대뇌 피질
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
대뇌 피질은 뇌의 가장 바깥쪽 층으로, 주름진 구조를 가지며, 인지, 감각, 운동 기능을 담당하는 부위이다. 해부학적으로 이랑과 고랑으로 구분되며, 전두엽, 두정엽, 측두엽, 후두엽, 섬엽, 변연엽으로 나뉜다. 대뇌 피질은 6개의 층으로 이루어진 신피질과 3~4개의 층을 가진 구피질로 구성되며, 각 층은 특정한 신경세포와 연결을 가진다. 대뇌 피질의 발달은 태아기부터 시작되며, 유전자와 환경의 상호작용에 의해 영향을 받는다. 뇌졸중, 뇌전증, 알츠하이머병, 발달 장애 등 다양한 질환과 관련이 있으며, 뇌 기능 매핑 및 유전자 변이 연구를 통해 질병 진단 및 치료에 활용된다.
더 읽어볼만한 페이지
- 뇌과학 - 공감
공감은 타인의 감정을 이해하고 공유하는 심리적 과정으로, 정서적 공감(감정적 반응)과 인지적 공감(관점 이해)으로 나뉘며, 유전적, 환경적 요인과 뇌의 거울 뉴런 시스템에 영향을 받고, 사회적 상호작용, 도덕적 판단, 정신 질환과 관련되며, 다양한 방법으로 측정되고, 동물 연구 및 허구 매체에서도 중요한 요소로 다뤄진다. - 뇌과학 - 시냅스
시냅스는 신경세포 간 신호 전달을 담당하는 구조로, 화학적 시냅스와 전기적 시냅스로 구분되며 기억, 학습 등 뇌 기능에 중요한 역할을 한다. - 대뇌피질 - 피라미드 세포
피라미드 세포는 뇌의 뉴런으로, 원뿔 모양의 세포체, 단일 축삭, 큰 정단 수상돌기, 여러 개의 기저 수상돌기, 수상돌기 가시를 가지며, 대뇌 피질과 해마에서 발견되어 인지, 기억, 학습과 같은 기능을 수행하고, 발화 특성 및 유전자 발현에 따라 다양한 하위 유형으로 분류된다. - 대뇌피질 - 전전두피질
전전두피질은 전두엽의 가장 앞쪽에 위치한 뇌 영역으로, 외측과 내측으로 나뉘며 단기 기억, 언어, 집행 기능 등 다양한 인지 기능을 수행하고 정신 질환 및 행동 변화와 관련된다. - 대뇌 - 바깥섬유막
바깥섬유막은 그람 음성 세균 세포벽의 주요 구성 성분으로, 인지질 이중층과 리포다당류로 이루어져 세균 생존과 병원성에 중요한 역할을 하며, 특히 리포다당류는 외막 안정화와 숙주 면역 반응 유발에 기여한다. - 대뇌 - 기저핵
기저핵은 대뇌 심부에 위치한 회백질 덩어리 집합으로, 선조체, 창백핵, 흑질, 시상하핵의 네 가지 주요 구조로 나뉘어 운동 조절, 인지 기능, 보상 학습 등 다양한 기능에 관여하며 파킨슨병, 헌팅턴병과 같은 신경 퇴행성 질환 및 정신 질환에도 영향을 미친다.
![대뇌 피질, 측면 보기. [[Sulcus (neuroanatomy)|고랑]]과 [[Gyrus|이랑]] (주름과 융기)](https://cdn.onul.works/wiki/noimage.png)
2. 해부학적 구조
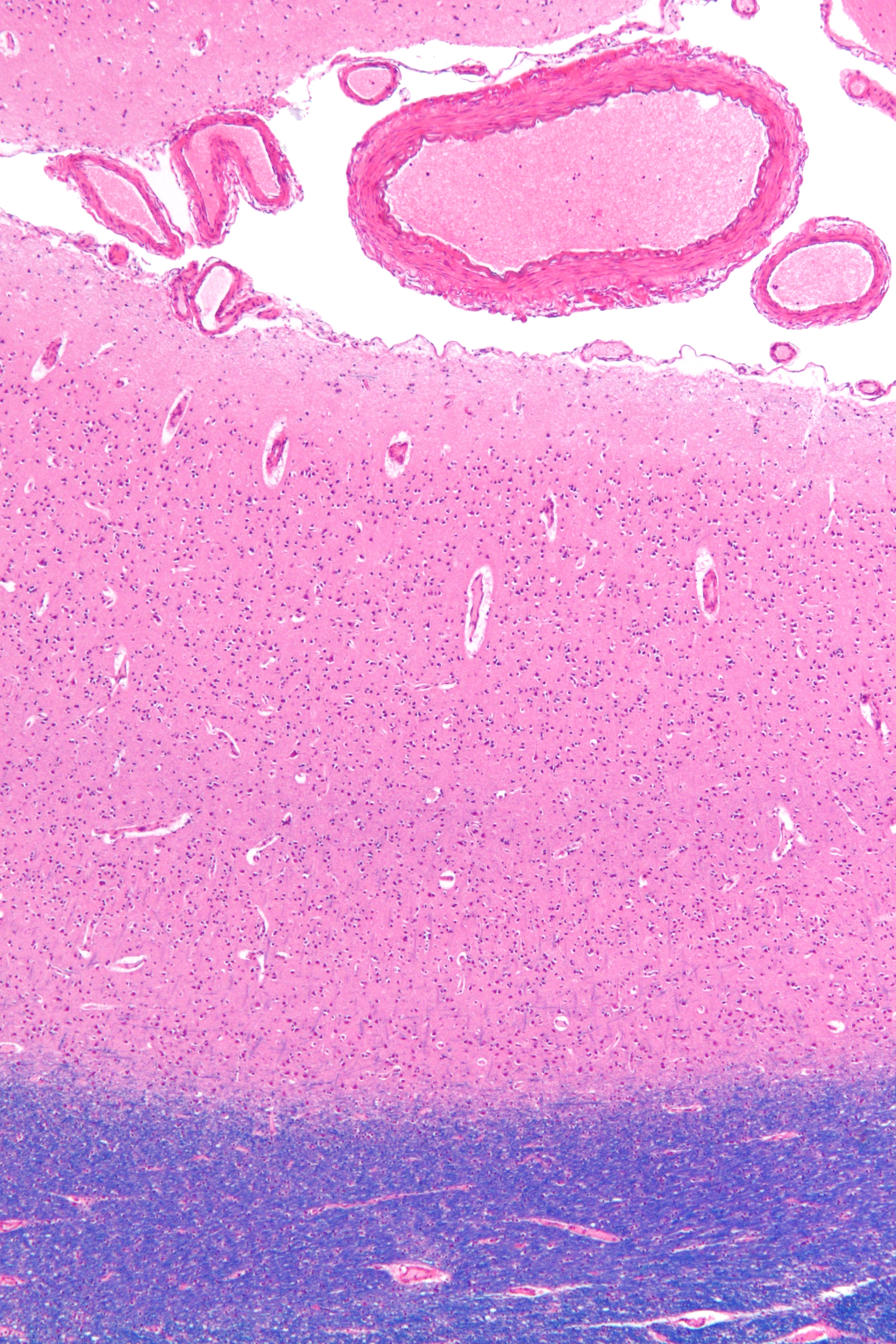
대뇌 피질은 뇌의 표면을 덮고 있는 신경 세포의 집합으로, 주름진 형태를 통해 표면적을 넓혀 더 많은 신경 세포를 수용한다. 대뇌 피질은 대뇌 반구 표면의 바깥 덮개이며, 이랑이라고 불리는 봉우리와 고랑이라고 불리는 홈으로 접혀 있다. 사람의 뇌에서 두께는 2mm~4mm이며,[6] 뇌 질량의 40%를 차지한다.[7] 대뇌 피질의 90%는 6층의 신피질이며, 나머지 10%는 3~4층의 구피질로 구성되어 있다.[7] 피질에는 140억~160억 개의 뉴런이 있다.[7]
대뇌 피질의 각 부분에는 여러 가지 명칭이 혼재되어 사용되고 있다.
첫째, 뇌구 등 육안으로 확인할 수 있는 구조를 기준으로 부여된 거시 해부학적인 명칭이 있다. (뇌엽, 뇌회, 뇌구 등 참조).
둘째, 현미경이나 염색 기술 등을 사용하여 확인되는 미세적인 구조를 기준으로 부여된 미세 해부학적·세포 구축학적인 명칭이 있다. (브로드만의 뇌 지도 등 참조).
셋째, 각 부위가 수행하는 기능적 역할에 따라 부여된 명칭이 있다. (일차 운동 피질, 체감각 피질, 시각 피질, 청각 피질, 언어 영역 등 참조).
그리고 이러한 명칭에 위, 아래, 앞, 뒤, 내측, 외측과 같은 방향을 나타내는 용어를 부가하여 위치를 더욱 세밀하게 지정한다.
또한 fMRI 등의 뇌 영상 기술을 사용한 연구에서는 데이터를 주고받기 위해 타일라흐 좌표 (Talairach coordinates)나 MNI 좌표 등의 삼차원 뇌 좌표계를 사용하여 x, y, z의 세 쌍의 수치로 피질 부위를 지정한다.
예를 들어 외측 슬상체로부터 시방선을 통해 시각 정보의 입력을 받는 대뇌 피질의 부위는, 거시 해부학적 분류에서는 후두엽 내측면의 일부, 또는 조두구 주변이며, 세포 구축학적 분류에서는 브로드만 영역 17영역(BA17)이며, 기능적인 분류라면 일차 시각 피질 (V1)이며, 타일라흐 좌표라면 (-3, -74, 7) [87]의 주변과 같은 점이다.
2. 1. 대뇌 피질의 주름
사람의 대뇌피질은 표면적이 매우 넓어 주름져 있으며, 바깥쪽으로 튀어나온 이랑과 안쪽으로 움푹 파인 고랑이 존재한다.[95][96] 이랑과 고랑은 대뇌피질 영역을 구분하는 경계가 되기도 한다.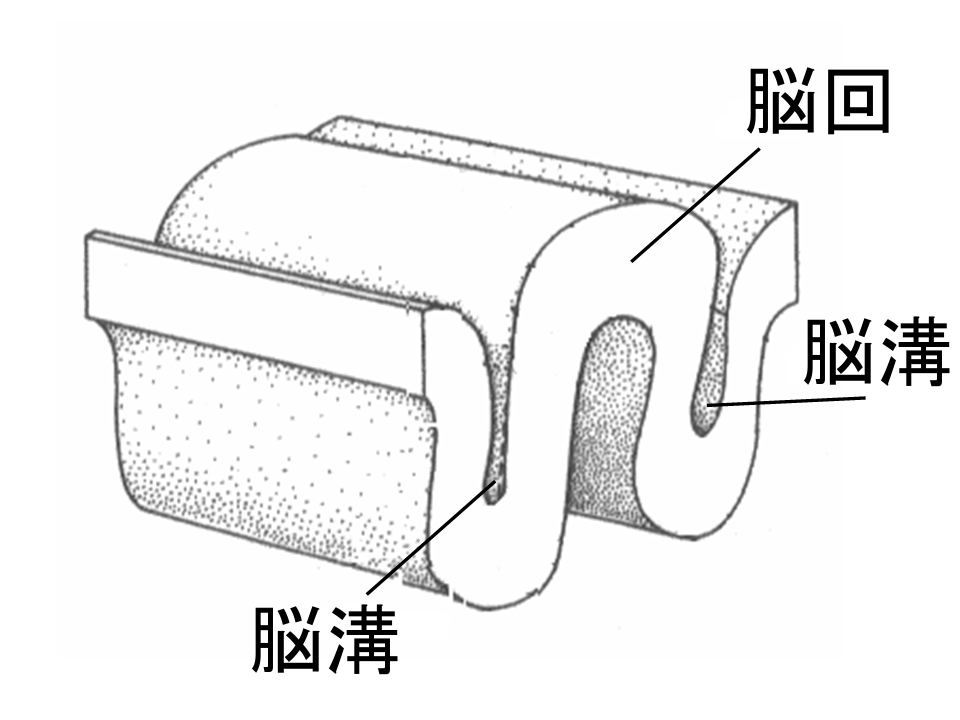
대뇌 표면에서 영역을 구분하는 중요한 지표는 다음과 같다.
- 중심고랑(Central sulcus): 대뇌 반구의 가쪽 면에서 이마엽과 마루엽의 경계가 되는 고랑이다. 앞쪽의 운동세포 쪽에 존재하며, 뒤쪽 부분의 감각 정보를 받는다. 깊이는 1cm 정도이다. 중심고랑을 기준으로 앞쪽은 운동 영역, 뒤쪽은 몸 감각 영역이 된다.
- 가쪽고랑(Lateral sulcus): 대뇌반구의 아랫쪽 표면과 옆쪽 표면을 구분한다. 전두엽(이마엽)과 측두엽(관자엽)을 구분한다.
- 마루뒤통수고랑(Parieto-occipital sulcus): 반구 위쪽 중앙 끝부분에서 시작하여 후두엽(뒤통수엽)까지 이어진다. 중앙 표면 아래로 내려와 새발톱고랑과 만난다. 두정엽(마루엽)과 후두엽을 구분한다.
- 새발톱고랑(Calcarine sulcus): 반구 중앙 표면에 있으며 뇌들보 뒤쪽 끝부분 아래에서 시작하여 뒤통수극에 이른다.
- 둘레섬고랑(circular sulcus of insula): 가쪽고랑을 벌렸을 때 관찰되는 고랑으로, 섬엽(insular lobe)을 거의 둘러싸고 있다. 섬엽을 나머지 전두엽·두정엽·측두엽과 구분한다.
가쪽고랑과 마루뒤통수고랑은 태아기 초기에 형성되며, 성인의 대뇌 고랑 중 가장 깊게 패여 있다.[95][96]

대뇌는 표면에서 보면 다수의 주름이 있는 것이 특징이다. 이 주름에 의한 함몰부를 고랑, 융기부를 이랑이라고 부른다. 특히 바깥쪽으로 가로로 뻗어 있는 외측구(''실비우스 열구'')와 세로로 뻗어 있는 중심구는 눈에 잘 띄며, 해부학적 구분의 지표로서 중요하다.
2. 2. 뇌엽
사람의 대뇌피질은 표면적이 매우 넓어 주름져 있으며, 솟아오른 이랑(gyrus)과 움푹 들어간 고랑(sulcus)을 경계로 영역을 구분하기도 한다. 특히 바깥쪽으로 가로로 뻗어 있는 '''외측구'''('''실비우스 열구''')와 세로로 뻗어 있는 '''중심구'''는 눈에 잘 띄며, 해부학적 구분의 지표로서 중요하다.[95][96]
주요 뇌엽에는 전두엽, 두정엽, 측두엽, 후두엽의 네 가지가 있다. 섬엽과 변연엽도 종종 포함된다.
대뇌 피질은 다음과 같은 영역으로 나눌 수 있다.
- '''전두엽''': 외측구보다 위쪽, 중심구보다 앞쪽 부분이다. 기억력, 사고력 등의 고등 행동, 추리, 계획, 운동, 감정, 문제 해결에 관여한다.
- '''두정엽''': 외측구보다 위쪽, 중심구보다 뒤쪽 부분으로, 두정후두구부터 각회 부근보다 앞쪽 부분이다. 촉각, 압각, 통증 등의 체감각 처리에 관여한다.
- '''측두엽''': 외측구보다 아래쪽, 각회 부근보다 앞쪽 부분이다. 청각 정보 처리와 감정, 사실적 기억, 시각적 기억 정보를 처리한다.
- '''후두엽''': 두정후두구 부근보다 뒤쪽 부분이다. 시각 정보 처리를 담당한다.
- '''섬엽''': 외측구가 안쪽으로 함몰되어 넓어져서 밖에서는 보이지 않는 부분이다.
- '''변연 피질''': 대상회, 해마곁이랑, 해마 등을 포함한다. 해마는 신피질은 아니지만, 대뇌 피질에 포함된다.
포유류의 경우, 더 큰 뇌는 더 두꺼운 피질을 갖는 경향이 있다.[14] 땃쥐와 같은 가장 작은 포유류는 대뇌 피질 두께가 약 0.5mm이며, 인간과 흰수염고래와 같이 가장 큰 뇌를 가진 동물은 2mm~4mm의 두께를 갖는다.[7][6]
2. 2. 1. 전두엽 (Frontal lobe)
대뇌반구의 앞부분에 있는 전두엽은 전전두엽 관련 영역을 가지고 있어 기억력, 사고력 등 고등행동을 관장하며, 다른 연합영역으로부터 들어오는 정보를 조정하고 행동을 조절한다. 또한 추리, 계획, 운동, 감정, 문제 해결에도 관여한다. 전두엽의 피질은 중앙전두피질, 안와전두피질, 후하전두피질, 등측면 전두피질로 세분화할 수 있다.[95][96]- 중앙전두피질 (혹은 중앙전전두영역): 각성과 자극 및 동기 부여에 중요한 역할을 한다. 이 부위에 병변이 생기면 주의력 결핍, 무관심, 반응이 느려지는 등의 무의지증이 나타난다.
- 안와전두피질 (혹은 안와전전두영역): 사회적 행동 조절을 돕는다. 안와전두피질에 병변이 생긴 환자는 정서적으로 불안정해지며, 나이가 들면 치매나 비이상적인 행동을 보이기도 한다. 이러한 변화는 전두엽, 특히 안와전두피질의 퇴화로 인해 나타날 수 있다.
- 좌측후하전두피질 (Left posteroinferior frontal cortex, 후하측전전두영역): 언어 표현 기능을 조절한다. 이 부분에 병변이 생기면 의사 표현에 문제가 발생한다.
- 등측면 전두피질 (Dorsolateral frontal cortex): 작업 기억과 같이 가장 최근에 획득한 정보를 다룬다. 이 영역에 병변이 생기면 정보를 기억하는 능력이 손상된다.
2. 2. 2. 두정엽 (Parietal lobe)
두정엽은 중심고랑과 마루뒤통수고랑 사이, 바깥쪽 틈새 상부에 있으며, 체감각 피질과 감각연합영역을 포함하여 촉각, 압각, 통증 등 체감각 처리에 관여한다. 피부, 근골격계, 내장, 미뢰로부터의 감각 신호를 담당하며, 기관에 운동 명령을 내리는 운동 중추가 있다.[95][96]- 앞쪽 두정엽: 뒤쪽 부분은 일차체성신경피질로, 체감각 자극을 인지하고 형태, 무게 등을 기억한다. 일차체성감각피질은 모든 체감각 자극을 받는다. 앞쪽 두정엽 병소는 촉각을 통한 사물 인지에 어려움을 유발할 수 있다.
- 중심후회(Postrolandic area): 후측면은 시각과 공간의 관계를 형성하고, 다른 감각을 통해 인지한 정보와 통합하여 움직이는 사물의 존재를 형성한다. 이 영역은 고유 수용성 감각(Propiroception)을 매개한다.
- 중앙두정엽(Midparietal lobe): 계산, 글쓰기, 손가락 인지 등의 능력과 관련 있다. 각회(Angular gyrus) 병소는 이러한 능력에 결함을 유발할 수 있다.
체감각 피질은 편두통 환자에게서 더 두껍다는 연구 결과가 있었으나,[17][18] 더 많은 환자 집단을 대상으로 한 후속 연구에서는 편두통 환자의 피질 두께에 변화가 없다고 보고되었다.[19]
2. 2. 3. 측두엽 (Temporal lobe)
대뇌반구의 양쪽 가에 있는 부분으로 청각연합영역과 청각피질이 있어 청각 정보 처리와 감정, 사실적 기억, 시각적 기억 정보를 처리한다. 오른쪽 측두엽이 손상된 환자는 비언어적 청각 자극(예: 음악)에 대한 해석 능력을 상실한다. 왼쪽 측두엽이 손상되면 언어의 형성과 기억, 인지 능력에 크게 방해받는다.[95][96]
2. 2. 4. 후두엽 (Occipital lobe)
마루뒤통수고랑에 의해 두정엽(마루엽)과 후두엽(뒤통수엽)이 서로 구분된다.[95][96] 후두엽은 바깥쪽 표면에서 두정후두고랑 위쪽 끝부분과 후두전패임을 잇는 가상적인 선의 뒤쪽 부분이고, 안쪽 표면에서는 두정후두고랑의 뒤쪽 부분이다. 시각연합영역과 일차시각피질이라고 하는 시각중추가 있어 시각 정보 처리를 담당한다. 눈으로 들어온 시각 정보가 시각 피질에 도착하면 사물의 위치, 모양, 운동 상태를 분석한다. 여기에 장애가 생기면 눈의 다른 부위에 이상이 없더라도 볼 수 없게 된다. 후두엽에서 발작이 일어나면 환시가 유발될 수 있다.]]
2. 2. 5. 섬엽 (Insular lobe)
가쪽고랑을 벌렸을 때 관찰되는 부분으로, 둘레섬고랑을 통해 주변 전두엽(이마엽), 두정엽(마루엽), 측두엽(관자엽)과 구분된다. 섬엽을 덮고 있는 주변 피질을 덮개(operculum)라고 부르며, 다음과 같이 구분할 수 있다.[96]- 이마덮개(frontal operculum): 전두엽 아래이마이랑 중 가쪽고랑의 앞가지와 오름가지 사이에 위치하는 세모부분에 해당한다.
- 이마마루덮개(frontoparietal operculum): 전두엽 아래이마이랑 중 가쪽고랑의 오름가지 뒤쪽에 위치하는 덮개부분, 중심앞이랑의 아래쪽 부분, 두정엽 중심뒤이랑의 아래쪽 부분, 그리고 아래마루소엽의 아래 앞쪽 부분을 포함한다.
- 관자덮개(temporal operculum): 가쪽고랑 아래쪽에 위치하는 위관자이랑 및 가로관자이랑에 해당한다.
2. 2. 6. 변연엽 (Limbic lobe)
대상회, 해마곁이랑, 해마 등을 포함한다.[88][89] 해마는 신피질은 아니지만, 대뇌 피질에 포함된다고 할 수 있다. 또한, "대뇌변연계"에는 대뇌 피질 이외의 부분도 포함된다.2. 3. 브로드만 영역 (Brodmann areas)
브로드만 영역은 독일의 해부학자인 Korbinian Brodmann이 세포 구조에 따라 대뇌 피질을 52개의 영역으로 구분한 것이다.[97] 브로드만은 신피질의 6층 구조를 자세히 관찰하여 피질 위치에 따라 일부 층이 두꺼워지거나 얇아지는 등 세포 구축이 균일하지 않다는 것을 발견했다. 그는 세포 구축의 차이를 바탕으로 대뇌 피질을 52개 영역(또는 야)으로 나누었으며, 이는 뇌 기능 국재론에서 위치를 나타내는 기준으로 자주 사용된다. 예를 들어, 후두부에 위치한 신피질의 17야에서는 외측 슬상체로부터 축삭을 받는 제4층이 두껍게 발달해 있다. 외측슬상체는 망막으로부터 입력을 받고, 17야는 시각 정보를 처리한다고 볼 수 있는 자료가 많기 때문에, 이 영역의 세포 구축적 특징과 처리하는 정보 사이에 일정한 관계가 있다고 생각된다.3. 대뇌 피질의 층 구조
대뇌 피질은 크게 6개의 층으로 구성된 신피질과 3~4개의 층으로 구성된 구피질로 나뉜다. 신피질은 대뇌 피질의 대부분(약 90%)을 차지하며, 각 층은 신경 세포의 종류와 연결 방식에서 차이를 보인다.[7] 이러한 층 구조는 대뇌 피질의 기능을 이해하는 데 중요한 역할을 한다.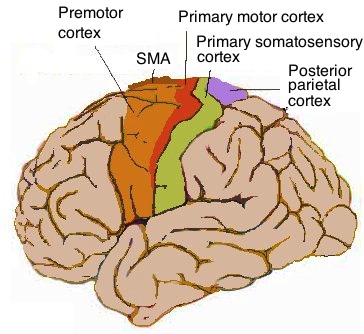
사람의 뇌에서 대뇌 피질은 2~4mm의 두께를 가지며,[6] 뇌 질량의 40%를 차지한다.[7] 피질에는 140억~160억 개의 뉴런이 존재하며,[7] 이 뉴런들은 피질 기둥과 피질 미니 기둥 형태로 배열되어 있다.[8][9]
신피질과 달리 구피질은 층 구조가 덜 분화되어 있다. 예를 들어, 해마는 3개의 세포층만을 갖는다.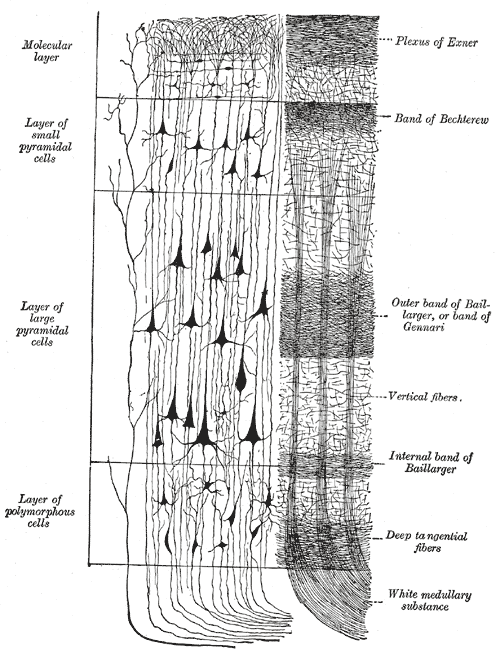
신피질과 구피질 사이에는 변연 피질이라는 전환 영역이 존재하며, 이곳에서는 2, 3, 4층이 합쳐져 있다.
3. 1. 신피질의 층 (Layers of neocortex)
신피질은 6개의 층으로 구성되며, 가장 바깥쪽 층인 I층(연질막에 가까움)부터 가장 안쪽 층인 VI층(백색질에 가까움)까지 번호가 매겨진다. 각 피질 층은 다른 뉴런 및 다른 피질 및 피질하 영역과의 연결이 특징적으로 분포되어 있다. 서로 다른 피질 영역 간의 직접적인 연결과 시상을 통한 간접적인 연결이 있다.피질 층상 구조의 가장 명확한 예 중 하나는 일차 시각 피질의 겐나리 선이다. 이것은 육안으로 볼 수 있는 흰색 조직 띠로, 후두엽의 갈고리 고랑에서 관찰할 수 있다. 겐나리 선은 시상에서 시각 정보를 시각 피질의 IV층으로 전달하는 축삭으로 구성된다.[35]
코르비니안 브로드만 (1909)의 연구는 포유류 신피질이 일관되게 6개의 층으로 나뉜다는 것을 확립했다.[34] 피질 층은 단순히 겹겹이 쌓여 있는 것이 아니라, 피질 전체 두께에 걸쳐 여러 층과 뉴런 유형 간의 특징적인 연결이 존재한다. 이러한 피질 미세 회로는 피질 기둥과 피질 소기둥으로 그룹화되며, 소기둥은 피질의 기본적인 기능 단위라고 제안되었다. 1957년, 버논 마운트캐슬은 피질의 기능적 특성이 측면 인접 지점 사이에서 갑자기 변하지만, 표면에 수직인 방향에서는 연속적임을 보여주었다. 이후 연구에서 시각 피질, 청각 피질, 연합 피질에서 기능적으로 구별되는 피질 기둥의 존재에 대한 증거가 제시되었다.
4층이 없는 피질 영역은 무과립 피질이라고 한다. 4층이 단지 초보적인 피질 영역은 이질과립이라고 한다.[36] 각 층 내의 정보 처리는 다른 시간 역학에 의해 결정되며, 2 Hz 신경 진동을 갖는 2/3층과 10–15 Hz 진동을 갖는 5층이 있다.[37]
대뇌 피질의 대부분 영역('''신피질''', 동종피질)에서는 신경 세포가 규칙적인 6층 구조를 이루며 정연하게 배열되어 있다. 이 6층은 바깥쪽부터 차례로 분자층, 외과립층, 외추체세포층, 내과립층, 내추체세포층(신경세포층), 다형세포층이라고 불린다.
코르비니안 브로드만은 신피질의 6층 구조를 자세히 관찰한 결과, 피질의 위치에 따라 일부 층이 두꺼워지거나 얇아지는 등 세포 구축이 균일하지 않다는 것을 발견했다. 브로드만은 세포 구축의 차이에 기반하여 대뇌 피질을 52개의 영역('''영역''' 또는 '''야''')으로 구분했다. 이 구분은 브로드만 영역이라고 불리며, 뇌 기능 국재론에서 위치를 나타내는 기준으로 자주 사용된다. 예를 들어, 후두엽에 위치한 신피질의 17야에서는 외측슬상체로부터 축삭을 받는 제4층이 두껍게 발달해 있는데, 외측슬상체는 망막으로부터 입력을 받으며, 17야는 시각 정보를 처리한다고 볼 수 있다.
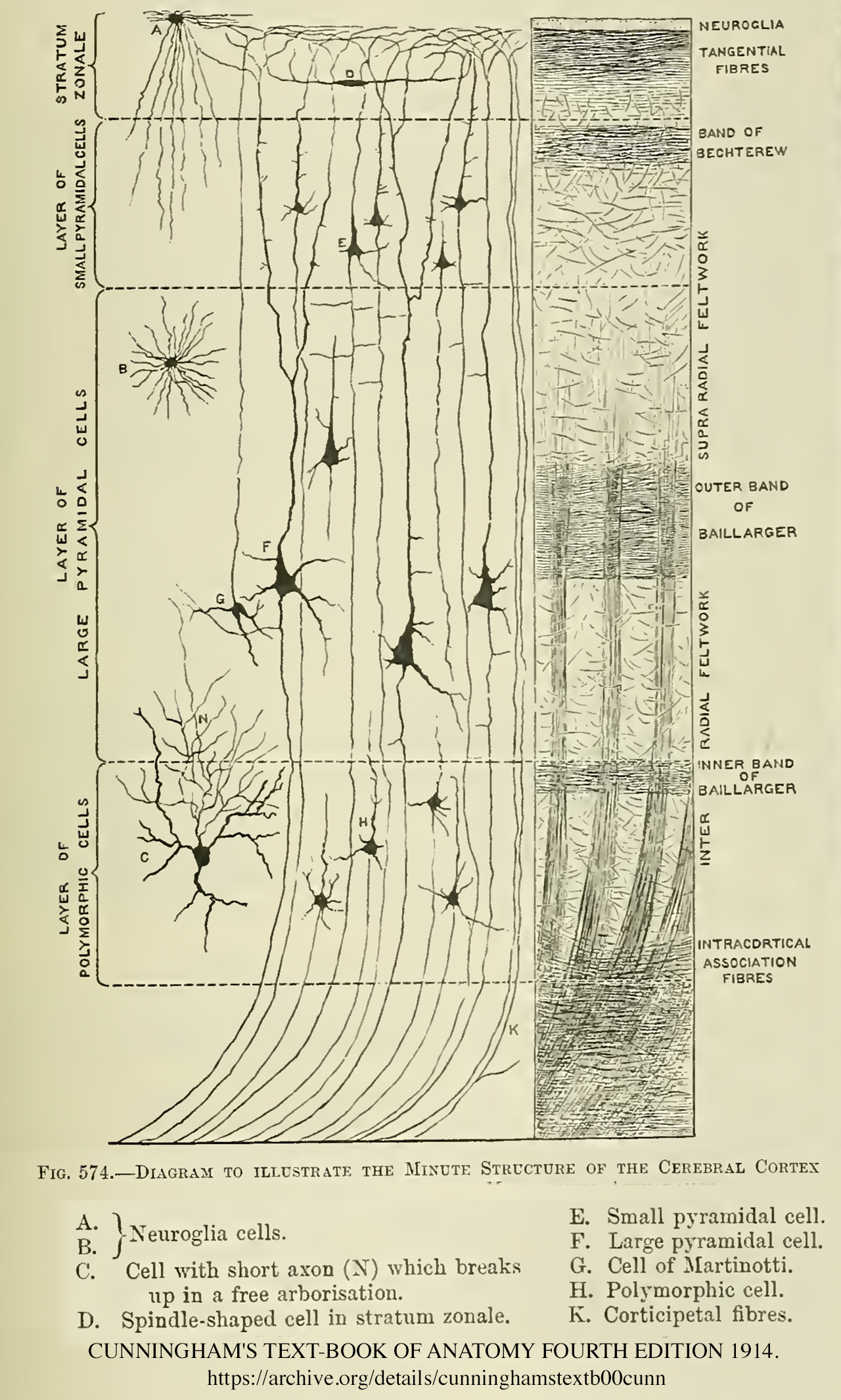
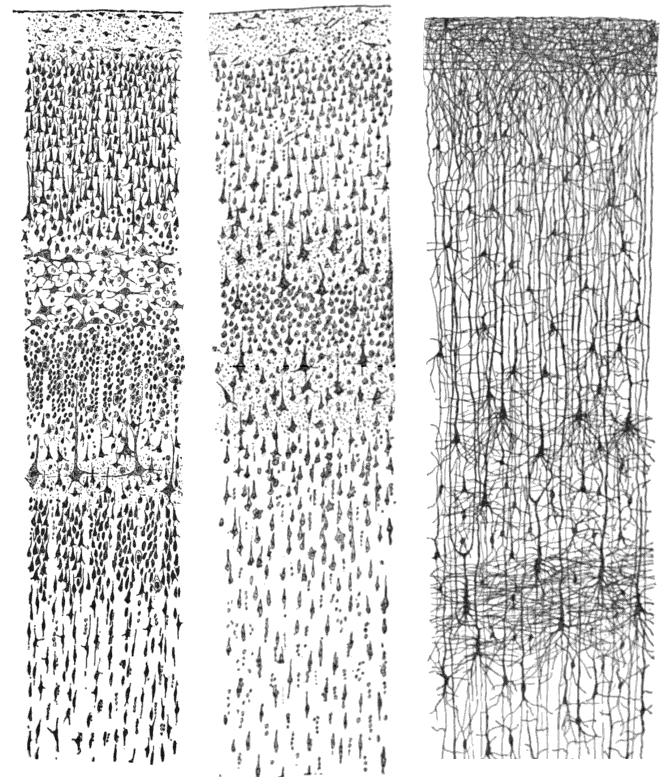
3. 1. 1. 1층: 분자층 (Molecular layer)
가장 바깥층으로, 신경세포가 거의 없다. 주로 가지돌기(dendrite)와 축삭(axon)의 종말가지로 이루어져 있다. 신경섬유에 대한 염색을 하였을 때 가지돌기와 축삭이 점상으로 관찰된다. 이 층에 분포하는 가지돌기는 피라밋세포(pyramidal neuron)에서 뻗어 나온 것이 대부분이다.[92] 반면 축삭은 같은쪽 및 반대쪽 대뇌반구의 대뇌피질 또는 시상에서 나온 것들로 구성되어 있다. 분자층은 주로 시냅스가 일어나는 층이라고 할 수 있다.I층은 '''분자층'''이며, GABA성 로즈힙 뉴런을 포함하여 드문드문 흩어져 있는 뉴런을 포함한다.[21] I층은 주로 피질 기둥 세포의 정단 수상돌기 뭉치와 수평으로 향하는 축삭의 연장선, 그리고 신경교세포로 구성된다.[22] 발생 과정에서 카할-레치우스 세포[23]와 아피아립 과립층 세포[24]가 이 층에 존재한다. 또한, 일부 가시 별세포도 여기서 발견될 수 있다. 정단 뭉치로의 입력은 연상 학습 및 주의와 관련된 대뇌 피질의 ''피드백'' 상호 작용에 중요한 역할을 하는 것으로 여겨진다.[25]
한때 I층으로의 입력이 피질 자체에서 오는 것으로 생각되었지만,[26] 현재는 대뇌 피질 전반의 I층이 IV층으로 가는 ''코어'' 또는 C형과는 대조적으로 ''매트릭스'' 또는 M형 시상 세포로부터 상당한 입력을 받는 것으로 알려져 있다.[27][28]
I층은 광범위한 정보를 수집하고 처리하는 중심 허브 역할을 하는 것으로 생각된다. 이는 상향 감각 입력과 하향 기대치를 통합하여 감각 인식이 예상 결과와 일치하는 방식을 조절한다. 또한 I층은 흥분성 입력을 분류, 지시 및 결합하여 신경 조절 신호와 통합한다. I층 내부 및 다른 피질 층에서 온 억제성 개재 뉴런은 이러한 신호를 게이트한다. 이러한 상호 작용은 신피질 전체의 정보 흐름을 동적으로 조정하여 지각과 경험을 형성한다.[29]
3. 1. 2. 2층: 바깥과립층 (External granular layer)
주로 별세포(성상세포)와 작은 피라미드 세포로 구성되어 있다. 이 세포들의 돌기들은 분자층에서 끝나며 축삭은 더 깊은 층으로 들어가거나 대뇌반구의 백질까지 들어간다.[1]3. 1. 3. 3층: 바깥피라밋층 (External pyramidal layer)
바깥피라밋층은 전형적인 피라밋세포로 구성된 층으로, 표면에서 깊어질수록 큰 신경세포가 분포하는 특징을 보인다. 이 층에 위치한 피라밋세포의 돌기들은 분자층으로 들어가고, 축삭은 대뇌 피질의 다른 부위로 뻗는 연합섬유 또는 맞교차섬유가 된다.[1] 제3층은 피질-피질 원심성 신경 섬유의 주요 공급원이며, 제1층부터 제3층까지는 교련 피질-피질 구심성 신경 섬유의 주요 표적이다.[1]3. 1. 4. 4층: 속과립층 (Internal granular layer)
주로 별세포로 이루어진 층이며, 다른 사이신경세포나 피라밋세포도 약간 섞여 있다. 시상으로부터 들어오는 구심성 경로의 종착지이기도 하다.[93] 내부 과립층에는 다양한 유형의 별세포와 피라미드 세포가 있으며, 시상피질 방사선으로부터의 시상 C형 뉴런(핵심형)의 구심성 섬유와 반구 내 피질-피질 구심성 섬유의 주요 표적이다. 제4층 위의 층은 상과립층(제1-III층)이라고도 하며, 아래 층은 하과립층(제V 및 VI층)이라고 한다. 아프리카 코끼리, 고래류, 하마는 제4층이 없다.3. 1. 5. 5층: 속피라밋층 (Internal pyramidal layer)
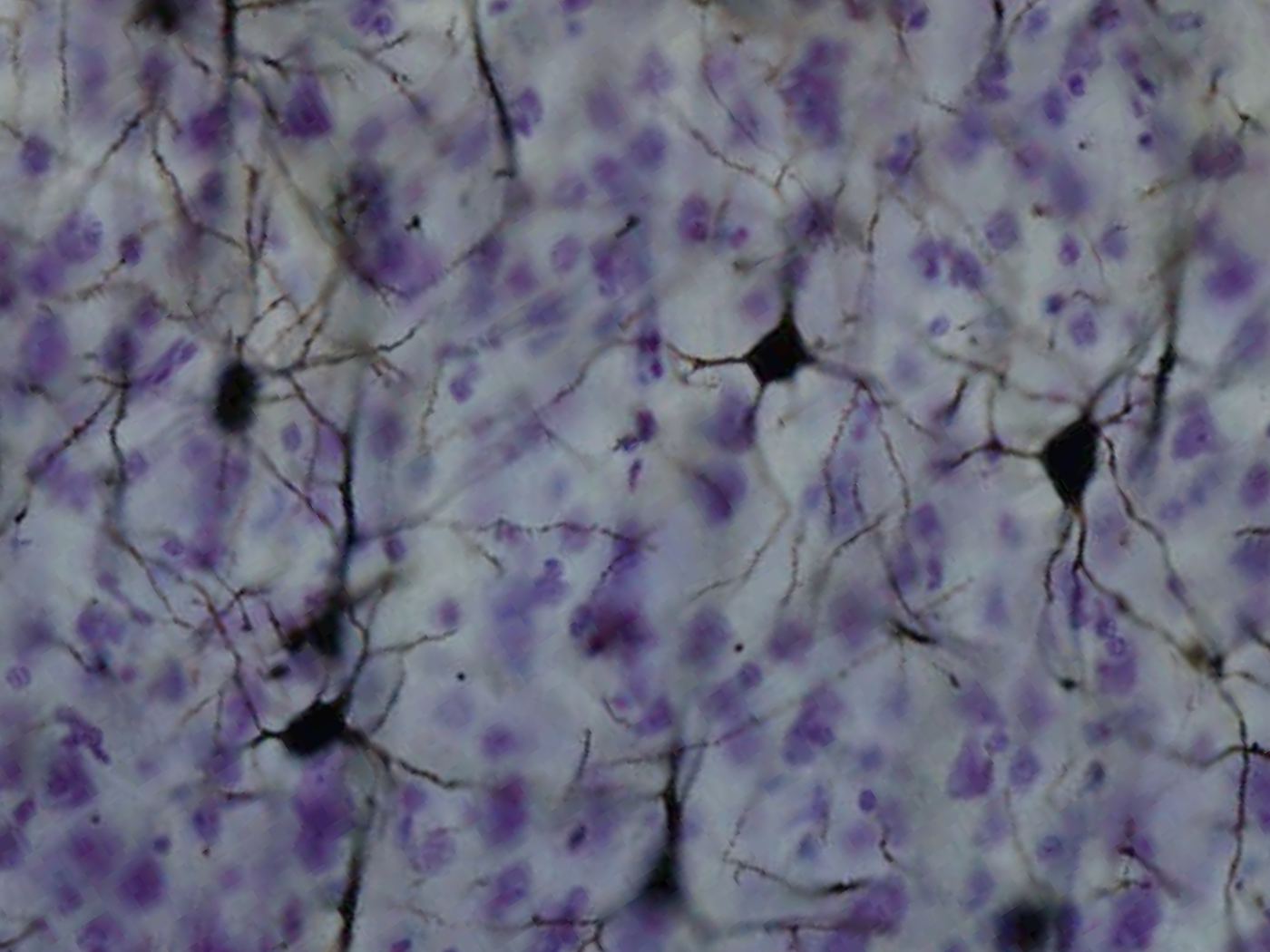
바깥피라밋층에 비해 직경이 큰 피라밋세포가 분포하는 층으로, 사이사이에 많은 사이신경세포가 섞여 있다. 속피라밋층에 위치한 피라밋세포의 축삭은 새줄무늬체, 뇌줄기, 척수 등의 피질하영역으로 뻗는 투사섬유가 된다.[43] 중심전회의 운동피질에서 속피라밋층의 피라밋세포들은 크고 베츠세포라고 불린다. 이러한 세포들은 피질척수로나 추체로의 투사섬유의 3% 정도를 차지한다.[43]
3. 1. 6. 6층: 뭇모양층 (Multiform layer)
이 층에는 방추세포가 분포하는 것이 특징이지만, 대부분의 세포들은 삼각형 모양의 세포체를 갖는 피라밋세포와 여러 종류의 사이신경세포도 섞여 있다. 말티노티세포들도 다모양층에 많이 있다. 백질로 들어가고 나오는 세포들이 많이 있으며, 특히 시상으로 내보내는 원심성 경로의 시점이기도 하다.[94]제6층은 소수의 큰 피라미드 뉴런과 많은 작은 방추형 및 다형 뉴런을 포함한다. 제6층은 원심성 신경 섬유를 시상으로 보내 뇌 피질과 시상 사이에 매우 정밀한 상호 연결을 설정한다.[31] 즉, 하나의 피질 기둥에서 제6층 뉴런은 동일한 피질 기둥에 입력을 제공하는 시상 뉴런과 연결된다. 이러한 연결은 흥분성이자 억제성이다. 뉴런은 흥분성 시냅스 후 전위 섬유를 시상의 뉴런으로 보내고 또한 이러한 동일한 시상 뉴런 또는 그에 인접한 뉴런을 억제성 시냅스 후 전위하는 시상 망상 핵으로 측부 섬유를 보낸다.[32] 한 가지 이론은 콜린성 입력에 의해 대뇌 피질로의 억제성 출력이 감소하기 때문에, 이것이 뇌간에 조절 가능한 "후주-내측 모서리 신경로 입력의 중계에 대한 게인 제어"를 제공한다는 것이다.[32]
3. 2. 구피질 (Allocortex)
층상 구조의 차이에 따라 대뇌 피질은 6개의 세포층을 가진 넓은 영역인 신피질과 3~4개의 층을 가진 훨씬 작은 영역인 구피질의 두 가지 유형으로 분류할 수 있다.[2] 구피질은 3~4개의 층을 가진 대뇌 피질의 일부이며, 후각 피질과 해마 등이 이에 해당한다. 구피질은 3개의 피질 층판을 가진 고피질, 4~5개의 층을 가진 궁피질, 그리고 구피질에 인접한 전환 영역인 주변피질의 3가지 아형으로 나뉜다.4. 기능
대뇌 피질은 감각, 운동, 인지 등 다양한 기능을 수행하며, 기능에 따라 여러 영역으로 나뉜다.
코르비니안 브로드만은 대뇌 피질을 세포구축학적 특징에 따라 52개의 브로드만 영역으로 나누었으며, 이들은 각기 다른 기능을 담당한다. 예를 들어, 브로드만 영역 17은 일차 시각 피질에 해당한다.
일반적으로 대뇌 피질은 감각 영역, 운동 영역, 연합 영역으로 구분된다.
- 감각 영역: 감각 정보를 받아들이고 처리하는 영역이다. 시상에서 오는 감각 입력을 받는 부분을 일차 감각 영역이라고 한다. 일차 시각 피질, 일차 청각 피질, 일차 체감각 피질 등이 여기에 속한다.
- 운동 영역: 몸의 움직임을 조절하는 영역이다. 일차 운동 피질은 실제 움직임을 실행하고, 보조 운동 영역과 전운동 피질은 움직임을 계획하고 선택한다.
- 연합 영역: 감각 정보와 기억을 통합하여 세상을 이해하고, 효과적으로 상호작용하며, 추상적인 사고와 언어를 사용할 수 있게 하는 영역이다. 두정엽, 측두엽, 후두엽은 감각 정보와 기억을 통합하고, 전두엽은 행동 계획과 추상적 사고를 담당한다.
대뇌 피질은 시상이나 기저핵과 같은 피질하 구조와 연결되어 정보를 주고받는다. 대부분의 감각 정보는 시상을 거쳐 피질로 전달되지만, 후각 정보는 후구를 통해 이상 피질로 바로 전달된다.
4. 1. 감각 영역 (Sensory areas)
감각 영역은 감각으로부터 정보를 수신하고 처리하는 피질 영역이다.[68] 시상으로부터 감각 입력을 받는 피질의 부분은 일차 감각 영역이라고 불린다. 시각, 청각 및 촉각은 각각 일차 시각 피질, 일차 청각 피질, 일차 체감각 피질에 의해 처리된다. 일반적으로 두 반구는 인체의 반대쪽(대측)에서 정보를 받는다. 예를 들어, 오른쪽 일차 체감각 피질은 왼쪽 사지로부터 정보를 받고, 오른쪽 시각 피질은 왼쪽 시각 수용 영역으로부터 정보를 받는다.[68]피질의 감각 지도의 구성은 지도식 표현으로 알려진 해당 감각 기관의 구성과 일치한다. 예를 들어, 일차 시각 피질의 인접한 지점은 망막의 인접한 지점에 해당한다. 이 지도식 표현은 망막국소지도라고 불린다. 마찬가지로 일차 청각 피질에는 음조지도가 있고, 일차 감각 피질에는 체감각지도가 있다. 몸의 이 마지막 지도식 표현이 후중심이랑에 나타나는데, 이는 체감각 피질 호문쿨루스라는 변형된 인간 표현으로, 서로 다른 신체 부위의 크기는 그들의 신경 지배의 상대적 밀도를 반영한다. 손가락 끝과 입술과 같이 감각 신경 지배가 많은 영역은 더 미세한 감각을 처리하기 위해 더 많은 피질 영역이 필요하다.[68]
4. 2. 운동 영역 (Motor areas)
운동 영역은 대뇌 피질의 양쪽 반구에 위치하며, 자발적인 움직임, 특히 손으로 수행하는 미세하고 분절적인 움직임을 제어하는 데 매우 밀접하게 관련되어 있다. 운동 영역의 오른쪽 반구는 신체의 왼쪽을 제어하고 그 반대도 마찬가지이다.
일반적으로 피질의 두 영역이 운동 영역으로 불린다.
- 일차 운동 피질은 자발적인 움직임을 ''실행''한다.
- 보조 운동 영역과 전운동 피질은 자발적인 움직임을 ''선택''한다.
다음과 같은 영역에서도 운동 기능이 설명된다.
- 후두정 피질은 공간에서 자발적인 움직임을 안내한다.
- 배외측 전전두피질은 고차원적인 지시, 규칙 및 자기 생성된 생각을 바탕으로 어떤 자발적인 움직임을 할지 결정한다.
대뇌 피질 바로 아래에는 기저핵이라고 불리는 회백질의 상호 연결된 피질하 덩어리가 있다. 기저핵은 중뇌의 흑색질과 대뇌 피질의 운동 영역으로부터 입력을 받아 이 두 위치로 신호를 다시 보낸다. 기저핵은 운동 제어에 관여하며, 시상의 외측에서 발견된다.
4. 3. 연합 영역 (Association areas)
연합 영역은 대뇌 피질에서 일차적인 감각 영역이나 운동 영역에 속하지 않는 부분을 말한다. 이 영역들은 감각 정보와 기억에 저장된 정보를 통합하여, 우리가 세상을 의미 있게 이해하고, 효과적으로 상호작용하며, 추상적인 사고와 언어를 사용할 수 있도록 돕는다.두정엽, 측두엽, 후두엽은 모두 대뇌 피질의 뒤쪽에 위치하며, 감각 정보와 기억을 통합하는 역할을 한다. 전두엽 또는 전전두 연합 복합체는 행동과 움직임을 계획하고 추상적인 사고를 하는 데 관여한다. 전체적으로 연합 영역은 뇌의 여러 부분에 흩어져 있는 네트워크로 구성되어 있다.[73] 각 네트워크는 서로 멀리 떨어진 영역들을 연결하며, 이 네트워크들은 서로 복잡하게 얽혀 있다.
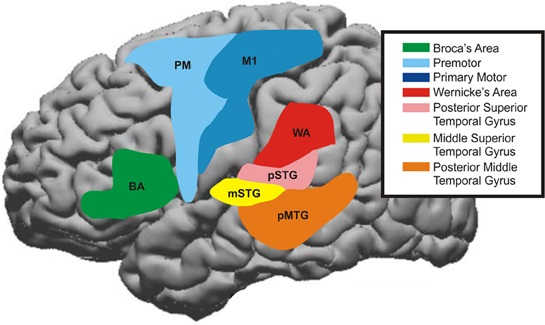
특히 인간의 경우, 연합 네트워크는 언어 기능에 매우 중요하다. 과거에는 언어 능력이 주로 좌측 하전두이랑의 브로카 영역(BA44, BA45)과 베르니케 영역(BA22)에 국한된다고 생각했다. 그러나 최근 연구에 따르면 언어 표현과 이해 과정은 가쪽고랑 주변뿐만 아니라 전두엽, 기저핵, 소뇌, 교 등 다른 영역에서도 일어나는 것으로 밝혀졌다.[74]
5. 발생
대뇌 피질은 태아의 발달 과정 중 신경관의 가장 앞쪽 부분인 앞뇌 영역에서 발달한다.[39][40] 신경판이 접혀서 닫히면서 신경관을 형성하며, 신경관 내부의 강에서 뇌실계가 발달하고, 벽의 신경상피 세포에서 신경계의 뉴런과 신경교세포가 발달한다. 신경관 형성이 시작되기 전 신경판의 가장 앞쪽 부분인 전뇌는 대뇌 반구와 이후 피질을 생성한다.[41]
뇌실 구역 내, 뇌실 옆에서 생성되는 대뇌 피질 뉴런은 처음에는 신경 줄기 세포 상태였다가, 방사상 교세포—전구 세포로 전환되어 분열하여 교세포와 뉴런을 생성한다.[42]
5. 1. 피질 규칙화 (Cortical patterning)
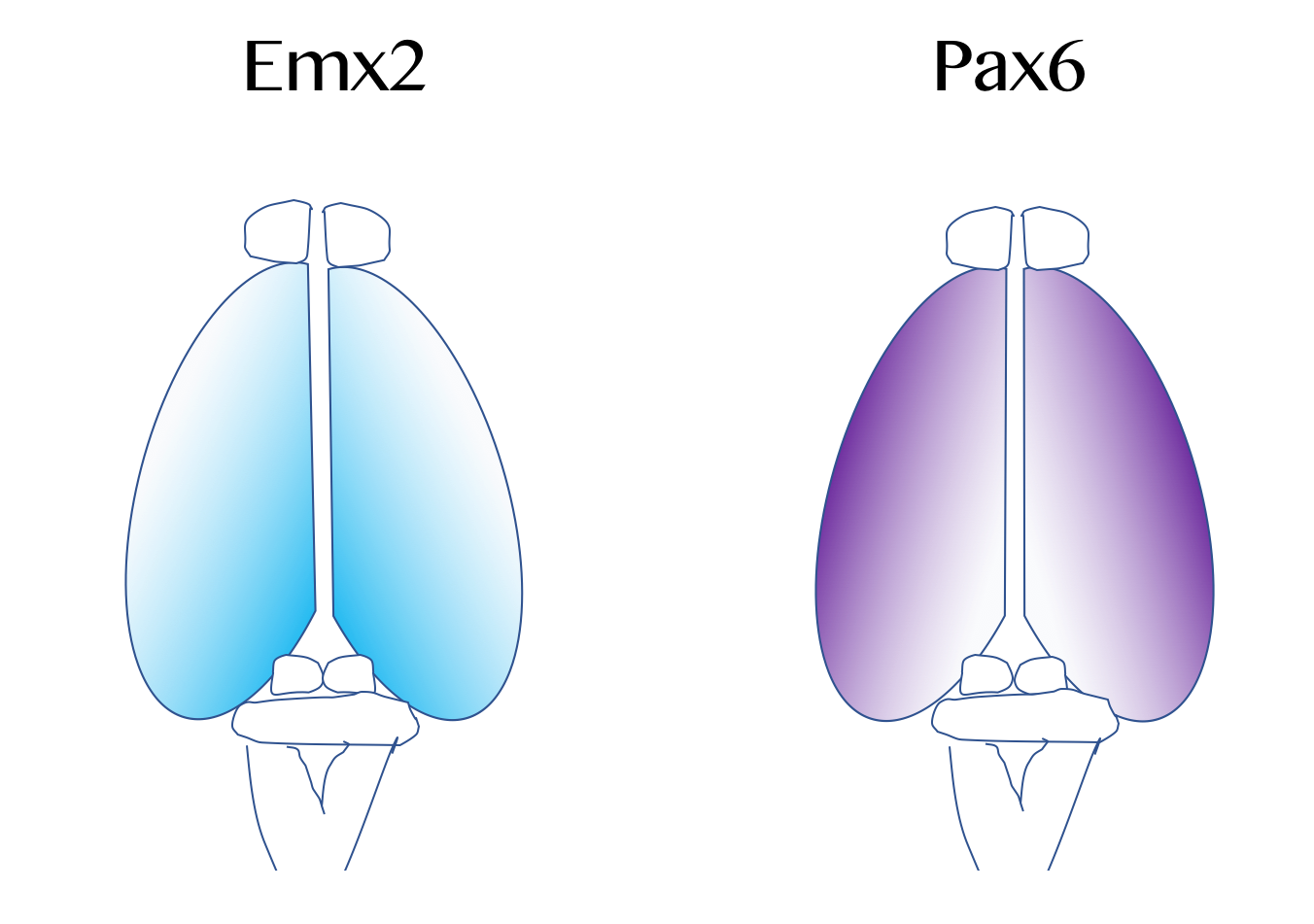
대뇌 피질은 일차운동피질이나 시각피질처럼 각 영역이 특별한 기능에 대응되는 경향이 있다. 이러한 해부-기능 간 대응 관계는 원지도(protomap)라고 하여 태아 발달 초기 단계에서 FGF8 등 섬유아세포 성장인자라는 분자의 분포에 따라 미리 결정된다.[99][100][101] 섬유아세포 성장인자의 분포에 따라 대뇌 피질의 크기, 형태, 위치 등이 결정되는데, 이를 피질 규칙화(cortical patterning)라 한다.EMX2와 PAX6이 대표적인 예시다.[102] 이들은 서로 피질 내에서 농도 기울기가 반대로 분포하고 있는데, EMX2는 중앙/후방으로 갈수록 그 농도가 높고 PAX6는 가측/전방으로 갈수록 그 농도가 높다. 만일 이 농도 기울기가 이 상태로 유지되지 않는다면 대뇌피질이 정상적으로 발달하지 않는다는 것이 밝혀졌다. 일례로, PAX6에 돌연변이가 생긴 경우 EMX2가 비정상적으로 많이 분비되어 시각 피질 등 중앙/후방에 배치된 피질의 영역이 비정상적으로 커지게 된다. 그 반대의 경우도 마찬가지로, PAX6가 과분비되는 경우 전두엽과 운동피질의 영역이 비정상적으로 커진다. 따라서 이 두 물질들은 대뇌피질의 발달에 관여하는 대표적인 세포 신호전달물질로 거론된다.[103] FGF8이 대뇌 전방에서 과분비될 경우 후방의 EMX2 농도가 과다분비된다.[103]
6. 임상적 의의
대뇌 피질의 손상이나 질병은 다양한 신경학적, 정신적 문제를 일으킬 수 있다. 알츠하이머병과 같은 신경퇴행성 질환은 대뇌 피질의 회백질 위축을 보인다.[76]
뇌 손상은 전두엽 장애와 같이 특정 엽에 손상을 일으켜 관련 기능에 영향을 미칠 수 있다. 뇌를 감염으로부터 보호하는 혈액-뇌 장벽이 손상되면 병원체가 유입될 수 있다.
피질 자극 매핑이라고 하는 뇌피질전도검사는 피질의 특정 영역의 기능을 국소화하기 위해 뇌에 직접 전극을 배치하는 침습적 절차이다. 이는 수술 전 매핑을 포함한 임상 및 치료 분야에 사용된다.[78]
6. 1. 신경퇴행성 질환
알츠하이머병과 라포라병은 신경퇴행성 질환으로 대뇌 피질의 위축을 동반한다. 따라서 대뇌 피질을 관찰하여 이러한 질병을 판단할 수 있다.[76]6. 2. 뇌졸중
뇌졸중과 같은 뇌혈관 질환으로 인해 대뇌 피질에 혈액 공급이 차단되면 해당 영역의 기능이 손상될 수 있다.6. 3. 뇌전증 (간질)
뇌전증과 같은 신경 질환은 대뇌 피질과 관련된 중추 신경계 질환 중 하나이다.[76] 뇌전증은 대뇌 피질의 비정상적인 전기 활동으로 인해 발작이 발생할 수 있다는 특징을 가진다.6. 4. 발달 장애
태아 발달 중인 태아는 선천적 결함 및 후기 발달 문제를 일으킬 수 있는 다양한 환경적 요인에 취약하다. 예를 들어, 산모의 알코올 섭취는 태아 알코올 스펙트럼 장애를 유발할 수 있다.[77] 신경 발달 질환을 유발할 수 있는 다른 요인으로는 독성 물질(예: 약물)과 X-ray와 같은 방사선 노출이 있다. 감염 또한 피질 발달에 영향을 미칠 수 있다. 바이러스 감염은 뇌이랑 없음증의 원인 중 하나이며, 이는 이랑 형성이 없는 매끄러운 피질을 초래한다.대뇌 피질의 다양한 유전 질환을 유발할 수 있는 많은 수의 유전자 변이가 있는데, 여기에는 소두증, 분열뇌증 및 여러 유형의 활뇌증이 포함된다.[79] 염색체 이상 또한 취약 X 증후군 및 레트 증후군과 같은 여러 신경 발달 장애를 유발할 수 있다.
MCPH1은 소두증을 암호화하며, 이 유전자와 ASPM의 장애는 소두증과 관련이 있다.[79] nibrin을 암호화하는 유전자 NBS1의 돌연변이는 소두증이 특징인 네이메헨 파열 증후군을 유발할 수 있다.[79]
EMX2,[80] 및 COL4A1의 돌연변이는 대뇌 반구의 큰 부분의 부재가 특징인 질환인 분열뇌증과 관련이 있다.[81]
6. 5. 기타 질환
알츠하이머병과 같은 신경퇴행성 질환은 대뇌 피질의 회백질 위축을 지표로 보여준다.[76]다른 중추 신경계 질환에는 뇌전증, 운동 질환, 다양한 유형의 실어증 (언어 표현 또는 이해의 어려움)과 같은 신경 질환이 포함된다.
질병이나 외상으로 인한 뇌 손상은 전두엽 장애와 같이 특정 엽에 손상을 일으켜 관련 기능에 영향을 미칠 수 있다. 뇌를 감염으로부터 보호하는 혈액-뇌 장벽이 손상되면 병원체가 유입될 수 있다.
태아 발달 중인 태아는 선천적 결함 및 후기 발달 문제를 일으킬 수 있는 다양한 환경적 요인에 취약하다. 예를 들어, 산모의 알코올 섭취는 태아 알코올 스펙트럼 장애를 유발할 수 있다.[77] 신경발달 질환을 유발할 수 있는 다른 요인으로는 독성 물질 (예: 약물)과 X-ray와 같은 방사선 노출이 있다. 감염 또한 피질 발달에 영향을 미칠 수 있다. 바이러스 감염은 뇌이랑 없음증의 원인 중 하나이며, 이는 이랑 형성이 없는 매끄러운 피질을 초래한다.
피질 자극 매핑이라고 하는 한 유형의 뇌피질전도검사는 피질의 특정 영역의 기능을 국소화하기 위해 노출된 뇌에 직접 전극을 배치하는 침습적 절차이다. 이는 수술 전 매핑을 포함한 임상 및 치료 적용 분야에 사용된다.[78]
참조
[1]
웹사이트
cerebral mantle
https://www.thefreed[...]
2024-05-09
[2]
서적
Noback's Human Nervous System
Humana Press
2012
[3]
논문
Cerebral cortex expansion and folding: what have we learned?
2016-05
[4]
논문
Evolution of the neocortex: a perspective from developmental biology
2009-10
[5]
서적
Principles of neural science
https://archive.org/[...]
McGraw-Hill, Health Professions Division
2000-01-05
[6]
서적
Neuroanatomy
Springer-Verlag
1992
[7]
서적
Human anatomy
McGraw-Hill
2011
[8]
논문
Generating neuronal diversity in the mammalian cerebral cortex
2015-11-13
[9]
논문
Gene-environment interactions in cortical interneuron development and dysfunction: A review of preclinical studies
2017-01
[10]
서적
Core text of neuroanatomy
Williams & Wilkins
1985
[11]
논문
Brain size and folding of the human cerebral cortex
2008-10
[12]
서적
Evolution of the Primate Brain
2012
[13]
서적
Principles of anatomy & physiology.
Wiley
2011
[14]
서적
The central nervous system of vertebrates, Volume 1
Springer
[15]
논문
Analyzing the neocortical fine-structure
2003-09
[16]
논문
Relationships between IQ and regional cortical gray matter thickness in healthy adults
2007-09
[17]
논문
Thickening in the somatosensory cortex of patients with migraine
2007-11
[18]
뉴스
Migraine Sufferers Have Thicker Brain Cortex
http://www.medicalne[...]
2007-11-20
[19]
논문
Absence of changes in cortical thickness in patients with migraine
2011-10
[20]
논문
The neurological basis of developmental dyslexia: an overview and working hypothesis
2000-12
[21]
웹사이트
Scientists identify a new kind of human brain cell
https://www.allenins[...]
2018-08-27
[22]
논문
Structure and function of the cerebral cortex
2007-06
[23]
논문
What is a Cajal-Retzius cell? A reassessment of a classical cell type based on recent observations in the developing neocortex
1999-12
[24]
논문
The discovery of the subpial granular layer in the human cerebral cortex
2010
[25]
논문
Brain states: top-down influences in sensory processing
2007-06
[26]
논문
Layer I of primary sensory neocortex: where top-down converges upon bottom-up
1995-11
[27]
논문
Thalamic input to distal apical dendrites in neocortical layer 1 is massive and highly convergent
2009-10
[28]
논문
Viewpoint: the core and matrix of thalamic organization
1998-07
[29]
논문
Layer 1 neocortex: Gating and integrating multidimensional signals
2024-01
[30]
논문
Where Do Core Thalamocortical Axons Terminate in Mammalian Neocortex When There Is No Cytoarchitecturally Distinct Layer 4?
2024-07
[31]
간행물
Cortex Cerebri
Springer-Verlag
1995
[32]
논문
Functional organization of the somatosensory cortical layer 6 feedback to the thalamus
2010-01
[33]
논문
Neocortical neurogenesis is not really "neo": a new evolutionary model derived from a comparative study of chick pallial development
2013-01
[34]
논문
The columnar organization of the neocortex
1997-04
[35]
논문
Receptive fields of single neurones in the cat's striate cortex
1959-10
[36]
간행물
Quantitative Architecture Distinguishes Prefrontal Cortical Systems in the Rhesus Monkey
http://cercor.oxford[...]
Cereb. Cortex
2008-08-29
[37]
논문
Layer-specific network oscillation and spatiotemporal receptive field in the visual cortex
2009-10
[38]
논문
Temporal specification and bilaterality of human neocortical topographic gene expression
2014-01
[39]
서적
Principles of Development
Oxford University Press
2015
[40]
논문
The transcription factor, Pax6, is required for cell proliferation and differentiation in the developing cerebral cortex
1999-09
[41]
서적
Human Embryology
Churchill Livingstone
2001
[42]
논문
Neurons derived from radial glial cells establish radial units in neocortex
2001-02
[43]
논문
Development and plasticity of cortical areas and networks
2001-04
[44]
서적
Development of the Nervous System
Elsevier Inc.
[45]
논문
Evolution of the neocortex: a perspective from developmental biology
2009-10
[46]
논문
Extrinsic cytological determinants of basket and stellate cell dendritic pattern in the cerebellar molecular layer
1972-11
[47]
서적
Developmental Biology
Sinauer Associates Publishers
2006
[48]
논문
Selective lengthening of the cell cycle in the neurogenic subpopulation of neural progenitor cells during mouse brain development
2005-07
[49]
논문
Specification of cerebral cortical areas
1988-07
[50]
논문
Epigenetic control on cell fate choice in neural stem cells
2012-04
[51]
논문
Developmental history of the transient subplate zone in the visual and somatosensory cortex of the macaque monkey and human brain
1990-07
[52]
논문
Neurons in rhesus monkey visual cortex: systematic relation between time of origin and eventual disposition
1974-02
[53]
논문
Development of layer I neurons in the primate cerebral cortex
2001-08
[54]
논문
Specification of cerebral cortical areas
1988-07
[55]
논문
Neocortex patterning by the secreted signaling molecule FGF8
2001-11
[56]
논문
Molecular regionalization of the neocortex is disrupted in Fgf8 hypomorphic mutants
2003-05
[57]
논문
Regulation of area identity in the mammalian neocortex by Emx2 and Pax6
2000-04
[58]
논문
FGF signaling expands embryonic cortical surface area by regulating Notch-dependent neurogenesis
2011-10
[59]
논문
Local tissue growth patterns underlying normal fetal human brain gyrification quantified in utero
2011-02
[60]
논문
Development and evolution of the human neocortex
2011-07
[61]
논문
Trnp1 regulates expansion and folding of the mammalian cerebral cortex by control of radial glial fate
2013-04
[62]
논문
Hedgehog signaling promotes basal progenitor expansion and the growth and folding of the neocortex
2016-07
[63]
논문
Cortical gyrification induced by fibroblast growth factor 2 in the mouse brain
2013-06
[64]
논문
Specification of cerebral cortical areas
1988-07
[65]
논문
Neocortex patterning by the secreted signaling molecule FGF8
2001-11
[66]
논문
Regulation of area identity in the mammalian neocortex by Emx2 and Pax6
2000-04
[67]
논문
Generating the cerebral cortical area map
2003
[68]
서적
Cortex: Statistics and Geometry of Neuronal Connectivity.
Springer Science & Business Media
1998
[69]
서적
Functional and Clinical Neuroanatomy: A Guide for Health Care Professionals
Academic Press
2020-01
[70]
서적
Encyclopedia of Behavioral Neuroscience
Academic Press
2010-01
[71]
서적
Anatomy and Physiology: The Unity of Form and Function, 5th Ed.
McGraw-Hill Companies Inc.
2010
[72]
서적
Dorland's Medical Dictionary for Health Consumers
2008
[73]
논문
The organization of the human cerebral cortex estimated by intrinsic functional connectivity
2011-09
[74]
논문
The anatomy of language: contributions from functional neuroimaging
2000-10
[75]
논문
Detection of spreading depolarizations in a middle cerebral artery occlusion model in swine
2020-03
[76]
논문
Multiple-region grey matter atrophy as a predictor for the development of dementia in a community: the Hisayama Study
2022-03
[77]
논문
Fetal alcohol spectrum disorder: an overview
2006-06
[78]
논문
Preoperative multimodal motor mapping: a comparison of magnetoencephalography imaging, navigated transcranial magnetic stimulation, and direct cortical stimulation
2012-08
[79]
논문
Genetic basis of developmental malformations of the cerebral cortex
2004-05
[80]
웹사이트
EMX2 empty spiracles homeobox 2 [Homo sapiens (human)]
https://www.ncbi.nlm[...]
National Center for Biotechnology Information, U.S. National Library of Medicine
[81]
논문
Novel COL4A1 mutation in an infant with severe dysmorphic syndrome with schizencephaly, periventricular calcifications, and cataract resembling congenital infection
2016-04
[82]
서적
Principles of Neural Science
McGraw-Hill Medical
2013
[83]
논문
A cortex-like canonical circuit in the avian forebrain
2020-09
[84]
논문
A neural correlate of sensory consciousness in a corvid bird
2020-09
[85]
논문
Birds do have a brain cortex-and think
2020-09
[86]
논문
Profiling by image registration reveals common origin of annelid mushroom bodies and vertebrate pallium
2010-09
[87]
웹사이트
BrainVoyager Brain Tutor
http://www.brainvoya[...]
[88]
사전
전뇌
[89]
웹사이트
대뇌변연계
https://www.excite.c[...]
2021-03-11
[90]
서적
Human anatomy
https://archive.org/[...]
McGraw-Hill
2011
[91]
서적
Noback's Human Nervous System, Seventh Edition
Humana Press
2012
[92]
저널
Structure and function of the cerebral cortex
2007-06-17
[93]
저널
Viewpoint: the core and matrix of thalamic organization
[94]
서적
Cortex Cerebri
Springer-Verlag
1995
[95]
서적
Clinical neuroanatomy
https://www.worldcat[...]
McGraw-Hill Medical
2009
[96]
서적
Gray's clinical neuroanatomy : the anatomic basis for clinical neuroscience
https://www.worldcat[...]
Elsevier/Saunders
2019-04-12
[97]
저널
Vergleichende Lokalisationslehre der Grosshirnrinde
Johann Ambrosius Barth
[98]
저널
Temporal Specification and Bilaterality of Human Neocortical Topographic Gene Expression
2014-01-22
[99]
저널
Specification of cerebral cortical areas
1988-07-08
[100]
저널
Neocortex patterning by the secreted signaling molecule FGF8
2001-11-02
[101]
저널
Molecular regionalization of the neocortex is disrupted in Fgf8 hypomorphic mutants.
2003-05
[102]
저널
Regulation of area identity in the mammalian neocortex by Emx2 and Pax6
2000-04-14
[103]
서적
Development of the Nervous System
https://archive.org/[...]
Elsevier Inc.
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com