[60]
2006년 탄소 동위 원소 분석 결과에 따르면, ''P. robustus''는 계절에 따라 C4 사바나 식물과 C3 숲 식물을 번갈아 섭취했던 것으로 보인다. 이는 식단의 계절적 변화 또는 계절에 따른 서식지 이동(숲과 사바나 사이)을 시사한다.[63] 또한, 동시대의 ''호모 에르가스터/호모 에렉투스'' 역시 ''P. robustus''와 비슷한 비율로 C3 및 C4 기반 음식을 섭취한 것으로 나타났다.[62]
''P. robustus''는 씨앗이나 견과류 같은 단단한 음식을 상당히 자주 먹었을 가능성이 있는데, 이는 치아 파절률이 중간 정도(239개 표본 중 약 12%)로 나타났기 때문이다. 이는 ''P. boisei''에서는 거의 발견되지 않는 특징이다.[63][64] 높은 충치 발생률은 꿀과 같은 당분이 많은 음식을 섭취했을 가능성을 시사한다.[89]
어린 ''P. robustus'' 개체는 성체보다 덩이줄기에 더 의존했을 수 있다. Swartkrans 동굴에서 발견된 어린 개체 치아의 스트론튬 수치가 높은데, 이는 해당 지역에서 덩이줄기를 통해 가장 많이 섭취될 수 있는 원소이기 때문이다. 어린 개체 치아의 상아질 노출은 일찍 젖을 뗐거나, 성체보다 더 거친 음식을 먹어 시멘트질과 에나멜 코팅이 마모되었음을 나타낼 수 있다. 또는 땅에서 파낸 음식에 묻은 모래를 제대로 제거하지 못했을 가능성도 있다.[71] 초기 추정에서는 견과류나 괴근과 같은 단단한 음식을 주로 먹었을 것으로 여겨지기도 했다.
4. 2. 사회 구조
현생 유인원과는 해부학적 및 신체적으로 뚜렷한 차이가 있어, 오스트랄로피테쿠스류의 사회 구조를 현생 영장류와 직접 비교하여 추론하는 것은 매우 어렵고 추측에 기댈 수밖에 없다.[66][67]2007년 인류학자 찰스 록우드와 동료들은 ''파란트로푸스 로부스투스''에서 수컷이 암컷보다 현저히 큰 뚜렷한 성적 이형성이 나타난다는 점에 주목했다. 이러한 특징은 일반적으로 수컷이 지배하는 일부다처제 사회, 특히 고릴라의 실버백처럼 한 마리의 수컷이 여러 암컷에 대한 번식 독점권을 가지는 하렘 사회와 관련이 깊다. ''P. 로부스투스'' 수컷과 암컷의 크기 차이(얼굴 치수 기준)는 고릴라와 유사하며, 어린 수컷은 나이 든 수컷보다 덜 건장한 모습을 보였다. 이는 고릴라에게서도 나타나는 성숙 지연 현상이다. 성별이 확인된 ''P. 로부스투스'' 표본 중 수컷(또는 수컷으로 추정되는 개체)이 대다수를 차지하는데, 이는 수컷의 사망률이 암컷보다 높았음을 시사할 수 있다. 하렘 사회에서는 암컷을 둘러싼 수컷 간 경쟁이 치열해지면서 무리에서 쫓겨나는 수컷이 발생할 가능성이 높고, 홀로 남겨진 수컷은 포식의 위험에 더 크게 노출되었을 수 있다. 이 가설에 따르면, 태어난 집단을 떠난 암컷은 혼자 있는 시간이 거의 없이 즉시 다른 기존 집단으로 합류했을 것이다.[65]

그러나 2011년 고인류학자 샌디 코플랜드와 동료들은 스트론튬 동위원소 비율 분석을 통해 다른 결론에 도달했다. 남아프리카 스테르크폰테인 계곡의 백운암 지대에서 발견된 ''P. 로부스투스'' 치아를 분석한 결과, 다른 유인원 사회와는 달리 암컷이 태어난 곳을 떠날 가능성이 더 높은 부계 거주 사회였을 가능성이 제기되었다.[66] 이는 수컷 간 경쟁이 심화될 경우 모계 거주 사회가 될 가능성이 높은 하렘 사회 가설과는 배치되는 결과이다. 수컷들은 태어난 계곡에서 멀리 벗어나지 않은 것으로 보이는데, 이는 활동 반경이 작았거나, 동굴이 풍부하거나 식물 성장에 유리한 백운암 지형을 선호했음을 시사할 수 있다.[66]
이에 더해 2016년 폴란드 인류학자 카타르지나 카시치카는 성숙 지연 현상이 붉은털원숭이와 같이 여러 수컷이 함께 생활하는 사회에서도 나타나므로, 이것만으로 사회 구조를 단정하기는 어렵다고 지적했다. 만약 ''P. 로부스투스''가 사바나 환경을 선호했다면, 여러 수컷이 함께 있는 사회가 탁 트인 환경에서 포식자로부터 무리를 방어하는 데 더 유리했을 수 있으며, 이는 사바나에 사는 개코원숭이와 유사한 형태이다. 여러 수컷 사회에서도 경쟁에서 밀려난 수컷이 무리에서 쫓겨날 가능성이 있으며, 이는 앞서 언급된 수컷 편향적인 사망률을 설명할 수 있는 또 다른 가능성을 제시한다.[67]
한편, 2017년 인류학자 캐서린 발로리아와 동료들은 시상능의 역할에 주목했다. 수컷 비인간 유인원(특히 고릴라와 오랑우탄)이 암컷보다 더 큰 시상능을 가지는 경향이 있다는 점에서, 시상능이 단순히 씹는 근육을 지지하는 기능 외에도 성적 선택의 영향을 받았을 수 있다고 보았다. 실제로 수컷 서부로랜드고릴라의 시상능 크기는 번식 성공률과 관련이 있는 것으로 나타났다. 발로리아 등은 이러한 해석을 ''파란트로푸스'' 수컷에게도 적용하여, 시상능과 그로 인해 커 보이는 머리가 일종의 과시 행동에 사용되었을 수 있다고 추정했다. 이는 일반적으로 확대된 송곳니를 과시하는 다른 영장류와는 다른 방식인데, ''파란트로푸스''는 송곳니가 비교적 작았기 때문에 이러한 과시 행동이 어려웠을 것이다. 다만, 수컷 고릴라와 오랑우탄의 경우 더 넓은 입 벌림으로 송곳니를 효과적으로 과시하기 위해 더 큰 측두근이 필요했고, 그 결과 시상능이 발달했을 가능성도 있다.[68]
4. 3. 기술
''P. robustus''의 손은 인간과 유사한 정밀 파지 능력을 갖추고 있었던 것으로 보이며, 이는 비인간 영장류의 도구 사용보다 더 복잡한 운동 기능을 요구하는 도구의 생산이나 사용이 가능했음을 시사한다.[43]동굴 유적지인 인류의 요람 지역에서는 종종 석기와 뼈 도구가 함께 발견된다. 석기는 주로 초기 호모 속 인류에게 귀속되지만, 뼈 도구는 일반적으로 ''P. robustus''와 연관 지어진다. 이는 뼈 도구가 ''P. robustus''의 유해가 호모 속 유해보다 훨씬 많이 발견되는 지층에서 가장 풍부하게 나타나기 때문이다. 오스트랄로피테쿠스의 뼈 기술은 1950년대 다트(Dart)에 의해 처음 제안되었는데, 그는 이를 골치치석 문화라고 부르며 약 300만 년에서 260만 년 전 마카판스갓에서 발견된 ''A. africanus''에게 귀속시켰다. 그러나 현재 이 뼈들은 도구로 여겨지지 않으며, 골치치석 문화의 존재는 지지받지 못한다. 최초의 뼈 도구는 1959년 스테르크폰테인 멤버 5에서 로빈슨(Robinson)에 의해 보고되었다. 이후 1980년대 후반과 1990년대 초반, 남아프리카 고생물학자 찰스 킴벌린 브레인이 스와르트크란스에서 진행한 발굴에서 84개의 유사한 뼈 도구가 추가로 발견되었고, 키저(Keyser)가 드리몰렌에서 발굴한 결과 23개가 더 나왔다. 이 도구들은 대부분 아슐리안 석기와 함께 발견되었으나, 올도완 석기가 나온 스와르트크란스 멤버 1에서 발견된 예외도 있다. 따라서 이 지역에서 총 108개의 뼈 도구 표본이 확인되었으며, 크롬드라이 B에서도 추가로 2개가 발견되었을 가능성이 있다. 크롬드라이 B에서 발견된 두 개의 석기("발달된 올도완" 또는 "초기 아슐리안")는 해당 지층에서 호모 속 인류가 확실히 확인되지 않았기 때문에 ''P. robustus''에 귀속될 수 있다. 물론 이 석기들이 재가공된 화석일 가능성(즉, 원래 다른 곳에 있다가 나중에 해당 지층으로 옮겨왔을 가능성)도 배제할 수는 없다.
뼈 도구는 주로 식물을 자르거나 가공하고,[72] 과일(특히 마룰라 열매)을 처리하거나, 나무껍질을 벗기거나,[69] 땅속줄기나 흰개미를 파내는 데 사용되었을 것으로 추정된다.[72][69][70] ''P. robustus''의 앞니 형태가 ''H. erectus''와 현생 인류의 중간 정도로 보이는 점은, 이들이 간단한 도구를 사용하여 음식을 미리 처리했기 때문에 큰 덩어리를 직접 물어뜯을 필요성이 줄었을 수 있음을 암시한다.[71] 뼈 도구는 일반적으로 중대형 포유류의 뼈대 긴뼈로 만들어졌지만, 아래턱, 갈비뼈, 뿔 심으로 만든 도구도 발견되었다. 이 도구들은 특정 작업을 위해 의도적으로 모양을 다듬거나 제작된 것은 아니지만, 풍화 흔적이 없고 특정 종류의 뼈를 선호한 경향이 나타나는 것으로 보아, 재료를 고를 때 특별히 손으로 선택했을 가능성이 높다. 이는 사용 전에 특정 모양으로 수정하고 직접 잘라서 만든 것으로 보이는 동아프리카의 뼈 도구와는 대조적인 특징이다.[72]
1988년, 브레인과 남아프리카 고고학자 A. 쉴렌트(Sillen)는 스와르트크란스 멤버 3에서 발견된 59,488개의 뼛조각을 분석하여 그중 270개가 불에 탄 흔적이 있음을 발견했다. 불에 탄 뼈는 주로 중형 영양, 얼룩말, 멧돼지, 비비, 그리고 ''P. robustus''의 것이었다. 이 흔적들은 멤버 3 지층 전체에 걸쳐 발견되어, 불이 해당 퇴적 기간 동안 주기적으로 사용되었음을 시사한다. 색상과 구조 변화를 분석한 결과, 46개는 300°C 미만, 52개는 300°C에서 400°C 사이, 45개는 400°C에서 500°C 사이, 그리고 127개는 그보다 높은 온도로 가열된 것으로 나타났다. 연구자들은 이것이 "화석 기록상 불 사용의 가장 초기 직접적인 증거"라고 결론지으며, 이 온도들을 동굴 근처에서 흔히 자라는 화이트 스팅크우드 나무를 태운 실험적인 모닥불에서 얻어진 온도와 비교했다. 일부 뼈에서는 도살과 일치하는 절단 자국이 발견되었지만, 연구자들은 호미닌이 음식을 요리하는 대신 포식자를 쫓거나 단순히 몸을 데우기 위해 불을 피웠을 가능성도 제기했다. 동굴에서 ''P. robustus''와 ''H. ergaster''/''H. erectus''가 모두 발견되었기 때문에, 어느 종이 불을 사용했는지는 확실하지 않다.[73] 호미닌 활동의 대안으로는, 뼈가 동굴 안에서 직접 타지 않고 주기적으로 발생하는 산불에 의해 자연적으로 탄 후(건조한 사바나 환경, 동굴 내 구아노나 식물 축적물 등으로 인해 산불에 취약했을 수 있음) 멤버 3 지층으로 쓸려 들어갔을 가능성도 제기된다.[74][75] 현재 불 사용에 대한 가장 오래된 증거 주장은 2011년 남아프리카 고고학자 피터 보몬트가 남아프리카 원더워크 동굴에서 발견한 170만 년 전의 흔적으로, 그는 이를 ''H. ergaster''/''H. erectus''에 의한 것으로 보았다.[76]
4. 4. 포식자

오스트랄로피테신의 뼈가 동굴에서 발견되는 것은 대형 육식동물이 사체를 동굴로 가져오면서 쌓였기 때문일 수 있다. 이러한 가능성은 1981년 고인류학자 C. K. 브레인의 저서 ''사냥꾼인가, 사냥감인가?: 아프리카 동굴 유해론 소개''에서 처음 자세히 연구되었다. 실제로 어린 ''파란트로푸스 로부스투스''의 두개골 조각인 SK 54에는 두 개의 뚜렷한 구멍 자국이 있는데, 이는 같은 퇴적층에서 발견된 표범 표본(SK 349)의 아래 송곳니와 일치하는 것으로 보인다.
브레인은 처음에 ''디노펠리스''와 사냥 하이에나가 오스트랄로피테신을 전문적으로 사냥했을 것이라고 추정했다.[104] 하지만 탄소 동위원소 분석 결과, 이들 포식자는 주로 큰 초식동물을 먹었으며, 오히려 표범, 검치호의 일종인 ''메간테레온'', 그리고 점박이 하이에나가 ''파란트로푸스 로부스투스''를 정기적으로 잡아먹었을 가능성이 더 높은 것으로 밝혀졌다.[103] 브레인은 이 포식자들이 ''파란트로푸스 로부스투스''를 사냥하여 동굴로 끌고 갔는지, 아니면 동굴 깊은 곳에 서식하며 들어오는 개체를 매복 공격했는지에 대해서는 확신하지 못했다. 오늘날 해당 지역의 개코원숭이는 특히 추운 겨울밤에 싱크홀에서 피난처를 찾기도 하지만, 브레인은 오스트랄로피테신이 계절에 따라 고원과 더 따뜻한 부시벨트 지역 사이를 이동하며 봄과 가을에만 동굴을 피난처로 이용했을 수도 있다고 제안했다.[104]
포식 위협에 대한 방어 행동으로, 오늘날 개코원숭이가 임팔라와 같은 중간 크기 이상의 초식동물과 함께 무리 지어 다니는 것처럼, 개방된 환경에 살았던 ''파란트로푸스 로부스투스''나 다른 초기 호미닌들도 비슷한 행동을 했을 가능성이 제기된다. 이는 화석 유적지에서 중간 크기 이상의 소과 및 말과 동물 유해가 풍부하게 발견되는 점과 관련이 있을 수 있다.[105]
5. 진화
파란트로푸스 로부스투스는 플라이스토세 초기 남아프리카에 등장한 고인류의 한 종이다. 약 207만 년 전, ''A. africanus''가 사라질 무렵 나타난 것으로 보이며[94], '강건한(robust)'이라는 이름처럼 오스트랄로피테쿠스보다 더 튼튼한 턱과 큰 어금니를 가진 것이 특징이다. 이는 로버트 브룸이 1938년 처음 화석을 발견했을 때 기존에 알려진 ''A. africanus''와 뚜렷하게 구분되는 점이었다.[107]
''P. robustus''의 정확한 진화 경로는 아직 명확하지 않다. 일부 학자들은 스텔크폰테인에서 발견된 StW 505 두개골을 근거로 ''A. africanus''에서 직접 진화했을 가능성을 제기하기도 했으나, 일반적으로 이 화석은 ''A. africanus''로 분류된다.[10] ''P. robustus''의 등장은 인류의 진화가 하나의 직선적인 경로가 아니라 여러 갈래로 나뉘어 진행되었음을 보여주는 중요한 증거로 여겨진다.[107] 또한, 동아프리카의 ''P. boisei''와 유사한 특징을 보이는 큰 어금니 화석(GDA-2)의 발견은[101] ''파란트로푸스'' 속 내의 다양성 또는 종 분류에 대한 추가적인 연구 필요성을 시사한다.
''P. robustus''는 초기 호모 속(''H. erectus'' 또는 ''H. ergaster'') 및 ''A. sediba''와 같은 다른 호미닌들과 동시대, 동일 지역에서 공존했다.[55][74][94] 이들 간의 상호작용과 경쟁 관계는 당시 생태 환경을 이해하는 데 중요한 연구 주제이다.[95] (자세한 내용은 #다른 호미닌과의 관계 참조)
이후 ''P. robustus''는 중기 플라이스토세 전이 시기의 급격한 기후 변화와 서식지 축소, 그리고 다른 호미닌과의 경쟁 등으로 인해 약 100만 년 전에서 60만 년 전 사이에 멸종한 것으로 추정된다.[106] (자세한 내용은 #멸종 참조)
5. 1. 다른 호미닌과의 관계
''P. robustus''는 플라이스토세 초기 남아프리카의 인류의 요람 지역에서 동시대의 다른 호미닌들과 공존했다. 특히 약 207만 년 전 ''A. africanus''가 사라질 무렵 등장하여[94], 초기 호모 에렉투스(또는 ''H. ergaster'')와 같은 지역에서 함께 서식한 증거가 발견되었다.[55][74][94] 또한, 약 200만 년 전 말라파에서는 ''A. sediba''와도 공존했던 것으로 보인다.[94]스와르트크란스, 드림롤렌 동굴 등 여러 유적지에서 ''P. robustus''와 초기 호모 속 화석이 함께 발견되었다. 예를 들어 스와르트크란스 동굴 유적의 멤버 1~3 지층에서는 ''P. robustus''와 초기 ''호모'' 속 화석이 같이 출토되었는데, 이 초기 ''호모'' 화석이 정확히 어떤 종(''H. erectus'', ''H. habilis'', ''H. rudolfensis'' 등)에 해당하는지에 대해서는 학자들 간의 논쟁이 계속되고 있다.[97][96][10] 드림롤렌 동굴에서도 ''P. robustus'' 화석과 함께 ''H. ergaster''/''H. erectus''로 추정되는 두개골(DNH 134)이 발견되어[94] 두 종의 공존 사실을 뒷받침한다.
이처럼 여러 호미닌 종들이 같은 시기, 같은 장소에 살았다는 사실은 이들 사이의 상호작용(공생, 중립, 경쟁 등)이 어떠했는지에 대한 궁금증을 낳았고, 이는 현재까지도 활발히 연구되는 주제이다.[95] 약 200만 년 전 남아프리카는 주요 기후 변동을 겪기 시작했고, ''호모'' 속과 ''파란트로푸스'' 속이라는 새로운 경쟁자들이 등장하면서 기존의 ''오스트랄로피테쿠스''에게는 마지막 서식지인 피난처가 되었을 가능성이 있다.[94]
''P. robustus''와 ''A. africanus''의 직접적인 진화적 관계 또한 명확히 밝혀지지 않았다. 고인류학자 로널드 J. 클라크는 스텔크폰테인 멤버 4 지층에서 발견된 StW 505 두개골이 ''A. africanus''에서 ''P. robustus''로 이어지는 과도기적 형태일 수 있다고 주장했지만, 이 화석은 일반적으로 ''A. africanus''로 분류된다.[10]
로버트 브룸이 1938년 ''P. robustus''를 처음 발견했을 때, 이는 레이먼드 다트가 발견했던 ''A. africanus''와는 확연히 다른, 훨씬 튼튼한(robust) 형태의 고인류였다. 이 발견은 인류의 진화가 하나의 직선적인 경로를 따른 것이 아니라, 여러 갈래로 나뉘어 다양하게 발전해왔음을 보여주는 중요한 증거가 되었다.[107] 또한, 곤돌린 동굴에서 발견된 GDA-2 어금니 화석은 크기가 매우 커서 일반적인 ''P. robustus''의 범위를 벗어나 동아프리카의 ''P. boisei''와 유사한 특징을 보이는데, 이는 ''파란트로푸스'' 속 내에서도 상당한 다양성이 존재했거나 종 분류에 대한 추가 연구가 필요함을 시사한다.[101]
5. 2. 멸종
''파란트로푸스 로부스투스''는 환경 변화에 대한 내성이 있는 강인한 종이었지만, 주로 숲이 우거진 환경을 선호했던 것으로 보인다. 실제로 대부분의 ''파란트로푸스 로부스투스'' 유해는 이러한 환경이 우세했던 200만 년 전에서 175만 년 전 사이 남아프리카의 습윤기에 해당하는 지층에서 발견된다.[106]''파란트로푸스 로부스투스''의 멸종은 중기 플라이스토세 전이와 빙하기 주기가 두 배로 길어진 시기와 일치한다. 빙하기 동안 극지방에 더 많은 얼음이 갇히면서 열대 우림대가 적도 쪽으로 축소되었고, 이로 인해 ''파란트로푸스 로부스투스''가 선호했던 습지와 숲 환경이 줄어들었다. 중기 플라이스토세 전이 이전에는 21,000년 주기로 기후가 변하면서, ''파란트로푸스 로부스투스'' 개체군이 특정 숲 피난처로 축소되었다가 습윤 주기가 돌아오면 다시 확장하는 과정을 반복했을 수 있다. 그러나 점차 건조한 주기가 길어지고 강도가 심해지면서, 이들은 약 100만 년 전에서 60만 년 전 사이(더 정확하게는 90만 년 전 무렵)에 화석 기록에서 마지막으로 나타나며 멸종에 이르렀을 것으로 추정된다.[106]
멸종의 또 다른 요인으로는 동시대에 살았던 다른 고인류와의 경쟁 가능성이 제기된다. ''파란트로푸스 로부스투스''는 인류의 요람 지역에서 ''호모 에르가스터'' 또는 ''호모 에렉투스''와 공존했으며,[55][74][94] 약 200만 년 전에는 말라파에서 ''오스트랄로피테쿠스 세디바''와 함께 살았다.[94] 또한, 가장 마지막까지 생존했던 ''오스트랄로피테쿠스 아프리카누스'' (Sts 5 개체)는 약 207만 년 전까지 존재했는데, 이는 ''파란트로푸스 로부스투스''와 ''호모 에렉투스''가 등장하는 시기와 겹친다.[94] 이들 사이의 관계가 공생적이었는지, 중립적이었는지, 아니면 경쟁적이었는지에 대해서는 아직 논쟁이 있다.[95] 일부 학자들은 남아프리카가 약 200만 년 전까지 ''오스트랄로피테쿠스''의 마지막 피난처였을 수 있으며, 이후 주요 기후 변동과 더불어 ''호모'' 및 ''파란트로푸스''와의 경쟁이 시작되었을 가능성을 제시한다.[94] 상대적으로 ''호모'' 속은 ''파란트로푸스 로부스투스''보다 훨씬 넓은 지리적 범위에 분포했기 때문에, 불리한 기후 변화 속에서도 적합한 피난처를 찾아 생존할 가능성이 더 높았다.[106]
그러나 현재까지 발견된 ''파란트로푸스 로부스투스''의 화석 기록은 지리적으로 약 500km2 범위에 한정되어 있다. 이는 현존하는 유인원 중 서식 범위가 가장 좁은 멸종위기종인 동부 고릴라 (70000km2)나 서부 고릴라 (700000km2), 또는 취약종인 침팬지 (2.6e6km2)의 서식 범위와 비교하면 매우 좁은 면적이다. 따라서 현재의 화석 분포가 이 종의 실제 서식 범위를 온전히 반영한다고 보기는 어렵다. 이러한 화석 기록의 한계(시그너-립스 효과) 때문에, ''파란트로푸스 로부스투스''가 주요 화석 발견지인 인류의 요람 지역 외의 다른 곳에서는 훨씬 더 최근까지 생존했을 가능성도 배제할 수 없다.[106]
참조
[1]
웹사이트
Hominin Taxic Diversity
https://oxfordre.com[...]
2020-05-29
[2]
간행물
The TM 1517 odontoskeletal assemblage from Kromdraai B, South Africa, and the maturational pattern of ''Paranthropus robustus''
https://kar.kent.ac.[...]
[3]
논문
The Pleistocene Anthropoid Apes of South Africa
[4]
웹사이트
"''Paranthropus''"
https://www.merriam-[...]
Merriam–Webster Dictionary
2020-06-20
[5]
논문
Ape-Like ''Australopithecus'' After Seventy Years: Was It a Hominid?
[6]
논문
Another new type of fossil ape-man
[7]
논문
The genera and species of the South African fossil ape-men
[8]
논문
A Systematic Assessment of Early African Hominids
[9]
논문
Evolutionary Importance of the South African 'Man-apes'
[10]
서적
Miscelanea en Homenaje a Emiliano Aguirre
https://mds.marshall[...]
Museo Arqueológico Regional
[11]
논문
The genera and species of the australopithecinae
[12]
논문
Evidence for increased hominid diversity in the Early to Middle Pleistocene of Indonesia
https://kar.kent.ac.[...]
[13]
논문
The Nature of ''Telanthropus capensis''
[14]
논문
New Discoveries in Tanganyika: Their Bearing on Hominid Evolution
[15]
논문
The dentition of the Transvaal Pleistocene anthropoids, ''Plesianthropus'' and ''Paranthropus''
https://journals.co.[...]
[16]
논문
"''Gigantopithecus blacki'': a giant ape from the Pleistocene of Asia revisited"
[17]
논문
The skull of StW 573, a 3.67 Ma ''Australopithecus prometheus'' skeleton from Sterkfontein Caves, South Africa
[18]
논문
Hominin Taxonomy and Phylogeny: What's In A Name?
https://www.nature.c[...]
[19]
논문
2.5-Myr Australopithecus boisei from west of Lake Turkana, Kenya
[20]
논문
"''Paranthropus boisei'': Fifty years of evidence and analysis"
https://mds.marshall[...]
[21]
논문
Oldest genetic data from a human relative found in 2-million-year-old teeth
https://www.nature.c[...]
2023-07-10
[22]
논문
Three-dimensional molar enamel distribution and thickness in ''Australopithecus'' and ''Paranthropus''
[23]
서적
The Human Lineage
https://books.google[...]
John Wiley and Sons
[24]
논문
Drimolen cranium DNH 155 documents microevolution in an early hominin species
[25]
논문
The bony labyrinth of StW 573 ("Little Foot"): Implications for early hominin evolution and paleobiology
[26]
논문
Reconstructed ''Homo habilis'' type OH 7 suggests deep-rooted species diversity in early ''Homo''
[27]
논문
Observations on the anatomy of the fossil Australopithecinae
[28]
논문
A new graphic reconstruction of the type specimen of ''Australopithecus robustus'' from Kromdraai, South Africa - TM 1517
https://www.research[...]
[29]
논문
New australopithecine endocast, SK 1585, from Swartkrans, South Africa
[30]
논문
Early hominid brain evolution: a new look at old endocasts
https://pdfs.semanti[...]
[31]
논문
Les veines méningées moyennes des Australopithèques
https://www.persee.f[...]
[32]
논문
New ''Australopithecus boisei'' calvaria from East Lake Turkana, Kenya
[33]
논문
The cranial venous sinus system in ''Australopithecus afarensis''
[34]
서적
Evolutionary History of the "Robust" Australopithecines
Routledge
[35]
논문
3D shape analyses of extant primate and fossil hominin vertebrae support the ancestral shape hypothesis for intervertebral disc herniation
[36]
논문
Comparative morphometric study of the australopithecine vertebral series Stw-H8/H41
http://pdfs.semantic[...]
[37]
논문
A fragmentary pelvis of ''Paranthropus robustus'' of the Plio-Pleistocene site of Drimolen Republic of South Africa
[38]
논문
Hip bone trabecular architecture shows uniquely distinctive locomotor behaviour in South African australopithecines
1999-02
[39]
논문
The humerus of ''Paranthropus robustus''
[40]
논문
Radius of ''Paranthropus robustus'' from member 1, Swartkrans Formation, South Africa
[41]
논문
New hominid fossils from the Swartkrans formation (1979–1986 excavations): Postcranial specimens
[42]
논문
First Partial Skeleton of a 1.34-Million-Year-Old ''Paranthropus boisei'' from Bed II, Olduvai Gorge, Tanzania
[43]
논문
Hand of ''Paranthropus robustus'' From Member 1, Swartkrans: Fossil Evidence for Tool Behavior
[44]
논문
Human evolution: taxonomy and paleobiology
[45]
논문
Human-like hip joint loading in ''Australopithecus africanus'' and ''Paranthropus robustus''
https://www.reposito[...]
[46]
논문
Evidence for habitual climbing in a Pleistocene hominin in South Africa
[47]
논문
The SKX 1084 hominin patella from Swartkrans Member 2, South Africa: An integrated analysis of its outer morphology and inner structure
[48]
논문
New first metatarsal (SKX 5017) from Swartkrans and the gait of ''Paranthropus robustus''
[49]
논문
Reviewed Work: Early Hominid Posture and Locomotion by John T. Robinson
[50]
논문
Size and Scaling in Human Evolution
[51]
서적
Evolutionary History of the "Robust" Australopithecines
Routledge
[52]
논문
Petite bodies of the "robust" australopithecines
[53]
논문
Stretching the time span of hominin evolution at Kromdraai (Gauteng, South Africa): Recent discoveries
[54]
논문
Femoral lengths and stature in Plio-Pleistocene hominids
[55]
논문
Recently identified postcranial remains of ''Paranthropus'' and Early ''Homo'' from Swartkrans Cave, South Africa
[56]
논문
Body Size, Body Shape, and the Circumscription of the Genus ''Homo''
[57]
논문
Body mass estimates of hominin fossils and the evolution of human body size
[58]
논문
Diet and teeth. Dietary hypotheses and human evolution
[59]
논문
Patterns of resource use in early ''Homo'' and ''Paranthropus''
[60]
논문
The rise of the hominids as an adaptive shift in fallback foods: Plant underground storage organs (USOs) and australopith origins
[61]
논문
Bears, pigs, and Plio-Pleistocene hominids: A case for the exploitation of belowground food resources
[62]
논문
The hunters and the hunted revisited
[63]
논문
Isotopic Evidence for Dietary Variability in the Early Hominin ''Paranthropus robustus''
[64]
논문
Mechanisms of Tooth Damage in ''Paranthropus'' Dietary Reconstruction
[65]
논문
Extended male growth in a fossil hominin species
http://doc.rero.ch/r[...]
[66]
논문
Strontium isotope evidence for landscape use by early hominins
[67]
논문
''Australopithecus robustus'' societies - one-male or multimale?
[68]
논문
Sagittal crest formation in great apes and gibbons
[69]
논문
Assessing the function of early hominin bone tools
[70]
논문
Evidence of termite foraging by Swartkrans early hominids
[71]
논문
Dietary proclivities of ''Paranthropus robustus'' from Swartkrans, South Africa
[72]
논문
The first bone tools from Kromdraai and stone tools from Drimolen, and the place of bone tools in the South African Earlier Stone Age
[73]
논문
Evidence from the Swartkrans cave for the earliest use of fire
[74]
논문
What's new is old: comments on (more) archaeological evidence of one-million-year-old fire from South Africa
https://www.research[...]
[75]
논문
Earliest fire in Africa: towards the convergence of archaeological evidence and the cooking hypothesis
[76]
논문
The Edge: More on Fire-Making by about 1.7 Million Years Ago at Wonderwerk Cave in South Africa
[77]
논문
Life-History Inference in the Early Hominins ''Australopithecus'' and ''Paranthropus''
[78]
논문
Mandibular development in ''Australopithecus robustus''
[79]
논문
The eruption pattern of the permanent incisors and first permanent molars in ''Australopithecus (Paranthropus) robustus''
[80]
PhD
The Paleodemography of Australopithecus
University of California, Berkeley
[81]
논문
Survivorship in gracile and robust ''Australopithecines'': A demographic comparison and a proposed birth model
[82]
논문
Newborn Size and Pelvic Dimensions of ''Australopithecus''
[83]
논문
Gestation Period for ''Australopithecus''
[84]
논문
Gestation Period and Birth Weight of ''Australopithecus''
[85]
논문
Behavioral ecological implications of early hominid body size
[86]
논문
Tertiary Dentine Frequencies in Extant Great Apes and Fossil Hominins
[87]
논문
A probable genetic origin for pitting enamel hypoplasia on the molars of ''Paranthropus robustus''
http://researchonlin[...]
[88]
논문
Dental caries in human evolution: frequency of carious lesions in South African fossil hominins
[89]
논문
Root caries on a ''Paranthropus robustus'' third molar from Drimolen
https://www.biorxiv.[...]
[90]
논문
The Hard Evidence of Alveolar Bone Loss in Early Hominids of Southern Africa
[91]
논문
An Attempt to Reconstruct the Behaviour of Australopithecines: The Evidence for Interpersonal Violence
[92]
논문
Macromammalian faunas, biochronology and palaeoecology of the early Pleistocene Main Quarry hominin-bearing deposits of the Drimolen Palaeocave System, South Africa
[93]
논문
Bipedality and hair loss in human evolution revisited: The impact of altitude and activity scheduling
[94]
논문
Contemporaneity of ''Australopithecus'', ''Paranthropus'', and early ''Homo erectus'' in South Africa
[95]
논문
Paleoecology of Early Hominidae in Africa
https://pdfs.semanti[...]
[96]
논문
Early ''Homo'' at Swartkrans, South Africa : a review of the evidence and an evaluation of recently proposed morphs
[97]
논문
A multi-disciplinary seriation of early ''Homo'' and ''Paranthropus'' bearing palaeocaves in southern Africa
[98]
논문
Comogenic nuclide burial dating of hominin-bearing Pleistocene cave deposits at Swartkrans, South Africa
[99]
논문
Stratigraphy, artefact industries and hominid associations for Sterkfontein, Member 5
[100]
논문
A new partial temporal bone of a juvenile hominin from the site of Kromdraai B (South Africa)
[101]
논문
First record of hominid teeth from the Plio-Pleistocene site of Gondolin, South Africa
[102]
논문
The Drimolen skull: the most complete australopithecine cranium and mandible to date
https://www.research[...]
[103]
논문
The hunters and the hunted revisited
[104]
서적
The Hunters Or the Hunted?: An Introduction to African Cave Taphonomy
https://books.google[...]
University of Chicago Press
[105]
논문
Possible predator avoidance behaviour of hominins in South Africa
[106]
논문
A two-million-year-long hydroclimatic context for hominin evolution in southeastern Africa
http://dro.dur.ac.uk[...]
[107]
웹사이트
Paranthropus robustus
http://humanorigins.[...]
Smithsonian Institution
2015-12-19
[108]
논문
The Drimolen skull: the most complete australopithecine cranium and mandible to date
http://servicios.edu[...]
2015-12-19
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com