하플로그룹 D (Y-DNA)
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
하플로그룹 D (Y-DNA)는 이전에는 D-M174로 불렸으며, Y 염색체 상의 유전적 특징에 따라 분류되는 하플로그룹 중 하나이다. 하플로그룹 DE에서 갈라져 나왔으며, D-M174는 아시아에서 주로 발견되었으나, 2019년 나이지리아에서 D2 (D-A5580.2)가 발견되면서 아프리카 기원설이 제기되었다. D1, D2로 하위 분류되며, D1은 다시 D1a와 D1b로 나뉜다. 현재 일본, 티베트, 안다만 제도, 중국 남부 등 동아시아와 동남아시아 지역에 주로 분포하며, D2는 아프리카, 서아시아에서도 발견된다.

더 읽어볼만한 페이지
- 인류 Y-DNA 하플로그룹 - 하플로그룹 K (Y-DNA)
하플로그룹 K는 약 47,000~50,000년 전 중동 또는 중앙아시아에서 기원한 것으로 추정되는 Y-염색체 DNA 하플로그룹으로, 하위 계통인 L, T, NO, M, S, P 등을 통해 유라시아, 오세아니아, 아메리카 대륙 등 전 세계에 분포한다. - 인류 Y-DNA 하플로그룹 - Y염색체 아담
Y염색체 아담은 현존하는 모든 인류의 부계 혈통을 따라 올라갈 때 만나는 가장 최근의 공통 조상을 지칭하는 비유적인 용어이며, 단 한 명의 남성이 아닌 Y염색체에 돌연변이가 발생하기 이전의 남성 집단을 의미한다.

2. 기원 및 확산
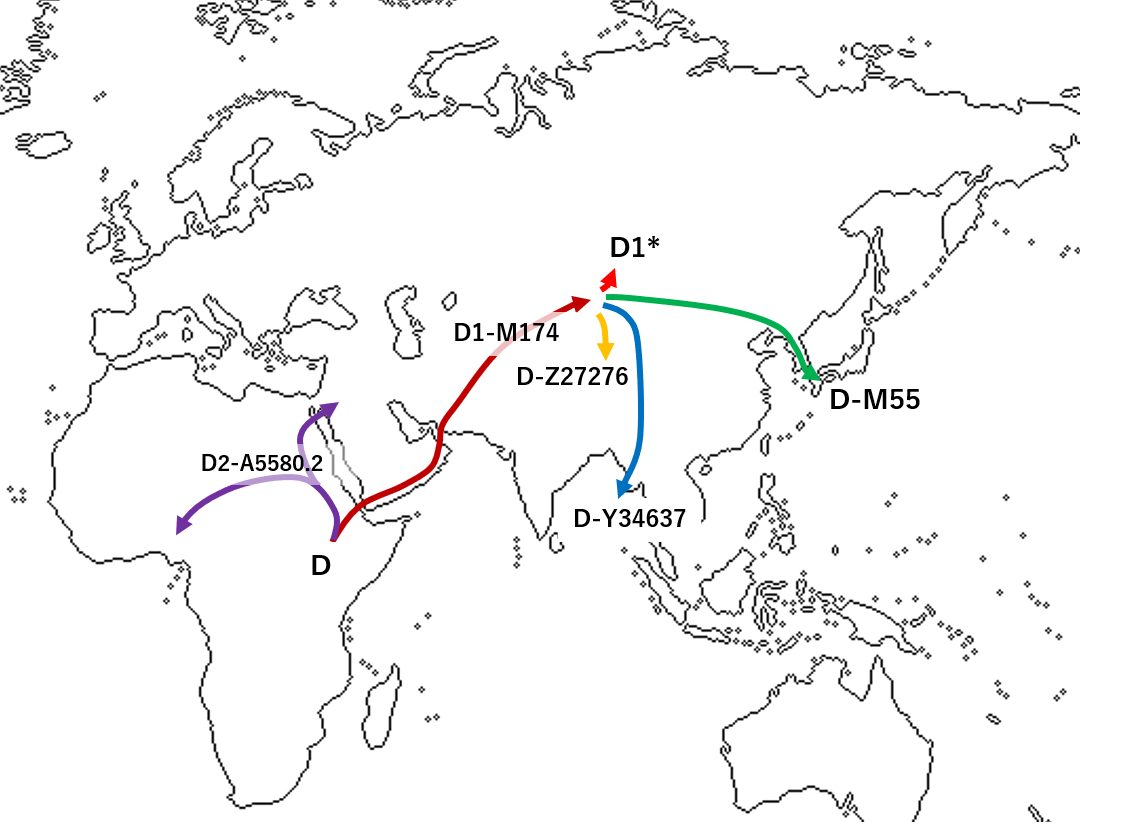
하플로그룹 D는 이전에는 D 계통 D-M174의 명칭이었다. D의 상위 그룹인 하플로그룹 DE의 기원에 대해서는 아프리카 기원설[2]과 아시아 기원설[3] 등 다양한 주장이 존재했으나, 최근 연구 결과 아프리카 기원설이 유력해졌다.
2019년 하버 외(Haber et al.)는 나이지리아인 3명에게서 "D0"(이후 D2로 명명)으로 명명된 하플로그룹을 확인했다. SNP A5580.2로 정의되는 "D0" 하플로그룹은 M174 밖에 있지만, D 계통에 속하며, E에는 없는 7개의 SNP를 D-M174와 공유하며, D 분기에서 초기에 갈라진 것으로 확인되었다.[1]
하버 외는 "D0" 하플로그룹의 깊은 뿌리와 D0와 DE의 초기 분기 시간(약 76,000~71,000년 전)을 근거로 D0와 DE, 그리고 D0와 D-M174의 공통 조상(현재 D-CTS3946 또는 "D"로 알려짐)의 아프리카 기원을 지지했다. DE*, E, D0의 분기 시간을 모두 약 76,000~71,000년 전으로 추정하며, 현대 유라시아인의 조상이 아프리카에서 탈출한 시기(이후 네안데르탈인과의 혼혈)를 약 50,300~59,400년 전으로 추정하여, 아프리카 기원설을 뒷받침했다.[1]
D-CTS3946은 아프리카 내부와 외부 모두에 퍼진 것으로 추정된다. 한 분기는 아프리카에서 D0(이후 D2로 명명)로 분기되었고, 다른 분기는 아프리카를 떠나 결국 D-M174(이후 D1으로 명명)로 분기되었다. 즉, M174 돌연변이는 나중에 아시아로 퍼진 D-CTS3946에서 발생했다.
할라스트 외 (2020)는 하플로그룹 CT (DE의 조상) 내에서의 초기 분기가 아프리카에서 발생했다고 주장하며, 비아프리카 Y 염색체의 계통 지리학적 분석 결과는 기저 하플로그룹 D 및 기타 기저 Y 계통의 아프리카로부터의 초기 이주(70,000~55,000년 전) 이후, 55,000~50,000년 전에 알려진 모든 생존하는 비아프리카 남성 계통(최근 이주민 제외)의 기원이 동/동남아시아임을 보여준다고 주장한다.
결론적으로, 하플로그룹 D는 약 7만 3천 년 전 아프리카에서 하플로그룹 DE에서 발생했으며, 하위 계통인 D1만이 아프리카를 벗어나 동아시아로 향하게 되었다.[38]
2. 1. 아프리카 기원설
하플로그룹 D는 이전에는 D 계통 D-M174의 명칭이었다. D의 상위 그룹인 하플로그룹 DE의 기원에 대해서는 아프리카 기원설[2]과 아시아 기원설[3] 등 다양한 주장이 존재한다.
2019년 하버 외(Haber et al.)는 나이지리아인 3명에게서 "D0"(이후 D2로 변경)로 명명된 하플로그룹을 확인했다.[1] SNP A5580.2로 정의되는 "D0" 하플로그룹은 M174 밖에 있지만, D 계통에 속하며, E에는 없는 7개의 SNP를 D-M174와 공유하며, D 분기에서 초기에 갈라진 것으로 확인되었다.
하버 외는 아프리카 기원과 아시아 기원을 포함한 여러 가능성을 고려했지만, "D0" 하플로그룹의 깊은 뿌리와 D0와 DE의 초기 분기 시간(약 76,000~71,000년 전)을 근거로 D0와 DE, 그리고 D0와 D-M174의 공통 조상(현재 D-CTS3946 또는 "D"로 알려짐)의 아프리카 기원을 지지했다.[1] D0는 D/E 분기 근처, D 분기에 있는 DE 계통의 분기로, 약 71,000년 전에 분기되었다.
하버 외는 DE*, E, D0의 분기 시간을 모두 약 76,000~71,000년 전으로 추정하며, 현대 유라시아인의 조상이 아프리카에서 탈출한 시기(이후 네안데르탈인과의 혼혈)를 약 50,300~59,400년 전으로 추정하며, 이는 해당 하플로그룹의 아프리카 기원을 뒷받침한다고 주장한다.[1]
따라서 D-CTS3946은 아프리카 내부와 외부 모두에 퍼진 것으로 추정된다. 한 분기는 아프리카에서 D0로 분기되었고, 다른 분기는 아프리카를 떠나 결국 D-M174로 분기되었다(즉, M174 돌연변이는 나중에 아시아로 퍼진 D-CTS3946에서 발생).
2019년 연구(Haber et al. 2019)[29]에 따르면, 기존 DE의 하위 그룹으로 D 계통에도 E 계통에도 속하지 않는 파라그룹 DE* (어느 계통에 가까운지 미상)로 여겨졌던 나이지리아인 3개 샘플[35]이 하플로그룹 E보다 하플로그룹 D에 더 가깝다는 것이 밝혀졌다. 이 샘플은 하플로그룹 E가 갖지 않는 SNP를 하플로그룹 D와 7개 공유하며, "D2"로 하플로그룹 D에 편입되었다.
이 발견으로 기존 하플로그룹 D (M174 변이로 정의)는 하플로그룹 D1으로 명칭이 변경되었고, 새롭게 발견된 D2 (D-A5580.2)와 하플로그룹 D1 (M174)을 합하여 하플로그룹 D는 CT3946 변이에 의해 정의되는 그룹으로 확대되었다. 이 D2는 서아시아 (사우디아라비아, 시리아)에서도 발견되었다[36]。
이처럼 하플로그룹 D2가 아프리카에서 발견되었기 때문에 하플로그룹 D (CT3946)는 아프리카에서 이미 탄생했다고 추정된다[37]。
따라서 하플로그룹 D는 약 7만 3천 년 전 아프리카[29]에서 하플로그룹 DE에서 발생, 하위 계통인 하플로그룹 D1만이 아프리카를 벗어나 내륙 루트를 통해 동아시아로 향하게 되었다[38].(D2 발견 이전에는 하플로그룹 DE와 D가 아시아에서 발상했다는 이설[39]도 있었다.)
할라스트 외 (2020)는 하플로그룹 CT (DE의 조상) 내에서의 초기 분기가 아프리카에서 발생했다고 주장한다. 또한 비아프리카 Y 염색체의 계통 지리학적 분석 결과는 모두 기저 하플로그룹 D 및 기타 기저 Y 계통의 아프리카로부터의 초기 이주(70,000~55,000년 전) 이후, 55,000~50,000년 전에 알려진 모든 생존하는 비아프리카 남성 계통(최근 이주민 제외)의 기원이 동/동남아시아임을 보여준다고 주장한다.
2. 2. 아시아 기원설 (반박됨)
2019년 연구(Haber et al. 2019)[29]에 따르면, 나이지리아인 3개 샘플[35]이 하플로그룹 E보다 하플로그룹 D에 더 가깝다는 것이 밝혀졌다. 이 샘플은 "D2"로 명명되었고, 하플로그룹 D에 편입되었다.이 발견으로 기존 하플로그룹 D (M174 변이)는 하플로그룹 D1으로 명칭이 변경되었고, D2 (D-A5580.2)와 하플로그룹 D1 (M174)을 합하여 하플로그룹 D는 CT3946 변이에 의해 정의되는 그룹으로 확대되었다. 이 D2는 사우디아라비아, 시리아에서도 발견되었다[36]。
이처럼 하플로그룹 D2가 아프리카에서 발견되었기 때문에, D2 발견 이전에는 하플로그룹 DE와 D가 아시아에서 발상했다는 이설[39]도 있었으나 하플로그룹 D (CT3946)는 아프리카에서 탄생했다고 추정된다[37]。
2. 3. 확산 경로
하플로그룹 D는 이전에 D 계통 D-M174의 명칭이었다. D의 상위 그룹인 하플로그룹 DE의 기원에 대해서는 아프리카 기원설[2]과 아시아 기원설[3] 등 다양한 주장이 존재한다. 그러나 D-M174는 아시아 기원으로 간주되었으며, 일반적으로 아시아에서만 발견된다.
하버 외 (2019)는 나이지리아인 3명에게서 "D0"으로 명명된 하플로그룹을 확인했다. SNP A5580.2로 정의되는 "D0" 하플로그룹은 M174 밖에 있지만, D 계통에 속하며, E에는 없는 7개의 SNP를 D-M174와 공유하며, D 분기에서 초기에 갈라진 것으로 확인되었다. 하버 외 (2019)는 아프리카 기원과 아시아 기원을 포함한 여러 가능성을 고려했지만, "D0" 하플로그룹의 깊은 뿌리와 D0와 그 상위 하플로그룹 DE의 초기 분기 시간(약 71,000년 전)을 근거로 D0와 DE, 그리고 D0와 D-M174의 공통 조상(현재 D-CTS3946 또는 "D"로 알려짐)의 아프리카 기원을 지지하는 결론을 내렸다.[1] 이들은 DE*, E, D0의 분기 시간을 모두 약 76,000~71,000년 전으로 추정하며, 현대 유라시아인의 조상이 아프리카에서 탈출한 시기를 약 50,300~59,400년 전으로 추정하는데, 이는 해당 하플로그룹들의 아프리카 기원을 뒷받침한다고 주장한다.[1]
따라서 D-CTS3946은 아프리카 내부와 외부 모두에 퍼진 것으로 추정된다. 한 분기는 아프리카에서 D0 (D2)로 분기되었고, 다른 분기는 아프리카를 떠나 결국 D-M174 (D1)로 분기되었다(즉, M174 돌연변이는 나중에 아시아로 퍼진 D-CTS3946에서 발생).
할라스트 외 (2020)는 하플로그룹 C, D 및 FT 염기 서열의 계통 발생 분석을 통해, "드문 깊은 뿌리 D0"를 고려할 때 하플로그룹 CT (DE의 조상) 내에서의 초기 분기가 아프리카에서 발생했다고 주장한다. 또한, 비아프리카 Y 염색체의 계통 지리학적 분석 결과는 기저 하플로그룹 D 및 기타 기저 Y 계통이 아프리카로부터 초기 이주(70,000~55,000년 전)한 이후, 55,000~50,000년 전에 동/동남아시아에서 기원했음을 보여준다고 주장한다. 이들은 이 계통들이 유라시아 전역으로 빠르게 확장된 다음, 동남아시아에서 다양해지고, 약 55,000~50,000년 전에 서쪽으로 확장되어 유라시아 내 다른 지역 계통을 대체했으며, 하플로그룹 D (D-M174)가 동아시아 인구 내에서 빠르게 확장되어 약 45,000년 전에 형성된 5개의 다른 분기로 구성된다고 결론 내렸다. 이들은 현재 이 하플로그룹들이 동아시아(동/동남아시아)에서 가장 큰 다양성을 가지고 있으며, 특히 동아시아와 동남아시아의 티베토버마어족 인구에서 가장 높은 다양성이 발견되었다고 밝혔다.[4]
2019년 연구(Haber et al. 2019)[29]에 따르면, 나이지리아인 3개 샘플[35]이 하플로그룹 E보다 하플로그룹 D에 더 가깝다는 것이 밝혀졌다. 이 샘플은 "D2"로 하플로그룹 D에 편입되었다.
이 발견으로 기존 하플로그룹 D (M174 변이)는 하플로그룹 D1, 새롭게 발견된 D2 (D-A5580.2)와 D1 (M174)을 합하여 하플로그룹 D는 CT3946 변이에 의해 정의되는 그룹으로 확대되었다. D2는 서아시아 (사우디아라비아, 시리아)에서도 발견되었다.[36]
하플로그룹 D2가 아프리카에서 발견되었기 때문에 하플로그룹 D (CT3946)는 아프리카에서 탄생했다고 추정된다.[37]
하플로그룹 D는 약 7만 3천 년 전 아프리카[29]에서 DE에서 발생, 하위 계통인 D1만이 아프리카를 벗어나 동아시아로 향하게 되었다.[38] (D2 발견 이전에는 DE와 D가 아시아에서 발상했다는 설[39]도 있었다.)
파라그룹 DE*는 티베트인[33]과 다른 서아프리카 샘플[40]에서도 검출되었지만, 상세한 분석은 이루어지지 않았다.
3. 분포
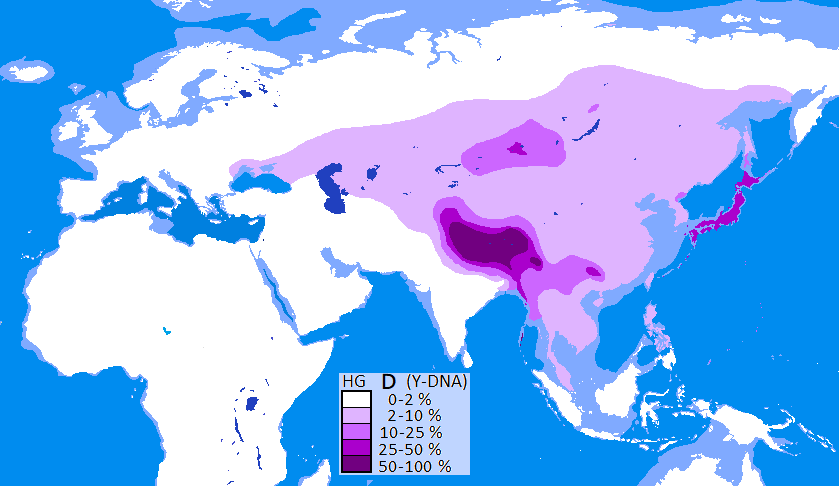
하플로그룹 D는 하플로그룹 DE에서 갈라져 나왔으며, 하플로그룹 E와는 형제 관계이다. DE의 기원에 대해서는 아프리카 기원설[2]과 아시아 기원설[3]이 대립했지만, D-M174는 아시아에서만 주로 발견되어 아시아 기원으로 간주된다.
하버 외 (2019)는 나이지리아인 3명에게서 발견된 "D0" 하플로그룹을 통해 D의 기원을 추적했다. D0는 D-M174와 7개의 SNP를 공유하며, D 분기에서 초기에 갈라진 것으로 확인되었다. 하버 외는 D0, DE, D-CTS3946 (D의 공통 조상)의 아프리카 기원을 지지하며, D0는 약 71,000년 전에 분기되었다고 주장한다. D-CTS3946은 아프리카 내외부로 퍼져나가, 아프리카 내에서는 D0로, 아프리카 외부에서는 D-M174로 분기되었다.[1]
할라스트 외 (2020)는 하플로그룹 CT (DE의 조상) 내 초기 분기가 아프리카에서 발생했다고 주장한다. 또한 비아프리카 Y 염색체의 계통 지리학적 분석 결과, 기저 하플로그룹 D 및 기타 Y 계통은 70,000~55,000년 전 아프리카에서 이주한 후, 55,000~50,000년 전 동/동남아시아에서 기원했다고 본다. 이들은 동남아시아에서 다양해진 후 서쪽으로 확장되었으며, 하플로그룹 D (D-M174)는 동아시아에서 5개의 분기로 빠르게 확장되었다고 결론지었다. 특히 티베토버마어족 인구에서 높은 다양성이 발견된다.[4]
FTDNA는 2019년에 사우디 아라비아와 시리아에서 D2 표본을 추가로 발견했다.[5][10][11] 예멘에서도 시리아 표본과 수천 년의 친족 관계를 가진 D2b-FT51782 표본이 발견되었다.[5][6] 또한 아프리카계 미국인에게서도 D2 표본이 발견되었는데,[8][9] 이 중 일부는 나이지리아인 표본과 한 갈래를 공유한다.[8] D2는 DE 분기에 가깝지만 D 분기에 속하며, M174 돌연변이가 없는 매우 분화된 하플로그룹이다.[10]
하플로그룹 D는 현재 일본 열도, 남서 제도, 안다만 제도, 티베트 고원, 중국 남부, 윈난성 등에서 고빈도로 관찰되며, 그 외 지역에서는 매우 드물게 발견된다. 티베트에서는 D1a1-Z27276, 일본에서는 D1a2a-M55, 안다만 제도에서는 D1a2b-Y34537[31]이 고빈도로 나타난다. 이들은 같은 하플로그룹 D에 속하지만, 하위 그룹이 달라 5만 3000년 이상의 분기 역사를 가진다.[32]
하플로그룹 D는 중국, 조선 및 동남아시아에서 다수인 하플로그룹 O 계통 및 기타 유라시아 계통과는 7만 년 이상 분리된 고립된 계통이다. 동아시아에서 가장 오래된 유형으로 추정되지만,[33] O 계통의 유입으로 인해 섬나라인 일본이나 산악 티베트에만 D 계통이 남았다는 설이 있다.
DE에서 갈라진 하플로그룹 E는 아프리카 대륙에서 고빈도로, 중동이나 지중해 지역에서 중저빈도로 나타난다. 티베트인에게서는 DE*가 극히 소수 발견된다.[34]
3. 1. 동아시아
일본 열도, 남서 제도, 안다만 제도, 티베트 고원, 중국 남부, 윈난성 등에서 하플로그룹 D (Y-DNA)가 높은 빈도로 발견된다. 특히 티베트에서는 D1a1-Z27276이, 일본에서는 D1a2a-M55가 높은 빈도로 나타난다. 이들은 같은 하플로그룹 D에 속하지만, 하위 그룹이 달라 5만 3000년 이상의 분기 역사를 가진다.[32]하플로그룹 D는 현재 중국, 조선(한반도), 동남아시아에서 다수를 차지하는 하플로그룹 O 계통 및 기타 유라시아 계통과는 7만 년 이상 분리된, 매우 고립적인 계통이다. 동아시아에서 가장 오래된 유형으로 추정되지만,[33] O 계통의 유입으로 인해 섬나라인 일본이나 산악 지형인 티베트에 주로 남아있게 되었다는 설이 있다.
중국중국 내 여러 소수민족에게서 하플로그룹 D가 발견된다.
일본일본 열도에서 발견되는 D1a2a 하플로그룹은 D 하플로그룹 중에서도 4만 년 이상 전에 분기된 하위 그룹이다.
- 아이누: 87.5% (D1a2a* 81.25%, D1a2a1a 6.25%)[55]
- 일본 본토 (규슈, 혼슈, 시코쿠): 약 32%[56]- 39%[57] (D1a2a)
- 주고쿠 지방, 시코쿠, 북부 규슈 등 서일본 일부 지역에서는 비교적 적게 나타난다.
- 간토 지방, 도호쿠 지방 등 동일본과 남부 규슈에 많이 분포한다.
- 오키나와
- 오키나와 본섬:
- 이토만시: 47.5% (YAP+ 하플로타입 IIa)[59] (80명 중 38명)
- 구시켄촌: 36.8% (YAP+ 하플로타입 IIa)[59] (19명 중 7명)
- 요미탄촌: 31.3% (YAP+ 하플로타입 IIa)[59] (32명 중 10명)
- 카츠렌초: 27.6% (YAP+ 하플로타입 IIa)[59] (29명 중 8명)
- 오키나와현 전체: 55.6% D-M55 (D-M55(xM116) 4.4%, D-M116(xM125) 31.1%, D-M125(xP42) 15.6%, D-P42 4.4%)[60] (45명 중 25명)
- 야에야마 제도:
- 이리오모테섬: 5.0% (YAP+ 하플로타입)[59] (20명 중 1명)
- 하테루마섬: 0% (YAP+ 하플로타입)[59] (7명 중 0명)
- 오키나와현립 야에야마 고등학교 및 오키나와현립 야에야마 상업고등학교: 32.7% (하플로그룹 DE)[61] (49명 중 16명)
- 미야코섬 오키나와현립 미야코 고등학교: 34.2% (하플로그룹 DE)[29] (38명 중 13명)
- 오키나와 본섬 시마지리군 하에바루정 오키나와현립 카이호 고등학교: 36.1% (하플로그룹 DE)[29] (36명 중 13명)
일본 본토에서는 티베트 등 중국 남서부에 많은 D1a1a-M15도 극소수 발견된다. (일본 1/23 D-M15[58], 미야기현 1/7 D-M15[29], 오사카시 성인 남성 1/241 D-M15[29], 가와사키시 대학생 1/321 D-M15[29], 삿포로시 대학생 1/302 D-M15)[29]
한국한반도에서는 하플로그룹 D 계통이 발견되는데, 이는 근세에 티베트에서 몽골을 경유하여 유입된 D1a1이나, 야요이 시대에 일본 열도에서 한반도로 북상한 D1a2a 계통으로 추정된다.
3. 2. 동남아시아
필리핀에서는 막탄섬에서 5.6%(1/18)[31], 루손섬 중부에서 2.9%(3/102)[31]로 D1b (L1378)가 발견되었다. 안다만 제도에서는 D1a2b (Y34637)[29] 빈도가 높으며, 특히 온간계 온게족(23/23), 자라와족(4/4)에서 100%를 차지하지만, 언어 계통이 다른 대안다만인에서는 0%(0/10)이다.[51]3. 3. 중앙아시아
튀르크계 남부 알타이인(알타이 키지) 표본 120명 중 6명(5.0%)에게서 D-P47이 관찰되었다.[62] 다른 연구에서는 코시 아가치 마을의 남부 알타이인 7명 중 1명(14%), 클라다 마을의 남부 알타이인 46명 중 5명(10.9%), 합계 남부 알타이인 96명 중 6명(6.3%)이 하플로그룹 D-M174(xM15)에 속하는 것으로 나타났다.[63] 중앙아시아의 튀르크계 민족과 몽골계 민족 사이에서도 D-P47과 D-M15가 소수 발견된다.3. 4. 서아시아 및 아프리카
FTDNA는 2019년에 D2의 다른 세 개의 표본을 발견했는데, 사우디 아라비아 서해안의 알 와지에서 2개[10][11], 시리아인(D2b-FT51782)에게서 1개[5][10]가 발견되었다. 또한 D2b-FT51782 표본이 예멘 바이다의 알 키르비에서 발견되었는데[6], 이 표본은 시리아 표본과 수천 년의 친족 관계를 가지고 있는 것으로 밝혀졌다.[5] 인접한 샤브와 주에 성씨가 알려지지 않은 다른 한 사람이 있는데, 이 사람은 최초의 표본과 1700년 전의 TMRCA(최근 공통 조상)를 가진다.[7] 2020년에 FTDNA의 마이클 세이거는 또 다른 표본이 아프리카계 미국인[8]과 다른 아프리카계 미국인에서 발견되었다고 발표했다.[8][9] 시리아 후손과 아프리카계 미국인 중 한 명에게서 발견된 표본은 현재까지 D2의 가장 기저 표본이다.[8][10][11][9] 다른 아프리카계 미국인 표본은 세 명의 나이지리아인과 한 갈래를 공유한다.[8] 하버 등이 제안한 최근 증거에 따르면 D2는 DE 분열에 가깝지만 D 분기에 속하며 다른 알려진 D 계통(그 자매 계통인 D-M174에 속함)이 가진 M174 돌연변이가 없는 매우 분화된 하플로그룹이다.[10]아프리카(나이지리아), 서아시아(시리아, 사우디 아라비아)에서는 하플로그룹 D 중 가장 오래 전에 분기된 계통인 D2가 최근 새롭게 발견되었다.[29][36]
3. 5. 아메리카 대륙
하플로그룹 C와 달리, 하플로그룹 D는 아메리카 대륙(아메리카 원주민)에서는 전혀 발견되지 않는다.[14]4. 하위 계통
하플로그룹 D는 크게 D1과 D2 두 가지 하위 계통으로 나뉜다.
- '''D1 (M174)'''
중앙아시아, 인도차이나반도, 동아시아, 안다만 제도 등에서 주로 발견된다.
- '''D1a (CTS11577)'''
- '''D1a1 (F6251/Z27276)''': 티베트, 러시아, 조지아 등
- '''D1a1a (M15)''': 알타이 공화국, 일본, 중앙아시아, 안다만 제도, 몽골, 중국 대륙, 동남아시아 등
- '''D1a1a1 (F849)'''
- '''D1a1a1a (N1)'''
- '''D1a1a1a1 (Z27269)''': 일본, 중국, 티베트[69]
- '''D1a1a1a1a (PH4979)'''
- '''D1a1a1a1a1 (BY15119/Z29428)'''
- '''D1a1a1a1a2 (F729)'''
- '''D1a1a1a1a2a (F17412, MF10280)''': 티베트인(창두), 대만, 중국(쓰촨), 일본(오사카)
- '''D1a1a1a1a2b (Y62194)'''
- '''D1a1a1a1a2b1 (F17409)''': 티베트(창두), 중국(쓰촨)
- '''D1a1a1a1a2b2 (Y62517)'''
- '''D1a1a1a1a2b2a (F16077)''': 티베트(르카쩌, 산난, 라싸)
- '''D1a1a1a1a2b2b (Y69263)'''
- '''D1a1a1a1a2b2b1 (Y161914)''': 티베트(창두), 중국(저장)
- '''D1a1a1a1a2b2b2''': 우즈베키스탄
- '''D1a1a1a2 (Z31591)''': 티베트(르카쩌, 라싸)
- '''D1a1a2 (F1070)''': 중국(광둥, 장쑤, 광시), 다이족(시솽반나), 싱가포르
- '''D1a1b (P99)''': 티베트, 몽골, 중앙아시아[69], 알타이 공화국, 중국 대륙
- '''D1a2 (Z3660)'''
- '''D1a2a (M64.1/Page44.1, M55)''': 한국, 일본(야마토 민족, 류큐인, 아이누족)[69]
- '''D1a2a1 (M116.1)''': 한국(가덕도 장항 유적 8호 인골)[70]
- '''D1a2a1a (M125)'''
- '''D1a2a1a1 (P42)'''
- '''D1a2a1a2 (IMS-JST022457)''': 일본 황실
- '''D1a2a1a3 (CTS10972)''': 일본(도쿄도), 중국(지린성)
- '''D1a2a1b (P120)'''
- '''D1a2a1c (CTS6609)''': 한국(충청남도), 일본(도쿄도)
- '''D1a2a1c1 (Z1574)''': 일본(도쿄도)
- '''D1a2a1c1a (CTS11032)'''
- '''D1a2a1c1a1 (Z1527)''': 일본
- '''D1a2a1c1a1a (CTS6909)''': 일본(도쿄도)
- '''D1a2a1c1a1b (CTS3033)''': 일본(도치기현, 도쿄도)
- '''D1a2a1c1a1c (Z30644)'''
- '''D1a2a2 (CTS131)'''
- '''D1a2a2a (CTS220)'''
- '''D1a2a2b (CTS68)''': 일본(레분 섬)
- '''D1a2b (Y34637)''': 안다만 제도[71]
- '''D1b (L1378)''': 필리핀(막탄 섬, 루손 섬)[72]
하플로그룹 D는 하플로그룹 DE에서 갈라져 나왔으며, D1은 아시아에서, D2는 아프리카에서 주로 발견된다. 특히 D1a는 일본, 티베트, 안다만 제도 등에서 높은 빈도로 나타난다.
하플로그룹 D는 다른 주요 하플로그룹([하플로그룹 O] 등)과는 7만 년 이상 전에 분기된 매우 고립적인 계통으로 알려져 있다.
4. 1. D1 (M174)
D1 (M174)은 SNP M174에 의해 정의되며, 이전에는 D-M174로 불렸다. D1은 D-CTS3946의 하위 그룹이다.2019년 연구(Haber et al. 2019)에서 나이지리아인 3개 샘플이 하플로그룹 E보다 D에 가깝다는 것이 밝혀졌다. 이 샘플들은 E가 갖지 않는 SNP를 D와 7개 공유하며, "D2"로 D에 편입되었다.[29] 이 발견으로 기존 D (M174)는 D1으로, 새롭게 발견된 D2 (D-A5580.2)와 D1 (M174)을 합하여 D는 CT3946 변이에 의해 정의되는 그룹으로 확대되었다.
D1은 현재 일본 열도, 남서 제도, 안다만 제도, 티베트 고원, 중국 남부, 윈난성 등에서 고빈도로 발견되며, 그 외 아시아와 아프리카의 매우 제한된 지역에서 산발적으로 발견된다.
D1은 하플로그룹 O 등과는 7만 년 이상 분기된 매우 고립적인 계통이다. 동아시아에서 가장 오래된 유형으로 추정되지만, O 계통이 동아시아 및 동남아시아에 널리 유입되면서 섬나라인 일본이나 산악 지대인 티베트에만 D 계통이 남았다는 설도 있다.[33]
하위 그룹으로는 D1a (CTS11577)와 D1b (L1378) 등이 있다.
4. 1. 1. D1a (CTS11577)
일본 열도, 남서 제도, 안다만 제도, 티베트 고원, 중국 남부, 윈난성에서 높은 빈도로 관찰되는 것 외에는 아시아, 아프리카의 매우 제한된 지역에서 드물게 발견된다.[31]티베트에서는 D1a1-Z27276이, 일본에서는 D1a2a-M55이, 안다만 제도에서는 D1a2b-Y34537이 높은 빈도로 나타난다.[31]
이들 하플로그룹은 같은 하플로그룹 D에 속해 있어도 하위 그룹이 다르기 때문에 5만 3000년 이상 전에 분기되었다.[32]
4. 1. 2. D1b (L1378)
주어진 소스에는 'D1b (L1378)'에 대한 정보가 없으며, 필리핀과의 관련성도 언급되지 않았다. 따라서 주어진 소스와 요약 정보로는 해당 섹션을 작성할 수 없다. 섹션 제목과 일치하는 내용을 소스에서 찾을 수 없으므로, 빈 문자열을 반환한다.4. 2. D2 (A5580.2)
하플로그룹 D2는 하플로그룹 D-CTS3946의 구석기 시대 하위 그룹으로, D0라고도 불린다. D-M174는 이 발견으로 인해 D1으로 이름이 변경되었다.[10][11]2019년 연구(Haber et al. 2019)에 따르면,[29] DE의 하위 그룹 중 D 계통에도 E 계통에도 속하지 않아 어느 계통에 가까운지 불분명했던 파라그룹 DE*로 여겨졌던 나이지리아인 3명의 샘플이[35] 하플로그룹 E보다 하플로그룹 D에 더 가깝다는 사실이 밝혀졌다. 이 샘플들은 하플로그룹 E에는 없는 SNP 7개를 하플로그룹 D와 공유하며, "D2"로 하플로그룹 D에 편입되었다.
이 발견으로 기존의 하플로그룹 D (M174 변이로 정의)는 하플로그룹 D1으로 명칭이 변경되었고, 새롭게 발견된 D2 (D-A5580.2)와 하플로그룹 D1 (M174)을 포함하는 하플로그룹 D는 CT3946 변이로 정의되는 그룹으로 확대되었다. D2는 서아시아 (사우디아라비아, 시리아)에서도 발견되었다.[36]
이처럼 하플로그룹 D2가 아프리카에서 발견됨에 따라, 하플로그룹 D (CT3946)는 아프리카에서 탄생했다고 추정된다.[37]
5. 계통수
ISOGG 2019 계통수(ver.14.151)는 다음과 같다.[68]
'''D1a''' (CTS11577)
*'''D1a1''' (F6251/Z27276)
**'''D1a1a''' (M15) 중국 (특히 먀오족, 이족, 티베트인 등), 동남아시아, 몽골, 중앙아시아
***'''D1a1a1''' (F849)
'''D1a1a1a''' (N1)
*'''D1a1a1a1''' (Z27269) 일본, 중국, 티베트[69]
**'''D1a1a1a1a''' (PH4979)
***'''D1a1a1a1a1''' (BY15119/Z29428)
'''D1a1a1a2''' (Z31591) 네팔(타망족), 카자흐스탄, 중국(이족)
***'''D1a1a2''' (F1070) 중국, 태국
**'''D1a1b''' (P99) 티베트에서 높은 빈도로 발견되며, 때때로 중국, 몽골,[69] 중앙아시아,[69] 및 알타이 공화국의 다른 지역에서도 발견된다.
*'''D1a2'''(Z3660)
**'''D1a2a''' (M64.1/Page44.1, M55) 일본[69]
***'''D1a2a1''' (M116.1)
***'''D1a2a2''' (CTS131)
'''D1a2a2a''' (CTS220)
'''D1a2a2b''' (CTS68) 일본(레분 섬)
**'''D1a2b''' (Y34637) 안다만 제도[71]
'''D1b''' (L1378) 필리핀[72]
인류 Y-염색체 DNA 하플로그룹 계보는 다음과 같다.
참조
[1]
논문
A Rare Deep-Rooting D0 African Y-Chromosomal Haplogroup and Its Implications for the Expansion of Modern Humans Out of Africa
2019-08
[2]
논문
Use of y chromosome and mitochondrial DNA population structure in tracing human migrations
[3]
간행물
Carriers of mitochondrial DNA macrohaplogroup L3 basic lineages migrated back to Africa from Asia around 70,000 years ago
2017
[4]
논문
A Southeast Asian origin for present-day non-African human Y chromosomes
2021-02
[5]
웹사이트
FamilyTreeDNA – Y-DNA D Haplogroup Project
https://www.familytr[...]
[6]
웹사이트
D YTree
https://www.yfull.co[...]
[7]
웹사이트
D-Y330435 YTree
https://www.yfull.co[...]
[8]
Youtube
The Tree of Mankind fromFTDNA (Mike Sager)
https://www.youtube.[...]
2020-02
[9]
웹사이트
New Discoveries Shed Light on Out of Africa Theory, and Beyond
https://dna-explaine[...]
2020-12-10
[10]
웹사이트
Exciting New Y DNA Haplogroup D Discoveries!
https://dna-explaine[...]
2019-07-06
[11]
웹사이트
Image
https://i2.wp.com/dn[...]
[12]
문서
Y-DNA Haplogroup D and its Subclades – 2019
https://docs.google.[...]
[13]
논문
"Dual origins of the Japanese: common ground for hunter-gatherer and farmer Y chromosomes"
2006
[14]
웹사이트
D YTree
https://www.yfull.co[...]
2019-09-03
[15]
문서
Y-DNA Haplogroup D and its Subclades – 2014
http://www.isogg.org[...]
[16]
논문
Seeing the wood for the trees: a minimal reference phylogeny for the human Y chromosome
[17]
웹사이트
'Y-DNA Haplogroup Tree 2015'
http://www.isogg.org[...]
2015-02-01
[18]
문서
[19]
문서
[20]
문서
[21]
문서
[22]
문서
[23]
문서
[24]
문서
[25]
문서
[26]
문서
[27]
문서
[28]
문서
[29]
논문
Carriers of mitochondrial DNA macrohaplogroup L3 basic lineages migrated back to Africa from Asia around 70,000 years ago.
https://www.biorxiv.[...]
2017-12-13
[30]
문서
[31]
문서
[32]
논문
Y-chromosomal sequences of diverse Indian populations and the ancestry of the Andamanese.
https://www.research[...]
2017
[33]
논문
Y chromosome evidence of earliest modern human settlement in East Asia and multiple origins of Tibetan and Japanese populations
http://www.biomedcen[...]
BioMed Central
2018-08-04
[34]
논문
"Rare deep-rooting Y chromosome lineages in humans: Lessons for Phylogeography"
2003-09
[35]
논문
"Rare deep-rooting Y chromosome lineages in humans: Lessons for Phylogeography"
https://www.ncbi.nlm[...]
2003-09
[36]
웹사이트
Exciting New Y DNA Haplogroup D Discoveries!
https://dna-explaine[...]
2019-06-21
[37]
논문
A Rare Deep-Rooting D0 African Y-Chromosomal Haplogroup and Its Implications for the Expansion of Modern Humans Out of Africa
2019-08
[38]
서적
DNA・考古・言語の学際研究が示す新・日本列島史
勉誠出版
2009
[39]
논문
Carriers of mitochondrial DNA macrohaplogroup L3 basic lineages migrated back to Africa from Asia around 70,000 years ago.
https://www.biorxiv.[...]
2017-12-13
[40]
논문
Little genetic differentiation as assessed by uniparental markers in the presence of substantial language variation in peoples of the Cross River region of Nigeria
[41]
논문
Y chromosome haplotypes reveal prehistorical migrations to the Himalayas
[42]
논문
The Northeast Indian Passageway: A Barrier or Corridor for Human Migrations?
[43]
논문
YAP insertion signature in South Asia
[44]
논문
Austro-Asiatic Tribes of Northeast India Provide Hitherto Missing Genetic Link between South and Southeast Asia
null
[45]
문서
D-M15以外のマーカーについては検査されていない。
[46]
논문
Analyses of genetic structure of Tibeto-Burman populations reveals sex-biased admixture in southern Tibeto-Burmans
2004
[47]
논문
Paternal Population History of East Asia: Sources, Patterns, and Microevolutionary Processes.
2001
[48]
논문
Male Demography in East Asia: A North–South Contrast in Human Population Expansion Times.
2006-04
[49]
웹사이트
Genetic origin of Kadai-speaking Gelong people on Hainan island viewed from Y chromosomes
http://www.nature.co[...]
[50]
논문
An updated tree of Y-chromosome Haplogroup O and revised phylogenetic positions of mutations P164 and PK4.
2011
[51]
논문
Genetic Affinities of the Andaman Islanders, a Vanishing Human Population
2003-01-21
[52]
웹사이트
East Asian ancestry in India
https://www.research[...]
null
2015-01
[53]
논문
Genome of a middle Holocene hunter-gatherer from Wallacea
https://www.ncbi.nlm[...]
2021
[54]
웹사이트
D Haplogroup (Y-DNA) Project
http://www.familytre[...]
FAMILY TREE DNA
2013-06-01
[55]
문서
田島等の2004年の論文"Genetic Origins of the Ainu inferred from combined DNA analyses of maternal and paternal lineages"による
[56]
논문
Overview of genetic variation in the Y chromosome of modern Japanese males.
[57]
논문
Y-chromosomal Binary Haplogroups in the Japanese Population and their Relationship to 16 Y-STR Polymorphisms
John Wiley & Sons
2007-02-02
[58]
논문
Y chromosome sequence variation and the history of human populations.
2000-11
[59]
논문
Genetic variations on the Y chromosome in the Japanese population and implications for modern human Y chromosome lineage.
1999
[60]
논문
Dual origins of the Japanese: common ground for hunter-gatherer and farmer Y chromosomes
2006
[61]
논문
A study of the geographic distribution of Y chromosomal and mitochondrial DNA haplogroups in Japanese population by Super Science High School Consortium (SSH).
2016
[62]
논문
Mitochondrial DNA and Y Chromosome Variation Provides Evidence for a Recent Common Ancestry between Native Americans and Indigenous Altaians.
2012-02-10
[63]
논문
Gene pool differences between Northern and Southern Altaians inferred from the data on Y-chromosomal haplogroups
[64]
논문
Genetic origins of the Ainu inferred from combined DNA analyses of maternal and paternal lineages
2004
[65]
웹사이트
Y-DNA Haplogroup D and its Subclades - 2019
https://docs.google.[...]
[66]
웹사이트
Y-DNA Haplogroup D and its Subclades - 2014
http://www.isogg.org[...]
[67]
논문
A Rare Deep-Rooting D0 African Y-Chromosomal Haplogroup and Its Implications for the Expansion of Modern Humans Out of Africa
2019-08
[68]
웹인용
Y-DNA Haplogroup D-M174 and its Subclades - 2019
https://docs.google.[...]
[69]
논문
Dual origins of the Japanese: common ground for hunter-gatherer and farmer Y chromosomes
2006
[70]
웹사이트
https://researchmap.[...]
[71]
웹사이트
D YTree
https://www.yfull.co[...]
2019-09-03
[72]
웹사이트
Y-DNA Haplogroup D and its Subclades - 2014
http://www.isogg.org[...]
[73]
논문
A Rare Deep-Rooting D0 African Y-Chromosomal Haplogroup and Its Implications for the Expansion of Modern Humans Out of Africa
2019-08
[74]
웹인용
Exciting New Y DNA Haplogroup D Discoveries!
https://dna-explaine[...]
2019-07-08
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com