오구동물
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
오구동물은 1에서 14cm 크기의 벌레 모양을 한 기생 동물로, 파충류, 조류, 포유류의 상부 호흡기에서 서식하며, 때로는 인간에게도 감염을 일으킨다. 오랫동안 절지동물과의 연관성이 제기되었으나, 분자계통분석을 통해 갑각류의 새미류와 근연관계임이 밝혀졌고, 어류흡충강으로 분류된다. 오구동물은 4개의 현존하는 목과 2개의 아강으로 나뉘며, 캄브리아기 화석 기록을 통해 오구동물의 초기 진화와 기생 생활의 시작을 알 수 있다.
더 읽어볼만한 페이지
- 소악강 - 따개비
따개비는 바다에 서식하며 단단한 표면에 붙어사는 갑각류로, 석회질 껍데기로 덮여 있고 만각을 이용해 플랑크톤을 섭취하며, 조간대에서 저조선까지 분포, 암수한몸이지만 교미를 통해 번식하며, 다양한 종이 존재하고 해안 생태계에서 중요한 역할을 하지만 기후 변화와 해양 오염으로 위협받고 있다. - 소악강 - 새미류
새미류는 어류나 개구리류에 기생하여 체액을 섭취하는 작은 갑각류 외부 기생 생물로, 원반 모양의 몸과 흡반, 유영 부속지를 가지며 양어장에서 어류에게 피해를 주기도 하고, 한국에서는 새우라고도 불리지만 갑각류 새우와는 다른 종류이다.

2. 계통 분류
오구동물의 생물학 및 계통 분류에 대한 초기 연구는 요제프 알로이스 프뢸리히,[2] 알렉산더 폰 훔볼트,[3] 카를 아스문트 루돌피,[4] 카를 모리츠 디에징,[5] 루돌프 로이카르트[6] 등에 의해 이루어졌다. 이후 루이스 웨스턴라 삼본,[7] 리하르트 하이몬스,[8] 존 라일리[9] 등이 중요한 요약 연구를 발표했으며, J. T. 셀프는 1969년까지의 연구 문헌과 함께 오구동물의 진화적 관계를 검토했다.[10]
오구동물의 계통학적 위치는 오랫동안 논쟁의 대상이었으나, 화석 증거는 이들의 고대 기원을 시사한다. 스웨덴 상부 캄브리아기 오르스텐 지역[24]과 캐나다 캄브리아기-오르도비스기 경계[25], 그린란드 중기 캄브리아기[26] 등에서 3차원적으로 잘 보존된 인산염화된 오구동물 화석이 발견되었다. 캄브리아기에서는 ''Aengapentastomum'', ''Boeckelericambria'', ''Dietericambria'', ''Haffnericambria'', ''Heymonsicambria'' 등 4개의 화석 속이 확인되었다. 이 화석들은 오구동물이 매우 이른 시기에 진화했음을 보여주며, 당시 개충강(원시 어류)과 같은 숙주에 기생했을 가능성을 제기한다.[25] 영국 실루리아기 해양 지층에서는 다섯 번째 화석 속인 ''Invavita''가 발견되었는데, 이 표본은 개충강 숙주(''Nymphatelina gravida'')에 부착된 상태로 발견되어,[27][28] 당시 오구동물의 기생 생활에 대한 직접적인 증거를 제공한다. ''Invavita''는 머리, 벌레 모양 몸통, 두 쌍의 다리를 가진 형태였다.[29]
현존하는 오구동물(Pentastomida)은 크게 4개의 목으로 분류된다.
- 두배동물목(Cephalobaenida)
- 포로세팔루스목(Porocephalida)
- 라일리엘라목(Raillietiellida)
- 레이가르티아목(Reighardiida)
2. 1. 범갑각류 내 계통
범갑각류 내에서 오구동물(Pentastomida, 설구동물)의 정확한 계통학적 위치는 오랫동안 논쟁의 대상이 되어 왔다. 범갑각류는 크게 빈갑류(Oligostraca)와 나머지 그룹으로 나뉘며, 이 나머지는 다시 다갑각류(Multicrustacea)와 Allotriocarida로 분류된다. 다갑각류는 초갑류(Thecostraca), 탄툴라쿠스류(Tantulocarida), 요각류(Copepoda), 연갑류(Malacostraca) 등을 포함한다. Allotriocarida는 두판류(Cephalocarida), 새각류(Branchiopoda), 요지류(Remipedia), 그리고 육각류(Hexapoda, 곤충 포함)를 아우른다.[61]역사적으로 오구동물은 다양한 기생충 그룹과 비교되었으며, 절지동물과 유사한 큐티클 구조 때문에 특히 혹응애(Eriophyidae)와의 유사성이 주목받았다.[11] 이후 연구에서는 지네나 노래기 같은 다지류, 유조동물, 완보동물(물곰)과의 유사성도 언급되었다. 일부 학자들은 오구동물을 환형동물과 절지동물 사이의 중간 형태로 보거나, 별도의 동물 문으로 분류해야 한다고 주장하기도 했다.[46] 오구동물은 탈피를 통해 성장하므로 탈피동물에 속하며, 유충 형태가 절지동물과 유사하다는 점도 확인되었다.[12] 과거에는 유조동물, 완보동물 등과 함께 측절지동물로 묶이기도 했으나[54], 이러한 분류는 제한적인 외부 또는 내부 구조 유사성에 기반한 것이라는 비판도 있었다.[55] 고생물학 분야에서는 엽족동물에서 파생되었다는 설도 제기되었으나 널리 받아들여지지는 않았다.[56][57]
현재 오구동물의 분류에 대해서는 크게 두 가지 해석이 존재한다. 첫째는 오구동물이 고도로 변형된 기생성 갑각류이며, 특히 새미류(물벼룩류)와 가깝다는 것이다. 이 주장은 피에르 조제프 반 베네덴이 오구동물을 기생성 요각류와 비교하면서 시작되었고,[13] 칼 게오르크 윙스트란드가 오구동물과 새미류 사이의 정자 구조 유사성을 발견하면서 더욱 구체화되었다.[14] 존 라일리 등도 오구동물을 갑각류에 포함시키는 상세한 근거를 제시했다.[15] 이후 로렌스 G. 에이블 등의 분자 계통 분석 연구[16][58] 와 후속 연구들[17][18][19][59][60][43] 은 이러한 결과를 뒷받침하며, 오구동물과 새미류를 묶어 '''어류흡충류'''(Ichthyostraca)라는 분류군이 제안되었다.[20][59] 정자 구조 등 형태학적 증거 역시 새미류와의 근연성을 지지하여[54], 현재 많은 갑각류 관련 문헌과 데이터베이스는 오구동물을 갑각류의 일원으로 포함하고 있다.[21]
다른 해석으로는 오구동물이 절지동물의 기원에 가까운 고대 절지동물 그룹이라는 주장이 있다. 이 관점의 비판론자들은 오구동물과 그 유충이 갑각류나 진정절지동물의 전형적인 특징을 보이지 않는다고 지적하며, 이들의 극도로 오래된 캄브리아기 기원을 언급하며 줄기군 절지동물로 해석한다.[22] 최근의 형태학적 분석에서는 오구동물을 절지동물 외부에 위치시키며, 선충류, 음경벌레 등과 자매군 관계에 있다고 보기도 한다.[23] 또한, 멸종된 동물인 ''파키벌미스''(Facivermis)가 오구동물과 밀접한 관련이 있을 수 있다는 가능성도 제기되었다.[23]
종합적으로 볼 때, 분자계통학 및 형태학적 증거에 기반하여 오구동물을 갑각류 내의 새미류와 가까운 그룹(어류흡충류)으로 분류하는 견해가 현재 학계에서 가장 널리 받아들여지고 있다.
2. 2. 오구동물 내 계통
오구동물의 계통학적 위치는 오랫동안 논란의 대상이었다. 역사적으로 오구동물은 다양한 기생충 그룹과 비교되었으며, 큐티클 구조가 절지동물과 유사하다는 점이 알려지면서 특히 혹응애와의 유사성이 주목받았다.[11] 혹응애는 오구동물보다 훨씬 작지만, 길쭉한 몸과 두 쌍의 다리를 가진다는 공통점이 있다. 이후 연구에서는 지네나 노래기와 같은 다지류, 유조동물, 완보동물과의 유사성도 제기되었다. 일부 학자들은 오구동물을 환형동물과 절지동물 사이의 중간 형태로 보기도 했고, 독립된 문으로 분류해야 한다고 주장하기도 했다. 오구동물은 탈피를 통해 성장하는데, 이는 이들이 탈피동물에 속함을 시사하며, 유충 형태가 절지동물과 유사하다는 연구 결과도 있다.[12]현재 오구동물의 계통에 대해서는 크게 두 가지 해석이 존재한다. 하나는 오구동물이 고도로 변형된 기생성 갑각류이며 어류 이와 관련이 깊다는 설이고, 다른 하나는 절지동물의 기원에 가까운 고대 절지동물 그룹이라는 설이다.
오구동물은 정자 구조, 난소 및 유생 형태 등에서 절지동물과의 유사성이 오래전부터 지적되었지만, 독특한 형태 때문에 절지동물로 인정받기 어려웠다. 이 때문에 오랫동안 독립된 동물 문(설형동물문)으로 취급되었으며[46], 유조동물, 완보동물 등과 함께 측절지동물로 묶이기도 했다.[54] 일본의 학자 카토(1967)는 이러한 논의에 대해, 과거에 오구동물과 유연성이 주장된 그룹으로 환형동물, 절지동물 내 다지류와 거미류 등을 언급하며, 이러한 주장들이 "단지 2~3개의 외부 또는 내부 구조의 유사성"에 근거한 것이어서 깊이 논할 가치가 부족하다고 지적했다. 그는 오구동물의 계통 파악이 어려운 이유로 이들이 아주 오래전에 기생 생활을 시작하여 퇴행적인 변화가 심하게 나타났기 때문이라고 설명했다.[55] 널리 받아들여지지는 않지만, 고생물학 분야에서는 엽족동물(절지동물, 유조동물, 완보동물의 공통 조상으로 여겨지는 고대 생물 그룹)에서 파생되었다는 설도 제기된 바 있다.[56][57]
그러나 1989년 Abele 연구팀[58]과 1997년 Zrzavý 연구팀[59] 등의 분자계통분석 연구 결과, 오구동물은 절지동물 문의 갑각류 아문에 속하며, 특히 새미류(충류)와 가까운 관계임이 밝혀졌다.[60][43] 이후 정자의 미세 구조 등 형태학적 연구에서도 새미류와의 근연성을 지지하는 증거들이 발견되어, 분자계통학과 형태학 양쪽에서 오구동물과 새미류의 가까운 관계가 확인되었다.[54] 이에 따라 오구동물은 새미류와 함께 갑각류 내의 어류흡충류 (Ichthyostraca)라는 분류군으로 묶이게 되었다.[59]
3. 형태
오구동물은 길이가 1cm에서 14cm에 달하는[46] 벌레 모양 또는 혀 모양의 동물이다. 몸 색깔은 주로 흰색이나 노란색이며, 투명한 경우도 많다. 몸은 키틴질의 표피로 덮여 있으며, 여러 개의 마디로 나뉜 것처럼 보이지만 이는 실제 체절 구조와는 다르다. 외견상 보이는 마디 수는 적게는 20개 미만에서 많게는 100개가 넘으며, 종을 구분하는 특징으로 사용된다.[46]
머리 부분(머리가슴) 앞쪽 끝에는 입이 있으며, 입 뒤 양옆으로 두 쌍의 갈고리가 있다.[46] 이 갈고리는 원래 신축성 있는 부속지였던 것으로 추정된다. 포로케팔루스목(Porcephalida)에서는 이 갈고리가 머리 복면에 직접 붙어 거의 일렬로 배열되어 있으며, 안쪽 쌍이 제1쌍, 바깥쪽 쌍이 제2쌍에 해당한다. 반면, 케파로바에나목(Cephalobaenida)이나 기반적인 화석 종에서는 머리 측면으로 돌출되어 앞뒤로 배열된 형태를 명확히 볼 수 있다. 현생종의 갈고리에는 관절이 없지만, 일부 화석 종에서는 짧은 세 마디로 나뉜 형태가 발견되기도 한다. 머리 부분은 입에서 유래한 선절 1개와 두 쌍의 부속지에서 유래한 체절 2개를 합쳐 최소 3개의 체절로 구성된 것으로 보이나(배아 발생 과정에서는 총 4개의 절이 관찰됨), 다른 절지동물의 머리 부분 체절 및 부속지와의 상동 관계는 아직 불확실하다.[48] 갈고리는 갈색 또는 금색을 띤다.
몸통은 가늘고 길며, 숙주의 폐에 기생하는 종은 원통형, 비강에 기생하는 종은 배 쪽이 납작한 경향이 있다. 발생학 및 고생물학적 증거에 따르면, 오구동물의 몸통은 본래 3개의 체절과 체절 구조가 없는 꼬리 부분으로 구성되었으며, 현생종 몸통의 외견상 많은 마디는 이 꼬리 부분이 길게 발달하고 분절된 결과로 여겨진다. 현생종의 몸통에는 부속지가 없지만, 일부 화석 종에서는 몸통의 두 번째와 세 번째 체절에 해당하는 부분에서 퇴화된 부속지가 발견되기도 한다.[48] 항문은 몸통의 뒤쪽 끝에 열린다.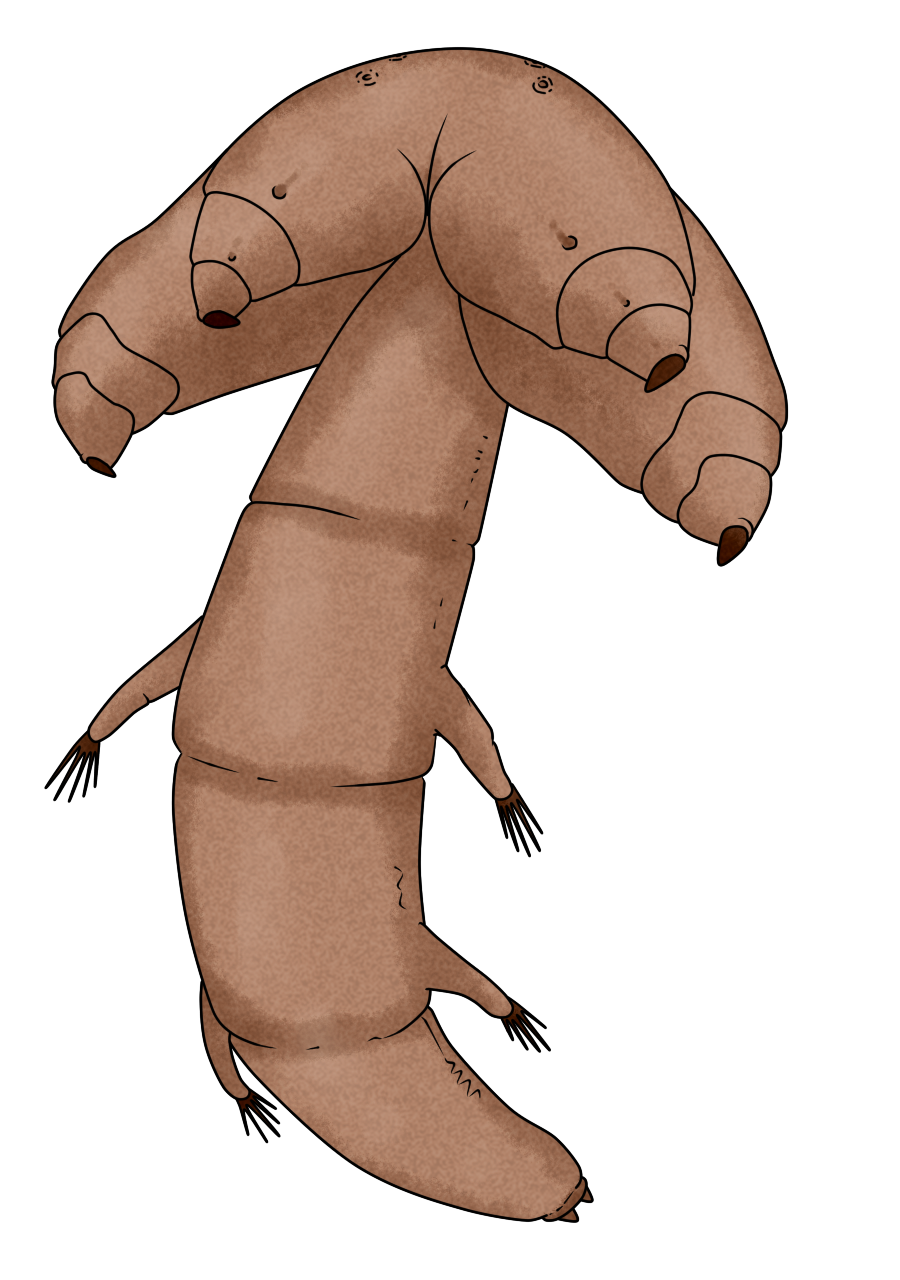
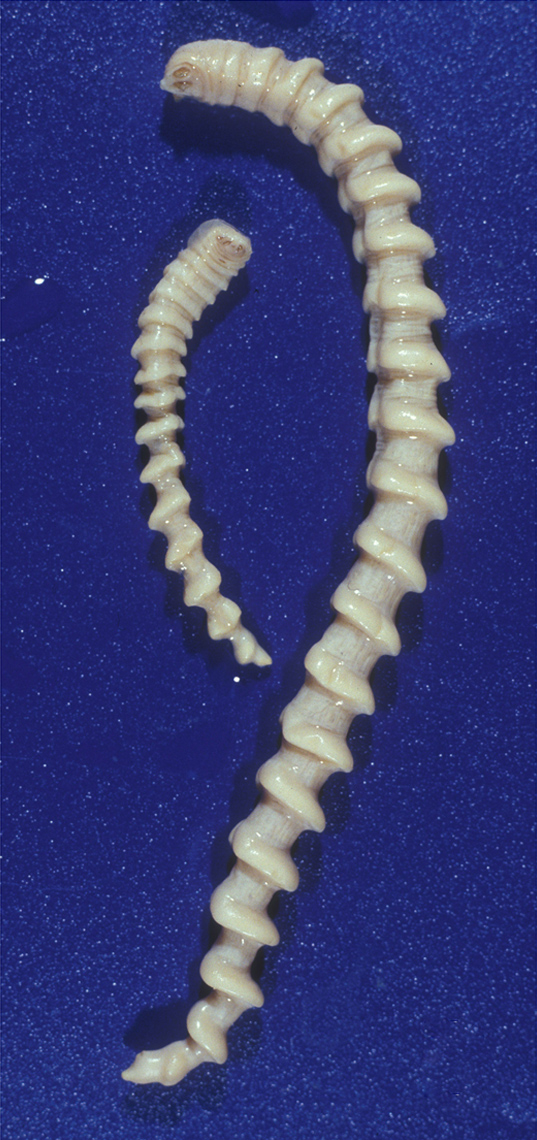
오구동물은 암수가 다른 성적 이형성을 보이며, 일반적으로 암컷이 수컷보다 크다. 생식 구멍은 암수 모두 몸통 앞쪽의 배 면에 열린다. 일부 종의 수컷은 생식 구멍 주변에 한 쌍의 침 모양 구조를 가지고 있다.[49][50]
3. 1. 내부 형태
오구동물은 길이가 1cm에서 14cm에 달하는 벌레 모양의 동물이다. 암컷이 수컷보다 크다. 몸은 분절되어 있으며 키틴질의 표피로 덮여 있다.소화관은 단순한 관 형태로, 주로 숙주의 혈액을 먹기 때문에 이러한 구조를 가진다. 다만, Linguatula 속은 육식성 포유류의 비강에 서식하며 주로 점액과 죽은 세포를 먹고,[30][31] 입은 근육질 펌프 형태로 약간 변형되어 액체를 빨아들이기 좋다.[32] 소화관은 입에서 항문까지 직선으로 이어지며, 전장, 중장, 후장으로 나눌 수 있다. 전장과 후장은 키틴질로 덮여 있고 짧으며, 소화관의 대부분은 중장이 차지한다.
신경계는 각 분절마다 신경절이 있는 복신경삭을 포함하며, 다른 절지동물의 신경계와 유사하다. 기본적으로 소화관 앞부분을 감싸는 뇌신경절과 여기서 몸의 배 쪽으로 뻗어 나가는 복신경삭으로 구성된다. 복신경삭은 세로로 틈이 있어, 기본적으로 사다리 모양 신경계에서 유래했음을 보여준다. 복신경삭에는 대략 3개의 신경절이 구분된다. 그러나 신경계는 퇴화하는 경향이 강하며, 특히 포로케팔리스목(Pentastomida)에서 이러한 경향이 뚜렷하다. 이 목에서는 복신경삭이 거의 퇴화하여 소화관 주변의 신경 덩어리만 남은 형태로 나타나기도 한다.
몸에는 혈강이 있지만, 순환계, 호흡계, 배설계는 없다.[32] 체강은 넓으며, 소화관에서 체벽으로 이어지는 결합 조직에 의해 나뉘어 배강과 복강으로 구분된다.
4. 생태
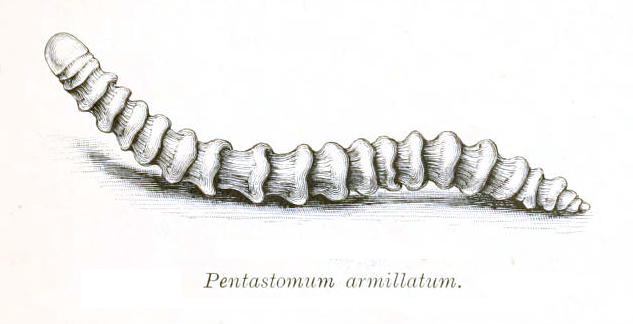
오구동물은 파충류, 조류, 포유류의 상부 호흡기에 서식하며 알을 낳는다. 자웅이체이며 내부 수정을 한다. 알은 숙주의 기침을 통해 배출되거나 소화계를 거쳐 몸 밖으로 나온다. 이 알은 일반적으로 어류나 작은 초식 포유류인 중간 숙주에게 섭취된다.[32]
유충은 중간 숙주 안에서 부화하여 장벽을 뚫고 나온 뒤, 중간 숙주의 몸 안에 낭종을 형성한다. 유충은 처음에는 둥근 모양에 4개 또는 6개의 짧은 다리를 가지지만, 여러 번의 탈피를 거쳐 성충의 형태를 갖추게 된다. 최소한 한 종인 ''Subtriquetra subtriquetra''는 자유 생활을 하는 유충 단계를 가진다.[33] 님프 단계를 포함하는 간접 발달과 직접 발달 방식이 모두 존재한다. 오구동물은 중간 숙주가 최종 숙주에게 먹혔을 때, 식도를 통해 호흡기로 이동하여 최종 숙주에 도달한다.[32][34]
모든 오구동물은 기생충으로 간주된다. 현존하는 종들은 주로 육상 척추동물, 특히 사지동물의 폐나 비강에 내부 기생하지만, 어류나 곤충을 숙주로 삼는 경우도 있다.[51]
알려진 화석종들은 모두 해양성이었다. 이들이 살았던 고생대 캄브리아기부터 실루리아기[52]에는 사지동물이 아직 번성하지 않았기 때문에, 당시의 다른 해양 생물에 기생했을 것으로 추정된다. 일부 화석종은 갯지렁이류에 외부 기생했던 것이 확인되었다.[53]
4. 1. 인간 감염

설충류는 때때로 인간에게 기생한다.[35] ''Sebekia'' 속이 피부염을 유발한다는 보고가 있지만,[36][37] 인간 내부 기생충 감염을 주로 일으키는 것은 ''Linguatula''와 ''Armillifer'' 두 속이다. 내장 설충증은 ''Linguatula serrata'', ''Armillifer armillatus'', ''Armillifer moniliformis'', ''Armillifer grandis'', 그리고 ''Porocephalus crotali''에 의해 발생할 수 있다.[38]

감염과 관련된 용어는 다음과 같다.
- ''Linguatula''에 의한 질병: 설충증 또는 설충병이라고 불린다.
- ''Porocephalus''에 의한 질병: 포로세팔리아증 또는 포로세팔로시스라고 불린다.
- ''Armillifer''에 의한 질병: 역시 포로세팔로시스라고 불린다.[39] (''Armillifer moniliformis''의 다른 이름은 ''Porocephalus moniliformis''이다.)[40]
- 설충류 감염 통칭: "설충증"
''Porocephalus''와 ''Armillifer''(둘 다 원통형이며 뱀에 서식)는 ''Linguatula''(납작하며 개와 늑대에 서식)보다 서로 더 유사하다.
5. 역사
역사적으로 혀벌레(오구동물)의 생물학 및 계통 분류에 대한 중요한 기록에는 요제프 알로이스 프뢸리히,[2] 알렉산더 폰 훔볼트,[3] 카를 아스문트 루돌피,[4] 카를 모리츠 디에징,[5] 루돌프 로이카르트[6] 등의 초기 연구가 포함된다.
루이스 웨스턴라 삼본,[7] 리하르트 하이몬스,[8] 존 라일리[9] 등이 중요한 내용을 요약하여 발표했으며, 1969년까지의 참고 문헌과 함께 오구동물의 진화적 관계를 검토한 연구는 J. T. 셀프에 의해 출판되었다.[10]
오구동물의 유연관계는 오랫동안 논쟁의 대상이 되어 왔다. 역사적으로 이들은 처음에는 다양한 기생충 그룹과 비교되었다. 이후 오구동물의 큐티클이 절지동물과 유사하다는 점이 알려지면서, 특히 혹응애(Eriophyidae)와의 유사점이 발견되었다.[11] 혹응애는 오구동물보다 훨씬 작지만, 길쭉한 체절형 몸체와 두 쌍의 다리를 가진다는 공통점이 있다. 이후의 연구에서는 지네와 노래기(Myriapoda), 벨벳벌레(Onychophora), 물곰(Tardigrada)과의 유사성이 언급되기도 했다. 일부 연구자들은 오구동물을 환형동물과 절지동물의 중간 형태로 보기도 했고, 다른 연구자들은 이들을 별도의 문으로 분류해야 한다고 주장했다. 오구동물은 탈피를 통해 성장하는데, 이는 이들이 탈피동물에 속한다는 점을 시사하며, 다른 연구에서는 이들의 유충이 절지동물과 유사하다는 점을 확인했다.[12]
현재는 일반적으로 두 가지 해석이 존재한다. 하나는 오구동물이 고도로 변형된 기생 갑각류이며, 어류 이와 관련이 있을 가능성이 높다는 것이다. 다른 하나는 오구동물이 절지동물의 기원에 가까운 고대 절지동물 그룹이라는 해석이다.
6. 논란
오구동물이 생물 분류 체계에서 어디에 속하는지는 오랫동안 학자들 사이에서 논쟁거리였다. 역사적으로 이들의 분류에 대한 중요한 연구는 요제프 알로이스 프뢸리히,[2] 알렉산더 폰 훔볼트,[3] 카를 아스문트 루돌피,[4] 카를 모리츠 디에징,[5] 루돌프 로이카르트[6] 등에 의해 초기 연구가 이루어졌다. 이후 루이스 웨스턴라 삼본,[7] 리하르트 하이몬스,[8] 존 라일리[9] 등이 중요한 내용을 요약 발표했으며, 1969년까지의 연구 문헌과 진화적 관계에 대한 검토는 J. T. 셀프가 출판했다.[10]
처음에는 오구동물을 다양한 기생충 그룹과 비교했다. 하지만 몸 표면의 단단한 껍질(큐티클)이 절지동물과 비슷하다는 점이 알려지면서, 특히 혹응애와의 유사점이 주목받았다.[11] 혹응애는 오구동물보다 훨씬 작지만, 길고 마디로 나뉜 몸과 두 쌍의 다리를 가졌다는 공통점이 있다. 이후 연구에서는 지네나 노래기, 벨벳벌레, 물곰과의 유사성도 언급되었다. 일부 학자들은 오구동물을 환형동물과 절지동물 사이의 중간 형태로 보기도 했고, 아예 독립적인 문으로 분류해야 한다고 주장하기도 했다. 오구동물은 탈피를 통해 성장하는데, 이는 이들이 탈피동물에 속한다는 것을 보여준다. 다른 연구에서는 오구동물의 유충이 절지동물과 비슷하다는 점을 확인했다.[12]
현재 오구동물의 분류에 대해서는 크게 두 가지 해석이 존재한다.
첫 번째는 오구동물이 고도로 변형된 기생성 갑각류이며, 특히 어류에 기생하는 물벼룩류와 가깝다는 설이다. 이러한 주장은 피에르 조제프 반 베네덴이 오구동물을 기생성 요각류와 비교하면서 시작되었다.[13] 이후 칼 게오르크 윙스트란드는 정자의 형태를 연구하여 오구동물과 물벼룩류의 정자 구조가 유사하다는 점을 발견했다.[14] 물벼룩류는 어류나 때로는 양서류에 기생하는 만각강 갑각류의 일종이다. 존 라일리와 동료들도 오구동물을 갑각류에 포함시키는 상세한 근거를 제시했다.[15] 로렌스 G. 에이블과 동료들의 DNA 등 분자 정보를 이용한 연구는 이 가설을 더욱 강력하게 뒷받침했다.[16] 이후 여러 분자 계통 연구에서도 비슷한 결과가 나왔으며,[17][18][19] 오구동물과 새예동물을 묶어 Ichthyostraca라는 분류군을 제안하기도 했다.[20] 이러한 연구 결과에 따라, 현재 갑각류에 대한 여러 주요 연구 서적과 데이터베이스에서는 오구동물을 갑각류의 일원으로 포함하고 있다.[21]
두 번째 해석은 오구동물이 절지동물의 기원에 가까운 고대의 절지동물 그룹이라는 설이다. 이 주장을 지지하는 학자들은 기생성 갑각류조차도 유충 단계에서는 갑각류의 특징을 보이는데, 오구동물의 유충은 갑각류나 심지어 진정절지동물의 전형적인 특징을 보이지 않는다는 점을 지적한다. 이들은 오구동물이 매우 오래된 캄브리아기에 기원했을 가능성을 언급하며, 이들을 절지동물의 초기 조상에 가까운 그룹(줄기군 절지동물)으로 해석한다.[22] 최근의 형태학적 분석에서는 오구동물을 선충류, 음경벌레 및 유사한 탈피동물 '벌레' 그룹을 포함하는 분류군과 자매군 관계로 보아, 절지동물 밖에 위치시킨다는 결과를 내놓았다.[23] 또한, 화석 자료를 추가하여 ''파키벌미스''(Facivermis)라는 멸종 동물이 오구동물과 가까운 관계일 수 있다고 제안했다. 하지만 이 연구가 오구동물과 갑각류의 관계를 직접적으로 검토하지는 않았다는 점은 유의해야 한다.
7. 화석 기록
뛰어난 보존 상태를 보이는, 3차원적이며 인산염화된 오구동물 화석이 스웨덴 상부 캄브리아기 오르스텐(Orsten) 라거슈테테[24]와 캐나다 캄브리아기/오르도비스기 경계 지층[25], 그리고 그린란드의 울루이안기(캄브리아기 중기) 지층[26]에서 발견되었다. 캄브리아기에서는 지금까지 ''Aengapentastomum'', ''Boeckelericambria'', ''Dietericambria'', ''Haffnericambria'', ''Heymonsicambria'' 등 5개의 화석 속이 확인되었다. 이러한 화석들은 오구동물이 매우 초기에 진화했음을 시사하며, 이 동물들이 당시 기생충이었는지, 만약 그렇다면 어떤 숙주에 기생했는지에 대한 의문을 제기한다. 이빨고기류(원시 어류)가 가능한 숙주로 언급되기도 한다.[25]
또 다른 속인 ''Invavita''는 영국의 실루리아기 해양 지층에서 발견되었다. ''Invavita'' 화석 표본은 ''Nymphatelina gravida''라는 종의 새엽류 숙주에 단단히 부착된 상태로 발견되었다.[27][28] 이 종은 머리, 벌레 모양의 몸통, 그리고 두 쌍의 다리를 가지고 있었다.[29]
참조
[1]
논문
Phylogenetic position of the Pentastomida and (pan)crustacean relationships
2004-03-07
[2]
논문
Beschreibung einiger neuer Eingeweidewürmer
[3]
서적
Voyage de Humboldt et Bonpand 2
Ptie. F. Schoell et G. Dufour, Paris
[4]
서적
Entozoorum Synopsis
Augustus Rücker Berlin
[5]
논문
Versuch einer Monographie der Gattung ''Pentastoma''
[6]
논문
Bau und Entwicklungsgeschichte der Pentastomen nach Untersuchungen besonders von ''Pent. taenoides'' und ''P. denticulatum''
C. F. Winter'sche Verlagshandlung, Leipzig
[7]
논문
A synopsis of the family Linguatulidae
[8]
서적
Klassen und Ordnungen des Tierreichs. Fünfter Band. IV Abteilung, 1. Buch
Akademische Verlagsgesellschaft m.b.H.
[9]
논문
The biology of pentastomids
[10]
논문
Biological relationships of the Pentastomida: a bibliography on the Pentastomida
[11]
논문
Ueber die Entwicklung des ''Pentastoma taenioides''
[12]
논문
"Arthropodencharaktere" bei einem Pentastomiden Embryo (''Reighhardia sernae'')
[13]
논문
Recherches sur l'organisation et le développement des Lingatules (Pentastoma Rud.), suivies de la description d'une espèce nouvelle provenant d'un Mandrill
[14]
논문
Comparative spermatology of a pentastomid, ''Raillietiella hemidactyli'', and a branchiuran crustacean, ''Argulus foliaceus'', with a discussion of pentastomid relationships
[15]
논문
The phylogenetic relationships of the Pentastomida: the case for their inclusion within the Crustacea
[16]
논문
Molecular evidence for inclusion of the Phylum Pentastomida in the Crustacea
http://mbe.oxfordjou[...]
[17]
논문
Phylogenetic position of the Pentastomida and (pan)crustacean relationships
http://www.eeob.iast[...]
2009-12-20
[18]
논문
First maxillae suction discs in Branchiura (Crustacea): development and evolution in light of the first molecular phylogeny of Branchiura, Pentastomida, and other "Maxillopoda"
[19]
논문
Phylotranscriptomics to bring the understudied into the fold: monophyletic Ostracoda, fossil placement, and Pancrustacean phylogeny
http://pdxscholar.li[...]
[20]
논문
The interrelationships of metazoan parasites: a review of phylum- and higher-level hypotheses from recent morphological and molecular phylogenetic analyses
[21]
서적
An Updated Classification of the Recent Crustacea
http://atiniui.nhm.o[...]
Natural History Museum of Los Angeles County
2009-12-20
[22]
논문
A new Late Cambrian pentastomid and a review of the relationships of this parasitic group
[23]
논문
Morphological support for the phylogenetic positioning of Pentastomida and related fossils
[24]
논문
Pentastomid parasites from the Lower Palaeozoic of Sweden
[25]
논문
An exceptionally preserved parasitic arthropod, ''Heymonsicambria taylori'' n. sp. (Arthropoda increate sedis: Pentastomida) from Cambrian – Ordovician boundary beds of Newfoundland
[26]
논문
The oldest tongue worm: A stem-group pentastomid arthropod from the early middle Cambrian (Wuliuan Stage) of North Greenland (Laurentia)
[27]
웹사이트
A 425-million-year-old parasite found attached to host
https://www.bbc.co.u[...]
2015-05-22
[28]
논문
A 425-Million-Year-Old Silurian Pentastomid Parasitic on Ostracods
[29]
웹사이트
Requiem for an ancient tongue worm
http://news.yale.edu[...]
2015-05-21
[30]
서적
Encyclopedic Reference of Parasitology: Biology, Structure, Function
https://books.google[...]
Springer
2001-05-21
[31]
논문
Characterization of tongue worms, Linguatula SPP. (Pentastomida) in Romania, with the first record of an unknown adult Linguatula from roe deer (Capreolus capreolus Linnaeus)
2022
[32]
서적
Invertebrate Zoology
Holt-Saunders International
[33]
서적
Parasitic Crustacea
2019
[34]
서적
Atlas of Crustacean Larvae
https://books.google[...]
JHU Press
2014-07
[35]
논문
The Pentastomida parasitic in man
[36]
논문
Human dermatitis caused by a nymph of ''Sebekia''
[37]
문서
Correct spelling: Sebakia --> Sebekia, See {{cite WoRMS |id=384727 |title=''Sebekia'' Sambon, 1922 }}
[38]
논문
Diagnosis of human visceral pentastomiasis
[39]
논문
Roentgen diagnosis of ''Armillifer armillatus'' infestation (porocephalosis) in man
1976-05
[40]
서적
Imaging of tropical diseases: with epidemiological, pathological, and clinical correlation
https://books.google[...]
Birkhäuser
2010-04-19
[41]
서적
甲殻亜門分類表
裳華房
[42]
논문
顎脚類(甲殻類)の分類と系統に関する研究の最近の動向
日本動物分類学会
[43]
논문
Phylotranscriptomics to Bring the Understudied into the Fold: Monophyletic Ostracoda, Fossil Placement, and Pancrustacean Phylogeny
https://academic.oup[...]
2012-09-12
[44]
논문
The Phylogeny and Evolutionary History of Arthropods
https://www.scienced[...]
2019-06-17
[45]
웹사이트
Pentastoma
https://medical-dict[...]
[46]
서적
無脊椎動物の多様性と系統
裳華房
[47]
논문
舌形動物および舌虫症に関する最近の知見――特に酪農学園大学野生動物医学センターWAMCで扱われた事例を中心に
酪農学園大学
[48]
논문
Pentastomid parasites from the Lower Palaeozoic of Sweden
https://www.cambridg[...]
1994-01
[49]
논문
Using Combined Morphological, Allometric and Molecular Approaches to Identify Species of the Genus Raillietiella (Pentastomida)
https://journals.plo[...]
2011-09-20
[50]
논문
The oldest tongue worm: a stem-group pentastomid arthropod from the early middle Cambrian (Wuliuan Stage) of North Greenland (Laurentia)
https://www.tandfonl[...]
2022-04-03
[51]
논문
A systematic monograph of the Recent Pentastomida, with a compilation of their hosts
https://repository.n[...]
2013-01-01
[52]
간행물
The Fossil Record of the Pancrustacea
https://doi.org/10.1[...]
Oxford University Press
2024-01-12
[53]
논문
A 425-Million-Year-Old Silurian Pentastomid Parasitic on Ostracods
https://doi.org/10.1[...]
2015-06
[54]
서적
無脊椎動物の多様性と系統
裳華房
[55]
서적
舌形動物門
中山書店
[56]
논문
Advances, diversions, possible relapses and additional problems in understanding the early evolution of the Articulata
https://www.tandfonl[...]
1998-01
[57]
논문
The xenusian-to-anomalocaridid transition within the lobopodians
http://paleoitalia.o[...]
[58]
논문
Molecular evidence for inclusion of the phylum pentastomida in the crustacea
https://doi.org/10.1[...]
Society for Molecular Biology and Evolution
[59]
간행물
Arthropod phylogeny: taxonomic congruence, total evidence and conditional combination approaches to morphological and molecular data sets
Chapman & Hall, London
[60]
논문
Arthropod relationships revealed by phylogenomic analysis of nuclear protein-coding sequences
https://www.nature.c[...]
2010-02
[61]
논문
Pancrustacean Evolution Illuminated by Taxon-Rich Genomic-Scale Data Sets with an Expanded Remipede Sampling
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com